Frequency of Hepatitis B Virus Resistance Mutations in Women Using Tenofovir Gel as Pre-Exposure Prophylaxis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants/Samples
2.2. Isolation and Amplification of HBV DNA
2.3. Sequencing of the HBV Pol Region
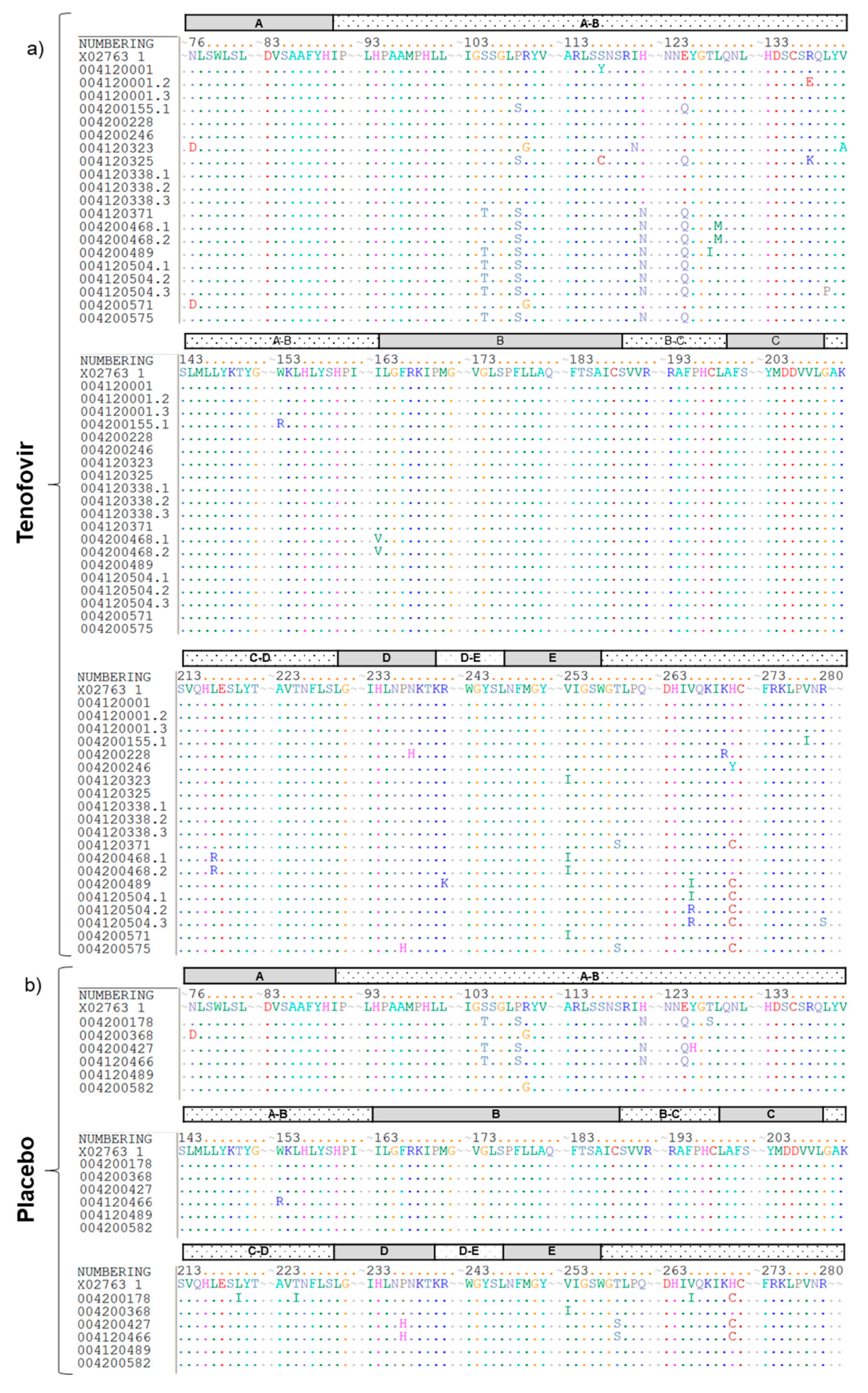
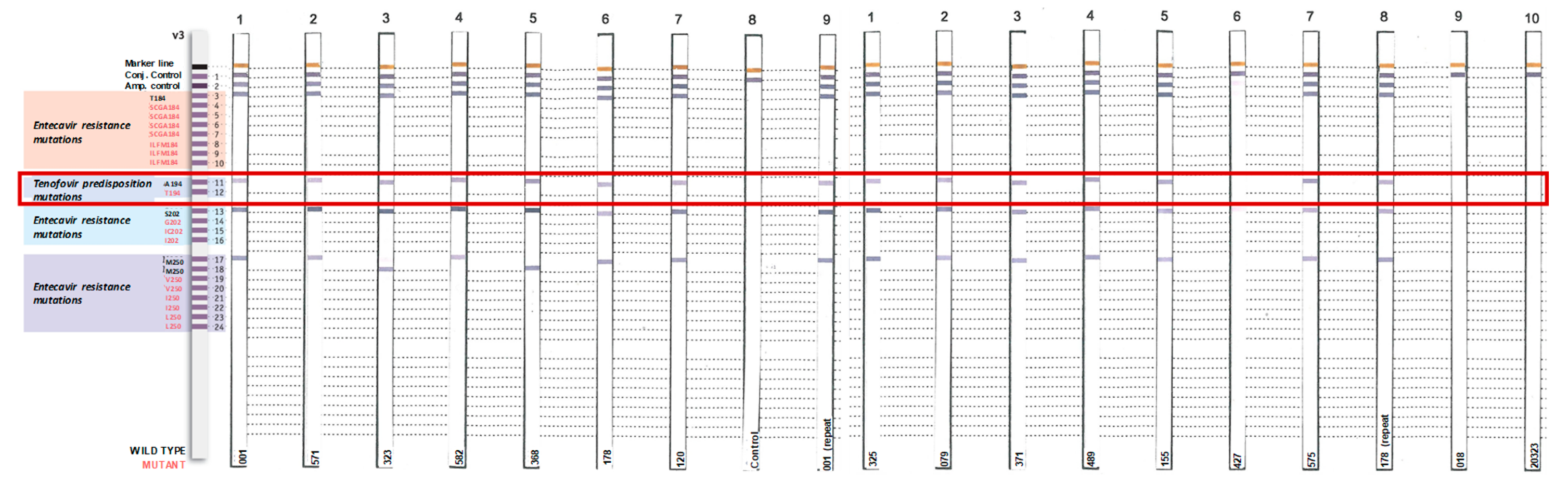
2.4. Identification of Tenofovir-Resistance Mutations
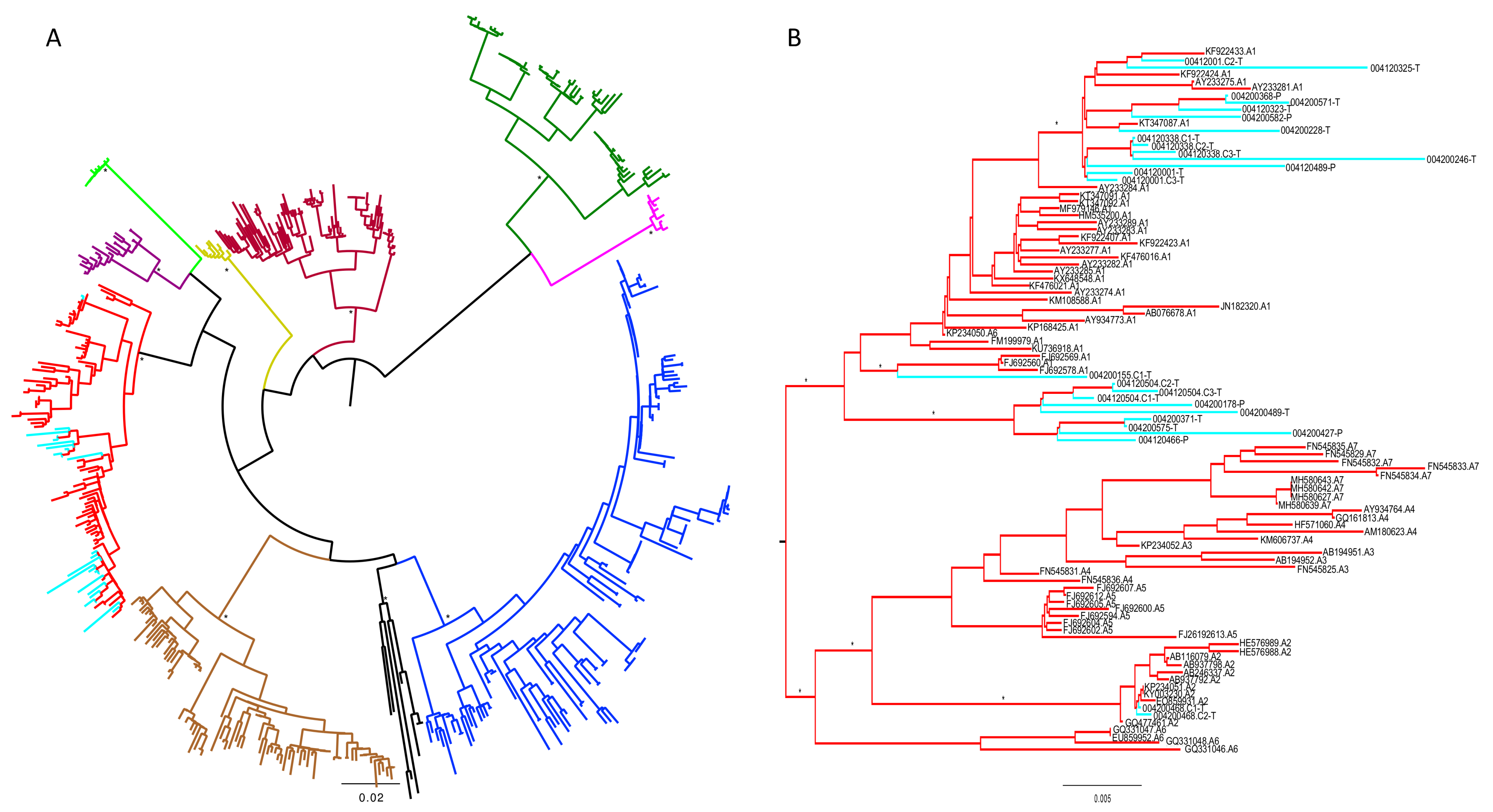
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- World Health Organisation. Global Hepatitis Report: 2017; World Health Organisation: Geneva, Switzerland, 2017; Available online: http://apps.who.int/iris/bitstream/handle/10665/255016/9789241565455-eng.pdf;jsessionid=5FECB02DFFA32EF581D009E369C9B9A5?sequence=1 (accessed on 14 November 2018).
- World Health Organisation. Hepatitis B. Key facts; World Health Organisation: Geneva, Switzerland, 2017; Available online: http://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 14 June 2018).
- Abdool Karim, Q.; Abdool Karim, S.S.; Frohlich, J.A.; Grobler, A.C.; Baxter, C.; Mansoor, L.E.; Kharsany, A.B.M.; Sibeko, S.; Mlisana, K.P.; Omar, Z.; et al. Effectiveness and Safety of Tenofovir Gel, an Antiretroviral Microbicide, for the Prevention of HIV Infection in Women. Science 2010, 329, 1168–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, R.M.; Lama, J.R.; Anderson, P.L.; McMahan, V.; Liu, A.Y.; Vargas, L.; Goicochea, P.; Casapia, M.; Guanira-Carranza, J.V.; Ramirez-Cardich, M.E.; et al. Preexposure chemoprophylaxis for HIV prevention in men who have sex with men. N. Engl. J. Med. 2010, 363, 2587–2599. [Google Scholar] [CrossRef] [PubMed]
- Baeten, J.M.; Donnell, D.; Ndase, P.; Mugo, N.R.; Campbell, J.D.; Wangisi, J.; Tappero, J.W.; Bukusi, E.A.; Cohen, C.R.; Katabira, E.; et al. Antiretroviral prophylaxis for HIV prevention in heterosexual men and women. N. Engl. J. Med. 2012, 367, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Hojilla, J.C.; Vlahov, D.; Crouch, P.C.; Dawson-Rose, C.; Freeborn, K.; Carrico, A. HIV Pre-exposure Prophylaxis (PrEP) Uptake and Retention Among Men Who Have Sex with Men in a Community-Based Sexual Health Clinic. AIDS Behav. 2018, 22, 1096–1099. [Google Scholar] [CrossRef] [PubMed]
- Snowden, J.M.; Chen, Y.H.; McFarland, W.; Raymond, H.F. Prevalence and characteristics of users of pre-exposure prophylaxis (PrEP) among men who have sex with men, San Francisco, 2014 in a cross-sectional survey: Implications for disparities. Sex. Transm. Infect. 2017, 93, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, B.; Moreira, R.I.; De Boni, R.B.; Kallas, E.G.; Madruga, J.V.; Vasconcelos, R.; Goulart, S.; Torres, T.S.; Marins, L.M.S.; Anderson, P.L.; et al. High pre-exposure prophylaxis uptake and early adherence among men who have sex with men and transgender women at risk for HIV Infection: The PrEP Brasil demonstration project. J. Int. AIDS Soc. 2017, 20, 21472. [Google Scholar] [CrossRef] [PubMed]
- Dienstag, J.L. Hepatitis B virus infection. N. Engl. J. Med. 2008, 359, 1486–1500. [Google Scholar] [CrossRef] [PubMed]
- Snow-Lampart, A.; Chappell, B.; Curtis, M.; Zhu, Y.; Myrick, F.; Schawalder, J.; Kitrinos, K.; Svarovskaia, E.S.; Miller, M.D.; Sorbel, J.; et al. No resistance to tenofovir disoproxil fumarate detected after up to 144 weeks of therapy in patients monoinfected with chronic hepatitis B virus. Hepatology 2011, 53, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Heathcote, E.J.; Marcellin, P.; Buti, M.; Gane, E.; De Man, R.A.; Krastev, Z.; Germanidis, G.; Lee, S.S.; Flisiak, R.; Kaita, K.; et al. Three-Year Efficacy and Safety of Tenofovir Disoproxil Fumarate Treatment for Chronic Hepatitis B. Gastroenterology 2011, 140, 132–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, C.; Yende-Zuma, N.; Tshabalala, P.; Abdool Karim, Q.; Abdool Karim, S.S. Safety of coitally administered tenofovir 1% gel, a vaginal microbicide, in chronic hepatitis B virus carriers: Results from the CAPRISA 004 trial. Antivir. Res. 2013, 99, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evol. Int. J. Org. Evol. 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Davila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, C.; Chen, B.A.; Guddera, V.; Hoesley, C.; Justman, J.; Nakabiito, C.; Salata, R.; Soto-Torres, L.; Patterson, K.; Minnis, A.M.; et al. MTN-001: Randomized Pharmacokinetic Cross-Over Study Comparing Tenofovir Vaginal Gel and Oral Tablets in Vaginal Tissue and Other Compartments. PLoS ONE 2013, 8, e55013. [Google Scholar] [CrossRef] [PubMed]
- Kitrinos, K.M.; Corsa, A.; Liu, Y.; Flaherty, J.; Snow-Lampart, A.; Marcellin, P.; Borroto-Esoda, K.; Miller, M.D. No detectable resistance to tenofovir disoproxil fumarate after 6 years of therapy in patients with chronic hepatitis B. Hepatology 2013, 59, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, L.; Corneli, A.; Ahmed, K.; Agot, K.; Lombaard, J.; Kapiga, S.; Malahleha, M.; Owino, F.; Manongi, R.; Onyango, J.; et al. For the FEM-PrEP Study Group, Preexposure Prophylaxis for HIV Infection among African Women. N. Engl. J. Med. 2012, 367, 411–422. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. HIV Drug Resistance Report; World Health Organisation: Geneva, Switzerland, 2017. [Google Scholar]
- Sharma, V.; Sharma, R.; Gaurav, G. First Report of Primary Tenofovir Resistance in a Hepatitis B Viral Hepatitis Patient from India without Human Immunodeficiency Virus Co-infection. J. Liver Res. Disord. Ther. 2016, 2, 00037. [Google Scholar] [CrossRef]


| Mutation Category | Tenofovir Arm (n = 20) | Placebo Arm (n = 6) |
|---|---|---|
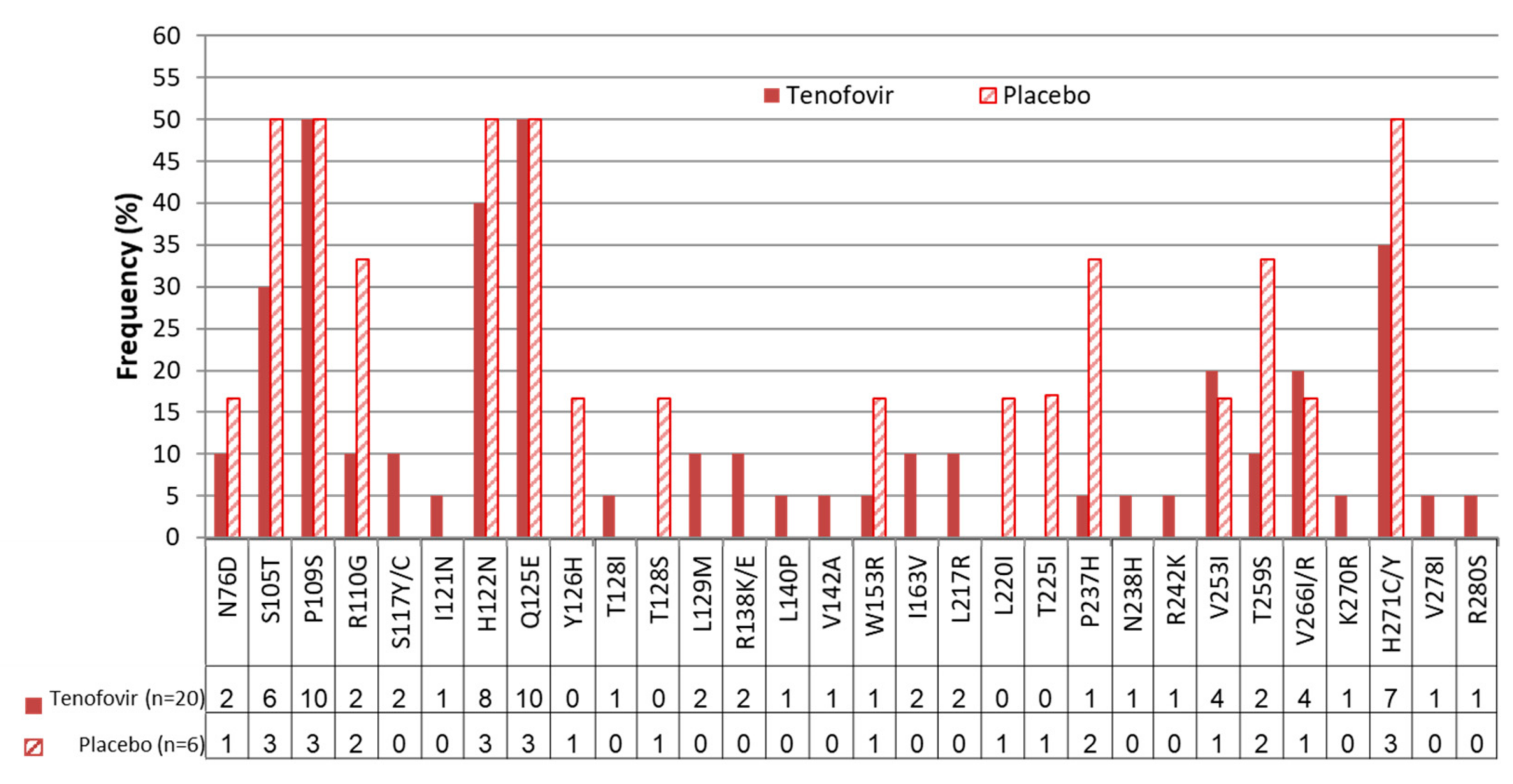
| rt Mutation | N76D(2), S105T(6), P109S(10), R110G(2), S117Y(1), S117C(1), I121N(1), N122H(8), Q125E(10), T128I(1), L129M(2), R138K(1), R138E(1), L140P(1), V142A(1), W153R(1), I163V(2), L217R(2), P237H(1), N238H(1), R242K(1), V253I(4), T259S(2), V266I(2), V266R(2), K270R(1), H271C(6), H271Y(1), V278I(1), R280S(1) | N76D(1), S105T(3), P109S(3), R110G(2), H122N(3), E125Q(3), Y126H(1), T128S(1), W153R(1), L220I(1), T225I(1), P237H(2), V253I(1), T259S(2), V266I(1), H271C(3) |
| Primary Tenofovir Resistance Mutation | None | None |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall | Tenofovir (N = 20) | Placebo (N = 17) | p Value | |
|---|---|---|---|---|
| Mean age, years (SD) | 24.7 (5.73) | 25.15 (5.84) | 24.2 (5.74) | 0.64 |
| % HBeAg positive (n/N) | 10.8 (4/37) | 10.0 (2/20) | 11.8 (2/17) | 0.63 |
| Median HBV DNA*Log10 copies/mL (IQR)$ | 2.63 (1.1–3.0) | 2.61 (1.1–3.2) | 1.83 (0.0–3.0) | 0.52 |
| Median ALT* (IU/L) (IQR) | 20.0 (15.0–22.0) | 19.5 (15.8–22.0) | 21.0 (15.0–23.0) | 0.66 |
| Median AST* (IU/L) (IQR) | 22.0 (19.0–25.0) | 23.5 (19.8–25.3) | 20.0 (19.0–24.0) | 0.31 |
| % with any grade 2^ or higher liver related adverse event during follow up (n/N) | 18.9 (7/37) | 10.0 (2/20) | 29.4 (5/17) | 0.21 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baxter, C.; Ngcapu, S.; Blackard, J.T.; Powell, E.A.; Penton, P.K.; Abdool Karim, S.S. Frequency of Hepatitis B Virus Resistance Mutations in Women Using Tenofovir Gel as Pre-Exposure Prophylaxis. Viruses 2019, 11, 569. https://0-doi-org.brum.beds.ac.uk/10.3390/v11060569
Baxter C, Ngcapu S, Blackard JT, Powell EA, Penton PK, Abdool Karim SS. Frequency of Hepatitis B Virus Resistance Mutations in Women Using Tenofovir Gel as Pre-Exposure Prophylaxis. Viruses. 2019; 11(6):569. https://0-doi-org.brum.beds.ac.uk/10.3390/v11060569
Chicago/Turabian StyleBaxter, Cheryl, Sinaye Ngcapu, Jason T Blackard, Eleanor A Powell, Patricia K Penton, and Salim S Abdool Karim. 2019. "Frequency of Hepatitis B Virus Resistance Mutations in Women Using Tenofovir Gel as Pre-Exposure Prophylaxis" Viruses 11, no. 6: 569. https://0-doi-org.brum.beds.ac.uk/10.3390/v11060569