Unravelling the Links between Phage Adsorption and Successful Infection in Clostridium difficile

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Phages Used and Their Growth Conditions
2.2. Phage Host Range Analysis and Efficiency of Plating
2.3. Phage Adsorption and Irreversible Phage Binding
- (i)
- Phage titer in control sample − S1 = total bound phage
- (ii)
- S2 × 100/total bound phage = percentage reversibly bound phage
- (iii)
- 100 − percentage reversibly bound phage = percentage of irreversibly bound phage.
2.4. Alignment and Phylogenetic Analysis
2.5. Statistical Analysis
3. Results
3.1. Phage Host Range and Effeciency of Plating
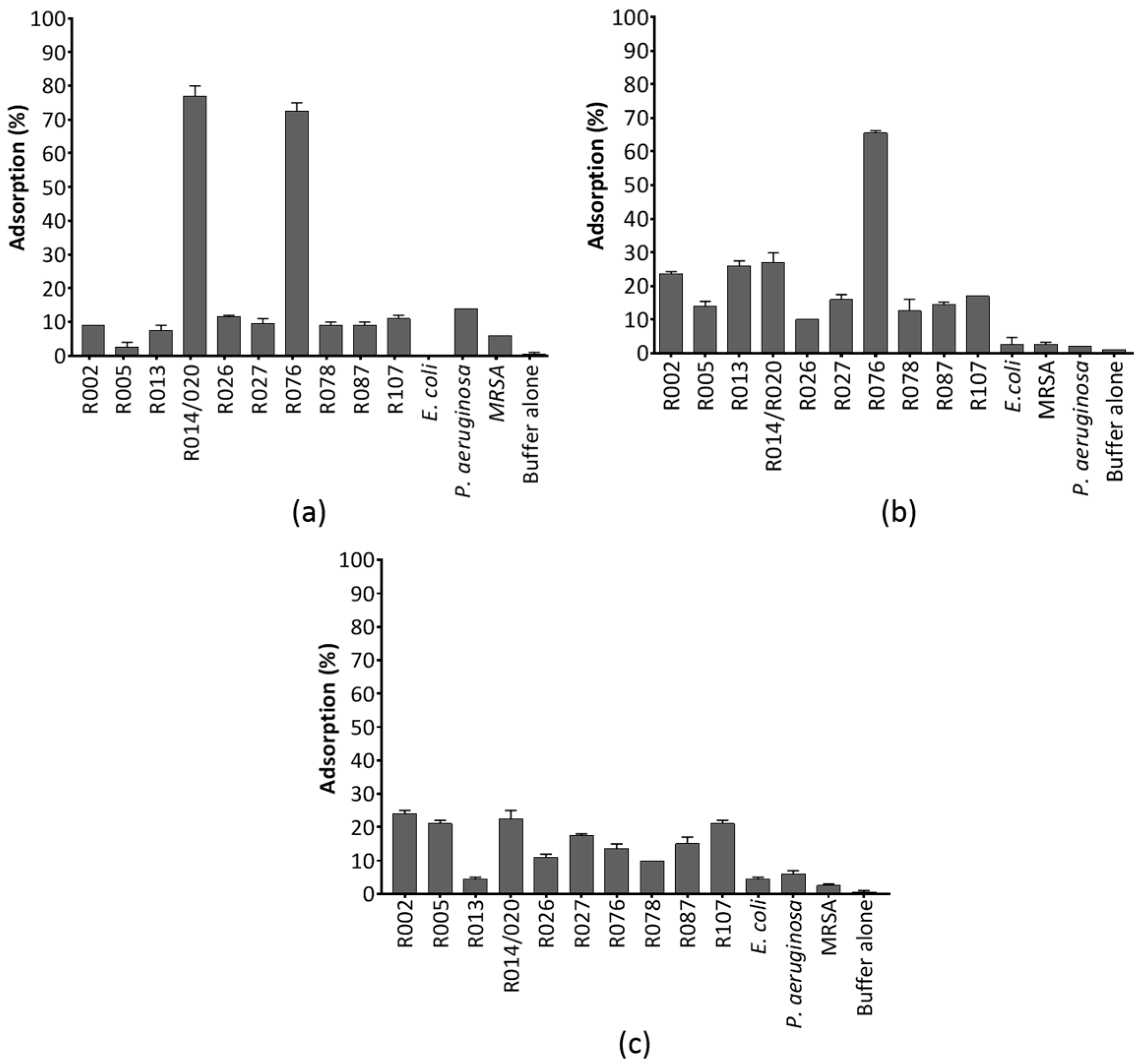
3.2. Adsorption of CDHM1, CDHM3 and CDHM6 to Relevent C. difficile Ribotypes and Other Pathogens
3.2.1. Adsorption Dynamics of CDHM1
3.2.2. Adsorption Dynamics of CDHM3
3.2.3. Adsorption Dynamics of CDHM6
3.3. Irreversible Binding of Phages CDHM1, CDHM3 and CDHM6 to Different C. difficile Ribotypes
3.4. Host Range, EOP, Adsorption and Irreversible Binding of Other C. difficile Phages to Different Ribotypes
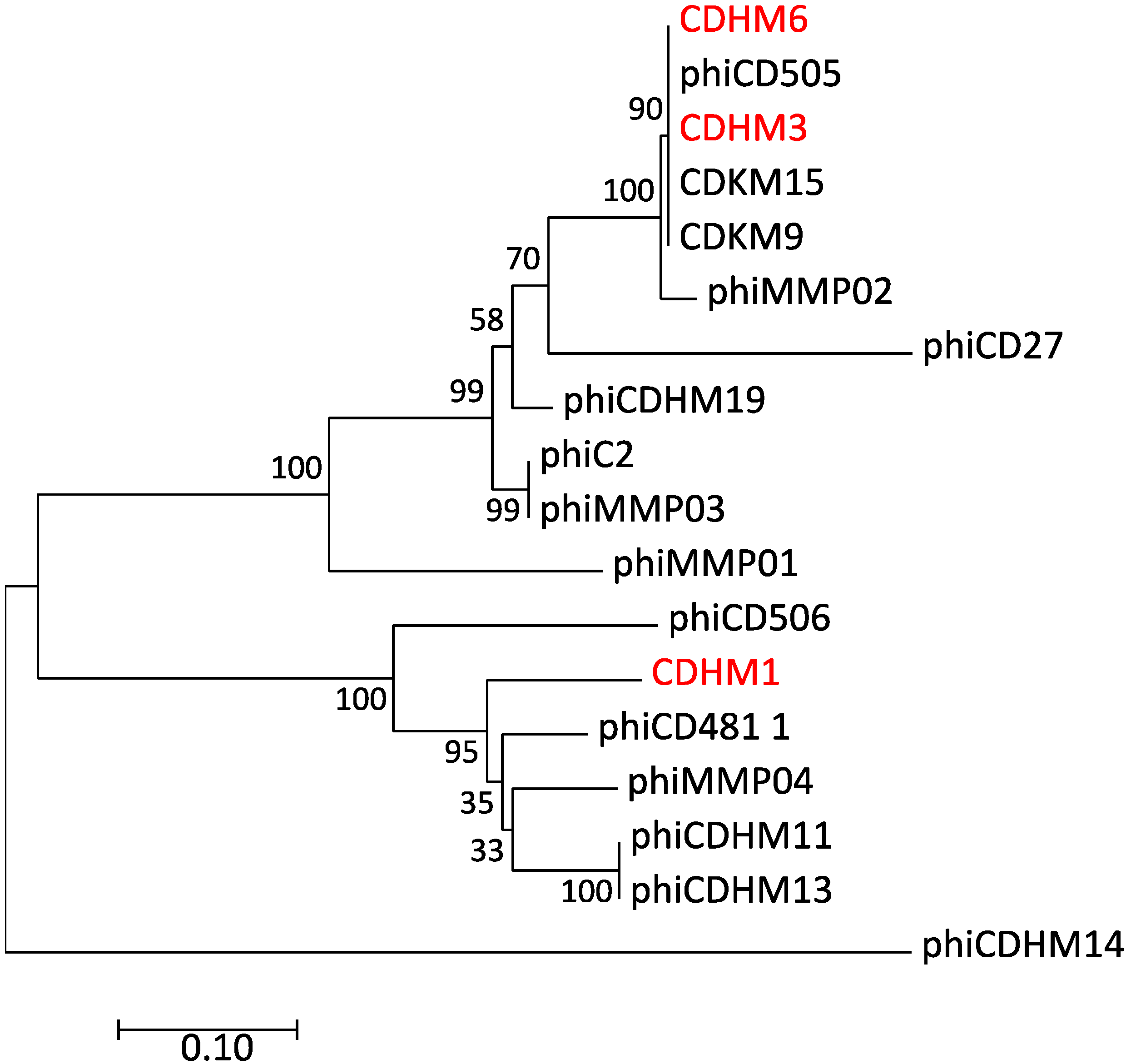
3.5. Alignement of Putative Phage Tail Fibers
3.6. Phylogentic Analysis of Putative Tail Fiber Proteins of C. diffiicle Phages
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile Infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Longo, D.L.; Leffler, D.A.; Lamont, J.T. Clostridium difficile Infection. N. Engl. J. Med. 2015, 372, 1539–1548. [Google Scholar]
- Vardakas, K.Z.; Polyzos, K.A.; Patouni, K.; Rafailidis, P.I.; Samonis, G.; Falagas, M.E. Treatment failure and recurrence of Clostridium difficile infection following treatment with vancomycin or metronidazole: A systematic review of the evidence. Int. J. Antimicrob. Agents 2012, 40, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N. Clostridium difficile infection: Epidemiology, risk factors and management. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, C.; Dionne, L.; Julien, A.; Longtin, Y. Clinical characteristics and outcome of patients with Clostridium difficile infection diagnosed by PCR versus a three-step algorithm. Clin. Microbiol. Infect. 2014, 20, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S. Editorial commentary: Changing epidemiology of Clostridium difficile and emergence of new virulent strains. Clin. Infect. Dis. 2014, 58, 1731–1733. [Google Scholar] [CrossRef] [PubMed]
- Agency, H.P. Clostridium difficile Ribotyping Network (CDRN) for England and Northern Ireland. 2011. Available online: http://www.hpa.org.uk/web/HPAwebFile/HPAweb_C/1296681523205 (accessed on 21 March 2018).
- Fawley, W.N.; Freeman, J.; Smith, C.; Harmanus, C.; van den Berg, R.J.; Kuijper, E.J.; Wilcox, M.H. Use of highly discriminatory fingerprinting to analyze clusters of Clostridium difficile infection cases due to epidemic ribotype 027 strains. J. Clin. Microbiol. 2008, 46, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, M. Clostridium difficile Ribotyping Network (CDRN) for England and Northern Ireland. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/491253/CDRN_2013-15_Report.pdf (accessed on 29 April 2018).
- Peng, Z.; Jin, D.; Kim, H.B.; Stratton, C.W.; Wu, B.; Tang, Y.W.; Suna, X. Update on antimicrobial resistance in Clostridium difficile: Resistance mechanisms and antimicrobial susceptibility testing. J. Clin. Microbiol. 2017, 55, 1998–2008. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Otieno, J.R.; Thanki, A.; Blades, M.J.; Millard, A.D.; Browne, H.P.; Lawley, T.D.; Clokie, M.R.J. As clear as mud? Determining the diversity and prevalence of prophages in the draft genomes of estuarine isolates of Clostridium difficile. Genome Biol. Evol. 2015, 7, 1842–1855. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Sillankorva, S.; Merabishvili, M.; Kluskens, L.D.; Azeredo, J. Unexploited opportunities for phage therapy. Front. Pharmacol. 2015, 6, 180. [Google Scholar] [CrossRef] [PubMed]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargreaves, K.R.; Clokie, M.R.J. Clostridium difficile phages: Still difficult? Front. Microbiol. 2014, 5, 184. [Google Scholar] [CrossRef] [PubMed]
- Nale, J.Y.; Spencer, J.; Hargreaves, K.R.; Buckley, A.M.; Trzepin, P. Bacteriophage Combinations Significantly Reduce Clostridium difficile Growth In Vitro and Proliferation In Vivo. Antimicrob. Agents Chemother. 2016, 60, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.; Ong, P.F.; Song, K.P.; Riley, T.V.; Chang, B.J. The complete genome sequence of Clostridium difficile phage phiC2 and comparisons to phiCD119 and inducible prophages of CD630. Microbiology 2007, 153, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.R.; Sekulovic, O.; Dupuy, B.; Soutourina, O.; Monot, M.; Fortier, L.C. High prevalence and genetic diversity of large phiCD211 (phiCDIF1296T)-like prophages in Clostridioides difficile. Appl. Environ. Microbiol. 2018, 84, e02164-17. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.C.; Moineau, S. Morphological and genetic diversity of temperate phages in Clostridium difficile. Appl. Environ. Microbiol. 2007, 73, 7358–7366. [Google Scholar] [CrossRef] [PubMed]
- Sekulovic, O.; Garneau, J.R.; Néron, A.; Fortier, L.C. Characterization of temperate phages infecting Clostridium difficile isolates of human and animal origins. Appl. Environ. Microbiol. 2014, 80, 2555–2563. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.; Riley, T.V.; Chang, B.J. Isolation and characterization of temperate bacteriophages of Clostridium difficile. Appl. Environ. Microbiol. 2005, 71, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.J.; Barylski, J.; Hargreaves, K.R.; Millard, A.A.; Vinner, G.K.; Clokie, M.R.J. Two novel myoviruses from the north of Iraq reveal insights into Clostridium difficile phage diversity and biology. Viruses 2016, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Mahony, D.E.; Bell, P.D.; Easterbrook, K.B. Two bacteriophages of Clostridium difficile. J. Clin. Microbiol. 1985, 21, 251–254. [Google Scholar] [PubMed]
- Ramesh, V.; Fralick, J.A.; Rolfe, R.D. Prevention of Clostridium difficile-induced ileocecitis with Bacteriophage. Anaerobe 1999, 5, 69–78. [Google Scholar] [CrossRef]
- Nale, J.Y.; Chutia, M.; Carr, P.; Hickenbotham, P.T.; Clokie, M.R.J. ‘Get in early’; Biofilm and wax moth (Galleria mellonella) models reveal new insights into the therapeutic potential of Clostridium difficile bacteriophages. Front. Microbiol. 2016, 7, 1383. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Ramachandran, A.; Thanki, A.M.; Vukusic, F.B.I.; Barylski, J.; Clokie, M.R.J. Bacteriophages are more virulent to bacteria with human cells than they are in bacterial culture; insights from HT-29 cells. Sci. Rep. 2018, 8, 5091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meader, E.; Mayer, M.J.; Gasson, M.J.; Steverding, D.; Carding, S.R.; Narbad, A. Bacteriophage treatment significantly reduces viable Clostridium difficile and prevents toxin production in an in vitro model system. Anaerobe 2010, 16, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Nale, J.; Redgwell, T.; Millard, A.; Clokie, M. Efficacy of an Optimised Bacteriophage Cocktail to Clear Clostridium difficile in a Batch Fermentation Model. Antibiotics 2018, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Meader, E.; Mayer, M.J.; Steverding, D.; Carding, S.R.; Narbad, A. Evaluation of bacteriophage therapy to control Clostridium difficile and toxin production in an in vitro human colon model system. Anaerobe 2013, 22, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Dowah, A.S.A.; Clokie, M.R.J. Review of the nature, diversity and structure of bacteriophage receptor binding proteins that target Gram-positive bacteria. Biophys. Rev. 2017, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.B.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef] [PubMed]
- De Smet, J.; Hendrix, H.; Blasdel, B.G.; Danis-Wlodarczyk, K.; Lavigne, R. Pseudomonas predators: Understanding and exploiting phage-host interactions. Nat. Rev. Microbiol. 2017, 15, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Quiberoni, A.; Guglielmotti, D.; Binetti, A.; Reinheimer, J. Characterization of three Lactobacillus delbrueckii subsp. bulgaricus phages and the physicochemical analysis of phage adsorption. J. Appl. Microbiol. 2004, 96, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, R.; Chapman-McQuiston, E.; Wu, X.L. On kinetics of phage adsorption. Biophys. J. 2007, 93, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Kasman, L.M.; Kasman, A.; Westwater, C.; Dolan, J.; Schmidt, M.G.; Norris, J.S. Overcoming the phage replication threshold: A mathematical model with implications for phage therapy. J. Virol. 2002, 76, 5557–5564. [Google Scholar] [CrossRef] [PubMed]
- Baptista, C.; Santos, M.A.; São-José, C. Phage SPP1 reversible adsorption to Bacillus subtilis cell wall teichoic acids accelerates virus recognition of membrane receptor YueB. J. Bacteriol. 2008, 190, 4989–4996. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, J.; Takemura, I.; Satoh, M.; Kato, S.; Ujihara, T.; Akechi, K.; Matsuzaki, S.; Daibata, M. Improved adsorption of an Enterococcus faecalis bacteriophage ΦEF24C with a spontaneous point mutation. PLoS ONE 2011, 6, e26648. [Google Scholar] [CrossRef] [PubMed]
- Jakutytė, L.; Lurz, R.; Baptista, C.; Carballido-Lopez, R.; São-José, C.; Tavares, P.; Daugelavičius, R. First steps of bacteriophage SPP1 entry into Bacillus subtilis. Virology 2012, 422, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; van Sinderen, D. Structural aspects of the interaction of dairy phages with their host bacteria. Viruses 2012, 4, 1410–1424. [Google Scholar] [CrossRef] [PubMed]
- Tokman, J.I.; Kent, D.J.; Wiedmann, M.; Denes, T. Temperature significantly affects the plaquing and adsorption efficiencies of Listeria phages. Front. Microbiol. 2016, 7, 631. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Patel, K.V.; Hickenbotham, P.T.; Nale, J.Y.; Hargreaves, K.R.; Clokie, M.R.J. Prophage carriage and diversity within clinically relevant strains of Clostridium difficile. Appl. Environ. Microbiol. 2012, 78, 6027–6034. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Flores, C.O.; Lawley, T.D.; Clokie, R.J. Abundant and Diverse Clustered Regularly Interspaced Short Palindromic Repeat Spacers in Clostridium difficile Strains and Prophages Target Multiple Phage Types within This Pathogen. MBio 2014, 5, e01045-13. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Colvin, H.V.; Patel, K.V.; Clokie, J.J.P.; Clokie, M.R.J. Genetically diverse Clostridium difficile strains harboring abundant prophages in an estuarine environment. Appl. Environ. Microbiol. 2013, 79, 6236–6243. [Google Scholar] [CrossRef] [PubMed]
- Van Twest, R.; Kropinski, A.M. Bacteriophage enrichment from water and soil. Methods Mol. Biol. 2009, 501, 15–21. [Google Scholar] [PubMed]
- Kutter, E. Phage host range and efficiency of plating. In Bacteriophages: Methods and Protocols; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: New York, NY, USA, 2009; Volume 501, pp. 141–149. [Google Scholar]
- Mirzaei, M.K.; Nilsson, A.S. Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.O.N.; Goldstein, B.; Holowka, D.; Baird, B. Brief communication the effect of receptor density on the forward rate constant for binding of ligands to cell surface receptors. Biophys. J. 1982, 52, 657–662. [Google Scholar]
- Hyman, P.; Abedon, S.T. Practical methods for determining phage growth parameters. In Bacteriophages: Methods and Protocols, Volume 1; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: New York, NY, USA, 2009; Volume 501, pp. 175–202. [Google Scholar]
- Li, X.; Koç, C.; Kühner, P.; Stierhof, Y.D.; Krismer, B.; Enright, M.C.; Penadés, J.R.; Wolz, C.; Stehle, T.; Cambillau, C.; et al. An essential role for the baseplate protein Gp45 in phage adsorption to Staphylococcus aureus. Sci. Rep. 2016, 6, 26455. [Google Scholar] [CrossRef] [PubMed]
- Bielmann, R.; Habann, M.; Eugster, M.R.; Lurz, R.; Calendar, R.; Klumpp, J.; Loessner, M.J. Receptor binding proteins of Listeria monocytogenes bacteriophages A118 and P35 recognize serovar-specific teichoic acids. Virology 2015, 477, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Fagan, R.P.; Fairweather, N.F. Biogenesis and functions of bacterial S-layers. Nat. Rev. Microbiol. 2014, 12, 211–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartual, S.G.; Otero, J.M.; Garcia-Doval, C.; Llamas-Saiz, A.L.; Kahn, R.; Fox, G.C.; van Raaij, M.J. Structure of the bacteriophage T4 long tail fiber receptor-binding tip. Proc. Natl. Acad. Sci. USA 2010, 107, 20287–20292. [Google Scholar] [CrossRef] [PubMed]
- Williamson, S.J.; McLaughlin, M.R.; Paul, J.H. Interaction of the PhiHSIC virus with its host: Lysogeny or pseudolysogeny? Appl. Environ. Microbiol. 2001, 67, 1682–1688. [Google Scholar] [CrossRef] [PubMed]
- Leiman, P.G.; Arisaka, F.; van Raaij, M.J.; Kostyuchenko, V.A.; Aksyuk, A.A.; Kanamaru, S.; Rossmann, M.G. Morphogenesis of the T4 tail and tail fibers. Virol. J. 2010, 7, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokhorov, N.S.; Riccio, C.; Zdorovenko, E.L.; Shneider, M.M.; Browning, C.; Knirel, Y.A.; Leiman, P.G.; Letarov, A.V. Function of bacteriophage G7C esterase tailspike in host cell adsorption. Mol. Microbiol. 2017, 105, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Nordstrom, K.; Forsgren, A. Effect of Protein A on Adsorption of Bacteriophages to Staphylococcus aureus. J. Virol. 1974, 14, 198–202. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| C. difficile Ribotype | Strain Identity | Infectivity of Phages 1 | Efficiency of Plating (%) | ||||
|---|---|---|---|---|---|---|---|
| CDHM1 | CDHM3 | CDHM6 | CDHM1 | CDHM3 | CDHM6 | ||
| 013 | ASH | − | + | − | 0 | 0 | 0 |
| 014/020 | ATJ | ++ | ++ | ++ | 100 | 73 | 81 |
| 026 | ARU | + | − | + | 0 | 0 | 0 |
| 027 | CD105LC1 | − | + | − | 0 | 0 | 0 |
| 076 | CD105HE1 | ++ | ++ | ++ | 100 | 100 | 100 |
| Ribotype | Strain Identity | Irreversible Binding (%) of C. difficile Phages 1 | ||
|---|---|---|---|---|
| CDHM1 | CDHM3 | CDHM6 | ||
| 002 | AKC | 13 | 4 | 14 |
| 005 | AIN | 18 | 12 | 15 |
| 013 | ASH | 3 | 51 | 21 |
| 014/020 | ATJ | 90 | 93 | 90 |
| 026 | ARU | 59 | 2 | 58 |
| 027 | CD105LC1 | 7 | 52 | 5 |
| 076 | CD105HE1 | 95 | 88 | 97 |
| 078 | ASS | 16 | 9 | 7 |
| 087 | APT | 29 | 8 | 10 |
| 107 | ARZ | 9 | 12 | 7 |
| Ribotype | Strain Identity | CDHM2 | CDHM5 | ||
|---|---|---|---|---|---|
| Adsorption (%) 1 | Irreversible Binding (%) 1 | Adsorption (%) 1 | Irreversible Binding (%) 1 | ||
| 002 | AKC | 22 | 29 | 33 | 29 |
| 005 | AIN | 11 | 6 | 10 | 12 |
| 013 | ASH | 3 | 11 | 13 | 9 |
| 014/020 | ATJ | 82 | 92 | 82 | 97 |
| 026 | ARU | 32 | 2 | 10 | 15 |
| 027 | CD105LC1 | 36 | 13 | 29 | 21 |
| 076 | CD105HE1 | 79 | 98 | 80 | 97 |
| 078 | ASS | 20 | 30 | 17 | 20 |
| 087 | APT | 17 | 8 | 10 | 30 |
| 107 | ARZ | 26 | 36 | 17 | 14 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thanki, A.M.; Taylor-Joyce, G.; Dowah, A.; Yakubu Nale, J.; Malik, D.; Rebecca Jane Clokie, M. Unravelling the Links between Phage Adsorption and Successful Infection in Clostridium difficile. Viruses 2018, 10, 411. https://0-doi-org.brum.beds.ac.uk/10.3390/v10080411
Thanki AM, Taylor-Joyce G, Dowah A, Yakubu Nale J, Malik D, Rebecca Jane Clokie M. Unravelling the Links between Phage Adsorption and Successful Infection in Clostridium difficile. Viruses. 2018; 10(8):411. https://0-doi-org.brum.beds.ac.uk/10.3390/v10080411
Chicago/Turabian StyleThanki, Anisha Mahendra, Grace Taylor-Joyce, Ahmed Dowah, Janet Yakubu Nale, Danish Malik, and Martha Rebecca Jane Clokie. 2018. "Unravelling the Links between Phage Adsorption and Successful Infection in Clostridium difficile" Viruses 10, no. 8: 411. https://0-doi-org.brum.beds.ac.uk/10.3390/v10080411