Suppression of nbe-miR1919c-5p Expression in Nicotiana benthamiana Enhances Tobacco Curly Shoot Virus and Its Betasatellite Co-Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Cloning and Vector Construction
2.2. Plant Growth, Virus Inoculation and Agro-infiltration
2.3. DNA Extraction and Virus DNA Accumulation Analysis
2.4. RNA Extraction and Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.5. Prediction and Quantification of Target Genes
3. Results
3.1. Leaf Curling Symptoms Induced by TbCSV/TbCSB Co-Infection in N. benthamiana
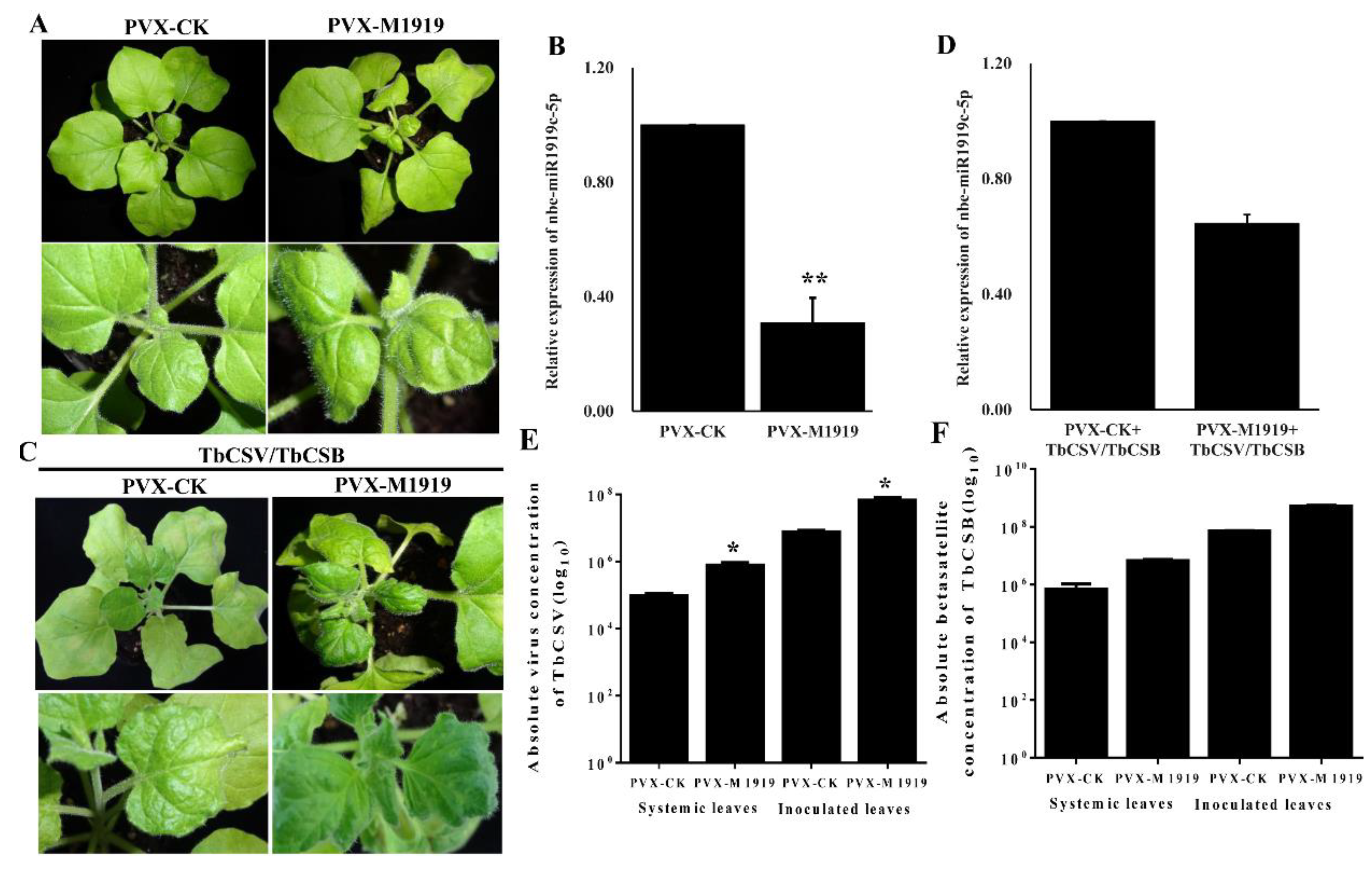
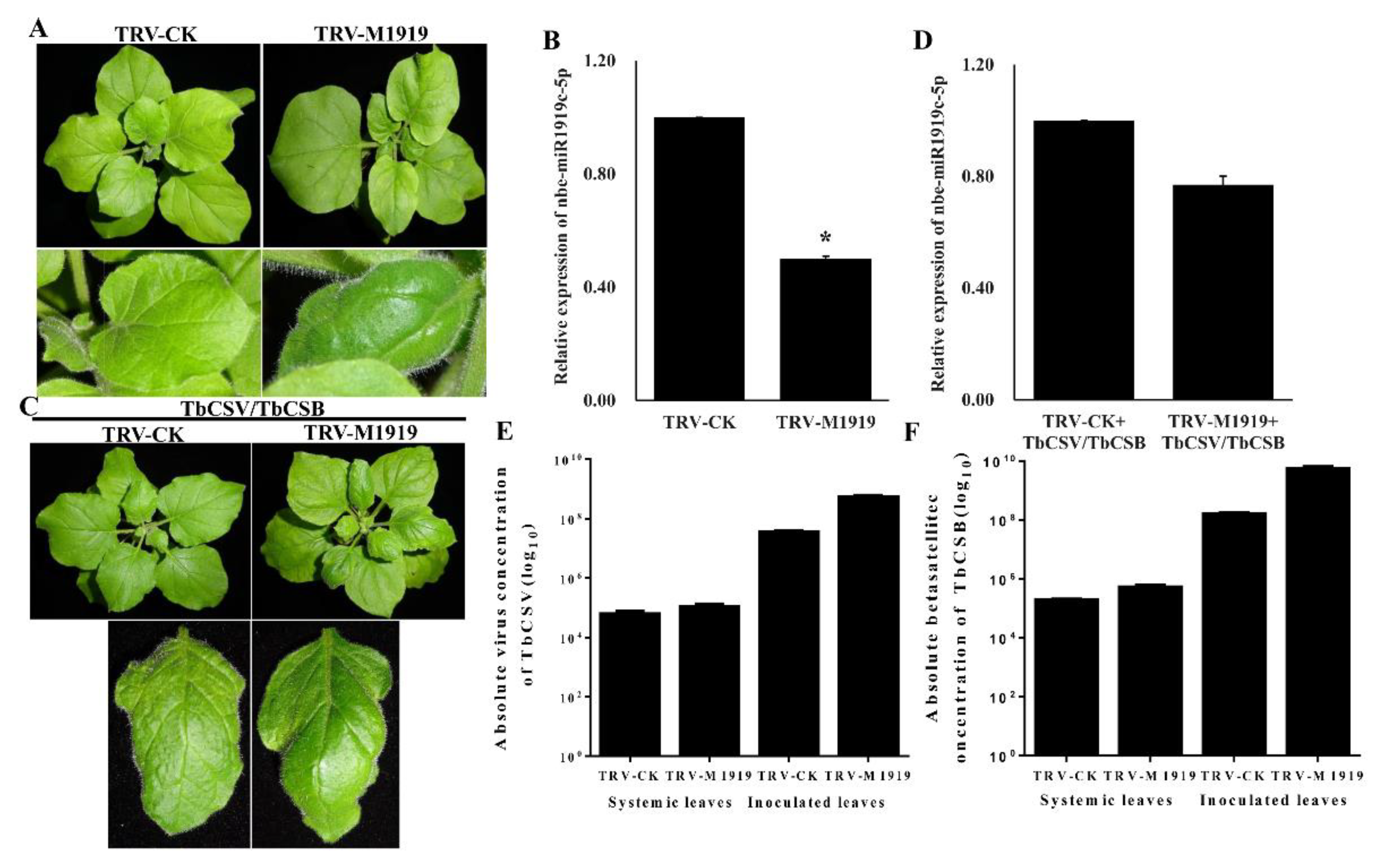
3.2. Suppression of Nbe-miR1919c-5p Expression Enhances Leaf Curling Symptoms Caused by TbCSV/TbCSB Co-Infection
3.3. Suppression of Nbe-miR1919c-5p Expression Enhances TbCSV and TbCSB DNA Accumulation in N. benthamiana Plants
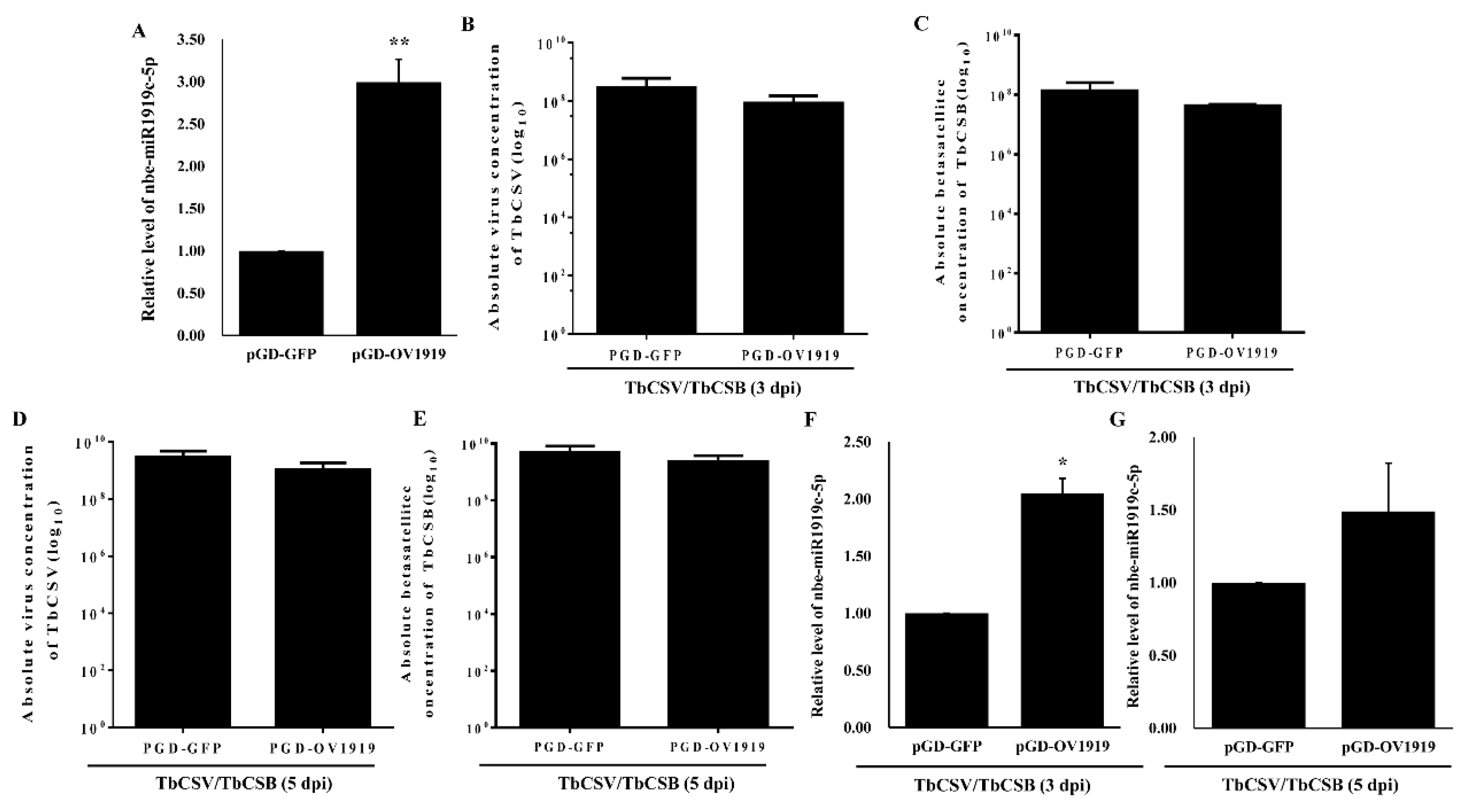
3.4. Over-Expression of Nbe-miR1919c-5p Reduced TbCSV and TbCSB DNA Accumulation in N. benthamiana Plants
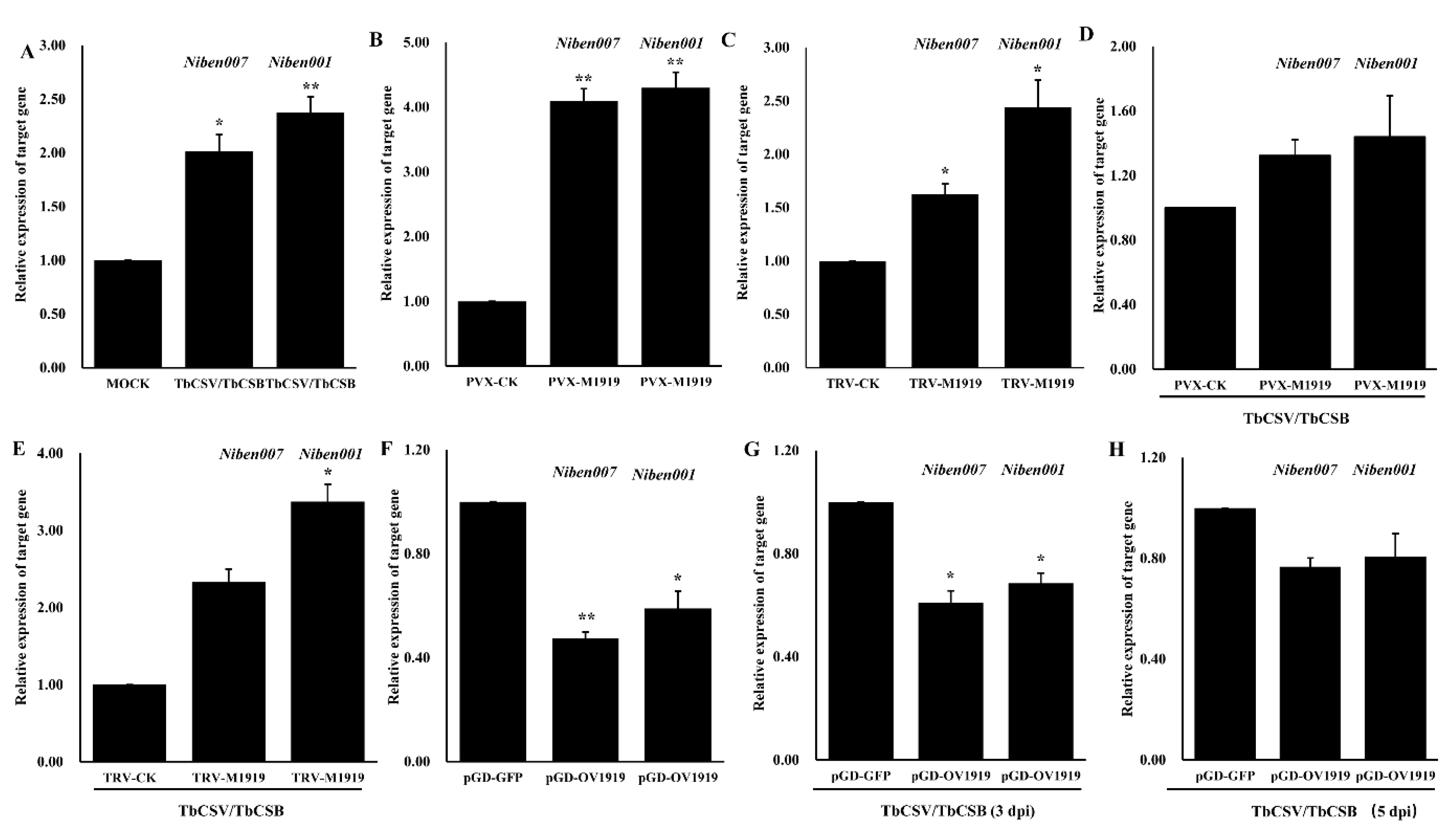
3.5. Quantification of Nbe-miR1919c-5p Target Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.W.; Wang, L.J.; Mao, Y.B.; Cai, W.J.; Xue, H.W.; Chen, X.Y. Control of root cap formation by MicroRNA-targeted auxin response factors in Arabidopsis. Plant. Cell 2005, 17, 2204–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Liu, S.; Wang, M.; Lang, Q.; Jin, C. Identification of microRNAs and their targets in tomato infected with Cucumber mosaic virus based on deep sequencing. Planta 2014, 240, 1335–1352. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ren, Y.; Zhang, Y.; Xu, J.; Zhang, Z.; Wang, Y. Genome-wide profiling of novel and conserved Populus microRNAs involved in pathogen stress response by deep sequencing. Planta 2012, 235, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Dong, Y.Y.; Yin, H.L.; Wang, N.; Yang, J.; Liu, X.M.; Wang, Y.F.; Wu, J.Y.; Li, X.K. Characterization of the stress associated microRNAs in Glycine max by deep sequencing. BMC Plant Biol. 2011, 11, 170. [Google Scholar] [CrossRef] [Green Version]
- Bazzini, A.A.; Hopp, H.E.; Beachy, R.N.; Asurmendi, S. Infection and coaccumulation of tobacco mosaic virus proteins alter microRNA levels, correlating with symptom and plant development. Proc. Natl. Acad. Sci. USA 2007, 104, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Ding, Z.; Wu, K.; Yang, L.; Li, Y.; Yang, Z.; Shi, S.; Liu, X.; Zhao, S.; Yang, Z.; et al. Suppression of Jasmonic Acid-Mediated Defense by Viral-Inducible MicroRNA319 Facilitates Virus Infection in Rice. Mol. Plant 2016, 9, 1302–1314. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, F.; Li, J.; Chen, J.P.; Zhang, H.M. Integrative Analysis of the microRNAome and Transcriptome Illuminates the Response of Susceptible Rice Plants to Rice Stripe Virus. PLoS ONE 2016, 11, e0146946. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Mou, G.; Wang, K.; Zhou, G. MicroRNAs responding to southern rice black-streaked dwarf virus infection and their target genes associated with symptom development in rice. Virus Res. 2014, 190, 60–68. [Google Scholar] [CrossRef]
- Wang, B.; Wang, L.; Chen, F.; Yang, X.; Ding, M.; Zhang, Z.; Liu, S.S.; Wang, X.W.; Zhou, X. MicroRNA profiling of the whitefly Bemisia tabaci Middle East-Aisa Minor I following the acquisition of Tomato yellow leaf curl China virus. Virol. J. 2016, 13, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.; Wang, Z.Q.; Liu, X.; Zhao, L.; Zhou, X.; Xie, Y. Identification and Analysis of Potential Genes Regulated by an Alphasatellite (TYLCCNA) that Contribute to Host Resistance against Tomato Yellow Leaf Curl China Virus and Its Betasatellite (TYLCCNV/TYLCCNB) Infection in Nicotiana benthamiana. Viruses 2019, 11, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, P.; Wu, J.; Zhang, J.; Zhao, S.; Zheng, H.; Gao, G.; Wei, L.; Li, Y. Viral infection induces expression of novel phased microRNAs from conserved cellular microRNA precursors. PLoS Pathog. 2011, 7, e1002176. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Qin, G. EXB1/WRKY71 transcription factor regulates both shoot branching and responses to abiotic stresses. Plant. Signal. Behav. 2016, 11, e1150404. [Google Scholar] [CrossRef] [PubMed]
- Amin, I.; Patil, B.L.; Briddon, R.W.; Mansoor, S.; Fauquet, C.M. Comparison of phenotypes produced in response to transient expression of genes encoded by four distinct begomoviruses in Nicotiana benthamiana and their correlation with the levels of developmental miRNAs. Virol. J. 2011, 8, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Guo, W.; Li, F.; Sunter, G.; Zhou, X. Geminivirus-Associated Betasatellites: Exploiting Chinks in the Antiviral Arsenal of Plants. Trends Plant Sci. 2019, 24, 519–529. [Google Scholar] [CrossRef]
- Amin, I.; Patil, B.L.; Briddon, R.W.; Mansoor, S.; Fauquet, C.M. A common set of developmental miRNAs are upregulated in Nicotiana benthamiana by diverse begomoviruses. Virol. J. 2011, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, A.R.; Haq, Q.M.R.; Mukherjee, S.K. MicroRNA profiling of tomato leaf curl new delhi virus (tolcndv) infected tomato leaves indicates that deregulation of mir159/319 and mir172 might be linked with leaf curl disease. Virol. J. 2010, 7, 281. [Google Scholar] [CrossRef] [Green Version]
- Romanel, E.; Silva, T.F.; Correa, R.L.; Farinelli, L.; Hawkins, J.S.; Schrago, C.E.; Vaslin, M.F. Global alteration of microRNAs and transposon-derived small RNAs in cotton (Gossypium hirsutum) during Cotton leafroll dwarf polerovirus (CLRDV) infection. Plant. Mol. Biol. 2012, 80, 443–460. [Google Scholar] [CrossRef]
- Du, J.; Wu, G.; Zhou, Z.; Zhang, J.; Li, M.; Sun, M.; Jiang, K.; Qing, L. Identification of microRNAs regulated by tobacco curly shoot virus co-infection with its betasatellite in Nicotiana benthamiana. Virol. J. 2019, 16, 130. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Guo, W.; Wu, G.; Lu, Y.; Peng, J.; Zheng, H.; Lin, L.; Chen, J. A virus-based miRNA suppression (VbMS) system for miRNA loss-of-function analysis in plants. Biotechnol. J. 2014, 9, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Tang, X. Short tandem target mimic: A long journey to the engineered molecular landmine for selective destruction/blockage of microRNAs in plants and animals. J. Genet. Genom. 2013, 40, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wu, G.; Li, M.; Ma, M.; Du, J.; Sun, M.; Sun, X.; Qing, L. Transcriptome analysis of Nicotiana benthamiana infected by Tobacco curly shoot virus. Virol. J. 2018, 15, 138. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xie, Y.; Zhou, X. Tobacco curly shoot virusDNAβ Is Not Necessary for Infection but Intensifies Symptoms in a Host-Dependent Manner. Phytopathology 2005, 95, 902–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, Z.; Chen, J. A high throughput DNA extraction method with high yield and quality. Plant. Methods 2012, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Zorzatto, C.; Machado, J.P.; Lopes, K.V.; Nascimento, K.J.; Pereira, W.A.; Brustolini, O.J.; Reis, P.A.; Calil, I.P.; Deguchi, M.; Sachetto-Martins, G. NIK1-mediated translation suppression functions as a plant antiviral immunity mechanism. Nature 2015, 520, 679. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nature Protocols 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Rodrígueznegrete, E.A.; Sánchezcampos, S.; Cañizares, M.C.; Navascastillo, J.; Moriones, E.; Bejarano, E.R.; Grandepérez, A. A sensitive method for the quantification of virion-sense and complementary-sense DNA strands of circular single-stranded DNA viruses. Sci. Rep. 2014, 4, 6438. [Google Scholar] [CrossRef] [Green Version]
- Caifu, C.; Ridzon, D.A.; Broomer, A.J.; Zhaohui, Z.; Lee, D.H.; Nguyen, J.T.; Maura, B.; Nan Lan, X.; Mahuvakar, V.R.; Andersen, M.R. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 2005, 33, e179. [Google Scholar]
- He, Q.; Peng, J.; Yan, F.; Lin, L.; Lu, Y.; Zheng, H.; Chen, H.; Chen, J. Intron retention and 3′-UTR analysis of Arabidopsis Dicer-like 2 transcripts. Mol Biol Rep. 2012, 39, 3271–3280. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Liu, Q.; Hu, P.; Jia, Q.; Liu, N.; Yin, K.; Cheng, Y.; Yan, F.; Chen, J.; Liu, Y. An efficient Potato virus X-based microRNA silencing in Nicotiana benthamiana. Sci. Rep. 2016, 6, 20573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sha, A.; Zhao, J.; Yin, K.; Tang, Y.; Wang, Y.; Wei, X.; Hong, Y.; Liu, Y. Virus-Based MicroRNA Silencing in Plants. Plant. Physiol. 2013, 164, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Baksa, I.; Nagy, T.; Barta, E.; Havelda, Z.; Várallyay, É.; Silhavy, D.; Burgyán, J.; Szittya, G. Identification of Nicotiana benthamiana microRNAs and their targets using high throughput sequencing and degradome analysis. BMC Genom. 2015, 16, 1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, A.; Yuan, Q.; Wang, S.; Peng, J.; Lu, Y.; Zheng, H.; Lin, L.; Chen, H.; Gong, Y.; Chen, J. Altered accumulation of osa-miR171b contributes to rice stripe virus infection by regulating disease symptoms. J. Exp. Bot. 2017, 68, 4357–4367. [Google Scholar] [CrossRef]
- Wang, S.; Cui, W.; Wu, X.; Yuan, Q.; Zhao, J.; Zheng, H.; Lu, Y.; Peng, J.; Lin, L.; Chen, J.; et al. Suppression of nbe-miR166h-p5 attenuates leaf yellowing symptoms of potato virus X on Nicotiana benthamiana and reduces virus accumulation. Mol. Plant. Pathol. 2018, 19, 2384–2396. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Target Gene | Expectation | UPE | miRNA Start | miRNA End | Target Start | Target End | miRNA Aligned Fragment | Target Aligned Fragment | Inhibition | Multiplicity |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nbe-miR1919c-5p | Niben101Scf04663g00007 | 2.0 | 15.475 | 1 | 21 | 254 | 274 | UGUCGCAGAUGACUUUCGCCC | AUGCGAAAGUCAUCUGCGACA | Cleavage | 1 |
| Nbe-miR1919c-5p | Niben101Scf02655g01001 | 2.0 | 16.74 | 1 | 21 | 485 | 505 | UGUCGCAGAUGACUUUCGCCC | AUGCGAAAGUCAUCUGCGACA | Cleavage | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, J.; Wu, R.; Liu, Z.; Sun, M.; Ghanem, H.; Li, M.; Wu, G.; Qing, L. Suppression of nbe-miR1919c-5p Expression in Nicotiana benthamiana Enhances Tobacco Curly Shoot Virus and Its Betasatellite Co-Infection. Viruses 2020, 12, 392. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040392
Du J, Wu R, Liu Z, Sun M, Ghanem H, Li M, Wu G, Qing L. Suppression of nbe-miR1919c-5p Expression in Nicotiana benthamiana Enhances Tobacco Curly Shoot Virus and Its Betasatellite Co-Infection. Viruses. 2020; 12(4):392. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040392
Chicago/Turabian StyleDu, Jiang, Rui Wu, Zhuoying Liu, Miao Sun, Hussein Ghanem, Mingjun Li, Gentu Wu, and Ling Qing. 2020. "Suppression of nbe-miR1919c-5p Expression in Nicotiana benthamiana Enhances Tobacco Curly Shoot Virus and Its Betasatellite Co-Infection" Viruses 12, no. 4: 392. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040392