Cellular Interleukin Enhancer-Binding Factor 2, ILF2, Inhibits Japanese Encephalitis Virus Replication In Vitro

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies
2.3. Plasmids
2.4. Lentivirus Preparation
2.5. MTS Assay
2.6. Co-Immunoprecipitation Analysis and LC-MS/MS
2.7. Indirect Immunofluorescence
2.8. Western Blot Assays
2.9. Quantitative Real-Time PCR
2.10. Plaque Assay
2.11. Luciferase Reporter Assay
2.12. Statistical Analysis
3. Results
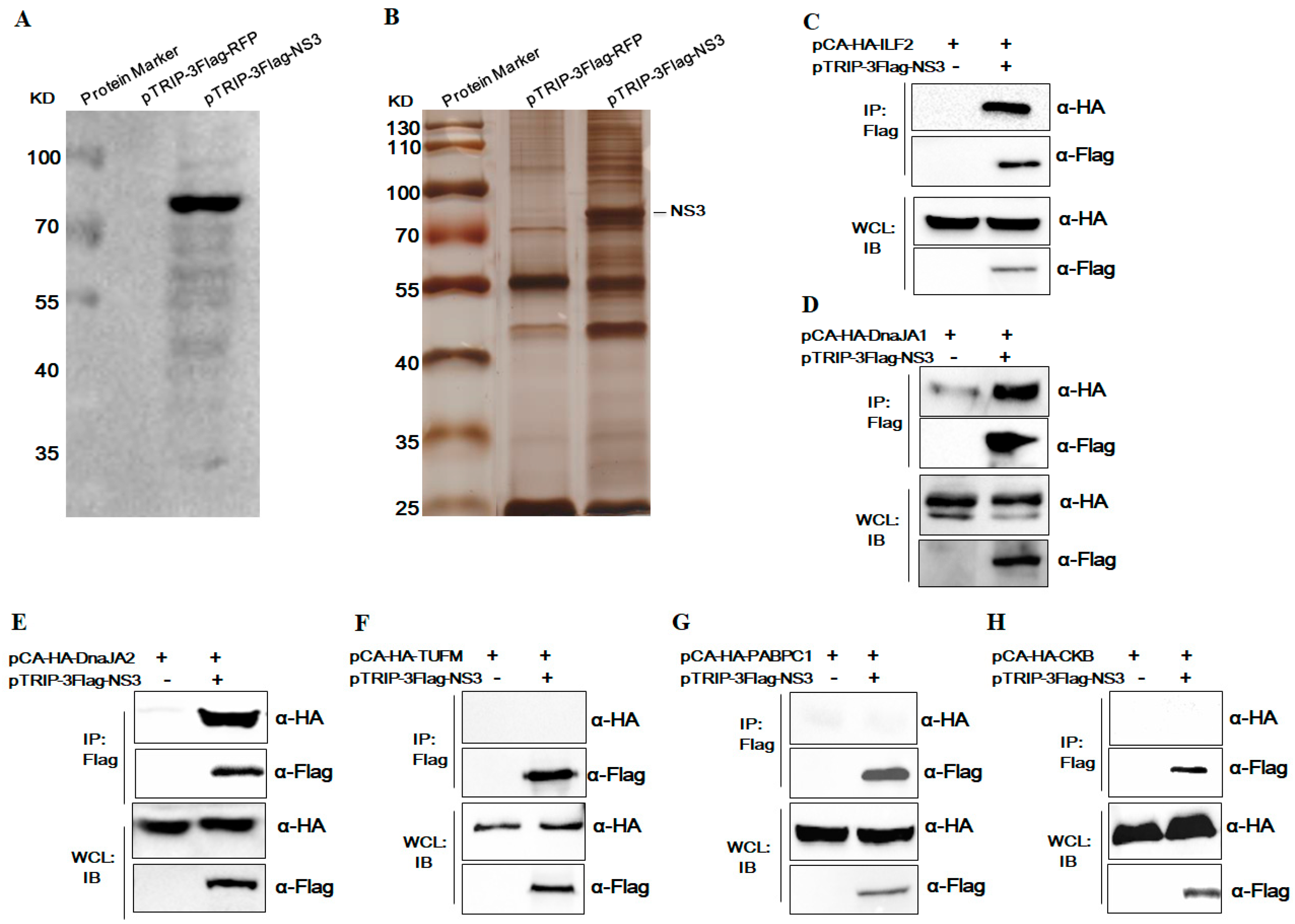
3.1. Identification of Host Cellular Proteins Interacting with JEV NS3
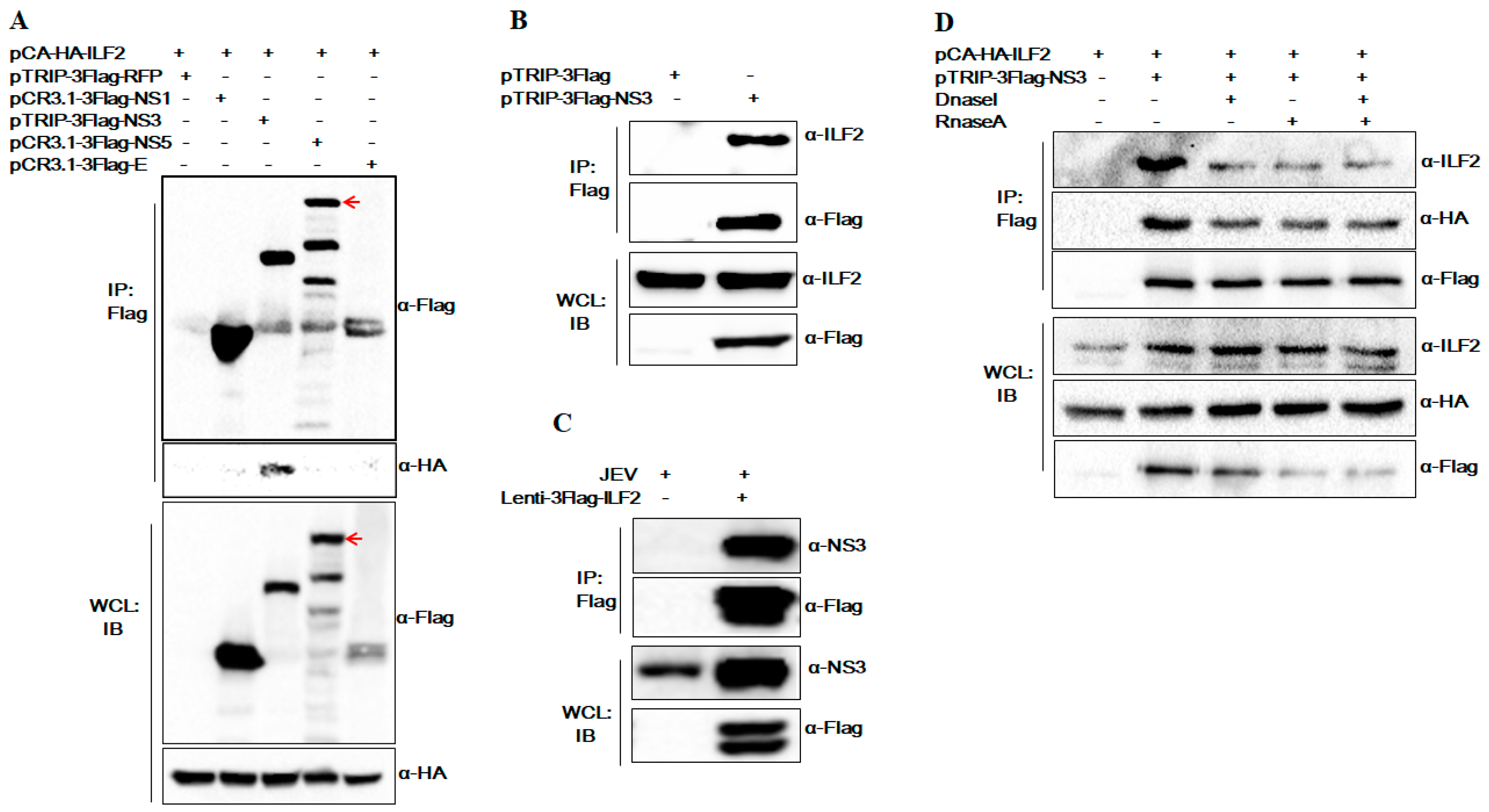
3.2. The Special Interaction of ILF2 and NS3 was not Mediated by DNA or RNA
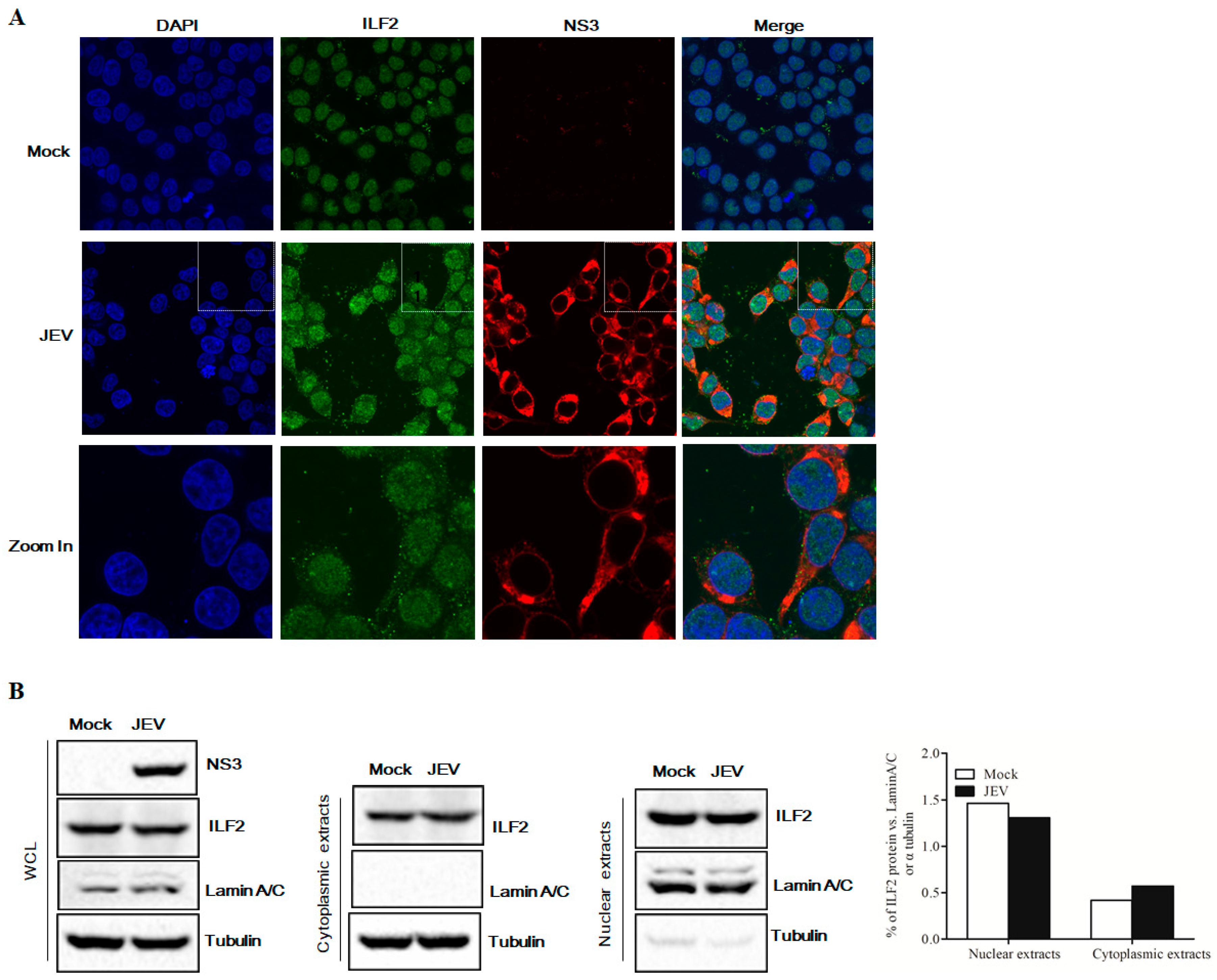
3.3. ILF2 Co-Localized with JEV NS3 during Virus Infection
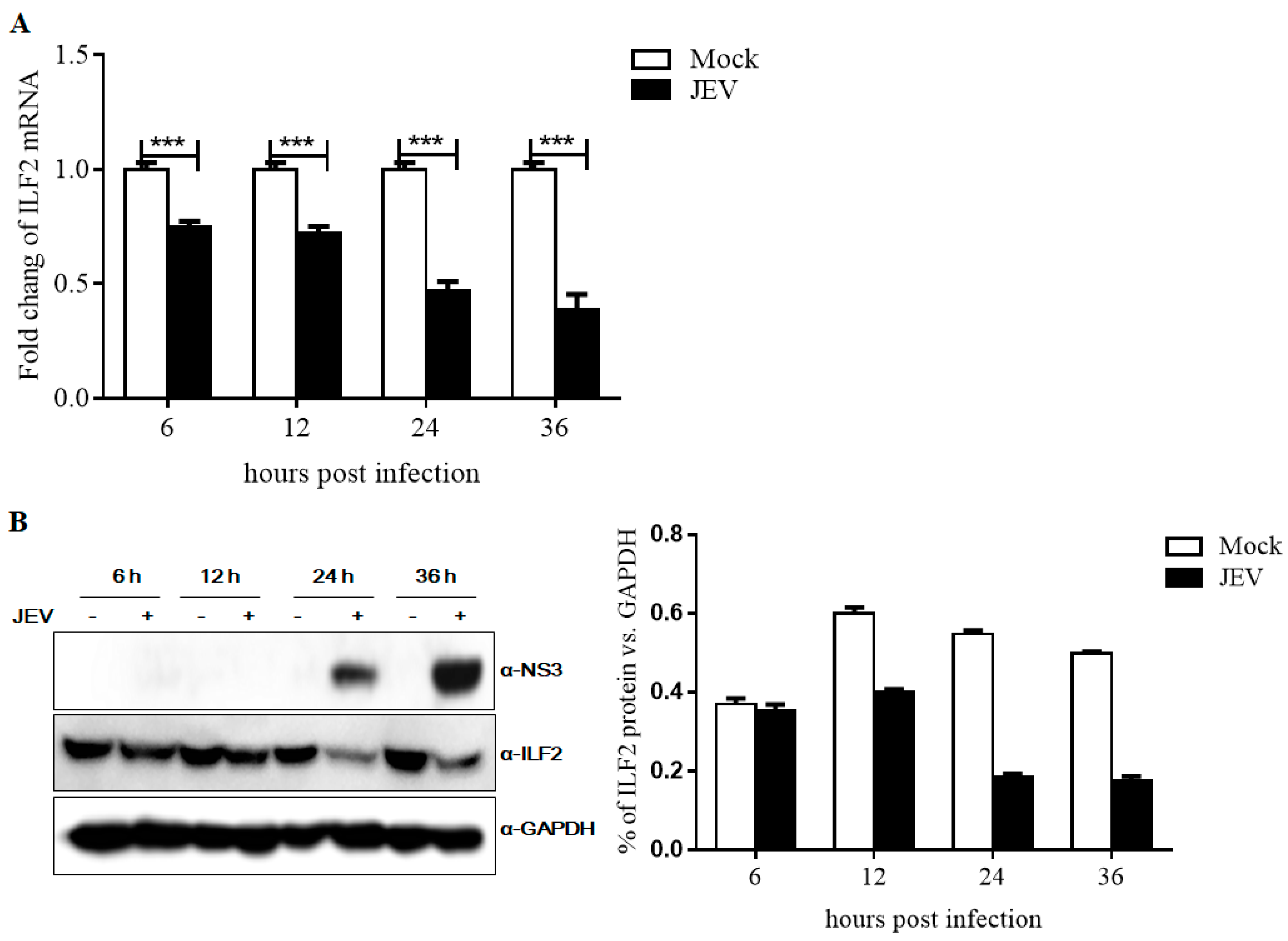
3.4. Expression of ILF2 in 293T Cells Infected with JEV
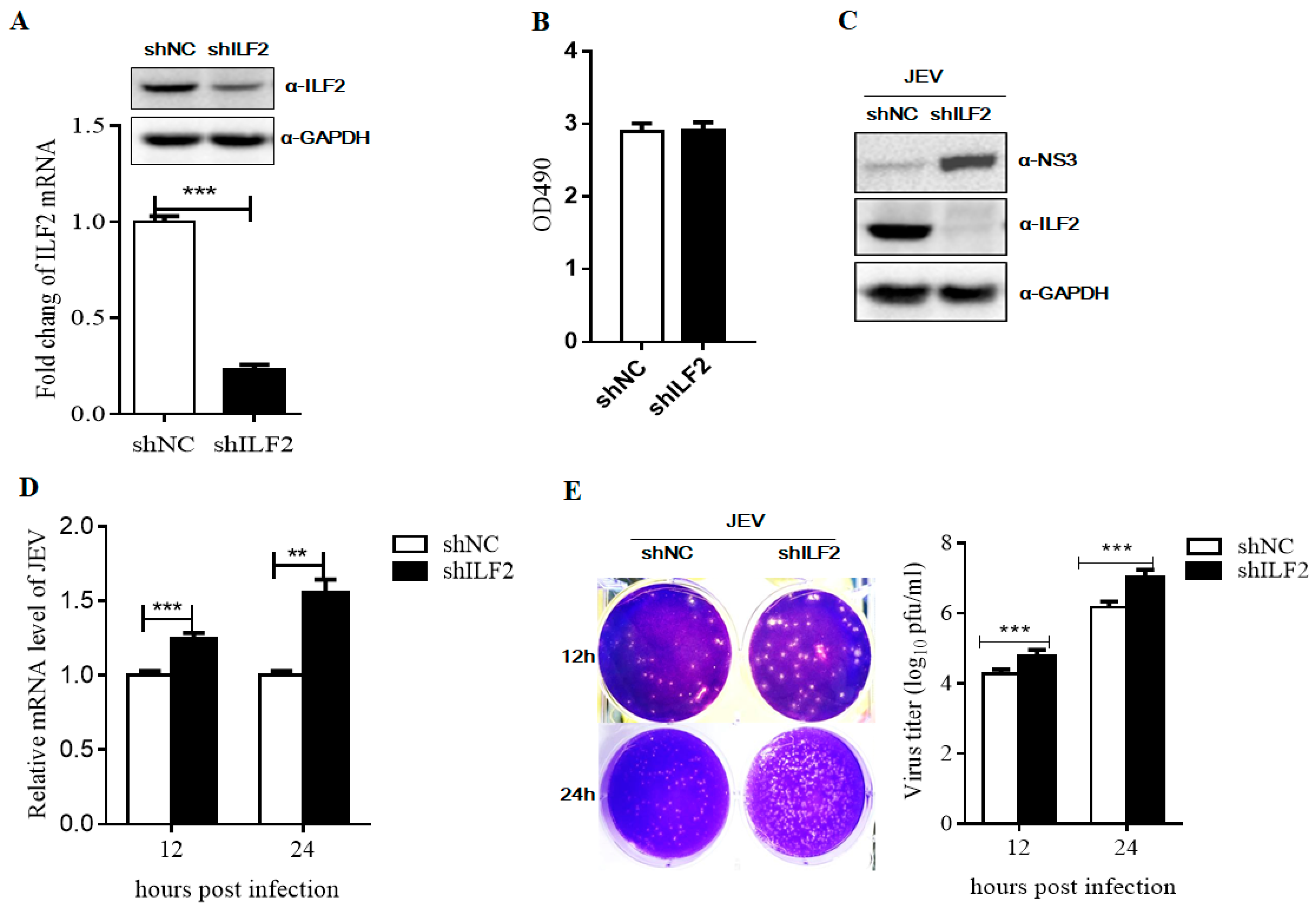
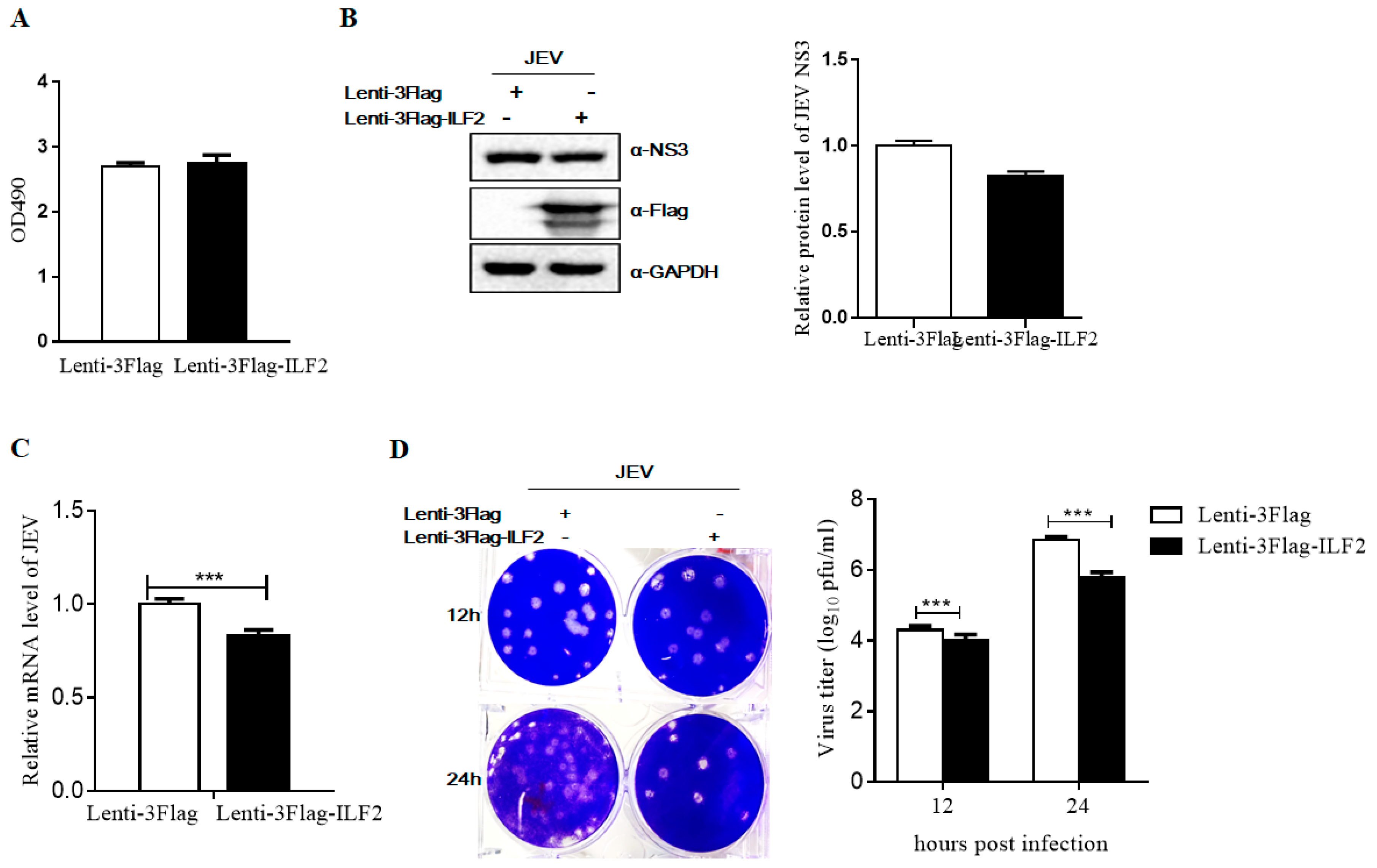
3.5. ILF2 Inhibits JEV Replication
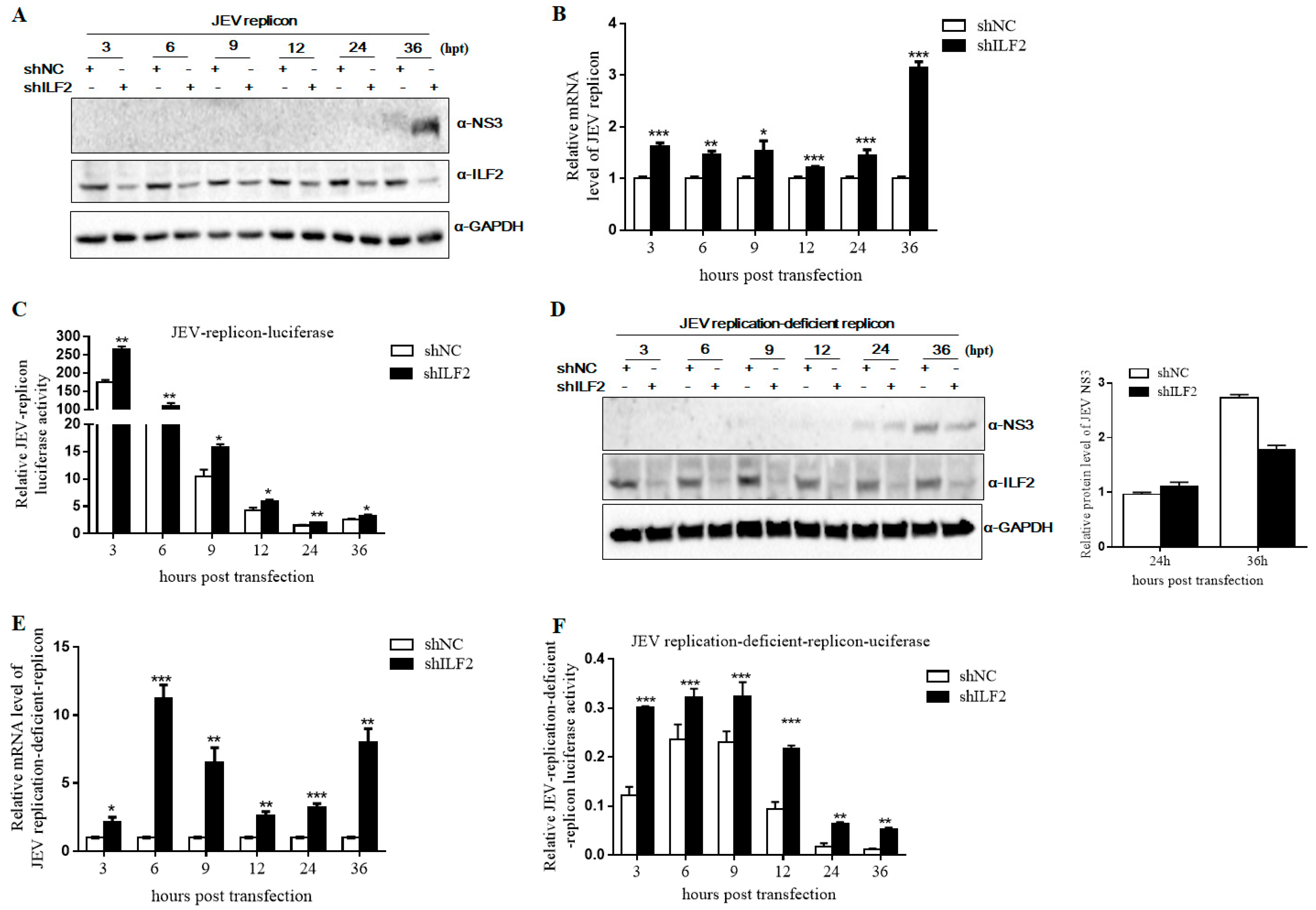
3.6. Knockdown of ILF2 Positively Regulates JEV Replication by Sub-Genomic Replicon System Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Misra, U.K.; Kalita, J. Overview: Japanese encephalitis. Prog. Neurobiol. 2010, 91, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Neyts, J.; Leyssen, P.; De Clercq, E. Infections with flaviviridae. Verh K Acad Geneeskd Belg 1999, 61, 661–697. [Google Scholar] [PubMed]
- Unni, S.K.; Růžek, D.; Chhatbar, C.; Mishra, R.; Johri, M.K.; Singh, S.K. Japanese encephalitis virus: From genome to infectome. Microbes Infect. 2011, 13, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Gong, P. A structural view of the RNA-dependent RNA polymerases from the Flavivirus genus. Virus Res. 2017, 234, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Unno, H.; Mori, Y.; Tani, H.; Moriishi, K.; Takamizawa, A.; Agoh, M.; Tsukihara, T.; Matsuura, Y. Crystal structure of the catalytic domain of Japanese encephalitis virus NS3helicase/nucleoside triphosphatase at a resolution of 1.8 A. Virology 2008, 373, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Yiang, G.T.; Chen, Y.H.; Chou, P.L.; Chang, W.J.; Wei, C.W.; Yu, Y.L. The NS3 protease and helicase domains of Japanese encephalitis virus trigger cell death via caspase-dependent and -independent pathways. Mol. Med. Rep. 2013, 7, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Kuo, M.D.; Chien, L.J.; Hsu, S.L.; Wang, Y.M.; Lin, J.H. RNA-protein interactions: Involvement of NS3, NS5, and 3′ noncoding regions of Japaneseencephalitis virus genomic RNA. J. Virol. Virol. 1997, 71, 3466–3473. [Google Scholar]
- Uchil, P.D.; Satchidanandam, V. Architecture of the flaviviral replication complex. Protease, nuclease, and detergents revealencasement within double-layered membrane compartments. J. Biol. Chem. 2003, 278, 24388–24398. [Google Scholar] [CrossRef]
- Ghosh, D.; Basu, A. Japanese encephalitis-a pathological and clinical perspective. PLoS Negl. Trop. Dis. 2009, 3, e437. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of Japanese encephalitis virus. Annu Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef]
- Olsen, S.J.; Supawat, K.; Campbell, A.P.; Anantapreecha, S.; Liamsuwan, S.; Tunlayadechanont, S.; Visudtibhan, A.; Lupthikulthum, S.; Dhiravibulya, K.; Viriyavejakul, A.; et al. Japanese encephalitis virus remains an important cause of encephalitis in Thailand. Int. J. Infect. Dis. 2010, 14, e888-92. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hou, L.; Du, J.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. Capsid, membrane and NS3 are the major viral proteins involved in autophagy induced by Japanese encephalitis virus. Vet. Microbiol. 2015, 178, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774, 774A–774E. [Google Scholar] [CrossRef] [PubMed]
- Heffelfinger, J.D.; Li, X.; Batmunkh, N.; Grabovac, V.; Diorditsa, S.; Liyanage, J.B.; Pattamadilok, S.; Bahl, S.; Vannice, K.S.; Hyde, T.B.; et al. Japanese Encephalitis Surveillance and Immunization - Asia and Western Pacific Regions, 2016. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Hegde, N.R.; Gore, M.M. Japanese encephalitis vaccines: Immunogenicity, protective efficacy, effectiveness, and impact on the burden of disease. Hum. Vaccin. Immunother. 2017, 13, 1–18. [Google Scholar] [CrossRef]
- Boldescu, V.; Behnam, M.A.M.; Vasilakis, N.; Klein, C.D. Broad-spectrum agents for flaviviral infections: Dengue, Zika and beyond. Nat. Rev. Drug Discov. 2017, 16, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Kaczor, A.; Matosiuk, D. Structure-based virtual screening for novel inhibitors of Japanese encephalitis virus NS3helicase/nucleoside triphosphatase. FEMS Immunol. Med. Microbiol. 2010, 58, 91–101. [Google Scholar] [CrossRef]
- Guan, D.; Altan-Bonnet, N.; Parrott, A.M.; Arrigo, C.J.; Li, Q.; Khaleduzzaman, M.; Li, H.; Lee, C.G.; Pe’ery, T.; Mathews, M.B. Nuclear factor 45 (NF45) is a regulatory subunit of complexes with NF90/110 involved in mitotic control. Mol. Cell. Biol. 2008, 28, 4629–4641. [Google Scholar] [CrossRef]
- Higuchi, T.; Todaka, H.; Sugiyama, Y.; Ono, M.; Tamaki, N.; Hatano, E.; Takezaki, Y.; Hanazaki, K.; Miwa, T.; Lai, S.; et al. Suppression of MicroRNA-7 (miR-7) Biogenesis by Nuclear Factor 90-Nuclear Factor 45Complex (NF90-NF45) Controls Cell Proliferation in Hepatocellular Carcinoma. J. Biol. Chem. 2016, 291, 21074–21084. [Google Scholar] [CrossRef]
- Kiesler, P.; Haynes, P.A.; Shi, L.; Kao, P.N.; Wysocki, V.H.; Vercelli, D. NF45 and NF90 regulate HS4-dependent interleukin-13 transcription in T cells. J. Biol. Chem. 2010, 285, 8256–8267. [Google Scholar] [CrossRef]
- Marchesini, M.; Ogoti, Y.; Fiorini, E.; Aktas Samur, A.; Nezi, L.; D’Anca, M.; Storti, P.; Samur, M.K.; Ganan-Gomez, I.; Fulciniti, M.T.; et al. ILF2 Is a Regulator of RNA Splicing and DNA Damage Response in 1q21-Amplified MultipleMyeloma. Cancer Cell 2017, 32, 88–100.e6. [Google Scholar] [CrossRef] [PubMed]
- Rigo, F.; Hua, Y.; Chun, S.J.; Prakash, T.P.; Krainer, A.R.; Bennett, C.F. Synthetic oligonucleotides recruit ILF2/3 to RNA transcripts to modulate splicing. Nat. Chem. Biol. 2012, 8, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamanna, R.A.; Hoque, M.; Lewis-Antes, A.; Azzam, E.I.; Lagunoff, D.; Pe’ery, T.; Mathews, M.B. The NF90/NF45 complex participates in DNA break repair via nonhomologous end joining. Mol. Cell. Biol. 2011, 31, 4832–4843. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.; Zhu, J.; Zhang, J.; Zhang, S.; Li, M.; Ni, R.; Liu, J.; Qiu, H.; Chen, W.; Wang, H.; et al. Expression and clinical role of NF45 as a novel cell cycle protein in esophageal squamouscell carcinoma (ESCC). Tumour Biol. 2015, 36, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Jiang, X.; Ding, C.; Du, C.; Owusu-Ansah, K.G.; Weng, X.; Hu, W.; Peng, C.; Lv, Z.; Tong, R.; et al. Expression and Critical Role of Interleukin Enhancer Binding Factor 2 in HepatocellularCarcinoma. Int. J. Mol. Sci. 2016, 17, 1373. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Yang, Y.; Yu, J.; Meng, F.; Zhang, T.; Gao, Y. The long noncoding RNA LINC00473, a target of microRNA 34a, promotes tumorigenesis by inhibiting ILF2 degradation in cervical cancer. Am. J. Cancer Res. 2017, 7, 2157–2168, eCollection 2017. [Google Scholar] [PubMed]
- Isken, O.; Baroth, M.; Grassmann, C.W.; Weinlich, S.; Ostareck, D.H.; Ostareck-Lederer, A.; Behrens, S.E. Nuclear factors are involved in hepatitis C virus RNA replication. RNA 2007, 13, 1675–1692. [Google Scholar] [CrossRef] [Green Version]
- Isken, O.; Grassmann, C.W.; Sarisky, R.T.; Kann, M.; Zhang, S.; Grosse, F.; Kao, P.N.; Behrens, S.E. Members of the NF90/NFAR protein group are involved in the life cycle of a positive-strandRNA virus. EMBO J. 2003, 22, 5655–5665. [Google Scholar] [CrossRef]
- Lee, J.W.; Liao, P.C.; Young, K.C.; Chang, C.L.; Chen, S.S.; Chang, T.T.; Lai, M.D.; Wang, S.W. Identification of hnRNPH1, NF45, and C14orf166 as novel host interacting partners of the mature hepatitis C virus core protein. J. Proteome Res. 2011, 10, 4522–4534. [Google Scholar] [CrossRef]
- Li, Y.; Belshan, M. NF45 and NF90 Bind HIV-1 RNA and Modulate HIV Gene Expression. Viruses 2016, 8, 47. [Google Scholar] [CrossRef]
- Shin, H.J.; Kim, S.S.; Cho, Y.H.; Lee, S.G.; Rho, H.M. Host cell proteins binding to the encapsidation signal epsilon in hepatitis B virus RNA. Arch. Virol. Virol. 2002, 147, 471–491. [Google Scholar] [CrossRef]
- Stricker, R.L.; Behrens, S.E.; Mundt, E. Nuclear factor NF45 interacts with viral proteins of infectious bursal disease virus and inhibits viral replication. J. Virol. Virol. 2010, 84, 10592–10605. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Bian, T.; Zhang, Z.; Zhou, L.; Ge, X.; Han, J.; Guo, X.; Yang, H.; Yu, K. Interleukin-2 enhancer binding factor 2 interacts with the nsp9 or nsp2 of porcinereproductive and respiratory syndrome virus and exerts negatively regulatory effect on the viral replication. Virol. J. 2017, 14, 125. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Li, A.; Wang, W.; Wu, J. Interleukin-enhanced binding factor 2 interacts with NLRP3 to inhibit the NLRP3 inflammasome activation. Biochem. Biophys. Res. Commun. 2018, 500, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.K.; Lin, Y.L.; Liao, C.L.; Lin, C.G.; Huang, Y.L.; Yeh, C.T.; Lai, S.C.; Jan, J.T.; Chin, C. Generation and characterization of organ-tropism mutants of Japanese encephalitis virus in vivo and in vitro. Virology 1996, 223, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Chien, H.L.; Liao, C.L.; Lin, Y.L. FUSE binding protein 1 interacts with untranslated regions of Japanese encephalitis virus RNA and negatively regulates viral replication. J. Virol. 2011, 85, 4698–4706. [Google Scholar] [CrossRef]
- Lin, C.W.; Huang, H.D.; Shiu, S.Y.; Chen, W.J.; Tsai, M.H.; Huang, S.H.; Wan, L.; Lin, Y.J. Functional determinants of NS2B for activation of Japanese encephalitis virus NS3 protease. Virus Res. 2007, 127, 88–94. [Google Scholar] [CrossRef]
- Martins, N.; Imler, J.L.; Meignin, C. Discovery of novel targets for antivirals: Learning from flies. Curr. Opin Virol. 2016, 20, 64–70. [Google Scholar] [CrossRef]
- Chiou, C.T.; Hu, C.C.; Chen, P.H.; Liao, C.L.; Lin, Y.L.; Wang, J.J. Association of Japanese encephalitis virus NS3 protein with microtubules and tumoursusceptibility gene 101 (TSG101) protein. J. Gen. Virol. 2003, 84, 2795–2805. [Google Scholar] [CrossRef]
- Chiu, H.P.; Chiu, H.; Yang, C.F.; Lee, Y.L.; Chiu, F.L.; Kuo, H.C.; Lin, R.J.; Lin, Y.L. Inhibition of Japanese encephalitis virus infection by the host zinc-finger antiviral protein. PLoS Pathog. 2018, 14, e1007166. [Google Scholar] [CrossRef]
- Li, C.; Ge, L.L.; Li, P.P.; Wang, Y.; Sun, M.X.; Huang, L.; Ishag, H.; Di, D.D.; Shen, Z.Q.; Fan, W.X.; et al. The DEAD-box RNA helicase DDX5 acts as a positive regulator of Japanese encephalitisvirus replication by binding to viral 3′ UTR. Antiviral Res. 2013, 100, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ge, L.L.; Li, P.P.; Wang, Y.; Dai, J.J.; Sun, M.X.; Huang, L.; Shen, Z.Q.; Hu, X.C.; Ishag, H.; et al. Cellular DDX3 regulates Japanese encephalitis virus replication by interacting with viral un-translated regions. Virology 2014, 449, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, F.; Zhang, J.W.; Li, W.; Zhao, D.M.; Wang, H.; Hua, R.H.; Bu, Z.G. Host Factor SPCS1 Regulates the Replication of Japanese Encephalitis Virus through Interactions with Transmembrane Domains of NS2B. J. Virol. 2018, 92, e00197-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.K.; Chai, F.; Li, H.Y.; Xiao, G.; Guo, L. Identification of host proteins involved in Japanese encephalitis virus infection by quantitative proteomics analysis. J. Proteome Res. 2013, 12, 2666–2678. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Miner, J.J.; Gorman, M.J.; Rausch, K.; Ramage, H.; White, J.P.; Zuiani, A.; Zhang, P.; Fernandez, E.; Zhang, Q.; et al. A CRISPR screen defines a signal peptide processing pathway required by flaviviruses. Nature 2016, 535, 164–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Wu, R.; Liu, H.; Wen, X.; Huang, X.; Wen, Y.; Ma, X.; Yan, Q.; Huang, Y.; Zhao, Q.; et al. The NS3 and NS4A genes as the targets of RNA interference inhibit replication of Japaneseencephalitis virus in vitro and in vivo. Gene 2016, 594, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, N.R.; Maizel, A.L.; Wang, F.; Sharma, S. Comparative analysis of NFAT (nuclear factor of activated T cells) complex in human T and Blymphocytes. J. Biol. Chem. 1993, 268, 14285–14293. [Google Scholar]
- Isken, O.; Grassmann, C.W.; Yu, H.; Behrens, S.E. Complex signals in the genomic 3′ nontranslated region of bovine viral diarrhea viruscoordinate translation and replication of the viral RNA. RNA 2004, 10, 1637–1652. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Oligonucleotide Sequence (5′–3′) |
|---|---|
| pCAGGS-HA-ILF2-F | GAATTCATGAGGGGTGACAGAGGC |
| pCAGGS-HA-ILF2-R | CTCGAGCTCCTGAGTTTCCATGC |
| pTRIP-ILF2-3Flag-F | ACTAGTATGAGGGGTGACAGAGGCCGTGG |
| pTRIP-ILF2-3Flag-R | GTCGACCTCCTGAGTTTCCATGCTTTC |
| pCAGGS-HA-Firefly-F | GAATTCATGGAAGATGCCAAAAACATTAAG |
| pCAGGS-HA-Firefly-R | CTCGAGCACGGCGATCTTGCCGCCCTTC |
| qILF2-F | AGTCGTGGAAAGCCTAAGAG |
| qILF2-R | GGTGGCACTGTTGTAATGAG |
| qFirefiy-F | GGCCCCAGCTAACGACATCTAC |
| qFirefly-R | GCAGCCCTTTCTTGCTCACG |
| qGAPDH-F | CACCCACTCCTCCACCTTTG |
| qGAPDH-R | CCACCACCCTGTTGCTGTAG |
| qJEV-C-F | GAGCTTGTTGGACGGCAGAG |
| qJEV-C-R | CACGGCGTCGATGAGTGTTC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, X.; Qian, P.; Rao, T.; Wei, Y.; Zhao, F.; Zhang, H.; Chen, H.; Li, X. Cellular Interleukin Enhancer-Binding Factor 2, ILF2, Inhibits Japanese Encephalitis Virus Replication In Vitro. Viruses 2019, 11, 559. https://0-doi-org.brum.beds.ac.uk/10.3390/v11060559
Cui X, Qian P, Rao T, Wei Y, Zhao F, Zhang H, Chen H, Li X. Cellular Interleukin Enhancer-Binding Factor 2, ILF2, Inhibits Japanese Encephalitis Virus Replication In Vitro. Viruses. 2019; 11(6):559. https://0-doi-org.brum.beds.ac.uk/10.3390/v11060559
Chicago/Turabian StyleCui, Xiaofang, Ping Qian, Tingting Rao, Yanming Wei, Fang Zhao, Huawei Zhang, Huanchun Chen, and Xiangmin Li. 2019. "Cellular Interleukin Enhancer-Binding Factor 2, ILF2, Inhibits Japanese Encephalitis Virus Replication In Vitro" Viruses 11, no. 6: 559. https://0-doi-org.brum.beds.ac.uk/10.3390/v11060559