Gray (Oreochromis niloticus x O. aureus) and Red (Oreochromis spp.) Tilapia Show Equal Susceptibility and Proinflammatory Cytokine Responses to Experimental Tilapia Lake Virus Infection

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. RNA Extraction and cDNA Synthesis
2.3. Quantification of TiLV by Quantitative Real Time PCR in Tissue Samples
2.4. Gene Expression Analysis
2.5. Statistical Analysis
3. Results
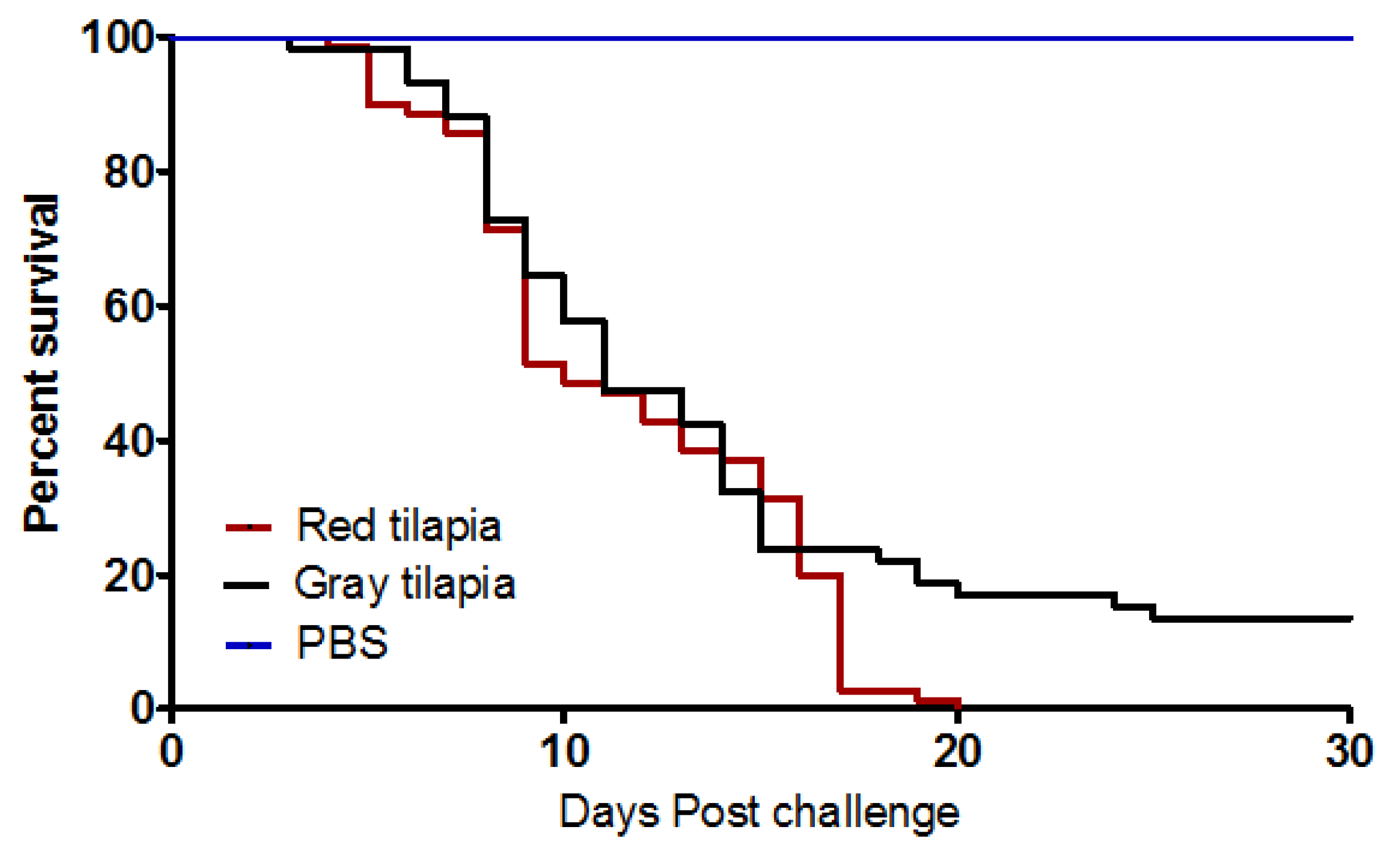
3.1. Clinical Observations and Kaplan Meyer’s Survival Analysis
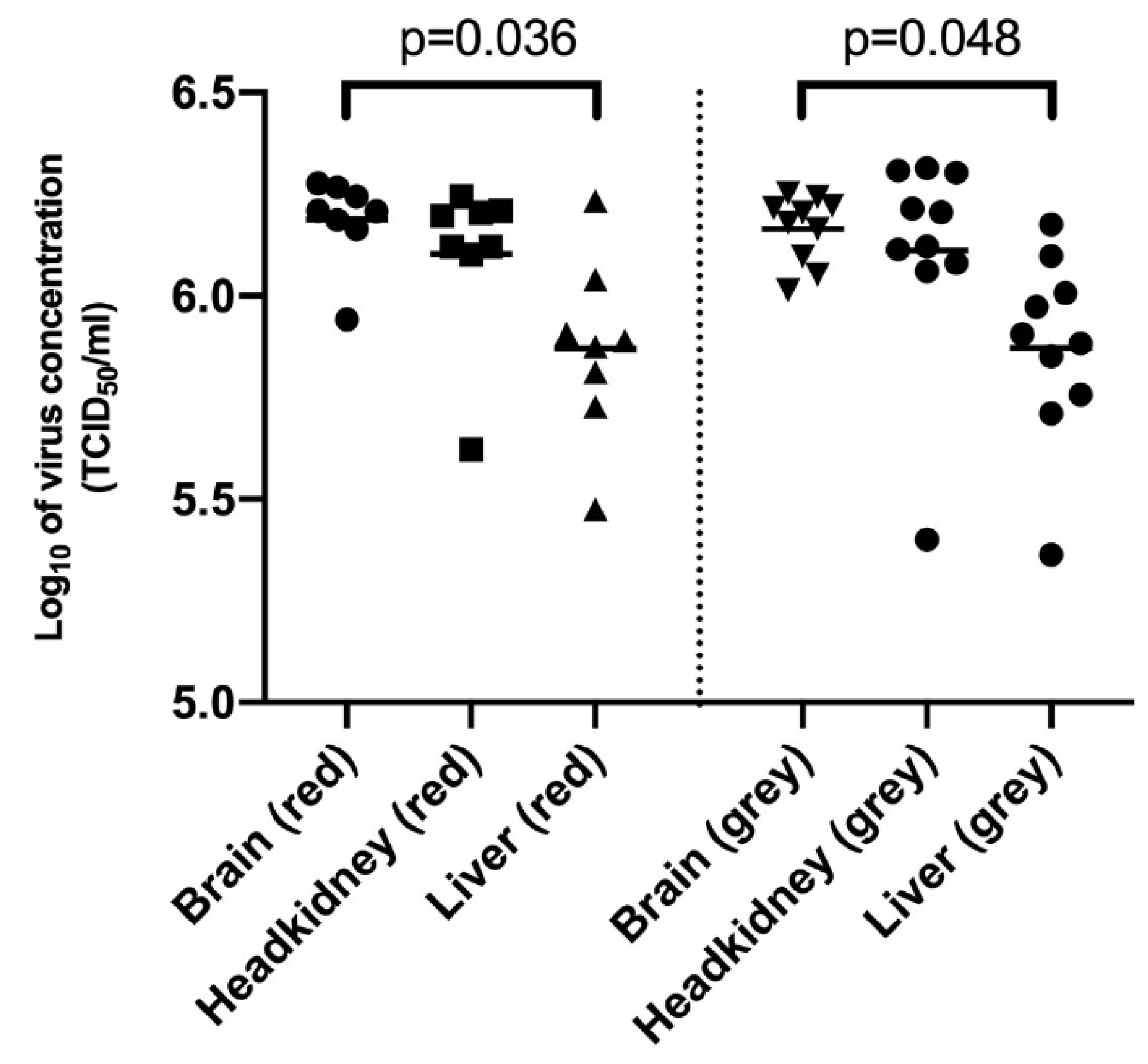
3.2. Comparison of Virus Concentration in Brain, Headkidney and Liver Samples
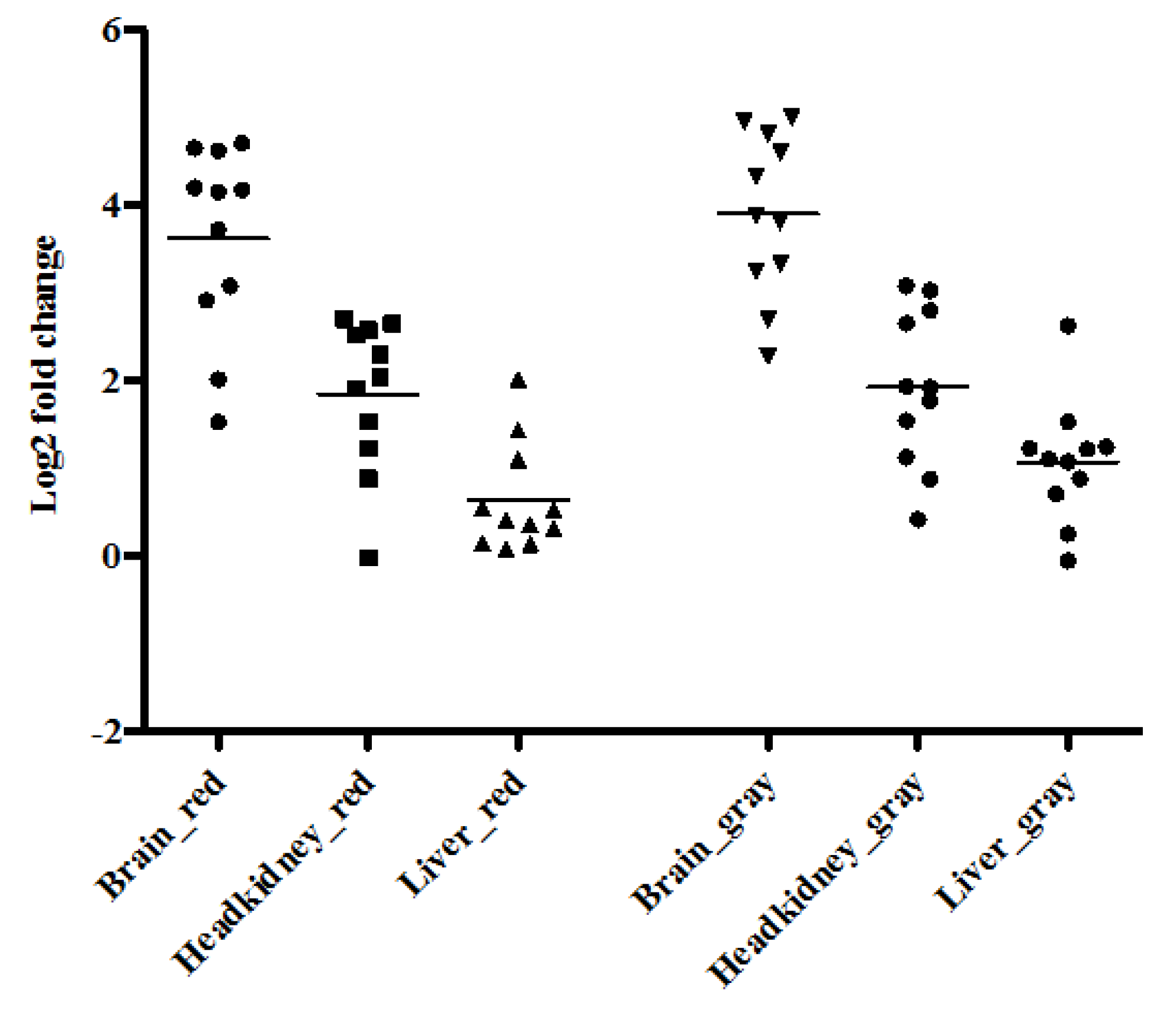
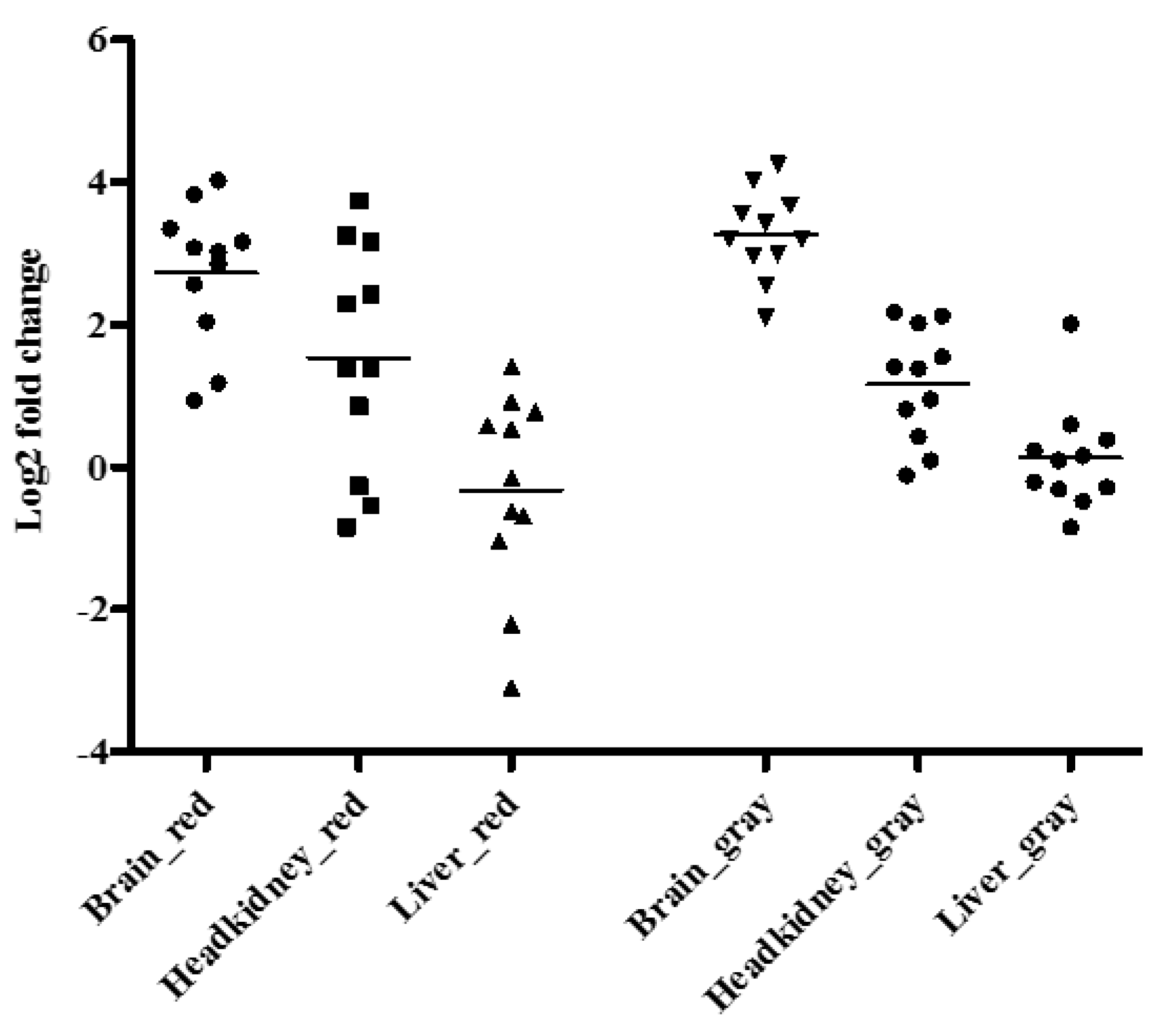
3.3. Gene Expression
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. F.a.A.O. State of the World Fisheries and Aquaculture; Food and Agriculture Organization, United Nations: Rome, Italy, 2018. [Google Scholar]
- Machimbirike, V.I.; Jansen, M.D.; Senapin, S.; Khunrae, P.; Rattanarojpong, T.; Dong, H.T. Viral infections in tilapines: More than just tilapia lake virus. Aquaculture 2019, 503, 508–518. [Google Scholar] [CrossRef]
- Eyngor, M.; Zamostiano, R.; Tsofack, J.E.K.; Berkowitz, A.; Bercovier, H.; Tinman, S.; Lev, M.; Hurvitz, A.; Galeotti, M.; Bacharach, E. Identification of a novel rna virus lethal to tilapia. J. Clin. Microbiol. 2014, 52, 4137–4146. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, H.; Kabuusu, R.; Beltran, S.; Reyes, E.; Lince, J.; del Pozo, J. Syncytial hepatitis of farmed tilapia, o reochromis niloticus (l.): A case report. J. Fish Dis. 2014, 37, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Bacharach, E.; Mishra, N.; Briese, T.; Zody, M.C.; Tsofack, J.E.K.; Zamostiano, R.; Berkowitz, A.; Ng, J.; Nitido, A.; Corvelo, A. Characterization of a novel orthomyxo-like virus causing mass die-offs of tilapia. MBio 2016, 7, e00431-16. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the international code of virus classification and nomenclature ratified by the international committee on taxonomy of viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.; Pradhan, P.; Swaminathan, T.; Sood, N.; Paria, P.; Das, A.; Verma, D.; Kumar, R.; Yadav, M.; Dev, A. Emergence of tilapia lake virus associated with mortalities of farmed nile tilapia oreochromis niloticus (linnaeus 1758) in India. Aquaculture 2018, 484, 168–174. [Google Scholar] [CrossRef]
- Dong, H.; Ataguba, G.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Evidence of tilv infection in tilapia hatcheries from 2012 to 2017 reveals probable global spread of the disease. Aquaculture 2017, 479, 579–583. [Google Scholar] [CrossRef]
- Surachetpong, W.; Janetanakit, T.; Nonthabenjawan, N.; Tattiyapong, P.; Sirikanchana, K.; Amonsin, A. Outbreaks of tilapia lake virus infection, Thailand, 2015–2016. Emerg. Infect. Dis. 2017, 23, 1031. [Google Scholar] [CrossRef]
- Pulido, L.L.H.; Chìo, M.M.; Chaparro, A.L.H.; Dong, H.T.; Senapin, S. Tilapia lake virus (tilv) from peru is genetically close to the israeli isolates. Aquaculture 2019, 510, 61–65. [Google Scholar] [CrossRef]
- Fathi, M.; Dickson, C.; Dickson, M.; Leschen, W.; Baily, J.; Muir, F.; Ulrich, K.; Weidmann, M. Identification of tilapia lake virus in Egypt in nile tilapia affected by ‘summer mortality’ syndrome. Aquaculture 2017, 473, 430–432. [Google Scholar] [CrossRef]
- Senapin, S.; Shyam, K.; Meemetta, W.; Rattanarojpong, T.; Dong, H.T. Inapparent infection cases of tilapia lake virus (tilv) in farmed tilapia. Aquaculture 2018, 487, 51–55. [Google Scholar] [CrossRef]
- Mugimba, K.; Chengula, A.; Wamala, S.; Mwega, E.; Kasanga, C.; Byarugaba, D.; Mdegela, R.; Tal, S.; Bornstein, B.; Dishon, A. Detection of tilapia lake virus (ti lv) infection by pcr in farmed and wild nile tilapia (Oreochromis niloticus) from lake Victoria. J. Fish Dis. 2018. [Google Scholar] [CrossRef]
- Tattiyapong, P.; Dachavichitlead, W.; Surachetpong, W. Experimental infection of tilapia lake virus (tilv) in nile tilapia (oreochromis niloticus) and red tilapia (Oreochromis spp.). Vet. Microbiol. 2017, 207, 170–177. [Google Scholar] [CrossRef]
- Jaemwimol, P.; Rawiwan, P.; Tattiyapong, P.; Saengnual, P.; Kamlangdee, A.; Surachetpong, W. Susceptibility of important warm water fish species to tilapia lake virus (tilv) infection. Aquaculture 2018, 497, 462–468. [Google Scholar] [CrossRef]
- Abdullah, A.; Ramly, R.; Ridzwan, M.S.M.; Sudirwan, F.; Abas, A.; Ahmad, K.; Murni, M.; Kua, B.C. First detection of tilapia lake virus (tilv) in wild river carp (barbonymus schwanenfeldii) at Timah Tasoh lake, Malaysia. J. Fish Dis. 2018, 41, 1459–1462. [Google Scholar] [CrossRef]
- Munang’andu, H.M.; Fredriksen, B.N.; Mutoloki, S.; Dalmo, R.A.; Evensen, O. The kinetics of cd4+ and cd8+ t-cell gene expression correlate with protection in atlantic salmon (Salmo salar L.) vaccinated against infectious pancreatic necrosis. Vaccine 2013, 31, 1956–1963. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Sandtro, A.; Mutoloki, S.; Brudeseth, B.E.; Santi, N.; Evensen, O. Immunogenicity and cross protective ability of the central vp2 amino acids of infectious pancreatic necrosis virus in atlantic salmon (Salmo salar L.). PLoS ONE 2013, 8, e54263. [Google Scholar] [CrossRef]
- Bustin, S.; Benes, V.; Nolan, T.; Pfaffl, M.W. Quantitative real-time RT-PCR—A perspective. J. Mol. Endocrinol. 2005, 34, 597–601. [Google Scholar] [CrossRef]
- Gadan, K.; Sandtro, A.; Marjara, I.S.; Santi, N.; Munang’andu, H.M.; Evensen, O. Stress-induced reversion to virulence of infectious pancreatic necrosis virus in naive fry of atlantic salmon (Salmo salar L.). PLoS ONE 2013, 8, e54656. [Google Scholar] [CrossRef]
- Chengula, A.A.; Mugimba, K.K.; Shlomit, T.S.; Dubey, D.; Mutoloki, S.; Dishon, A.; Evensen, Ø.; Munang’andu, H.M. Comparison of efficiency and sensitivity for primers targeting all segments of tilapia lake virus (TiLV) using quantitative real-time PCR. J. Fish. Dis. 2019. (Submitted). [Google Scholar]
- CLC Workbench Version 6. Available online: https://www.qiagenbioinformatics.com/products/clc-genomics-workbench/ (accessed on 9 February 2019).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Kembou, J.T.; Zamostiano, R.; Watted, S.; Berkowitz, A.; Rosenbluth, E.; Mishra, N.; Briese, T.; Lipkin, W.; Kabuusu, R.; Ferguson, H. Detection of tilapia lake virus in clinical samples by culturing and nested reverse transcription-pcr. J. Clin. Microbiol. 2017, 55, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Fredriksen, B.N.; Mutoloki, S.; Dalmo, R.A.; Evensen, O. Antigen dose and humoral immune response correspond with protection for inactivated infectious pancreatic necrosis virus vaccines in atlantic salmon (Salmo salar L.). Vet. Res. 2013, 44, 7. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Mutoloki, S.; Evensen, O. Acquired immunity and vaccination against infectious pancreatic necrosis virus of salmon. Dev. Comp. Immunol. 2014, 43, 184–196. [Google Scholar] [CrossRef]
- Zhi, T.; Xu, X.; Chen, J.; Zheng, Y.; Zhang, S.; Peng, J.; Brown, C.L.; Yang, T. Expression of immune-related genes of nile tilapia oreochromis niloticus after gyrodactylus cichlidarum and cichlidogyrus sclerosus infections demonstrating immunosupression in coinfection. Fish Shellfish Immunol. 2018, 80, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Jantrakajorn, S.; Wongtavatchai, J. Francisella infection in cultured tilapia in Thailand and the inflammatory cytokine response. J. Aquat. Anim. Health 2016, 28, 97–106. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | NCBI | Primer Sequence | Bp Length | Tm (°C) | |
|---|---|---|---|---|---|
| IL-1β | KF747686.1 | F | TGGAGGAGGTGACGGATAAA | 86 bp | 62 °C |
| R | GGTGTCGCGTTTGTAGAAGA | ||||
| TNFα | NM_001115056 | F | GGCTAGATTTCCTCTGCTGTATC | 79 bp | 62 °C |
| R | GCTATGACAGCACCTCTGTATC | ||||
| β-actin | KJ126772.1 | F | GTGGGTATGGGTCAGAAAGAC | 111 bp | 62 °C |
| R | GTCATCCCAGTTGGTCACAATA | ||||
| TiLV Seg 3 | KU751816 | F | TCCAGATCACCCTTCCTACTT | 109 bp | 62 °C |
| R | ATCCCAAGCAATCGGCTAAT |
| Parameters | Fish breed/Strain | |
|---|---|---|
| Gray Tilapia | Red Tilapia | |
| Controls | 60 | 60 |
| Number of fish tested | 76 | 73 |
| Number of tanks | 2 | 2 |
| Post challenge Mortality % | 80.4% | 100.0% |
| Hazard risk ratio | 16.98 | 19.51 |
| 95% Conf Interv Hazard risk | 09.40–30.68 | 11.62–32.96 |
| Media survival | 10 | 11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugimba, K.K.; Tal, S.; Dubey, S.; Mutoloki, S.; Dishon, A.; Evensen, Ø.; Munang’andu, H.M. Gray (Oreochromis niloticus x O. aureus) and Red (Oreochromis spp.) Tilapia Show Equal Susceptibility and Proinflammatory Cytokine Responses to Experimental Tilapia Lake Virus Infection. Viruses 2019, 11, 893. https://0-doi-org.brum.beds.ac.uk/10.3390/v11100893
Mugimba KK, Tal S, Dubey S, Mutoloki S, Dishon A, Evensen Ø, Munang’andu HM. Gray (Oreochromis niloticus x O. aureus) and Red (Oreochromis spp.) Tilapia Show Equal Susceptibility and Proinflammatory Cytokine Responses to Experimental Tilapia Lake Virus Infection. Viruses. 2019; 11(10):893. https://0-doi-org.brum.beds.ac.uk/10.3390/v11100893
Chicago/Turabian StyleMugimba, Kizito Kahoza, Shlomit Tal, Saurabh Dubey, Stephen Mutoloki, Arnon Dishon, Øystein Evensen, and Hetron M. Munang’andu. 2019. "Gray (Oreochromis niloticus x O. aureus) and Red (Oreochromis spp.) Tilapia Show Equal Susceptibility and Proinflammatory Cytokine Responses to Experimental Tilapia Lake Virus Infection" Viruses 11, no. 10: 893. https://0-doi-org.brum.beds.ac.uk/10.3390/v11100893