The Multifarious Role of 14-3-3 Family of Proteins in Viral Replication


1
School of Science, Monash University, Bandar Sunway, Selangor Darul Ehsan 47500, Malaysia
2
Tropical Medicine & Biology Platform, Monash University, Bandar Sunway, Selangor Darul Ehsan 47500, Malaysia
*
Author to whom correspondence should be addressed.
Viruses 2020, 12(4), 436; https://0-doi-org.brum.beds.ac.uk/10.3390/v12040436
Submission received: 13 March 2020
/
Revised: 6 April 2020
/
Accepted: 7 April 2020
/
Published: 13 April 2020
(This article belongs to the Special Issue Influenza A Virus: Host-Virus Relationship)
Abstract
:The 14-3-3 proteins are a family of ubiquitous and exclusively eukaryotic proteins with an astoundingly significant number of binding partners. Their binding alters the activity, stability, localization, and phosphorylation state of a target protein. The association of 14-3-3 proteins with the regulation of a wide range of general and specific signaling pathways suggests their crucial role in health and disease. Recent studies have linked 14-3-3 to several RNA and DNA viruses that may contribute to the pathogenesis and progression of infections. Therefore, comprehensive knowledge of host–virus interactions is vital for understanding the viral life cycle and developing effective therapeutic strategies. Moreover, pharmaceutical research is already moving towards targeting host proteins in the control of virus pathogenesis. As such, targeting the right host protein to interrupt host–virus interactions could be an effective therapeutic strategy. In this review, we generated a 14-3-3 protein interactions roadmap in viruses, using the freely available Virusmentha network, an online virus–virus or virus–host interaction tool. Furthermore, we summarize the role of the 14-3-3 family in RNA and DNA viruses. The participation of 14-3-3 in viral infections underlines its significance as a key regulator for the expression of host and viral proteins.
1. Introduction
Viruses are a leading source of death and have a significant impact on global health. They are unique pathogens which rely on living organisms to complete their life cycle [1]. Therefore, identifying the viral and host factors involved in viral infection is the main aim of virologists, in order to develop antiviral drugs. Drugs targeting viral proteins have clear disadvantages, such as viral subtype specificity, the rapid development of drug resistance, and low fidelity polymerases in the case of HIV and influenza viruses [2]. Therefore, considering the increasing threat of viral infections, focus is emerging on the need to identify novel, effective, and safe alternative therapies for viral infections. Targeting host proteins is an efficient alternative, since viruses are dependent on host proteins at multiple stages to achieve their replication cycles [3,4,5].
Recently, the 14-3-3 family of proteins has received much attention in the field of disease progression and drug development [6,7,8,9]. The 14-3-3 proteins belong to the highly conserved acidic protein family, which consists of seven mammalian isoforms (beta, epsilon, gamma, eta, sigma, tau, and zeta) [10]. Initially, they were identified in 1967 as abundant brain proteins [11]. In this review, we focus on the 14-3-3 family of proteins and their multifaceted roles in a variety of viral infections.
1.1. An Overview of the 14-3-3 Family
Historically, 14-3-3 proteins were named based on their elution and the migration pattern of their two-dimensional DEAE-cellulose chromatography and starch gel electrophoresis [12].
The 14-3-3 proteins are essential regulators of intracellular signaling pathways and upon interacting with their target protein, they modify its function and intracellular localization [13]. Furthermore, they are involved in many processes, including cell-cycle control, cell metabolism, apoptosis, and gene transcription control [10,14,15,16]. Hence, it is vital to understand the role of 14-3-3 during viral infection. In addition, the structure of 14-3-3 provides its great capability to bind to a multitude of functionally diverse signaling proteins. This property allows 14-3-3 proteins to affect multiple pathways in the host cell and thus become the target of a variety of viruses modulating them to alter cellular processes to their advantage.
1.2. Structure and General Function of the 14-3-3 Family
The crystal structure of 14-3-3 proteins comprises of highly helical and cup-shaped dimers [17]. The cup-like structure of 14-3-3 dimers is the key to their diverse functions and target binding properties. Moreover, the rigid structure of the 14-3-3 dimer, and the frequent presence of 14-3-3 binding motifs in a single target molecule and within disordered regions, are the standard features which are essential for 14-3-3 proteins to bind to and regulate their binding partners [18]. The 14-3-3 dimers contain nine α-helices, which are organized in an anti-parallel array. Among them, helices αA, αC and αD are involved in dimerization; however, helices αC, αE, αG and αI form a concave amphipathic groove at the ligand-binding site [19,20]. Typically, 14-3-3 proteins bind to phosphorylated serine and threonine residues, with two optimal binding motifs on their targets: RSXpSXP and RXXXpSXP [21]. In some cases, they can also bind to non-phosphorylated targets [21,22].
The general function of 14-3-3 proteins can be classified based on the following characteristics [14,21]: (A) conformational changes of its binding protein: the rigid structure of 14-3-3 proteins may serve as a stable base to support the reshaping of their binding partners and the altering of their activities; (B) masking of a phosphorylated region of a binding protein: 14-3-3 protein binding can also mask significant surface features of the target protein; (C) act as a scaffold molecule or adapter between two other binding proteins [14,21].
Furthermore, various post-translational modifications, such as phosphorylation, acetylation, acylation, polyglycylation and oxidative stress, are documented for 14-3-3 proteins [23,24]. Eventually, these post-translational changes of 14-3-3 isoforms, or their interacting proteins, may have several effects on 14-3-3 proteins, including dimerization specificity, functional regulation, and cellular localization [25]. All these diverse functions of 14-3-3 proteins contribute to an array of protein–protein interactions among the host proteins, and host–virus interactions.
2. Host–Virus Interactions
Viruses are a major contributor to the global burden of diseases because they cause acute and chronic infectious diseases [26,27]. Many studies have shown how viruses hijack cellular pathways and evade the host’s innate immune response, starting from entry, until the virus exits from host cells by modulating host factors and signaling pathways [28,29]. Therefore, comprehensive knowledge of host–virus interactions is important in order to understand the viral life cycle and to develop specific therapeutic approaches for the treatment of viral infections. Hence, the study of protein–protein interactions and their communication networks within infected cells has become an important tool for the understanding of the mechanisms of viral replication and, in the long run, leading to the discovery of new anti-viral targets.
2.1. Protein–Protein Interactions
Protein–protein interaction (PPI) is one of the primary components of system biology and a key mediator for host–virus interactions. PPI can be detected by in-vitro, in-vivo, and in-silico studies. The in-vitro methods include tandem affinity purification (TAP), co-immunoprecipitation (co-IP), protein fragment complementation, protein arrays, phage displays, X-ray crystallography and nuclear magnetic resonance (NMR) spectroscopy [29,30,31,32]. In contrast, in-vivo methods include yeast two-hybrid (Y2H) [33] and synthetic lethality techniques [34,35]. For in-silico techniques, usually, computer simulations are used to perform sequence- [36] and structure-based predictions [37], gene fusion [38], and in-silico two-hybrid [39], chromosome proximity [40] and gene expression-based approaches [41,42].
2.2. Protein–Protein Interaction Network
The interaction between proteins is one of the most critical processes in the execution of cellular processes. In system biology research, protein–protein interaction networking (PPIN) is an essential tool for the identification of interactions between protein pairs. Therefore, the importance of understanding this PPI has led to the development of experimental and computational interaction network tools. As discussed above, a wide variety of methods have been developed to study these interactions.
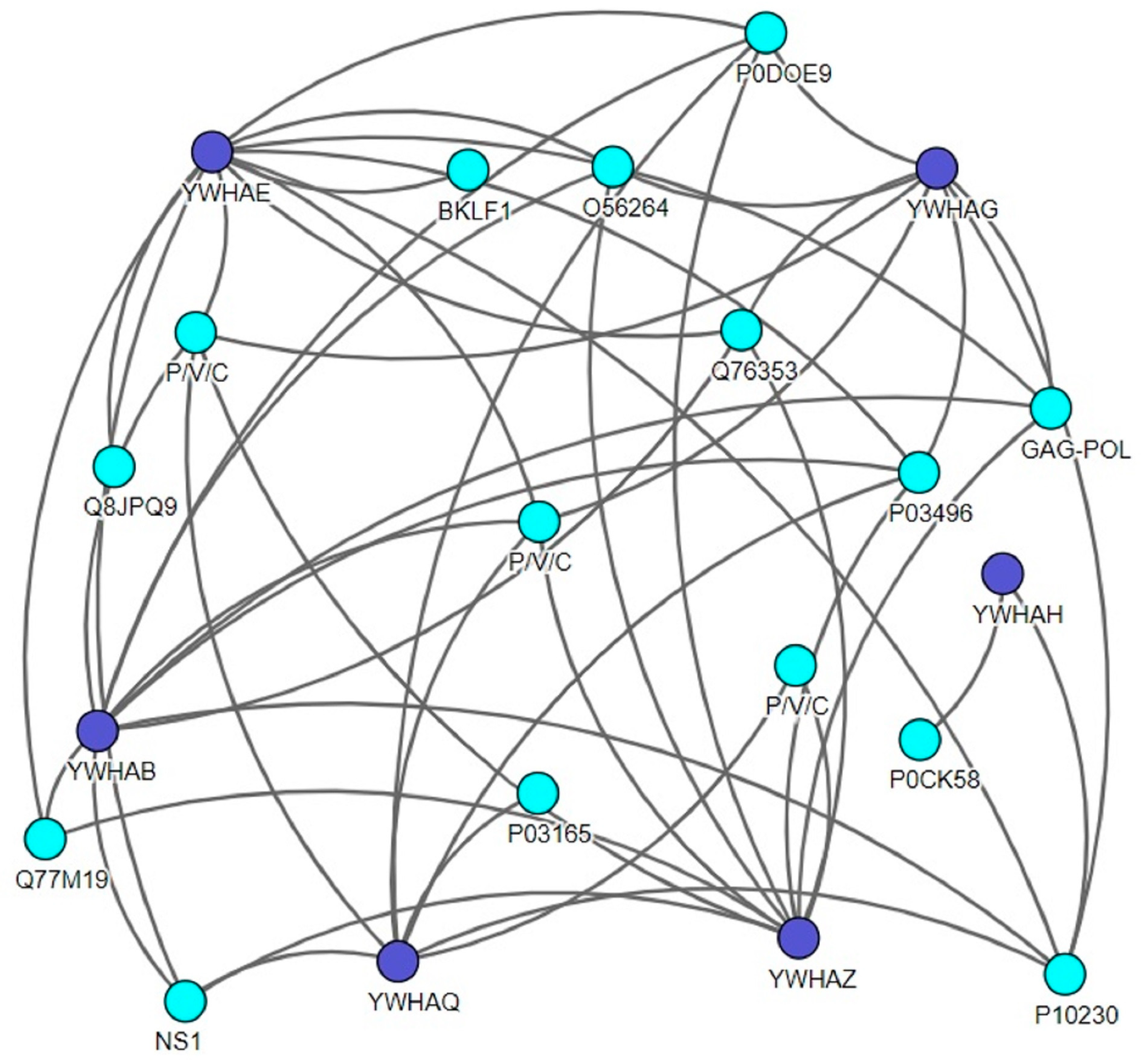
To date, there are many tools and web-based technologies that allow these data to be explored, providing different methods to visualize the interaction of the protein network. Here, to identify the direct host interaction partners for all viral proteins, we generated a protein–protein interaction map using the Virusmentha network (Figure 1). The Virusmentha database, which is freely available at https://virusmentha.uniroma2.it/, is the most comprehensive automatically updating resource available to date for viral interactions [42,43].
The isoforms of 14-3-3 proteins, like β, θ, ε, γ, η, and ζ, were found to interact with single-stranded RNA viruses (ssRNA) such as the influenza A virus (IAV), measles virus, human respiratory syncytial virus, human immunodeficiency virus (HIV), La Crosse virus, and double-stranded DNA (dsDNA) viruses like herpes simplex virus type I, human herpes 4, hepatitis B virus (HBV), Nipah virus, Hendra virus and Murid herpesvirus (Table 1). Investigators have confirmed these interactions using different approaches and methods, as described in Table 1. Among all the isoforms of the 14-3-3 family, ζ, ε, and θ are the most established and well-studied in the context of viral infections [46,47,48,49,50,51]. Similarly, co-IP using tandem affinity purification (TAP) and pull-down techniques are the most extensively used methods in the study of protein–protein interactions.
3. Role of 14-3-3 Family Members in Virus Infection
The 14-3-3 proteins are associated with many cellular processes, which could probably implicate them in many human illnesses. Therefore, the characterization of their functions at the molecular level is of the utmost importance. Given the multitude of its binding partners and its essential roles in numerous biological processes, the 14-3-3 family of proteins should be considered pathfinders for further exploration of closely related viruses.
Viruses have been shown to arrest host cells by hijacking and manipulating host protein complexes to promote the development of infection. Generally, viruses are classified based on the Baltimore Classification System invented by David Baltimore, which is primarily based on their genome composition and replication strategy [54]. Although there are seven different replication strategies in the system, the primary stages in viral replication are virus attachment, entry, uncoating, transcription, the synthesis of virus components, virion assembly, and the release of progeny virions [55]. However, they are unable to replicate without the machinery and metabolism of a host cell. To further explore the roles of host–virus interaction in the viral life cycle, we discuss the importance of 14-3-3 proteins in RNA and DNA viruses in the subsequent sections.
3.1. Role of 14-3-3 Family in RNA Viruses
RNA viruses have ssRNA or dsRNA as their genetic material [56]. In addition, based on the polarity of the RNA, they can be classified into a negative sense or a positive sense [57]. In contrast, dsRNA viruses are a diverse group of viruses that vary widely in their range of hosts (animals, bacteria, plants and fungi), genome segment numbers (one to twelve), and organization of virions (T-number and capsid layers) [58]. Like many viruses, host cells play an essential role in RNA virus replication as well. To date, several isoforms of 14-3-3 proteins are reported to interact with, and play an essential role in, RNA virus infection [59,60,61,62,63,64,65]. Moreover, discussions regarding the 14-3-3 family have dominated research in recent years for their promising progress, which also elucidates their enormous role in controlling cell cycles, apoptosis and signaling pathways.
In 2000, Aoki et al. demonstrated that 14-3-3 proteins associate with the hepatitis C virus (HCV) core protein to activate RAF-1 kinase and contribute to hepatocyte growth regulation [59]. Further, the HCV core protein induces Bax-mediated apoptosis through HCV core–14-3-3 ε protein interaction [60]. In general, the host factor contributes positively to the budding of many negative-stranded RNA viruses, as reviewed elsewhere [66,67]. However, in the case of parainfluenza virus 5 (PIV5), the M protein interacts with host protein 14-3-3 β and negatively affects the production of PIV5 particles [61]. This shows that the interaction of the M protein with 14-3-3 β is not related to virus budding.
The 14-3-3 proteins have a crucial role to play in the regulation of cell cycle progression [10,68]. They regulate cell cycle progression by altering the activities of cell division cycle 25C (Cdc25C), cyclin B1 (a regulatory protein involved in mitosis), checkpoint kinase 1 (Chk1), and wee1 [10,69,70]. The 14-3-3 proteins interact with the accessory protein Vpr (viral protein R) of human immunodeficiency virus type I (HIV-1) and significantly regulate cell cycle progression by associating with Cdc25C [62]. Hence, Vpr induces G2-M cell cycle arrest by increasing the association between 14-3-3 θ and cell cycle regulatory factors (Cdc25C, cyclin B1, and Cdk1) during HIV infection [63]. In addition, Vpr interferes with, and alters the activity of, forkhead transcription factors (Foxo3a) by inhibiting their association with 14-3-3 proteins [71].
Similarly, in the case of adeno-associated virus type 2 (AAV-2) infection, 14-3-3 ε and 14-3-3 γ interact with the Rep68 protein of AAV-2 by phosphorylation at serine 535 [64]. Interestingly, the Rep78 of AAV-2 does not interact with 14-3-3 proteins. Another key point is that the phosphorylation of Rep68 in the C-terminal seven amino acid (LARGHS) of Rep68 is necessary for the interaction of 14-3-3 with Rep68. Moreover, Yaffe et al. described that 14-3-3 proteins’ interaction with target proteins often entails the phosphorylation of a serine residue [11]. Thus, this observation may provide a hint to understanding the role of Rep68 in AAV infections. Furthermore, 14-3-3-Rep68 interaction down-regulates the DNA binding activity of the target protein in an AAV infection [65]. This suggests that 14-3-3 proteins are functionally involved in the regulation of the Rep68 protein during the viral life cycle.
Members of the14-3-3 family regulate numerous intracellular processes, one of which is immunity [72]. Immunity which is 14-3-3-mediated may guide the rational design of therapeutics, and this can be seen in many virus infections [73] as well as mosquito-transmitted Zika and dengue viruses [74,75,76]. In a very recent study, the NS3 protein of the Zika virus interacted with 14-3-3 ε and 14-3-3 η to inhibit RIG-I and the melanoma differentiation-associated protein 5 (MDA5) signaling pathway [75]. Notably, 14-3-3 η was also reported to promote antiviral signaling by MDA5 in HCV infection [77]. In the dengue virus, 14-3-3 ε binds to the NS3 protein and prevents RIG-I from translocating to the mitochondrial antiviral signaling (MAVS) protein adapter, thereby blocking antiviral signaling [76]. Liu et al. reported 14-3-3 ε as a critical mediator for the relocation of RIG-I from cytosol to mitochondrial-associated MAVS, as it establishes a translocon complex with RIG-I and TRIM25, eventually triggering an antiviral response [78].
Previous studies from our laboratory have shown that the 14-3-3 protein also binds to the severe acute respiratory syndrome (SARS) coronavirus nucleocapsid (N) protein [79]. This binding allows the regulation of nucleocytoplasmic shuttling of N protein in a phosphorylation-dependent manner. Additionally, our group has shown that N protein induces apoptosis by reducing the expression level of 14-3-3 θ in the absence of growth factors (serum), which contributes to the accumulation of the phosphorylated N protein in the nucleus [78]. Likewise, coronavirus 2, or COVID-19, originated from China sharing similar demographic profiles, and laboratory and radiological findings with SARS and Middle East respiratory syndrome (MERS) [80]. In SARS, our group has demonstrated that N protein is phosphorylated by multiple kinases (cyclin-dependent kinase, glycogen synthase kinase, and casein kinase II) and significantly interferes with cellular machinery by binding to 14-3-3 proteins through phosphorylation-dependent protein–protein interactions. This suggests that this discovery may provide new insights into a possible mechanism for the phosphorylation-dependent nucleocytoplasmic shuttling of the N protein and could be a potential antiviral strategy against COVID-19.
In addition, a proteomics report showed that the influenza A virus (IAV) NS1 protein interacts with several 14-3-3 isoforms, although the functional implication of this interaction remains unclear [73].
3.2. Role of 14-3-3 in DNA Viruses
DNA viruses have DNA as their genetic material and replicate either by host or by virally encoded DNA polymerases [81]. Isoforms of the 14-3-3 family are reported to play essential roles in the life cycle of DNA viruses like the hepatitis B virus (HBV), herpes simplex virus type (HSV-I) and Epstein–Barr virus (EBV) [51,82,83,84]. For instance, in the HBV life cycle, 14-3-3 ζ is responsible for maintaining the HBV protein x (HBx) expression in hepatocellular carcinoma cells (HCC). The knockdown of 14-3-3 ζ reduces the expression of HBx. This suggests that 14-3-3 ζ–HBx interaction could be a potential therapeutic target for HBV-related hepatocellular carcinoma [51]. Over the past few years, research has shown that 14-3-3 proteins are key regulators of many processes, including mitosis and apoptosis in animals [85]. According to Kim et al., HBx induces apoptosis by inhibiting the association between 14-3-3 ε and Bax, thereby enhancing mitochondrial–Bax translocation and cytochrome C release [86]. Consequently, a potential 14-3-3 binding motif of HBx is crucial for stress-activated protein kinases (SAPK)/ Jun amino-terminal kinases (JNK) activitity and Fas-mediated apoptosis protection [87].
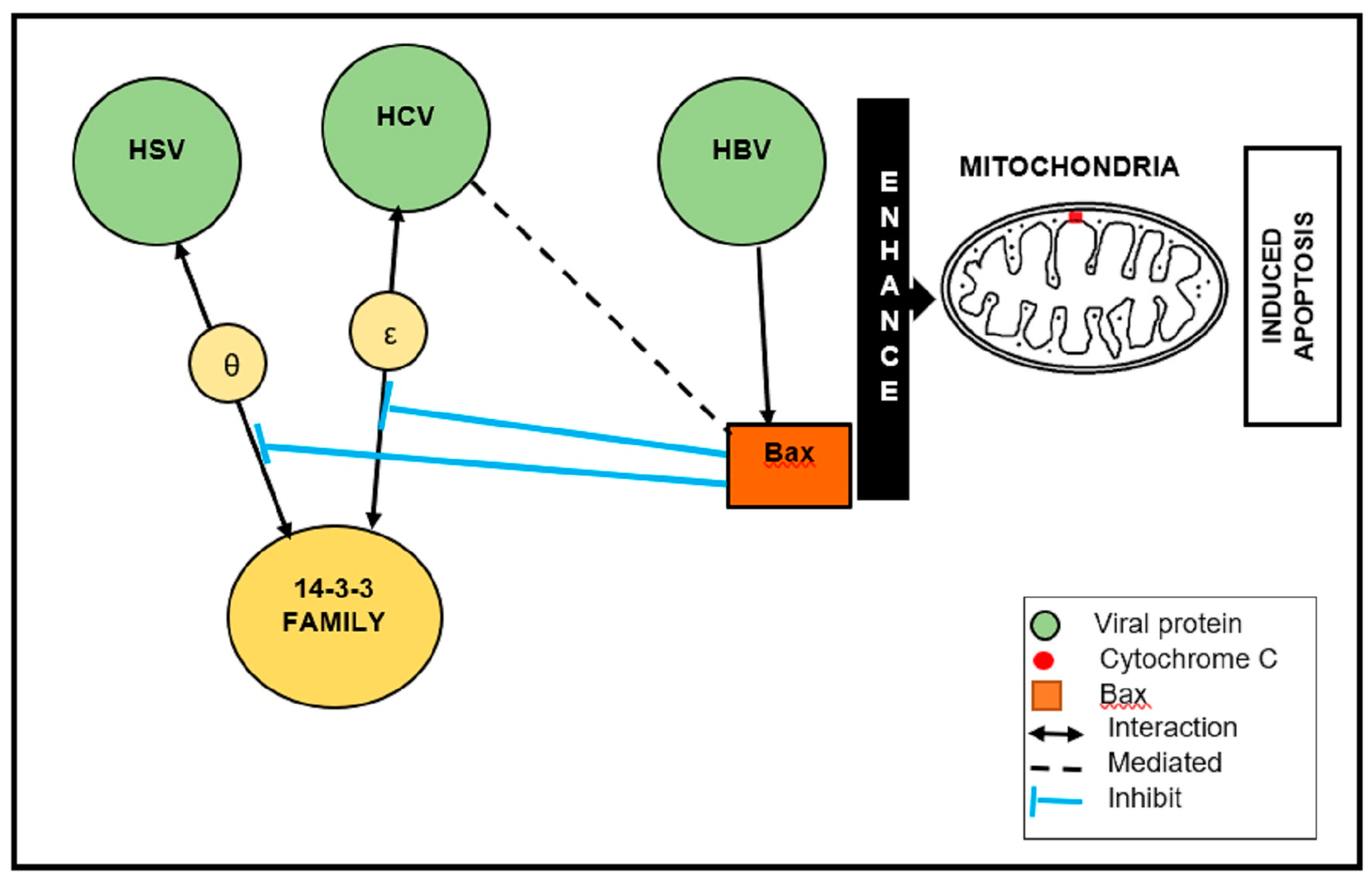
In the case of HSV-I, ICP27 is an essential protein for viral replication because it is involved in the nuclear export of viral mRNA and the suppression of host protein synthesis by inhibiting cellular mRNA splicing [82,83]. ICP27–14-3-3 θ interaction sequesters Bax to the cytoplasm. Further, ICP27 inhibits the interaction between 14-3-3 θ and Bax [82]. It is interesting to note that HBV, HCV, and HSV directly associate with, or inhibit, cellular protein 14-3-3 to induce apoptosis (Figure 2). Typically, (pro-apoptotic) Bax is a member of the Bcl-2 family and a key apoptosis regulator. Upon apoptotic stimulation, Bax is activated and oligomerized in the mitochondrial membrane (MOM) to mediate its permeability [88]. Therefore, 14-3-3 proteins are shown to promote Bax-mediated apoptosis, resulting in Bax being activated and MOM permeabilization.
Gupta et al. identified several members of the 14-3-3 protein family (ζ, ε, γ, β, η) as interacting partners of EBV-encoded large tegument protein deneddylase (BPLF1) using co-IP and mass spectrometry approaches [84]. Interactions between 14-3-3 and BPLF1 may participate in the regulation of many signaling pathways; ubiquitin ligases, cullin 1 (CUL1) and tripartite motif-containing protein 25 (TRIM25) are the possible partners in this regulation. Furthermore, 14-3-3 proteins and TRIM25 serve as vital co-factors in the signaling of viral nucleic acid sensors, such as retinoic acid-inducible gene I (RIG-I) and MDA5 [78]. In this case, 14-3-3 proteins stabilize the interaction of TRIM25 with RIG-I, thus facilitating the ubiquitination of RIG-I [89].
In addition, we also demonstrate and summarize all the roles of 14-3-3 proteins in different viruses (Table 2). The apoptosis pathway, cell cycle, cell signaling, ubiquitin ligase, and nucleocytoplasmic shuttling are the most common pathways which were usurped and manipulated by the viruses to favor their replication.
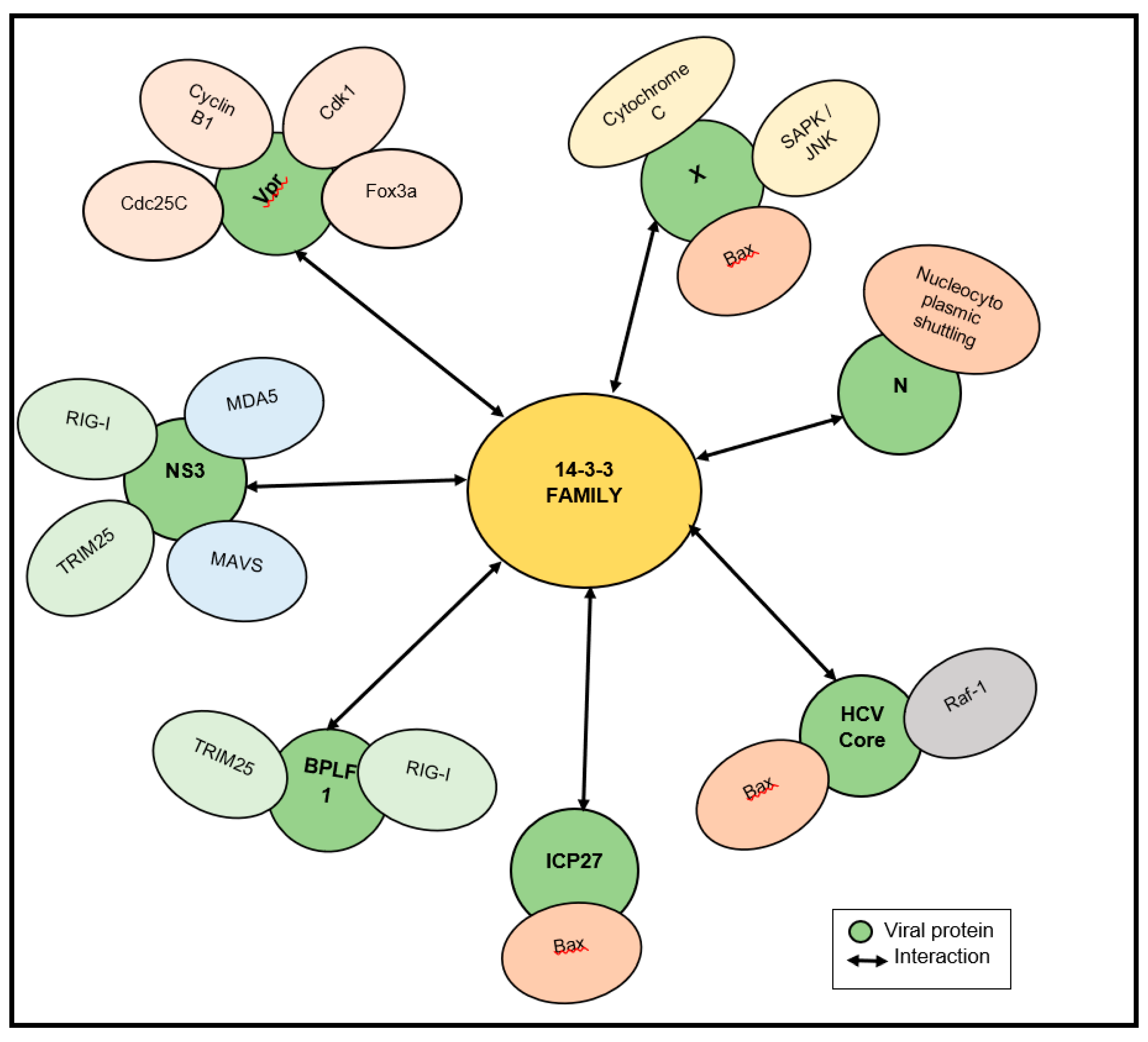
We also elaborate on all the signaling pathways that are manipulated by 14-3-3–virus interactions (Figure 3). It is interesting to note that cellular proteins are essential for the replication of many RNA and DNA viruses, and may serve as viable targets for treating viral infections.
4. Conclusions
Viruses exploit the molecular machinery of the infected host to support their replication. To achieve this, viruses establish virus-specific protein interactions to perturb several cellular processes in the infected host [90,91]. Thus, a comprehensive understanding of the perturbation of the host–virus relationship and virus infection are essential for the development of antiviral therapies. In order to provide an overview of host–virus interactions, we generated a host (14-3-3 proteins)-specific interaction map using Virusmentha, an online tool. Based on this database, 14-3-3 ζ, ε, and θ are well known in the context of viral infections.
Furthermore, 14-3-3 family members display a significant role in interacting with several RNA and DNA viruses through multiple pathways, including apoptosis, cell signaling, cell cycle, and ubiquitination. These viral–14-3-3 interactions may change the typical distribution and disrupt the original functions of 14-3-3 proteins. For example, HBV, HCV, and HSV viruses were directly associated with, or inhibited, 14-3-3 proteins from inducing apoptosis. Therefore, monitoring and manipulating 14-3-3 proteins may represent new diagnostic and therapeutic targets for virus infections. The development of therapeutics, including drugs and vaccines, is highly dependent on the knowledge gained from investigating host–virus interactions. As such, interrupting host–virus interactions by targeting the right host factor might be a highly effective strategy for treating viral infections.
Funding
This work was supported by an expired FRGS grant (grant#FRGS/1/2015/SKK08/MUSM/)01/1 from the Ministry of Higher Education (MOHE) that expired in November 2018.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Becker, K.; Hu, Y.; Biller-Andorno, N. Infectious diseases–a global challenge. Int. J. Med. Microbiol. 2006, 296, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.-H.; Spindler, K.R. Identifying Host Factors That Regulate Viral Infection. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.-Y.; Yen, H.-L. Targeting the Host or the Virus: Current and Novel Concepts for Antiviral Approaches against Influenza Virus Infection. Antivir. Res. 2012, 96, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Watanabe, S.; Kawaoka, Y. Cellular Networks Involved in the Influenza Virus Life Cycle. Cell Host Microbe 2010, 7, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Kawaoka, Y. Influenza Virus–Host Interactomes as a Basis for Antiviral Drug Development. Curr. Opin. Virol. 2015, 14, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masters, S.C.; Subramanian, R.R.; Truong, A.; Yang, H.; Fujii, K.; Zhang, H.; Fu, H. Survival-Promoting Functions of 14-3-3 Proteins. Biochem. Soc. Trans. 2002, 30, 360–365. [Google Scholar] [CrossRef]
- Molzan, M.; Kasper, S.; Röglin, L.; Skwarczynska, M.; Sassa, T.; Inoue, T.; Breitenbuecher, F.; Ohkanda, J.; Kato, N.; Schuler, M.; et al. Stabilization of Physical RAF/14-3-3 Interaction by Cotylenin A as Treatment Strategy for RAS Mutant Cancers. ACS Chem. Biol. 2013, 8, 1869–1875. [Google Scholar] [CrossRef]
- Bier, D.; Rose, R.; Bravo-Rodriguez, K.; Bartel, M.; Ramirez-Anguita, J.M.; Dutt, S.; Wilch, C.; Klärner, F.-G.; Sanchez-Garcia, E.; Schrader, T.; et al. Molecular Tweezers Modulate 14-3-3 Protein–Protein Interactions. Nat. Chem. 2013, 5, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Du, Y.; Horton, J.R.; Upadhyay, A.K.; Lou, B.; Bai, Y.; Zhang, X.; Du, L.; Li, M.; Wang, B.; et al. Discovery and Structural Characterization of a Small Molecule 14-3-3 Protein-Protein Interaction Inhibitor. Proc. Natl. Acad. Sci. USA 2011, 108, 16212–16216. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Subramaniam, R.R. 14-3-3 proteins: Structure, function, and regulation. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 617–647. [Google Scholar] [CrossRef]
- Yaffe, M.B.; Rittinger, K.; Volinia, S.; Caron, P.R.; Aitken, A.; Leffers, H.; Gamblin, S.J.; Smerdon, S.J.; Cantley, L.C. The Structural Basis for 14-3-3: Phosphopeptide Binding Specificity. Cell 1997, 91, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Skoulakis, E.M.C.; Davis, R.L. 14-3-3 Proteins in Neuronal Development and Function. Mol. Neurobiol. 1998, 16, 269–284. [Google Scholar] [CrossRef]
- Aghazadeh, Y.; Papadopoulos, V. The role of 14-3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Obsilova, V.; Silhan, J.; Boura, E.; Teisinger, J.; Obsil, T. 14-3-3 proteins: A family of versatile molecular regulators. Physiol. Res. 2018, 57, 11–21. [Google Scholar]
- Aitken, A. 14-3-3 Proteins: A Historic Overview. Semin. Cancer Biol. 2006, 16, 162–172. [Google Scholar] [CrossRef]
- Hermeking, H.; Benzinger, A. 14-3-3 proteins in cell cycle regulation. Semin. Cancer Biol. 2006, 16, 183–192. [Google Scholar] [CrossRef]
- Liu, D.; Bienkowska, J.; Petosa, C.; Collier, R.J.; Fu, H.; Liddington, R. Crystal Structure of the Zeta Isoform of the 14-3-3 Protein. Nature 1995, 376, 191–194. [Google Scholar] [CrossRef]
- Obsil, T.; Obsilova, V. Structural basis of 14-3-3 protein functions. Semin. Cell Dev. Biol. 2011, 663–672. [Google Scholar] [CrossRef]
- Xiao, B.; Smerdon, S.J.; Jones, D.H.; Dodson, G.G.; Soneji, Y.; Aitken, A.; Gamblin, S.J. Structure of a 14-3-3 Protein and Implications for Coordination of Multiple Signalling Pathways. Nature 1995, 376, 188–191. [Google Scholar] [CrossRef]
- Rittinger, K.; Budman, J.; Xu, J.; Volinia, S.; Cantley, L.C.; Smerdon, S.J.; Gamblin, S.J.; Yaffe, M.B. Structural Analysis of 14-3-3 Phosphopeptide Complexes Identifies a Dual Role for the Nuclear Export Signal of 14-3-3 in Ligand Binding. Mol. Cell 1999, 4, 153–166. [Google Scholar] [CrossRef]
- Campbell, J.K.; Gurung, R.; Romero, S.; Speed, C.J.; Andrews, R.K.; Berndt, M.C.; Mitchell, C.A. Activation of the 43 KDa Inositol Polyphosphate 5-Phosphatase by 14-3-3ζ†. Biochemistry 1997, 36, 15363–15370. [Google Scholar] [CrossRef] [PubMed]
- Muslin, A.J.; Tanner, J.; Allen, P.M.; Shaw, A.S. Interaction of 14-3-3 with Signaling Proteins Is Mediated by the Recognition of Phosphoserine. Cell 1996, 84, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Lalle, M.; Bavassano, C.; Fratini, F.; Cecchetti, S.; Boisguerin, P.; Crescenzi, M.; Pozio, E. Involvement of 14-3-3 Protein Post-Translational Modifications in Giardia Duodenalis Encystation. Int. J. Parasitol. 2010, 40, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Musiani, F.; Ciurli, S.; Dikiy, A. Interaction of Selenoprotein W with 14-3-3 Proteins: A Computational Approach. J. Proteome Res. 2011, 10, 968–976. [Google Scholar] [CrossRef]
- Legand, A.; Briand, S.; Shindo, N.; Brooks, W.A.; Jong, M.D.D.; Farrar, J.; Aguilera, X.; Hayden, F.G. Addressing the Public Health Burden of Respiratory Viruses: The Battle against Respiratory Viruses (BRaVe) Initiative. Future Virol. 2013, 8, 953–968. [Google Scholar] [CrossRef]
- Brown, C.R.; Maclachlan, J.H.; Cowie, B.C. Addressing the Increasing Global Burden of Viral Hepatitis. Hepatobiliary Surg. Nutr. 2017, 6, 274–276. [Google Scholar] [CrossRef] [Green Version]
- Davey, N.E.; Travé, G.; Gibson, T.J. How Viruses Hijack Cell Regulation. Trends Biochem. Sci. 2011, 36, 159–169. [Google Scholar] [CrossRef]
- Mayer, K.A.; Stöckl, J.; Zlabinger, G.J.; Gualdoni, G.A. Hijacking the Supplies: Metabolism as a Novel Facet of Virus-Host Interaction. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Aitken, A. Post-Translational Modification of 14-3-3 Isoforms and Regulation of Cellular Function. Semin. Cell Dev. Biol. 2011, 22, 673–680. [Google Scholar] [CrossRef]
- Gavin, A.-C.; Grandi, P.; Boesche, M.; Krause, R.; Leutwein, C.; Kuster, B.; Superti-Furga, G. Functional Organization of the Yeast Proteome by Systemic Analysis of Protein Complexes. GBM Annu. Fall Meet. Halle 2002 2002, 2002, 141–147. [Google Scholar] [CrossRef]
- Pitre, S.; Alamgir, M.; Green, J.R.; Dumontier, M.; Dehne, F.; Golshani, A. Computational Methods for Predicting Protein–Protein Interactions. Adv. Biochem. Eng./Biotechnol. 2008, 110, 247–267. [Google Scholar]
- Michnick, S.W.; Ear, P.H.; Landry, C.; Malleshaiah, M.K.; Messier, V. Protein-Fragment Complementation Assays for Large-Scale Analysis, Functional Dissection and Dynamic Studies of Protein–Protein Interactions in Living Cells. Methods Mol. Biol. Signal Trans. Protoc. 2011, 395–425. [Google Scholar] [CrossRef]
- Smith, G. Filamentous Fusion Phage: Novel Expression Vectors That Display Cloned Antigens on the Virion Surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Chiba, T.; Ozawa, R.; Yoshida, M.; Hattori, M.; Sakaki, Y. A Comprehensive Two-Hybrid Analysis to Explore the Yeast Protein Interactome. Proc. Natl. Acad. Sci. USA 2001, 98, 4569–4574. [Google Scholar] [CrossRef] [Green Version]
- Ooi, S.L.; Pan, X.; Peyser, B.D.; Ye, P.; Meluh, P.B.; Yuan, D.S.; Irizarry, R.A.; Bader, J.S.; Spencer, F.A.; Boeke, J.D. Global Synthetic-Lethality Analysis and Yeast Functional Profiling. Trends Genet. 2006, 22, 56–63. [Google Scholar] [CrossRef]
- Brown, J.A.; Sherlock, G.; Myers, C.L.; Burrows, N.M.; Deng, C.; Wu, H.I.; Mccann, K.E.; Troyanskaya, O.G.; Brown, J.M. Global Analysis of Gene Function in Yeast by Quantitative Phenotypic Profiling. Mol. Syst. Biol. 2006, 2. [Google Scholar] [CrossRef] [Green Version]
- Valente, G.T.; Acencio, M.L.; Martins, C.; Lemke, N. The Development of a Universal In Silico Predictor of Protein-Protein Interactions. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.; Henrick, K.; Nakamura, H.; Markley, J.L. The Worldwide Protein Data Bank (WwPDB): Ensuring a Single, Uniform Archive of PDB Data. Nucleic Acids Res. 2007, 35, D301–D303. [Google Scholar] [CrossRef] [Green Version]
- Enright, A.J.; Iliopoulos, I.; Kyrpides, N.C.; Ouzounis, C.A. Protein Interaction Maps for Complete Genomes Based on Gene Fusion Events. Nature 1999, 402, 86–90. [Google Scholar] [CrossRef]
- Pazos, F.; Valencia, A. In Silico Two-Hybrid System for the Selection of Physically Interacting Protein Pairs. Proteins Struct. Funct. Genet. 2002, 47, 219–227. [Google Scholar] [CrossRef]
- Raes, J.; Korbel, J.O.; Lercher, M.J.; Von Mering, C.; Bork, P. Prediction of effective genome size in metagenomic samples. Genome Biol. 2007, 8, R10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigoriev, A. A Relationship between Gene Expression and Protein Interactions on the Proteome Scale: Analysis of the Bacteriophage T7 and the Yeast Saccharomyces Cerevisiae. Nucleic Acids Res. 2001, 29, 3513–3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderone, A.; Licata, L.; Cesareni, G. An integrated approach: A New Resource for Virus-Host Protein Interactions. Nucleic Acids Res. 2014, 43. [Google Scholar] [CrossRef]
- Virus Mentha: Viral Interactomes Browser. Available online: https://virusmentha.uniroma2.it/ (accessed on 3 April 2020).
- Bridges, D.; Moorhead, G.B.G. 14-3-3 Proteins: A Number of Functions for a Numbered Protein. Sci. Signal. 2005, 2005. [Google Scholar] [CrossRef] [PubMed]
- Pichlmair, A.; Kandasamy, K.; Alvisi, G.; Mulhern, O.; Sacco, R.; Habjan, M.; Binder, M.; Stefanovic, A.; Eberle, C.-A.; Goncalves, A.; et al. Viral Immune Modulators Perturb the Human Molecular Network by Common and Unique Strategies. Nature 2012, 487, 486–490. [Google Scholar] [CrossRef]
- Komarova, A.V.; Combredet, C.; Meyniel-Schicklin, L.; Chapelle, M.; Caignard, G.; Camadro, J.-M.; Lotteau, V.; Vidalain, P.-O.; Tangy, F. Proteomic Analysis of Virus-Host Interactions in an Infectious Context Using Recombinant Viruses. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef] [Green Version]
- Jager, S.; Cimermancic, P.; Gulbahce, N.; Johnson, J.R.; McGovern, K.E.; Clarke, S.C.; Shales, M.; Mercenne, G.; Pache, L.; Li, K.; et al. Global landscape of HIV-human protein complexes. Nature 2011, 481, 365–370. [Google Scholar] [CrossRef]
- Khattar, E.; Mukherji, A.; Kumar, V. Akt Augments the Oncogenic Potential of the HBx Protein of Hepatitis B Virus by Phosphorylation. FEBS J. 2012, 279, 1220–1230. [Google Scholar] [CrossRef]
- Wu, W.; Tran, K.C.; Teng, M.N.; Heesom, K.J.; Matthews, D.A.; Barr, J.N.; Hiscox, J.A. The Interactome of the Human Respiratory Syncytial Virus NS1 Protein Highlights Multiple Effects on Host Cell Biology. J. Virol. 2012, 86, 7777–7789. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Salwinski, L.; Zhang, C.; Chu, D.; Sampankanpanich, C.; Reyes, N.A.; Vangeloff, A.; Xing, F.; Li, X.; Wu, T.-T.; et al. An Integrated Approach to Elucidate the Intra-Viral and Viral-Cellular Protein Interaction Networks of a Gamma-Herpesvirus. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef]
- Lin, A.E.; Greco, T.M.; Döhner, K.; Sodeik, B.; Cristea, I.M. A Proteomic Perspective of Inbuilt Viral Protein Regulation: pUL46 Tegument Protein Is Targeted for Degradation by ICP0 during Herpes Simplex Virus Type 1 Infection. Mol. Cell. Proteom. 2013, 12, 3237–3252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozenblatt-Rosen, O.; Deo, R.C.; Padi, M.; Adelmant, G.; Calderwood, M.A.; Rolland, T.; Grace, M.; Dricot, A.; Askenazi, M.; Tavares, M.; et al. Interpreting Cancer Genomes Using Systematic Host Network Perturbations by Tumour Virus Proteins. Nature 2012, 487, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Dimmock, N.J.; Easton, A.J.; Leppard, K. Introduction to Modern Virology; Blackwell Pub.: Malden, MA, USA, 2007. [Google Scholar]
- Brooks, G.F.; Jawetz, E.; Melnick, J.L.; Adelberg, E.A. Jawetz, Melnick & Adelbergs Medical Microbiology: A Lange Medical Book; McGraw-Hill Medical: New York, NY, USA, 2013. [Google Scholar]
- Poltronieri, P.; Sun, B.; Mallardo, M. RNA Viruses: RNA Roles in Pathogenesis, Coreplication and Viral Load. Curr. Genom. 2015, 16, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.; Haenni, A.-L. Expression Strategies of Ambisense Viruses. Virus Res. 2003, 93, 141–150. [Google Scholar] [CrossRef]
- Patton, J.T. Segmented Double-Stranded RNA Viruses Structure and Molecular Biology; Caister Academic Press: Norfolk, UK, 2008. [Google Scholar]
- Aoki, H.; Hayashi, J.; Moriyama, M.; Arakawa, Y.; Hino, O. Hepatitis C Virus Core Protein Interacts with 14-3-3 Protein and Activates the Kinase Raf-1. J. Virol. 2000, 74, 1736–1741. [Google Scholar] [CrossRef] [Green Version]
- Diao, J.; Khine, A.A.; Sarangi, F.; Hsu, E.; Iorio, C.; Tibbles, L.A.; Woodgett, J.R.; Penninger, J.; Richardson, C.D. X Protein of Hepatitis B Virus Inhibits Fas-Mediated Apoptosis and Is Associated with Up-Regulation of the SAPK/JNK Pathway. J. Biol. Chem. 2000, 276, 8328–8340. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.P.; Lamb, R.A. Escaping from the Cell: Assembly and Budding of Negative-Strand RNA Viruses. Current Topics in Microbiology and Immunology Biology of Negative Strand RNA Viruses: The Power of Reverse Genetics; Springer: Berlin, Germany, 2004; pp. 145–196. [Google Scholar]
- Graves, P.R.; Lovly, C.M.; Uy, G.L.; Piwnica-Worms, H. Localization of Human Cdc25C Is Regulated Both by Nuclear Export and 14-3-3 Protein Binding. Oncogene 2001, 20, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Kino, T.; Gragerov, A.; Valentin, A.; Tsopanomihalou, M.; Ilyina-Gragerova, G.; Erwin-Cohen, R.; Chrousos, G.P.; Pavlakis, G.N. Vpr Protein of Human Immunodeficiency Virus Type 1 Binds to 14-3-3 Proteins and Facilitates Complex Formation with Cdc25C: Implications for Cell Cycle Arrest. J. Virol. 2005, 79, 2780–2787. [Google Scholar] [CrossRef] [Green Version]
- Kino, T.; Martino, M.U.D.; Charmandari, E.; Ichijo, T.; Outas, T.; Chrousos, G.P. HIV-1 Accessory Protein Vpr Inhibits the Effect of Insulin on the Foxo Subfamily of Forkhead Transcription Factors by Interfering With Their Binding to 14-3-3 Proteins: Potential Clinical Implications Regarding the Insulin Resistance of HIV-1-Infected Patients. Diabetes 2004, 54, 23–31. [Google Scholar]
- Han, S.-I.; Kawano, M.-A.; Ishizu, K.-I.; Watanabe, H.; Hasegawa, M.; Kanesashi, S.-N.; Kim, Y.-S.; Nakanishi, A.; Kataoka, K.; Handa, H. Rep68 Protein of Adeno-Associated Virus Type 2 Interacts with 14-3-3 Proteins Depending on Phosphorylation at Serine 535. Virology 2004, 320, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Pena-Blanco, A.; Garcia-Saez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. FEBS J. 2017, 285, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Calistri, A.; Salata, C.; Parolin, C.; Palù, G. Role of Multivesicular Bodies and Their Components in the Egress of Enveloped RNA Viruses. Rev. Med. Virol. 2009, 19, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.; Harrison, M.S.; Schmitt, A.P. Parainfluenza Virus 5 M Protein Interaction with Host Protein 14-3-3 Negatively Affects Virus Particle Formation. J. Virol. 2010, 85, 2050–2059. [Google Scholar] [CrossRef] [Green Version]
- Dalal, S.N.; Schweitzer, C.M.; Gan, J.; Decaprio, J.A. Cytoplasmic Localization of Human cdc25C during Interphase Requires an Intact 14-3-3 Binding Site. Mol. Cell. Biol. 1999, 19, 4465–4479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Forbes, K.C.; Wu, Z.; Moreno, S.; Piwnica-Worms, H.; Enoch, T. Replication Checkpoint Requires Phosphorylation of the Phosphatase Cdc25 by Cds1 or Chk1. Nature 1998, 395, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Bolton, D.L.; Barnitz, R.A.; Sakai, K.; Lenardo, M.J. 14-3-3 Theta Binding to Cell Cycle Regulatory Factors Is Enhanced by HIV-1 Vpr. Biol. Direct 2008, 3, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzivion, G.; Shen, Y.H.; Zhu, J. 14-3-3 Proteins; Bringing New Definitions to Scaffolding. Oncogene 2001, 20, 6331–6338. [Google Scholar] [CrossRef] [Green Version]
- Riedl, W.; Acharya, D.; Lee, J.-H.; Liu, G.; Serman, T.; Chiang, C.; Chan, Y.K.; Diamond, M.S.; Gack, M.U. Zika Virus NS3 Mimics a Cellular 14-3-3-Binding Motif to Antagonize RIG-I- and MDA5-Mediated Innate Immunity. Cell Host Microbe 2019, 26. [Google Scholar] [CrossRef]
- Lin, J.-P.; Fan, Y.-K.; Liu, H.M. The 14-3-3η Chaperone Protein Promotes Antiviral Innate Immunity via Facilitating MDA5 Oligomerization and Intracellular Redistribution. PLoS Pathog. 2019, 15. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.K.; Gack, M.U. A Phosphomimetic-Based Mechanism of Dengue Virus to Antagonize Innate Immunity. Nat. Immunol. 2016, 17, 523–530. [Google Scholar] [CrossRef]
- Surjit, M.; Kumar, R.; Mishra, R.N.; Reddy, M.K.; Chow, V.T.K.; Lal, S.K. The Severe Acute Respiratory Syndrome Coronavirus Nucleocapsid Protein Is Phosphorylated and Localizes in the Cytoplasm by 14-3-3-Mediated Translocation. J. Virol. 2005, 79, 11476–11486. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.M.; Loo, Y.-M.; Horner, S.M.; Zornetzer, G.A.; Katze, M.G.; Gale, M. The Mitochondrial Targeting Chaperone 14-3-3ε Regulates a RIG-I Translocon That Mediates Membrane Association and Innate Antiviral Immunity. Cell Host Microbe 2012, 11, 528–537. [Google Scholar]
- Yan, G.; Pang, L.; Cook, A.R.; Ho, H.J.; Win, M.S.; Khoo, A.L.; Wong, J.G.; Lee, C.K.; Yan, B.; Jureen, R.; et al. Distinguishing Zika and Dengue Viruses through Simple Clinical Assessment, Singapore. Emerg. Infect. Dis. 2018, 24, 1565–1568. [Google Scholar] [CrossRef]
- Kuo, R.-L.; Li, Z.-H.; Li, L.-H.; Lee, K.-M.; Tam, E.-H.; Liu, H.M.; Liu, H.-P.; Shih, S.-R.; Wu, C.-C. Interactome Analysis of the NS1 Protein Encoded by Influenza A H1N1 Virus Reveals a Positive Regulatory Role of Host Protein PRP19 in Viral Replication. J. Proteome Res. 2016, 15, 1639–1648. [Google Scholar] [CrossRef]
- Petrosillo, N.; Viceconte, G.; Ergonul, O.; Ippolito, G.; Petersen, E. COVID-19, SARS and MERS: Are They Closely Related? Clin. Microbiol. Infect. 2020. [Google Scholar] [CrossRef]
- Payne, S. Introduction to DNA Viruses. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/B9780128031094000283?via%3Dihub (accessed on 12 March 2020).
- Lee, S.K.; Park, S.O.; Joe, C.O.; Kim, Y.S. Interaction of HCV Core Protein with 14-3-3ε Protein Releases Bax to Activate Apoptosis. Biochem. Biophys. Res. Commun. 2007, 352, 756–762. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, J.C.; Min, J.S.; Kang, I.; Oh, J.; Ahn, J.K. HSV-1 ICP27 Induces Apoptosis by Promoting Bax Translocation to Mitochondria through Interacting with 14-3-3θ. BMB Rep. 2017, 50, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Ylä-Anttila, P.; Callegari, S.; Tsai, M.-H.; Delecluse, H.-J.; Masucci, M.G. Herpesvirus Deconjugases Inhibit the IFN Response by Promoting TRIM25 Autoubiquitination and Functional Inactivation of the RIG-I Signalosome. PLoS Pathog. 2018, 14. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Zhang, Y.; Wang, C.; Sun, Z.; Li, L.; Dong, J.; Zhou, W. 14-3-3ζ Binds to Hepatitis B Virus Protein X and Maintains Its Protein Stability in Hepatocellular Carcinoma Cells. Cancer Med. 2018, 7, 5543–5553. [Google Scholar] [CrossRef] [Green Version]
- Rosenquist, M. 14-3-3 proteins in apoptosis. Braz. J. Med. Biol. Res. 2003, 36, 403–408. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, S.Y.; Kim, J.; Lee, H.; Choi, M.; Kim, J.K.; Ahn, J.K. Hepatitis B Virus X Protein Induces Apoptosis by Enhancing Translocation of Bax to Mitochondria. IUBMB Life 2008, 60, 473–480. [Google Scholar] [CrossRef]
- Chen, I.-H.B.; Li, L.; Silva, L.; Sandri-Goldin, R.M. ICP27 Recruits Aly/REF but Not TAP/NXF1 to Herpes Simplex Virus Type 1 Transcription Sites Although TAP/NXF1 Is Required for ICP27 Export. J. Virol. 2005, 79, 3949–3961. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.-R.; Kim, H.-I.; Choi, M.-S.; Yi, C.-M.; Inn, K.-S. Regulation of MDA5-MAVS Antiviral Signaling Axis by TRIM25 through TRAF6-Mediated NF-ΚB Activation. Mol. Cells 2015, 38, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Chisari, F.V. Unscrambling Hepatitis C Virus–Host Interactions. Nature 2005, 436, 930–932. [Google Scholar] [CrossRef]
- Lanford, R.E.; Guerra, B.; Lee, H.; Averett, D.R.; Pfeiffer, B.; Chavez, D.; Notvall, L.; Bigger, C. Antiviral Effect and Virus-Host Interactions in Response to Alpha Interferon, Gamma Interferon, Poly(I)-Poly(C), Tumor Necrosis Factor Alpha, and Ribavirin in Hepatitis C Virus Subgenomic Replicons. J. Virol. 2003, 77, 1092–1104. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The Virusmentha network showing a host (14-3-3s)–virus interaction map. Lines between the proteins indicate interactions between proteins. Purple nodes indicate the different 14-3-3 isoforms, and blue nodes indicate the viral proteins. The Virusmentha database is available at https://virusmentha.uniroma2.it/ [44,45].
Figure 1.
The Virusmentha network showing a host (14-3-3s)–virus interaction map. Lines between the proteins indicate interactions between proteins. Purple nodes indicate the different 14-3-3 isoforms, and blue nodes indicate the viral proteins. The Virusmentha database is available at https://virusmentha.uniroma2.it/ [44,45].

Figure 2.
Schematic illustration of the Bax-mediated apoptosis of herpes simplex virus (HBV), hepatitis C virus (HCV), and hepatitis B virus (HBV). HBV, HCV, and HSV directly associate with, or inhibit, cellular protein 14-3-3 to induce apoptosis.
Figure 2.
Schematic illustration of the Bax-mediated apoptosis of herpes simplex virus (HBV), hepatitis C virus (HCV), and hepatitis B virus (HBV). HBV, HCV, and HSV directly associate with, or inhibit, cellular protein 14-3-3 to induce apoptosis.

Figure 3.
Schematic illustration of the signaling pathways which are manipulated by virus–14-3-3 interactions. Yellow nodes indicate the 14-3-3 family, green nodes indicate the viral proteins and other nodes indicate the functional proteins in signaling pathways.
Figure 3.
Schematic illustration of the signaling pathways which are manipulated by virus–14-3-3 interactions. Yellow nodes indicate the 14-3-3 family, green nodes indicate the viral proteins and other nodes indicate the functional proteins in signaling pathways.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of virus–host protein (14-3-3) interactions using different methods.
| UniProt ID | Host Protein 14-3-3 | Method | Viruses | Target Gene/Protein | Ref |
|---|---|---|---|---|---|
| P03496 | Beta/alpha | TAP | Influenza A Virus (strain A/Puerto Rico/8/1934) | NS | [46] |
| Q77M19 | Beta/alpha, zeta, epsilon | Co-IP | Measles virus strain Schwarz | P | [47] |
| Q76353 | Beta/alpha, zeta, epsilon | Physical association by using the pull-down, anti-tag co-IP, | Human immunodeficiency virus I (HIV-I) | - | [48] |
| P10230 | Eta | Physical association by using Affinity chromatography technology | Herpes simplex virus (type 1 / strain 17) | Tegument protein UL46 | [52] |
| P03165 | Theta | Physical association by Anti-bait co-IP | HBV ayw/France/Tiollais/1979 | X | [49] |
| GAG-POL | Beta/alpha, gamma, epsilon, zeta | Physical association by using the Affinity chromatography technology | HIV -I | Gag-pol | [48] |
| P0DOE9 | Beta/alpha, zeta, epsilon, gamma, theta | co-IP | Human respiratory syncytial virus A2 | 1C | [50] |
| P0CK58 | Eta | TAP | Human herpesvirus 4 (strain B95-8) | Apoptosis regulator BALF1 | [53] |
| Q8JPQ9 | Epsilon | TAP | La Crosse virus L78 | N | [46] |
| Q997F2 | Theta, zeta | TAP | Nipah virus | P/V/C | [46] |
| P0C1C6 | Gamma, theta, epsilon, beta/alpha, zeta | TAP | Hendra virus horse/Australia/Hendra/1994 | P/V/C | [46] |
| P0C1C7 | Gamma, theta, epsilon, beta/alpha, zeta | TAP | Nipah virus | P/V/C | [46] |
| Q9WPI5 | Theta | TAP | IAV (A/Texas/36/1991(H1N1)) | NS1 | [46] |
| P88993 | Epsilon | Physical association by using the Two-hybrid array | Murid herpesvirus 4 | BKLF1 | [51] |
Table 2.
Summary of the 14-3-3 family’s roles in different viruses.
| 14-3-3 | Genome | Viruses | Target Protein | Pathway/Function | Ref |
|---|---|---|---|---|---|
| ζ, ε | dsDNA | Hepatitis B virus | Protein x | Bax-mediated apoptosis | [86] |
| ε | (+) ssRNA | Hepatitis C virus | HCV core | Bax-mediated apoptosis | [60] |
| θ | dsDNA | Herpes simplex virus type I | ICP27 | Bax-mediated apoptosis | [82,88] |
| β | (−) ssRNA | Parainfluenza virus 5 | M protein | Virus budding | [61] |
| θ | (+) ssRNA | Human immunodeficiency virus type I | Vpr | Cell cycle | [62,63] |
| ε, γ | ssDNA | Adeno-associated virus type 2 | Rep68 | Virus replication | [65] |
| ζ, ε, γ, β, η | dsDNA | Epstein-Barr virus | BPLF1 | Cell signaling and ubiquitin ligase | [78,89] |
| ε, η | (+) ssRNA | Zika virus | NS3 | Cell signaling and ubiquitin ligase | [75] |
| θ | (+) ssRNA | Coronavirus | N | Nucleocytoplasmic shuttling | [78,79] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nathan, K.G.; Lal, S.K. The Multifarious Role of 14-3-3 Family of Proteins in Viral Replication. Viruses 2020, 12, 436. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040436
AMA Style
Nathan KG, Lal SK. The Multifarious Role of 14-3-3 Family of Proteins in Viral Replication. Viruses. 2020; 12(4):436. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040436
Chicago/Turabian StyleNathan, Kavitha Ganesan, and Sunil K. Lal. 2020. "The Multifarious Role of 14-3-3 Family of Proteins in Viral Replication" Viruses 12, no. 4: 436. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040436
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.