The Susceptibility of Human Melanoma Cells to Infection with the Leningrad-16 Vaccine Strain of Measles Virus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures, Virus and Viral Propagation
2.2. Viral and Total RNA Extraction
2.3. Quantitative Real-Time PCR (qPCR)
2.4. MTT Assay
2.5. xCELLigence Real-Time Cell Proliferation Measurement
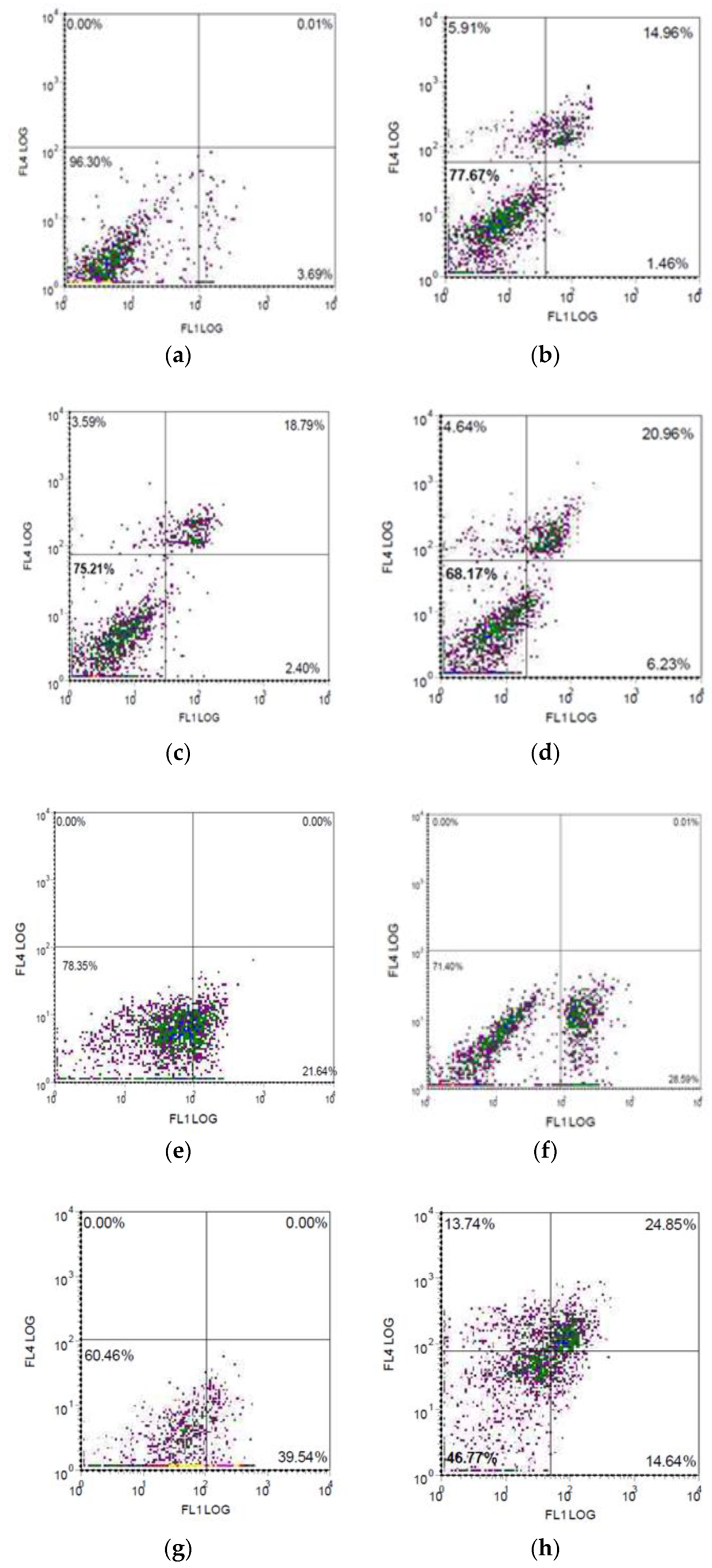
2.6. Flow Cytometry
2.7. ELISA
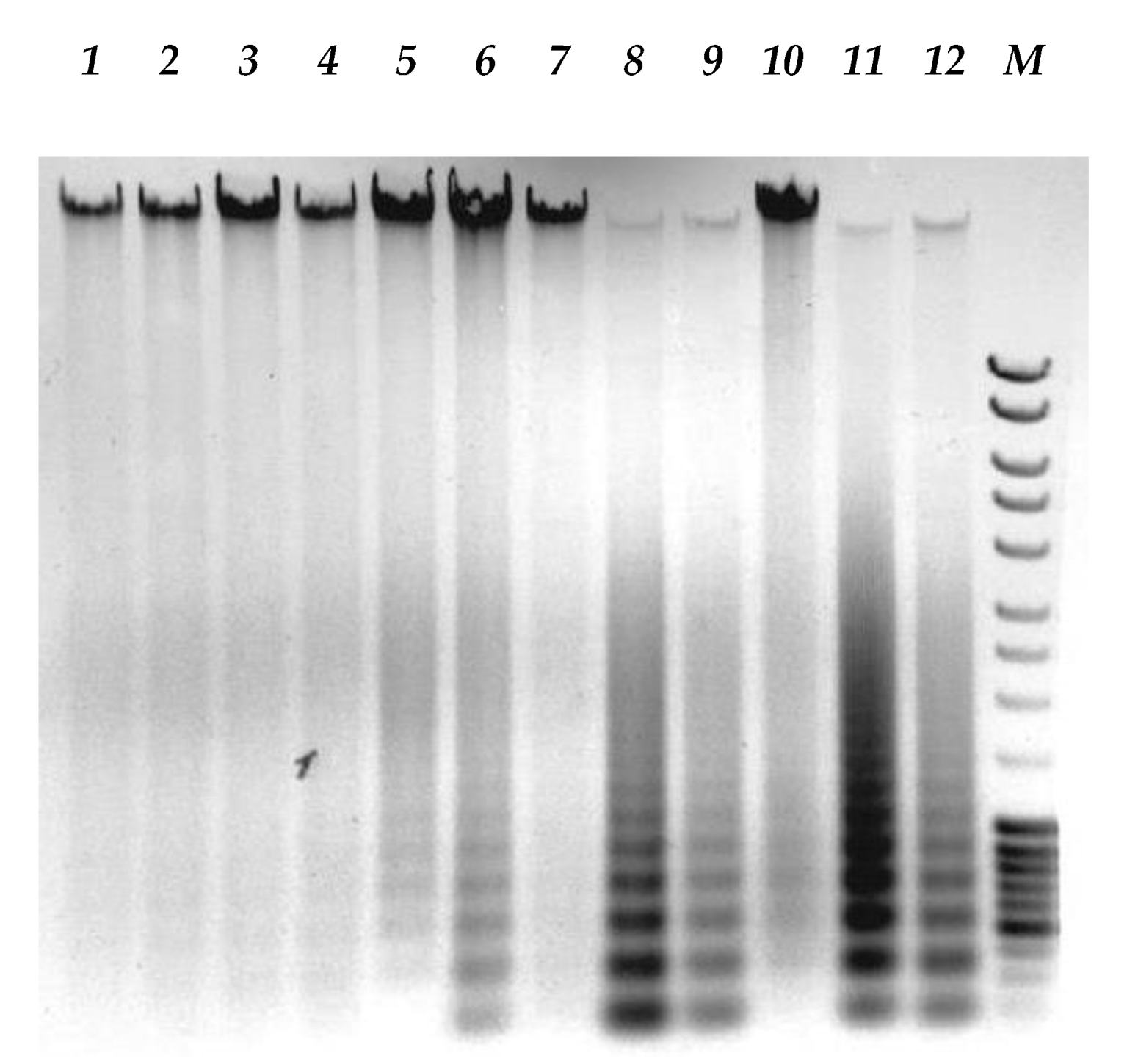
2.8. Agarose Gel Electrophoresis
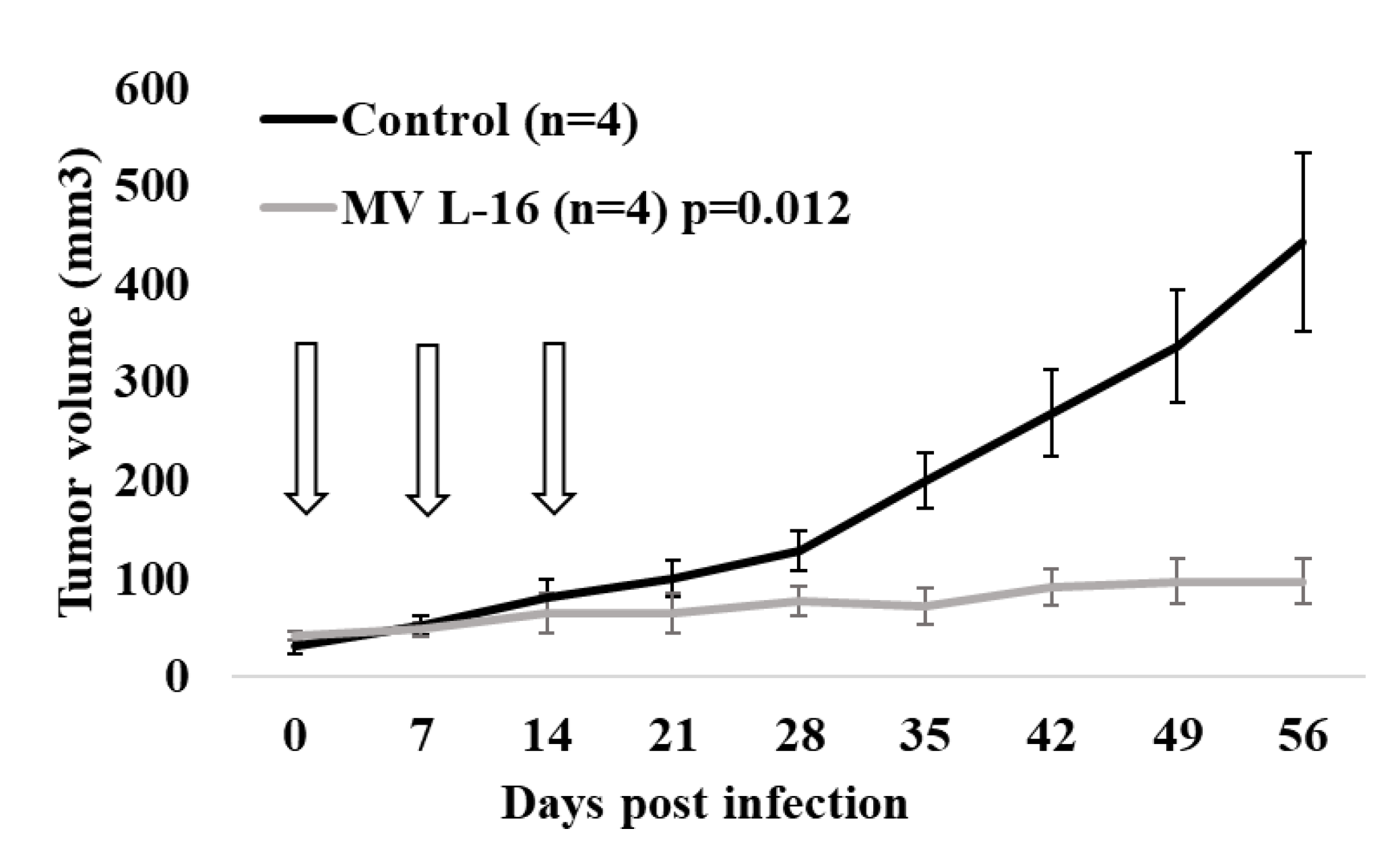
2.9. Animal Experiments
2.10. Statistical Analysis
3. Results
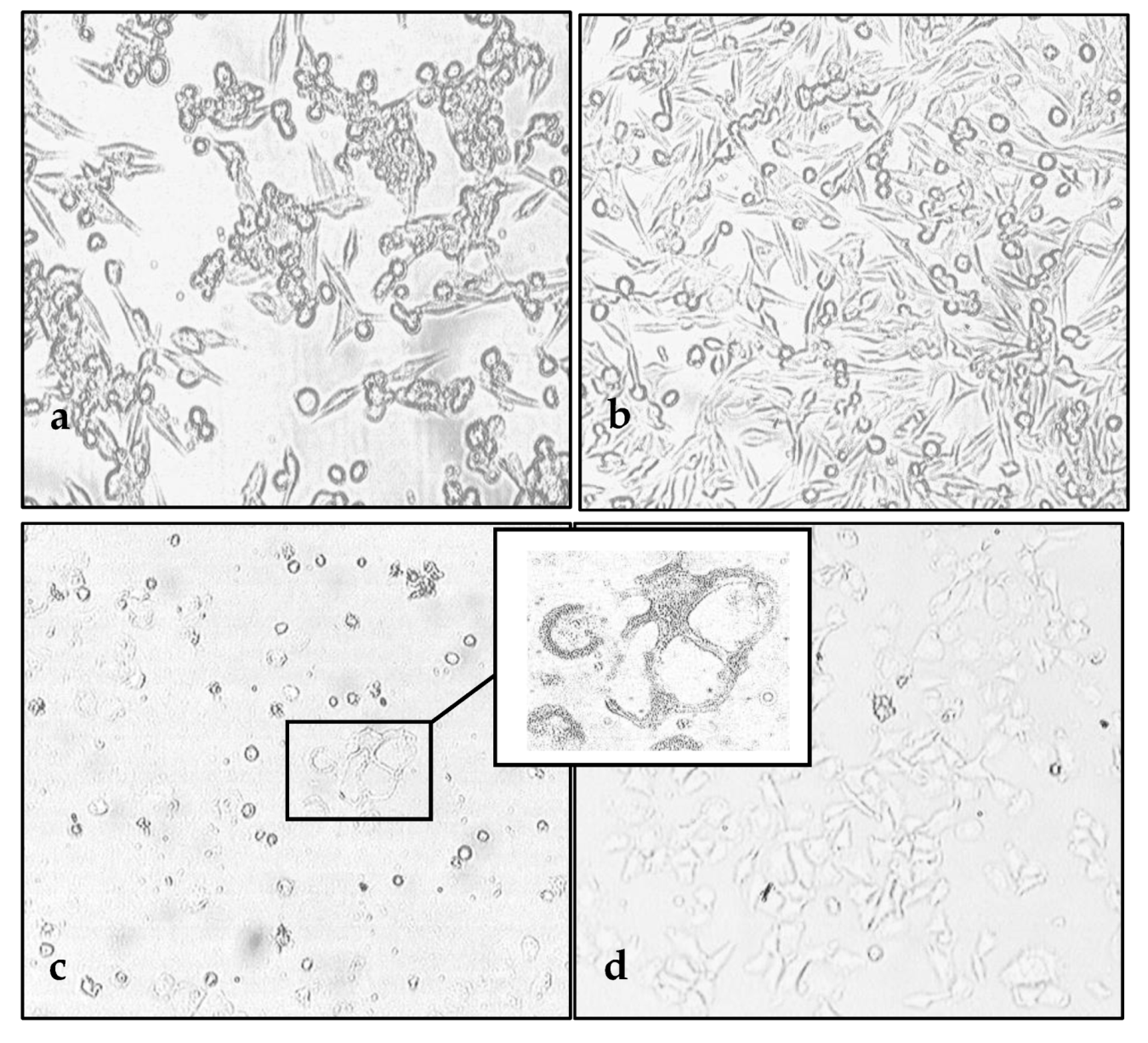
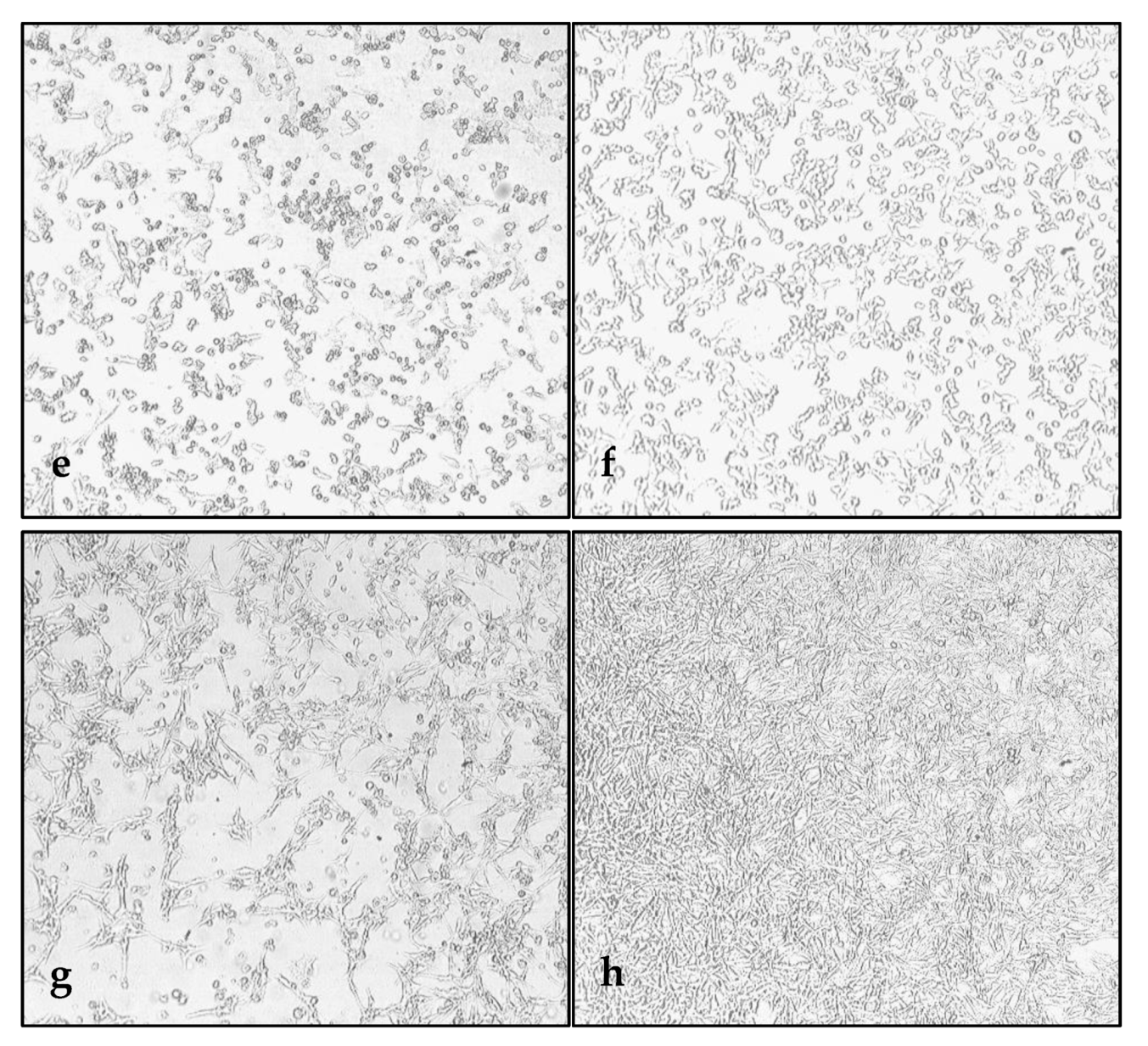
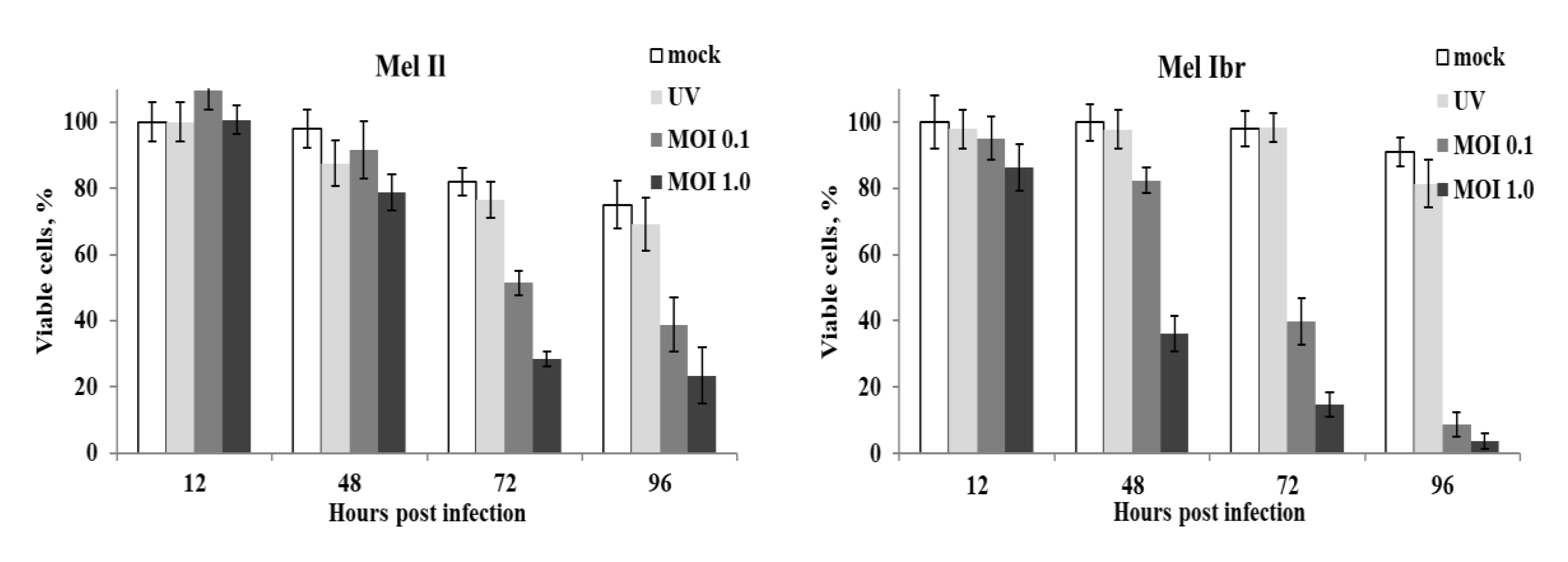
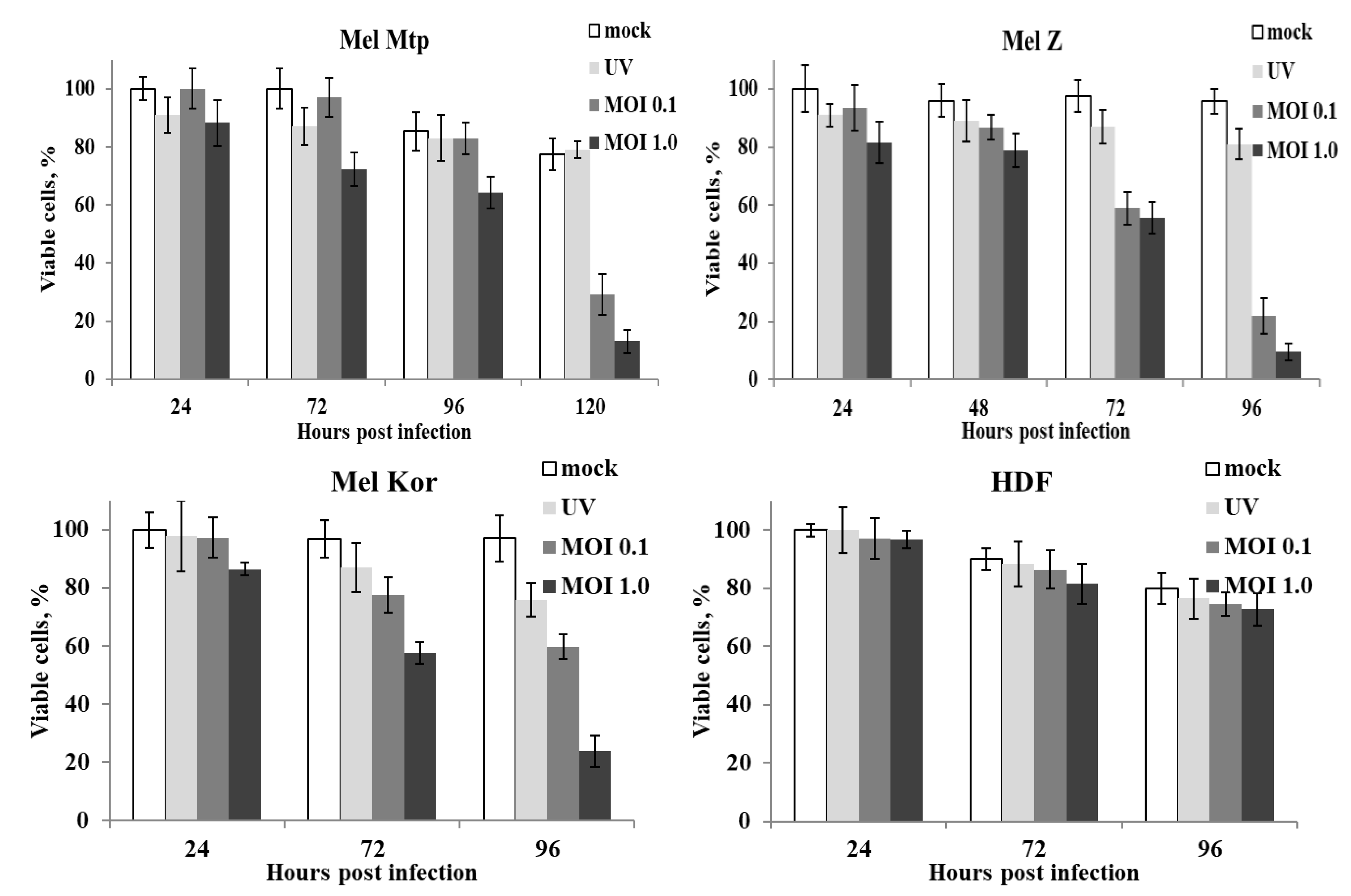
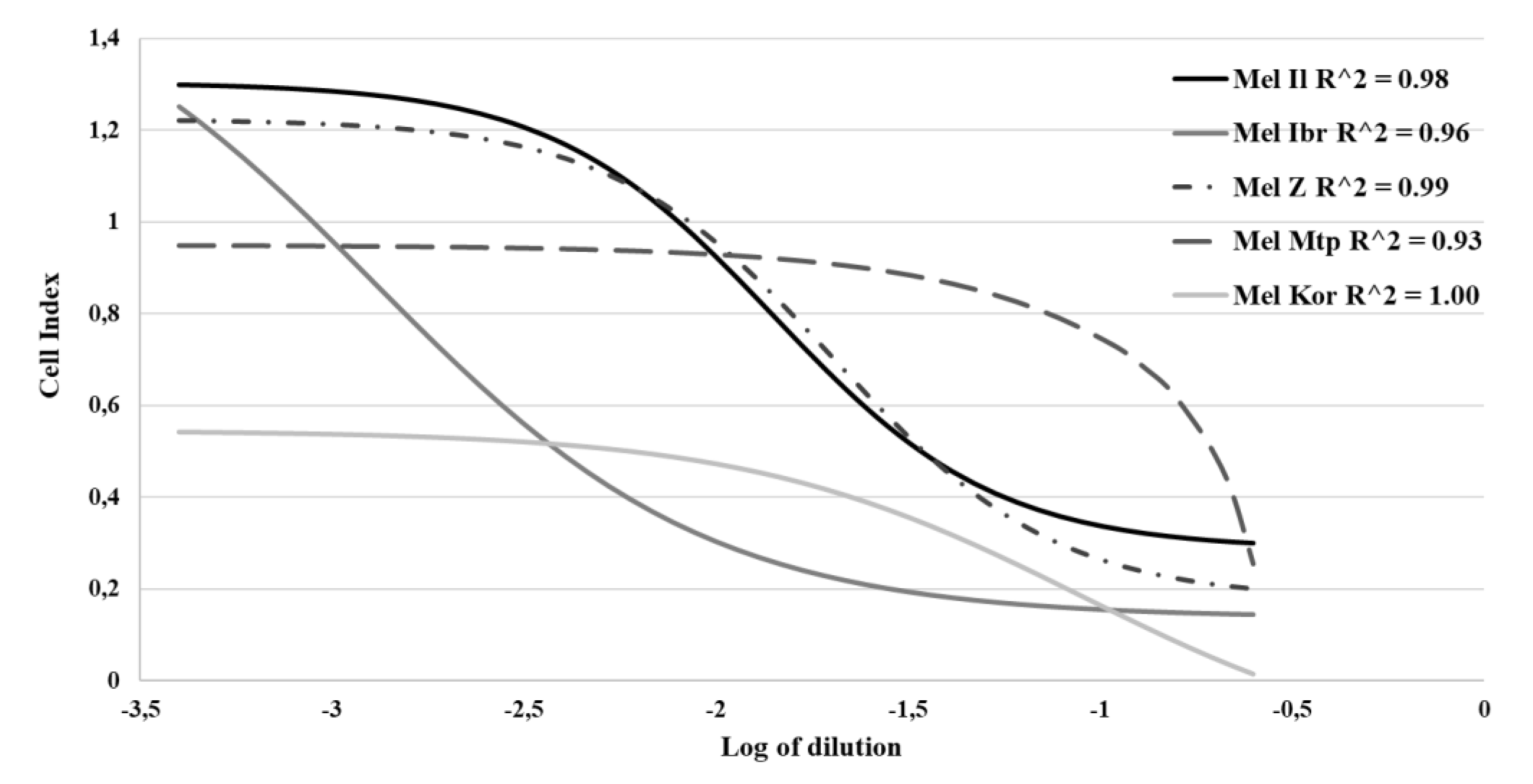
3.1. Susceptibility of Melanoma Cell Lines to the Oncolysis Induced by the Leningrad-16 Vaccine Strain of Measles Virus
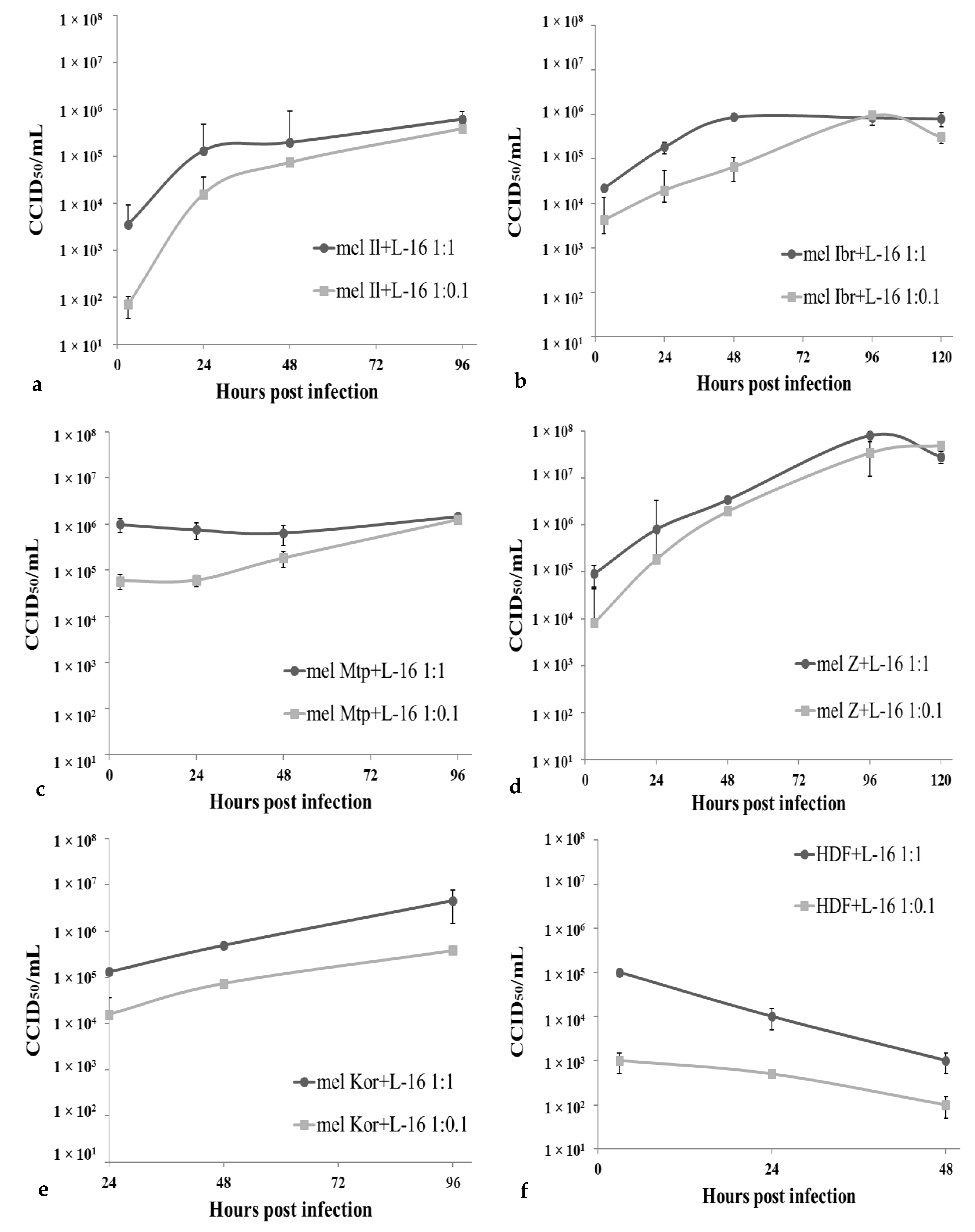
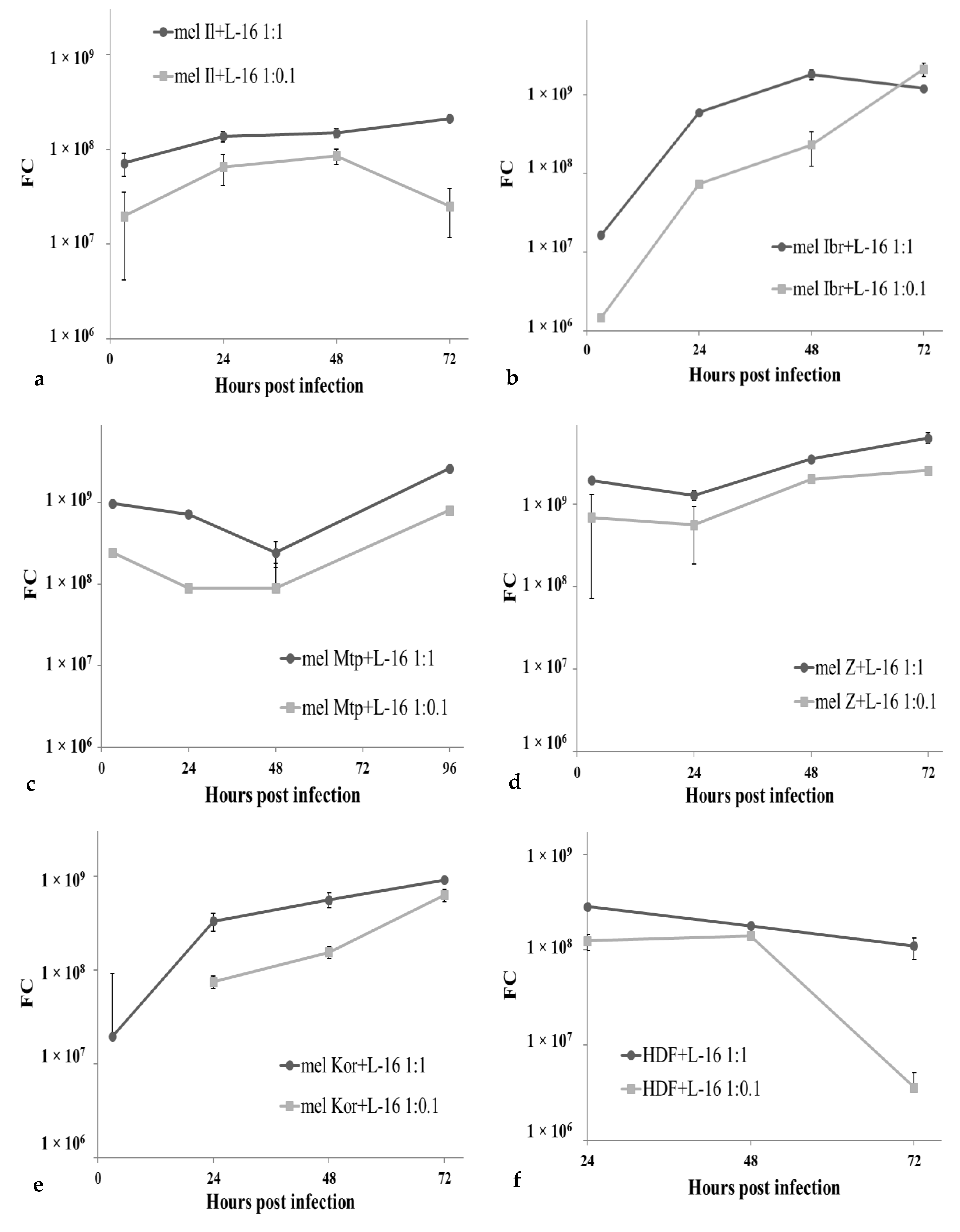
3.2. Permissiveness of Human Melanoma Cells to the Leningrad-16 Vaccine Strain of Measles Virus
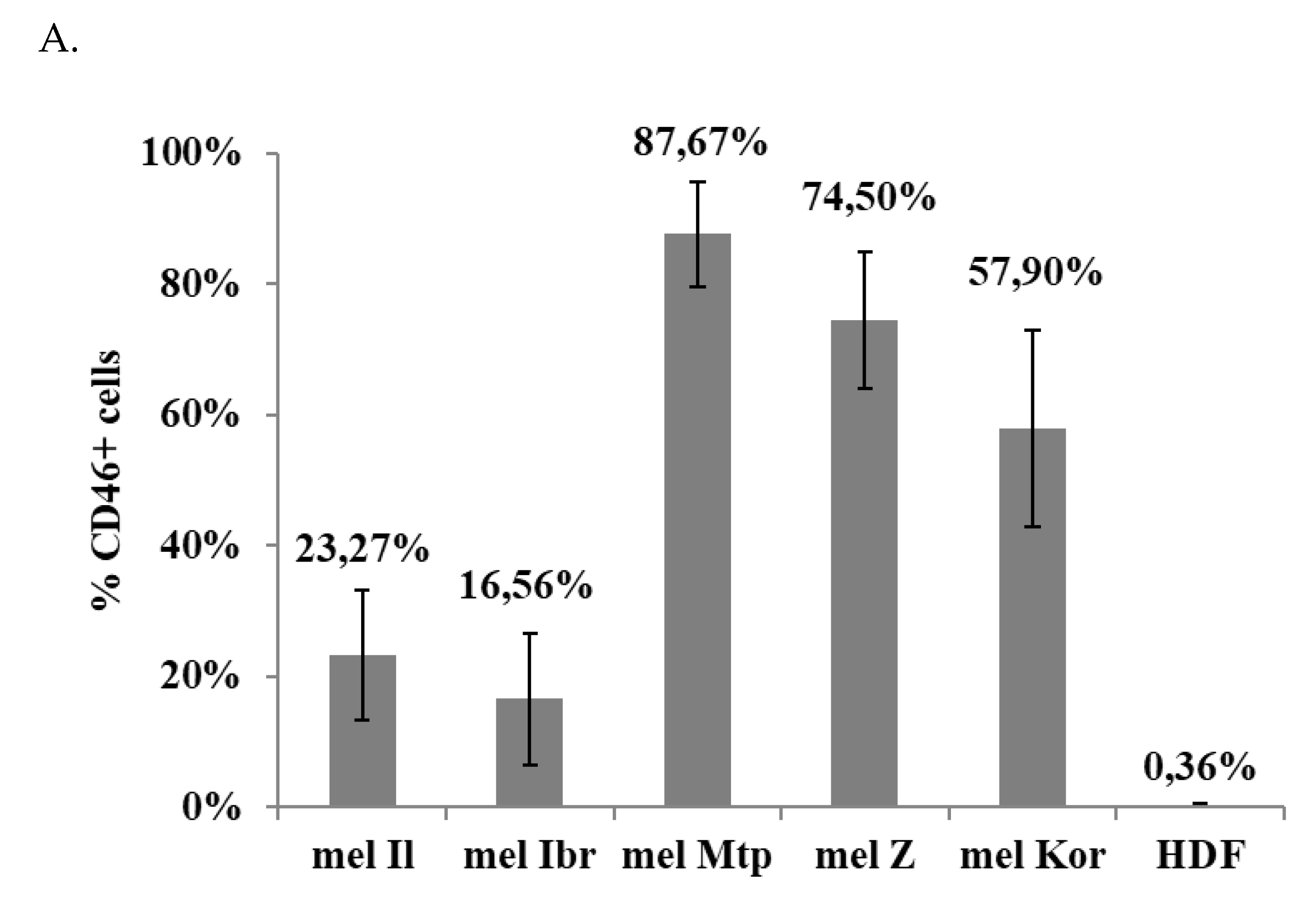
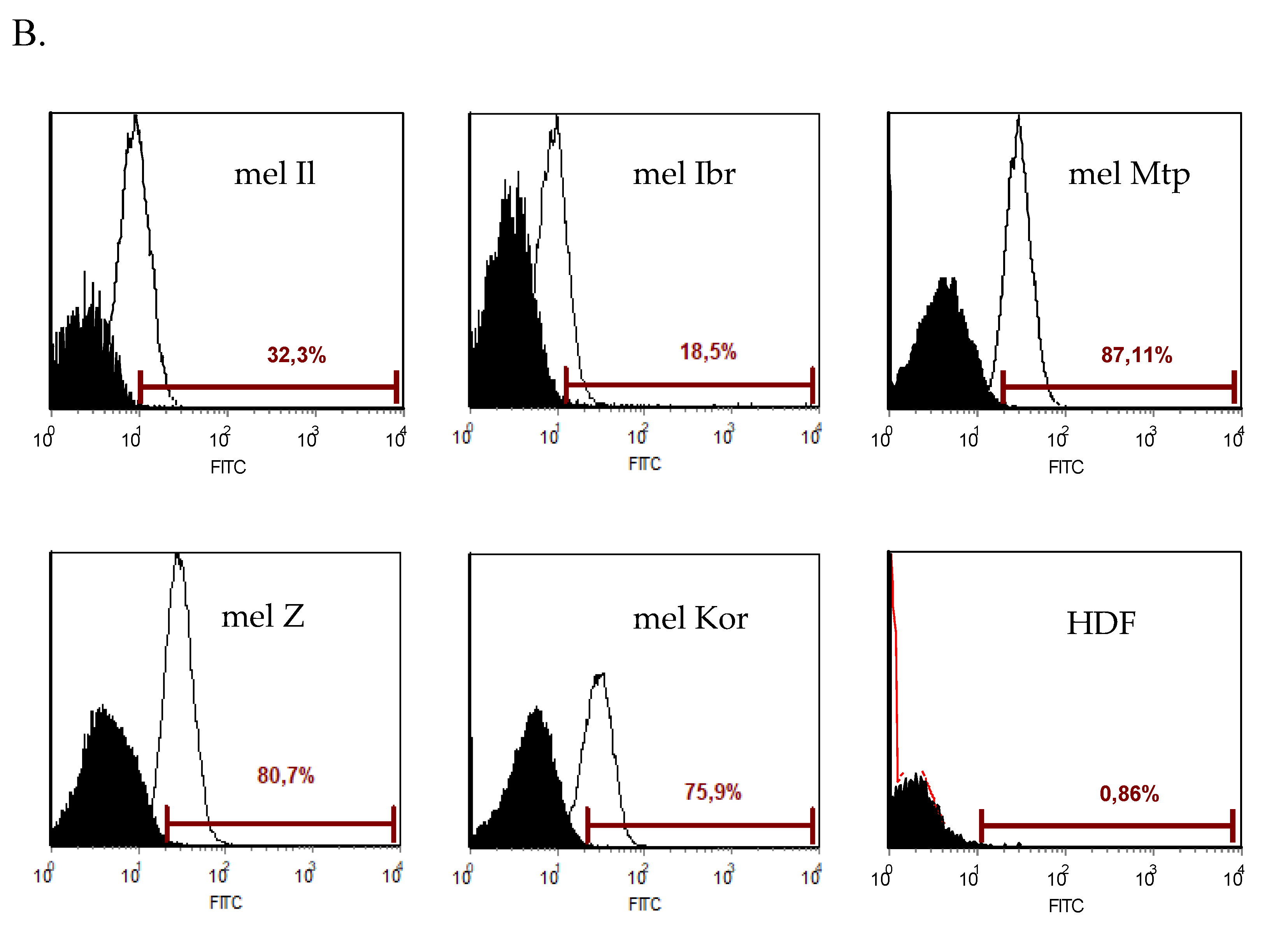
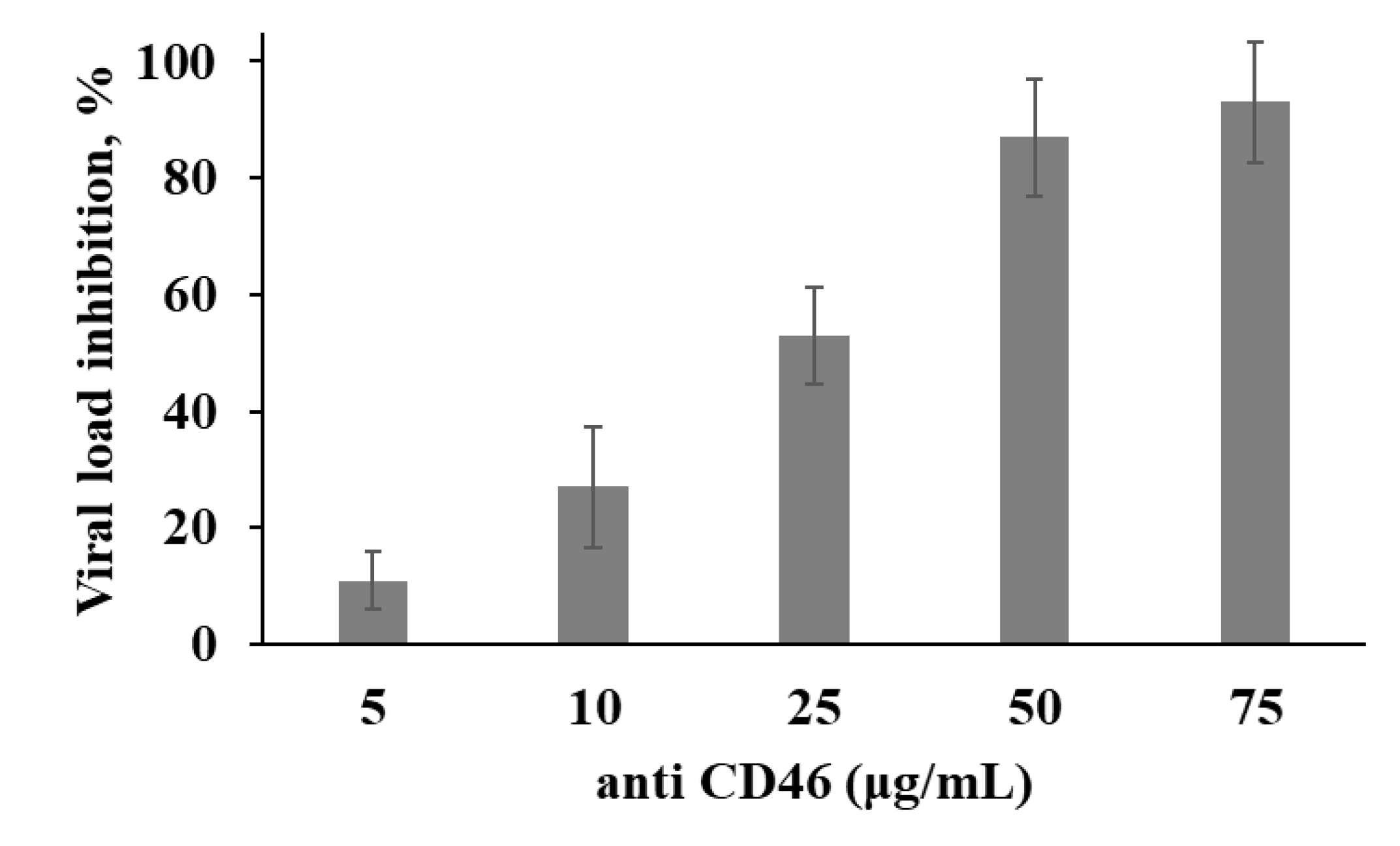
3.3. Expression of the Measles Virus Receptor, CD46, by the Human Melanoma Cell Lines
3.4. Expression of type I IFN Signaling Pathway Genes by Human Melanoma Cells in Response to Measles Virus Infection
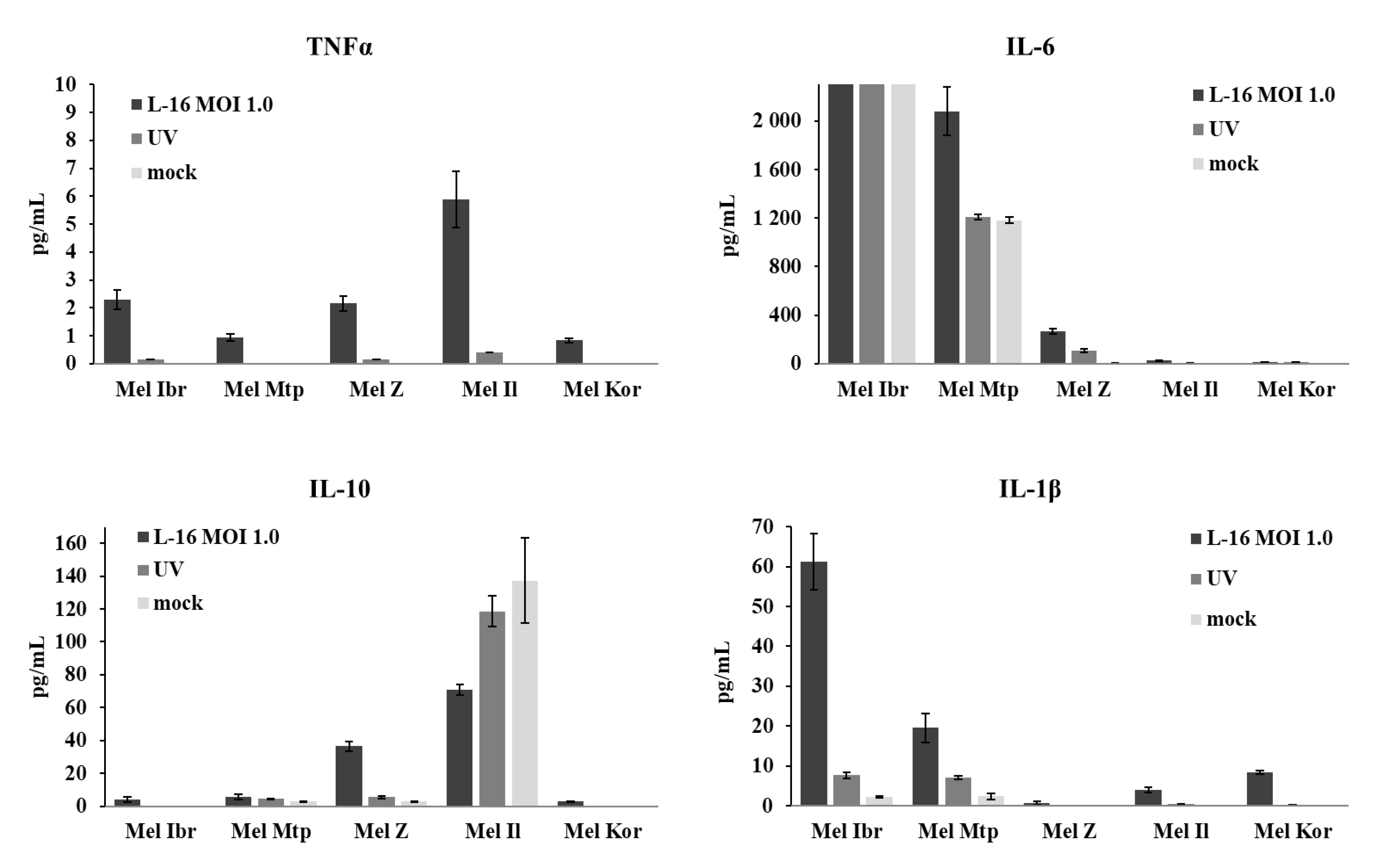
3.5. Release of Inflammatory Cytokines by Human Melanoma Cells After Infection with the Leningrad-16 Vaccine Strain of Measles Virus
3.6. Enhancement of Human Melanoma Cell Death Followed by Measles Virus Infection
3.7. Potent Antitumor Activity Induced by the Leningrad-16 Vaccine Strain of Measles Virus in Mouse Xenografts
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Malishevskaya, N.P.; Sokolova, A.V.; Demidov, L.V. The incidence of skin melanoma in the Russian Federation and federal districts. Med Counc. 2018, 10, 161–165. (In Russian) [Google Scholar] [CrossRef]
- Matthews, N.H.; Li, W.-Q.; Qureshi, A.A.; Weinstock, M.A.; Cho, E. Epidemiology of Melanoma. In Cutaneous Melanoma: Epidemiology and Therapy; Ward, W.H., Farma, J.M., Eds.; Codon Publications: Brisbane, Australia, 2017. [Google Scholar]
- Lacroix, J.; Kis, Z.; Josupeit, R.; Schlund, F.; Stroh-Dege, A.; Frank-Stöhr, M.; Leuchs, B.; Schlehofer, J.R.; Rommelaere, J.; Dinsart, C. Preclinical Testing of an Oncolytic Parvovirus in Ewing Sarcoma: Protoparvovirus H-1 Induces Apoptosis and Lytic Infection In Vitro but Fails to Improve Survival In Vivo. Viruses 2018, 10, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errington, F.; White, C.L.; Twigger, K.R.; Rose, A.; Scott, K.; Steele, L.; Ilett, L.J.; Prestwich, R.; Pandha, H.S.; Coffey, M.; et al. Inflammatory tumor cell killing by oncolytic reovirus for the treatment of melanoma. Gene Ther. 2008, 18, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.A.; Besmer, D.M.; Moerdyk-Schauwecker, M.; Moestl, N.; Ornelles, D.; Mukherjee, P.; Grdzelishvili, V.Z. Vesicular Stomatitis Virus as an Oncolytic Agent against Pancreatic Ductal Adenocarcinoma. J. Virol. 2012, 86, 3073–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Fan, J.; Ding, Y.; Zhang, J.; Zhou, B.; Zhang, Y.; Huang, B.; Hu, Z. Oncolytic cancer therapy with a vaccinia virus strain. Oncol. Rep. 2019, 41, 686–692. [Google Scholar] [CrossRef]
- Msaouel, P.; Opyrchal, M.; Dispenzieri, A.; Peng, K.W.; Federspiel, M.J.; Russell, S.J.; Galanis, E. Clinical Trials with Oncolytic Measles Virus: Current Status and Future Prospects. Curr. Cancer Drug Targets 2017, 18, 177–187. [Google Scholar] [CrossRef]
- Alberts, P.; Tilgase, A.; Rasa, A.; Bandere, K.; Venskus, D. The advent of oncolytic virotherapy in oncology: The Rigvir® story. Eur. J. Pharmacol. 2018, 837, 117–126. [Google Scholar] [CrossRef]
- Bayan, C.Y.; Lopez, A.T.; Gartrell, R.D.; Komatsubara, K.M.; Bogardus, M.; Rao, N.; Chen, C.; Hart, T.D.; Enzler, T.; Rizk, E.M.; et al. The Role of Oncolytic Viruses in the Treatment of Melanoma. Curr. Oncol. Rep. 2018, 20, 80–101. [Google Scholar] [CrossRef]
- Donnelly, O.G.; Errington-Mais, F.; Steele, L.; Hadac, E.; Jennings, V.; Scott, K.; Peach, H.; Phillips, R.M.; Bond, J.; Pandha, H.; et al. Measles virus causes immunogenic cell death in human melanoma. Gene Ther. 2013, 20, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Dharmadhikari, N.; Mehnert, J.M.; Kaufman, H.L. Oncolytic virus immunotherapy for melanoma. Curr. Treat. Options Oncol. 2015, 16, 326. [Google Scholar] [CrossRef]
- Ammour, Y.I.; Ryabaya, O.O.; Milovanova, A.V.; Sidorov, A.V.; Shohin, I.E.; Zverev, V.V.; Nasedkina, T.V. Oncolytic Properties of a Mumps Virus Vaccine Strain in Human Melanoma Cell Lines. Mol. Biol. 2018, 52, 570–576. [Google Scholar] [CrossRef]
- Anderson, B.D.; Nakamura, T.; Russell, S.J.; Peng, K.W. High CD46 Receptor Density Determines Preferential Killing of Tumor Cells by Oncolytic Measles Virus. Cancer Res. 2004, 64, 4919–4926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nechaeva, E.A.; Getmanova, T.N.; Senkina, T.Y.; Yurchenko, N.D. Study of Leningrad-16 Vaccine Strain of Measles Virus Reproduction in Cell Cultures Perspective for Biotechnology. New Dev. New Appl. Animal Cell Tech. 1998, 577–579. [Google Scholar]
- Mikhaylova, I.N.; Kovalevsky, D.A.; Morozova, L.F.; Golubeva, V.A.; Cheremushkin, E.A.; Lukashina, M.I.; Voronina, E.S.; Burova, O.S.; Utyashev, I.A.; Kiselev, S.L.; et al. Cancer/testis genes expression in human melanoma cell lines. Melanoma Res. 2008, 18, 303–313. [Google Scholar] [CrossRef]
- Mikhaĭlova, I.N.; Lukashina, M.I.; Baryshnikov, A.I.; Morozova, L.F.; Burova, O.S.; Palkina, T.N.; Kozlov, A.M.; Golubeva, V.A.; Cheremushkin, E.A.; Doroshenko, M.B.; et al. Melanoma cell lines as the basis for antitumor vaccine preparation. Vestn. Ross. Akad. Med. Nauk. 2005, 7, 37–40. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Ammour, Y.; Faizuloev, E.; Borisova, T.; Nikonova, A.; Dmitriev, G.; Lobodanov, S.; Zverev, V. Quantification of measles, mumps and rubella viruses using real-time quantitative TaqMan-based RT-PCR assay. J. Virol. Methods 2013, 187, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Emelyanova, M.; Ghukasyan, L.; Abramov, I.; Ryabaya, O.; Stepanova, E.; Kudryavtseva, A.; Sadritdinova, A.; Dzhumakova, C.; Belysheva, T.; Surzhikov, S.; et al. Detection of BRAF, NRAS, KIT, GNAQ, GNA11 and MAP2K1/2 mutations in Russian melanoma patients using LNA PCR clamp and biochip analysis. Oncotarget 2017, 8, 52304–52320. [Google Scholar] [CrossRef] [Green Version]
- Lichty, B.D.; Breitbach, C.J.; Stojdl, D.F.; Bell, J.C. Going viral with cancer immunotherapy. Nat. Rev. Cancer. 2014, 14, 559–567. [Google Scholar] [CrossRef]
- Calain, P.; Roux, L. Generation of Measles Virus Defective Interfering Particles and Their Presence in a Preparation of Attenuated Live-Virus Vaccine. J. Virol. 1988, 62, 2859–2866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Xu, L.; Junhong, S.; Peppelenbosch, M.P.; Pan, Q. Transcriptional Regulation of Antiviral Interferon-Stimulated Genes. Trends Microbiol. 2017, 25, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.F.; Tan, D.Q.; Jeyasekharan, A.D.; Hsieh, W.S.; Ho, A.S.; Ichiyama, K.; Ye, M.; Pang, B.; Ohba, K.; Liu, X.; et al. Combination of vaccine-strain measles and mumps virus synergistically kills a wide range of human hematological cancer cells: Special focus on acute myeloid leukemia. Cancer Lett. 2014, 354, 272–280. [Google Scholar] [CrossRef]
- Galanis, E.; Bateman, A.; Johnson, K.; Diaz, R.M.; James, C.D.; Vile, R.; Russell, S.J. Use of viral fusogenic membrane glycoproteins as novel therapeutic transgenes in gliomas. Hum. Gene Ther. 2001, 12, 811–821. [Google Scholar] [CrossRef]
- Wild, T.F.; Malvoisin, E.; Buckland, R. Measles virus: Both the haemagglutinin and fusion glycoproteins are required for fusion. J. Gen. Virol. 1991, 72, 439–442. [Google Scholar] [CrossRef]
- Ndoye, A.; Weeraratna, A.T. Autophagy—An emerging target for melanoma therapy. F1000 Res. 2016, 5, F1000 Faculty Rev-1888. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.R.; Jacobson, B.A.; Belgum, H.; Raza, A.; Sadiq, A.; Drees, J.; Wang, H.; Jay-Dixon, J.; Etchison, R.; Federspiel, M.J.; et al. Measles vaccine strains for virotherapy of non-small-cell lung carcinoma. J. Thorac. Oncol. 2014, 9, 1101–1110. [Google Scholar] [CrossRef] [Green Version]
- Achard, C.; Boisgerault, N.; Delaunay, T.; Roulois, D.; Nedellec, S.; Royer, P.; Pain, M.; Combredet, C.; Mesel-Lemoine, M.; Cellerin, L.; et al. Sensitivity of human pleural mesothelioma to oncolytic measles virus depends on defects of the type I interferon response. Oncotarget 2015, 6, 44892–44904. [Google Scholar] [CrossRef]
- Allagui, F.; Achard, C.; Panterne, C.; Combredet, C.; Labarrière, N.; Dréno, B.; Elgaaied, A.B.; Pouliquen, D.; Tangy, F.; Fonteneau, J.F.; et al. Modulation of the Type I Interferon Response Defines the Sensitivity of Human Melanoma Cells to Oncolytic Measles Virus. Curr. Gene Ther. 2016, 16, 419–428. [Google Scholar] [CrossRef]
- Kurokawa, C.; Iankov, I.D.; Anderson, S.K.; Aderca, I.; Leontovich, A.A.; Maurer, M.J.; Oberg, A.L.; Schroeder, M.A.; Giannini, C.; Greiner, S.M.; et al. Galanis, E. Constitutive Interferon Pathway Activation in Tumors as an Efficacy Determinant Following Oncolytic Virotherapy. J. Natl. Cancer Inst. 2018, 110, 1123–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajaraman, S.; Canjuga, D.; Ghosh, M.; Cosmin Codrea, M.; Sieger, R.; Wedekink, F.; Tatagiba, M.; Koch, M.; Lauer, U.M.; Nahnsen, S.; et al. Measles Virus-Based Treatments Trigger a Pro-inflammatory Cascade and a Distinctive Immunopeptidome in Glioblastoma. Mol. Ther. Oncolytics. 2019, 12, 147–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodbourn, S.; Didcock, L.; Randall, R.E. Interferons: Cell signaling, immune modulation, antiviral response and virus countermeasures. J. Gen. Virol. 2000, 81, 2341–2364. [Google Scholar] [CrossRef] [PubMed]
- Hiscott, J. Triggering the innate antiviral response through IRF-3 activation. J. Biol. Chem. 2007, 282, 15325–15329. [Google Scholar] [CrossRef] [Green Version]
- Nourbakhsh, M.; Hauser, H. Constitutive silencing of IFN-beta promoter is mediated by NRF (NF-kappaB-repressing factor), a nuclear inhibitor of NF-kappa B. EMBO J. 1999, 18, 6415–6425. [Google Scholar] [CrossRef] [Green Version]
- Le Tortorec, A.; Denis, H.; Satie, A.-P.; Patard, J.-J.; Ruffault, A.; Jegou, B.; Dejucq-Rainsford, N. Antiviral responses of human Leydig cells to mumps virus infection or poly I:C stimulation. Hum. Reprod. 2008, 23, 2095–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, C.K.; Radeke, M.J.; Cattaneo, R.; Samuel, C.E. Measles virus C protein impairs production of defective copyback double-stranded viral RNA and activation of protein kinase R. J. Virol. 2014, 88, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J. Oncogenes: The Passport for Viral Oncolysis Through PKR Inhibition. Biomark. Cancer 2016, 8, 101–110. [Google Scholar] [CrossRef]
- Rozières, A.; Viret, C.; Faure, M. Autophagy in Measles Virus Infection. Viruses 2017, 9, 359. [Google Scholar] [CrossRef] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Genome Mutations Determined by NGS [20] | Corresponding Protein Mutations |
|---|---|---|
| mel Il | BRAF c.1798_1799delGTinsAA | p.V600K |
| BRAF c.1165C > T | p.R389C | |
| TP53 c.326T > C | p.F109S | |
| mel Ibr | BRAF c.1799T > A | p.V600E |
| mel Mtp | NRAS c.182A > G | p.Q61R |
| mel Z | BRAF c.1799T > A | p.V600E |
| PDGFRA c.1432T > C | p.S478P | |
| mel Kor | NRAS c.182A > G | p.Q61R |
| Gene | Fold change | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mel Il+L-16 vs mel Il | mel Ibr+L-16 vs mel Ibr | mel Mtp+L-16 vs mel Mtp | mel Z+L-16 vs mel Z | HDF+L-16 vs HDF | |||||||||||
| 24 | 48 | 72 | 24 | 48 | 72 | 24 | 48 | 96 | 24 | 48 | 72 | 24 | 48 | 72 | |
| Intracytoplasmic sensors of viral dsRNA | |||||||||||||||
| DDX58 | 78.84 | 73.26 | 42.71 | 21.74 | 127.59 | 3.45 | 922.92 | 406.33 | 116.29 | 30.92 | 63.05 | 184.87 | 41.22 | 2.55 | −0.50 |
| IFIH1 | 68.51 | 25.57 | 13.98 | 24.49 | 80.11 | 3.71 | 477.59 | 519.54 | 548.76 | 14.25 | 110.16 | 270.05 | 24.99 | 2.10 | 0.27 |
| Cell signaling/ Regulation of transcription | |||||||||||||||
| MAVS | 1.23 | 0.05 | 0.02 | 2.86 | 12.13 | 1.71 | 0.46 | 1.06 | 0.81 | -0.05 | 0.38 | 0.51 | 1.46 | 0.00 | −0.40 |
| IRF3 | 1.14 | 0.08 | 0.01 | 0.68 | 5.90 | −0.16 | 5.73 | 7.48 | 10.22 | 1.25 | 2.63 | 6.01 | 3.29 | −0.11 | −0.78 |
| Interferon β and its receptors | |||||||||||||||
| IFN-β | 20.39 | 4.27 | 1.47 | −0.88 | 1.75 | 1.05 | 5792.62 | 2521.38 | 1305.15 | −0.33 | 0.27 | 1.54 | 17.38 | 0.14 | −0.35 |
| IFNAR1 | 2.47 | 0.11 | 0.02 | 0.24 | 1.91 | −1.06 | 0.37 | 0.72 | 1.06 | 0.78 | 0.45 | 1.94 | 9.56 | −0.39 | −0.38 |
| IFNAR2 | 3.88 | 1.14 | 0.05 | 0.15 | 7.02 | −0.56 | 4.86 | 4.24 | 3.36 | 1.35 | 0.86 | 1.32 | 8.85 | −0.79 | −1.22 |
| Interferon Stimulated Genes Cell defense/immune response/Apoptosis | |||||||||||||||
| OAS1 | 41.79 | 17.73 | 11.32 | 3.76 | 6.37 | 8.68 | 5802.42 | 2918.37 | 1402.09 | 299 | 11.96 | 7.97 | 1.47 | −0.43 | −0.13 |
| MxA | 36.25 | 23.65 | 16.76 | 9.20 | 106.77 | 95.82 | 1.64 | 5.63 | 5.60 | 2.24 | 6.80 | 18.08 | 1.74 | −0.11 | −0.48 |
| EIF2AK2 | 10.46 | 2.31 | 1.02 | 0.42 | 0.59 | 1.92 | −0.88 | 0.28 | 0.61 | 0.15 | 0.29 | 3.13 | 761.56 | 547.91 | −0.15 |
| TNFSF10 | 111.91 | 39.11 | 7.28 | 83.86 | 659.24 | 280.84 | 13.50 | 43.98 | 62.37 | 2.14 | 18.05 | 51.31 | −0.75 | 0.19 | −0.32 |
| XAF1 | 44.86 | 12.38 | 13.19 | 5.53 | 12.44 | 12.37 | 2.38 | 3147.85 | 7813.10 | −0.66 | 2.44 | 3.45 | 2.77 | 0.12 | 0.09 |
| IFIT2/ISG54 | 4.66 | 4.86 | 0.01 | 653.00 | 653.68 | 212.44 | 1.10 | 1.70 | 1.08 | 0.07 | 2.98 | 7.89 | n/a | n/a | n/a |
| ISG12/IFI27 | 2.78 | 10.88 | 17.97 | n/a | n/a | n/a | 24.85 | 186.39 | −1.89 | n/a | n/a | n/a | n/a | n/a | n/a |
| ISG15 | 5.14 | 4.60 | 0.22 | n/a | n/a | n/a | 6985.52 | 3144.69 | 255.73 | 1.04 | 12.79 | 26.90 | n/a | n/a | n/a |
| IFI6/G1P3 | 2.78 | 5.81 | 4.33 | 0.23 | 1.55 | 0.70 | 129.33 | 5.31 | −1.44 | 1.89 | 1.82 | 2.06 | n/a | n/a | n/a |
| Signal transducers and activators of transcription | |||||||||||||||
| STAT1 | 36.25 | 21.63 | 11.89 | 1.30 | 0.96 | −0.08 | 75.42 | 96.80 | −0.05 | 0.91 | −1.42 | 8.97 | n/a | n/a | n/a |
| STAT2 | 3.53 | 1.24 | 0.38 | 0.30 | −0.75 | 0.00 | 34.65 | 3.19 | −56.21 | 0.17 | 1.46 | 8.40 | n/a | n/a | n/a |
| STAT3 | n/a | n/a | n/a | n/a | n/a | n/a | 0.87 | −0.42 | −0.01 | 1.24 | −1.46 | −0.08 | n/a | n/a | n/a |
| Suppressors of cytokine signaling | |||||||||||||||
| SOCS3 | n/a | n/a | n/a | n/a | n/a | n/a | 65.53 | −39.54 | −1000.40 | −0.98 | 0.49 | 7.92 | n/a | n/a | n/a |
| FC < −1000, p value < 0.001 | FC < −10, p value < 0.005 | −1 > FC > −2 | 0 > FC > −1, p value > 0.05 | 0 < FC < 2.2, p value > 0.05 | |||||
| 2.2 < FC < 10, p value < 0.05 | 10 < FC < 30, p value <0.01 | 30 < FC < 60 | 60 < FC < 100, p value < 0.005 | FC > 100, p value < 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ammour, Y.; Ryabaya, O.; Shchetinina, Y.; Prokofeva, E.; Gavrilova, M.; Khochenkov, D.; Vorobyev, D.; Faizuloev, E.; Shohin, I.; Zverev, V.V.; et al. The Susceptibility of Human Melanoma Cells to Infection with the Leningrad-16 Vaccine Strain of Measles Virus. Viruses 2020, 12, 173. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020173
Ammour Y, Ryabaya O, Shchetinina Y, Prokofeva E, Gavrilova M, Khochenkov D, Vorobyev D, Faizuloev E, Shohin I, Zverev VV, et al. The Susceptibility of Human Melanoma Cells to Infection with the Leningrad-16 Vaccine Strain of Measles Virus. Viruses. 2020; 12(2):173. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020173
Chicago/Turabian StyleAmmour, Yulia, Oxana Ryabaya, Yulia Shchetinina, Elena Prokofeva, Marina Gavrilova, Dmitry Khochenkov, Denis Vorobyev, Evgeny Faizuloev, Igor Shohin, Vitaly V. Zverev, and et al. 2020. "The Susceptibility of Human Melanoma Cells to Infection with the Leningrad-16 Vaccine Strain of Measles Virus" Viruses 12, no. 2: 173. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020173