Efficacy of Heterologous Prime-Boost Vaccination with H3N2 Influenza Viruses in Pre-Immune Individuals: Studies in the Pig Model




Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Vaccines
2.2. Experimental Design
2.3. Serological Assays
2.4. Virus Titration and Lesion Scores
2.5. Tissue Collection and Processing for ELISPOT Assays
2.6. ASC and ISC ELISPOT
2.7. Statistics
3. Results
3.1. Serum Antibody Responses and Protection against Challenge after Vaccination of Infection-Immune Pigs
3.1.1. Serum Antibody Responses against the Strains Used for Infection and Vaccination
3.1.2. Serum Antibody Responses against Antigenically Distinct Influenza Strains
3.1.3. Protection against Challenge
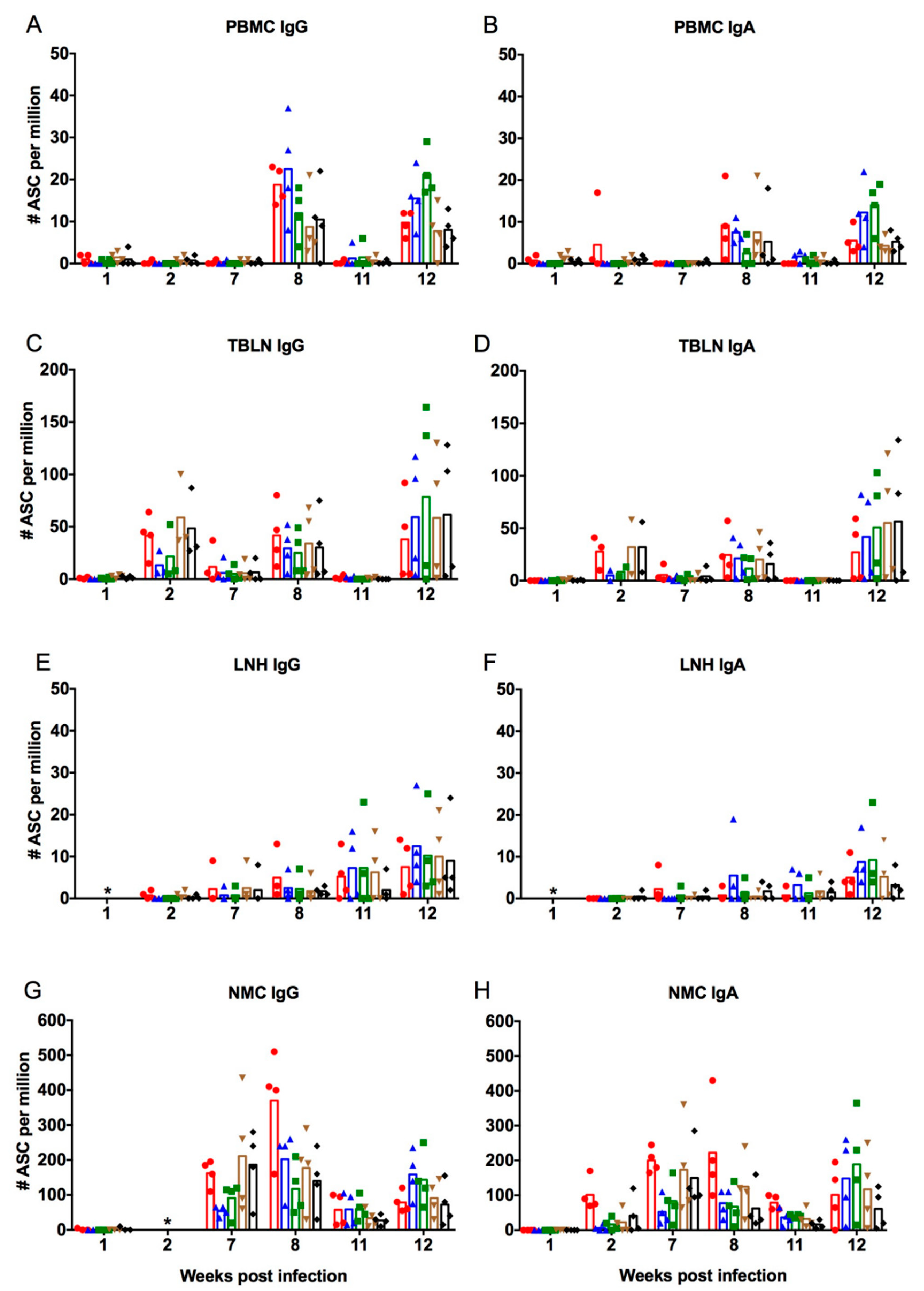
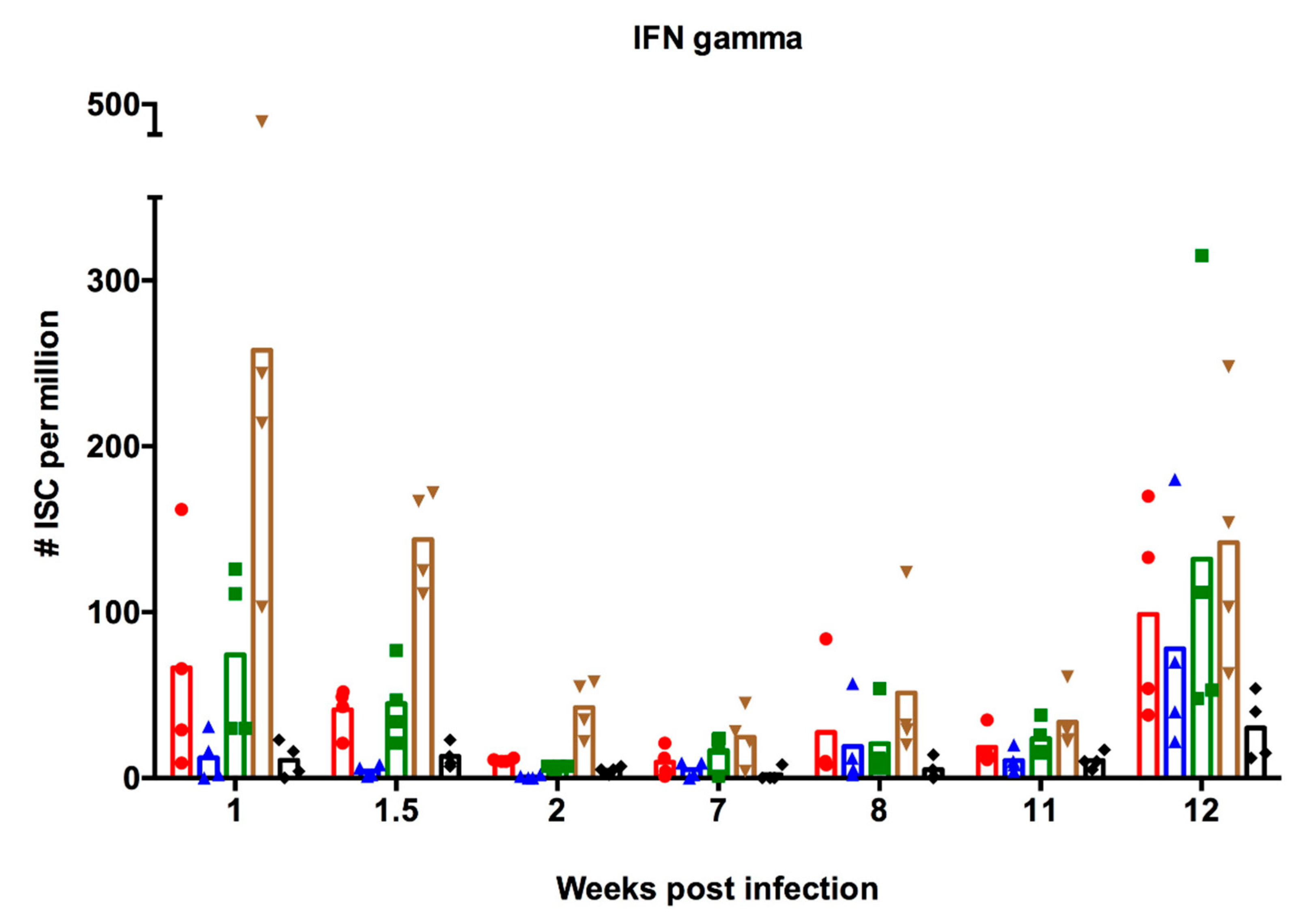
3.2. Antibody-Secreting and Interferon-Secreting Cell Responses after Vaccination of Infection-Immune Pigs
3.2.1. Antibody-Secreting Cell Responses
3.2.2. Interferon-Secreting Cell Responses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nelson, M.I.; Vincent, A.L. Reverse zoonosis of influenza to swine: New perspectives on the human-animal interface. Trends Microbiol. 2015, 23, 142–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Reeth, K.; Vincent, A. Influenza viruses. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Incorporated: Hoboken, NJ, USA, 2019; pp. 576–593. [Google Scholar]
- Nelson, M.I.; Wentworth, D.E.; Culhane, M.R.; Vincent, A.L.; Viboud, C.; LaPointe, M.P.; Lin, X.; Holmes, E.C.; Detmer, S.E. Introductions and evolution of human-origin seasonal influenza a viruses in multinational swine populations. J. Virol. 2014, 88, 10110–10119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haesebrouck, F.; Biront, P.; Pensaert, M.B.; Leunen, J. Epizootics of respiratory tract disease in swine in Belgium due to H3N2 influenza virus and experimental reproduction of disease. Am. J. Vet. Res. 1985, 46, 1926–1928. [Google Scholar] [PubMed]
- de Jong, J.C.; Smith, D.J.; Lapedes, A.S.; Donatelli, I.; Campitelli, L.; Barigazzi, G.; Van Reeth, K.; Jones, T.C.; Rimmelzwaan, G.F.; Osterhaus, A.D.; et al. Antigenic and genetic evolution of swine influenza A (H3N2) viruses in Europe. J. Virol. 2007, 81, 4315–4322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, N.S.; Russell, C.A.; Langat, P.; Anderson, T.K.; Berger, K.; Bielejec, F.; Burke, D.F.; Dudas, G.; Fonville, J.M.; Fouchier, R.A.; et al. The global antigenic diversity of swine influenza A viruses. Elife 2016, 5, e12217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, A.S.; Nelson, S.W.; Page, S.L.; Nolting, J.M.; Killian, M.L.; Sreevatsan, S.; Slemons, R.D. Swine-to-human transmission of influenza A (H3N2) virus at agricultural fairs, Ohio, USA, 2012. Emerg. Infect. Dis. 2014, 20, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Bowman, A.S.; Walia, R.R.; Nolting, J.M.; Vincent, A.L.; Killian, M.L.; Zentkovich, M.M.; Lorbach, J.N.; Lauterbach, S.E.; Anderson, T.K.; Davis, C.T.; et al. Influenza A (H3N2) virus in swine at agricultural fairs and transmission to humans, Michigan and Ohio, USA, 2016. Emerg. Infect. Dis. 2017, 23, 1551–1555. [Google Scholar] [CrossRef]
- Epperson, S.; Jhung, M.; Richards, S.; Quinlisk, P.; Ball, L.; Moll, M.; Boulton, R.; Haddy, L.; Biggerstaff, M.; Brammer, L.; et al. Human infections with influenza A (H3N2) variant virus in the United States, 2011–2012. Clin. Infect. Dis. 2013, 57, S4–S11. [Google Scholar] [CrossRef] [Green Version]
- Duwell, M.M.; Blythe, D.; Radebaugh, M.W.; Kough, E.M.; Bachaus, B.; Crum, D.A.; Perkins, K.A., Jr.; Blanton, L.; Davis, C.T.; Jang, Y.; et al. Influenza A(H3N2) variant virus outbreak at three Fairs—Maryland, 2017. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1169–1173. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, T.; Lambrecht, B.; Marche, S.; Steensels, M.; Van Borm, S.; Bublot, M. Influenza vaccines and vaccination strategies in birds. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 121–165. [Google Scholar] [CrossRef]
- Stephenson, I.; Nicholson, K.G.; Hoschler, K.; Zambon, M.C.; Hancock, K.; DeVos, J.; Katz, J.M.; Praus, M.; Banzhoff, A. Antigenically distinct MF59-adjuvanted vaccine to boost immunity to H5N1. N. Engl. J. Med. 2008, 359, 1631–1633. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.; Hancock, K.; Hoschler, K.; DeVos, J.; Praus, M.; Bardelli, M.; Malzone, C.; Castellino, F.; Gentile, C.; McNally, T.; et al. Fast rise of broadly cross-reactive antibodies after boosting long-lived human memory B cells primed by an MF59 adjuvanted prepandemic vaccine. Proc. Natl. Acad. Sci. USA 2009, 106, 7962–7967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Giudice, G.; Stittelaar, K.J.; van Amerongen, G.; Simon, J.; Osterhaus, A.D.; Stohr, K.; Rappuoli, R. Seasonal influenza vaccine provides priming for A/H1N1 immunization. Sci. Transl. Med. 2009, 1, 12re1. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Tan, G.S.; Hai, R.; Pica, N.; Petersen, E.; Moran, T.M.; Palese, P. Broadly protective monoclonal antibodies against H3 influenza viruses following sequential immunization with different hemagglutinins. PLoS Pathog. 2010, 6, e1000796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Brand, J.M.; Kreijtz, J.H.; Bodewes, R.; Stittelaar, K.J.; van Amerongen, G.; Kuiken, T.; Simon, J.; Fouchier, R.A.; Del Giudice, G.; Rappuoli, R.; et al. Efficacy of vaccination with different combinations of MF59-adjuvanted and nonadjuvanted seasonal and pandemic influenza vaccines against pandemic H1N1 (2009) influenza virus infection in ferrets. J. Virol. 2011, 85, 2851–2858. [Google Scholar] [CrossRef] [Green Version]
- Van Reeth, K.; Gracia, J.C.M.; Trus, I.; Sys, L.; Claes, G.; Versnaeyen, H.; Cox, E.; Krammer, F.; Qiu, Y. Heterologous prime-boost vaccination with H3N2 influenza viruses of swine favors cross-clade antibody responses and protection. NPJ Vaccines 2017, 2. [Google Scholar] [CrossRef]
- Erbelding, E.J.; Post, D.J.; Stemmy, E.J.; Roberts, P.C.; Augustine, A.D.; Ferguson, S.; Paules, C.I.; Graham, B.S.; Fauci, A.S. A Universal influenza vaccine: The strategic plan for the national institute of allergy and infectious diseases. J. Infect. Dis. 2018, 218, 347–354. [Google Scholar] [CrossRef]
- Francis, M.E.; King, M.L.; Kelvin, A.A. Back to the future for influenza preimmunity-looking back at influenza virus history to infer the outcome of future infections. Viruses 2019, 11, 122. [Google Scholar] [CrossRef] [Green Version]
- Guthmiller, J.J.; Wilson, P.C. Harnessing immune history to combat influenza viruses. Curr. Opin. Immunol. 2018, 53, 187–195. [Google Scholar] [CrossRef]
- Fonville, J.M.; Wilks, S.H.; James, S.L.; Fox, A.; Ventresca, M.; Aban, M.; Xue, L.; Jones, T.C.; Le, N.M.H.; Pham, Q.T.; et al. Antibody landscapes after influenza virus infection or vaccination. Science 2014, 346, 996–1000. [Google Scholar] [CrossRef] [Green Version]
- Lessler, J.; Riley, S.; Read, J.M.; Wang, S.; Zhu, H.; Smith, G.J.; Guan, Y.; Jiang, C.Q.; Cummings, D.A. Evidence for antigenic seniority in influenza A (H3N2) antibody responses in southern China. PLoS Pathog. 2012, 8, e1002802. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.S.; Gardner, T.J.; Krammer, F.; Aguado, L.C.; Tortorella, D.; Basler, C.F.; Palese, P. Neutralizing antibodies against previously encountered influenza virus strains increase over time: A longitudinal analysis. Sci. Transl. Med. 2013, 5, 198ra107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Van Reeth, K.; Nauwynck, H.; Pensaert, M. Bronchoalveolar interferon-alpha, tumor necrosis factor-alpha, interleukin-1, and inflammation during acute influenza in pigs: A possible model for humans? J. Infect. Dis. 1998, 177, 1076–1079. [Google Scholar] [CrossRef] [Green Version]
- Rajao, D.S.; Vincent, A.L. Swine as a model for influenza A virus infection and immunity. ILAR J. 2015, 56, 44–52. [Google Scholar] [CrossRef]
- Bodewes, R.; de Mutsert, G.; van der Klis, F.R.M.; Ventresca, M.; Wilks, S.; Smith, D.J.; Koopmans, M.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Rimmelzwaan, G.F. Prevalence of antibodies against seasonal Influenza A and B Viruses in children in Netherlands. Clin. Vaccine Immunol. 2011, 18, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Nunez, I.A.; Carlock, M.A.; Allen, J.D.; Owino, S.O.; Moehling, K.K.; Nowalk, P.; Susick, M.; Diagle, K.; Sweeney, K.; Mundle, S.; et al. Impact of age and pre-existing influenza immune responses in humans receiving split inactivated influenza vaccine on the induction of the breadth of antibodies to influenza a strains. PLoS ONE 2017, 12, e0185666. [Google Scholar] [CrossRef] [Green Version]
- Rajão, D.S.; Gauger, P.C.; Anderson, T.K.; Lewis, N.S.; Abente, E.J.; Killian, M.L.; Perez, D.R.; Sutton, T.C.; Zhang, J.; Vincent, A.L.; et al. Novel reassortant human-like H3N2 and H3N1 influenza A viruses detected in pigs are virulent and antigenically distinct from swine viruses endemic to the United States. J. Virol. 2015, 89, 11213–11222. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lambre, C.R.; Terzidis, H.; Greffard, A.; Webster, R.G. Measurement of anti-influenza neuraminidase antibody using a peroxidase-linked lectin and microtitre plates coated with natural substrates. J. Immunol. Methods 1990, 135, 49–57. [Google Scholar] [CrossRef]
- Van Reeth, K.; Gregory, V.; Hay, A.; Pensaert, M. Protection against a European H1N2 swine influenza virus in pigs previously infected with H1N1 and/or H3N2 subtypes. Vaccine 2003, 21, 1375–1381. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Richt, J.A.; Janke, B.H.; Sandbulte, M.R.; Gauger, P.C.; Loving, C.L.; Webby, R.J.; Garcia-Sastre, A. Live attenuated influenza vaccine provides superior protection from heterologous infection in pigs with maternal antibodies without inducing vaccine-associated enhanced respiratory disease. J. Virol. 2012, 86, 10597–10605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, D.L.; Karasin, A.; Zuckermann, F.; Olsen, C.W. Systemic and mucosal immune responses to H1N1 influenza virus infection in pigs. Vet. Microbiol. 2000, 74, 117–131. [Google Scholar] [CrossRef]
- Kitikoon, P.; Loving, C.L.; Vincent, A.L. Antibody secreting cell assay for influenza A virus in swine. Methods Mol. Biol. 2014, 1161, 347–353. [Google Scholar] [CrossRef]
- Brockmeier, S.L.; Loving, C.L.; Nelson, E.A.; Miller, L.C.; Nicholson, T.L.; Register, K.B.; Grubman, M.J.; Brough, D.E.; Kehrli, M.E., Jr. The presence of alpha interferon at the time of infection alters the innate and adaptive immune responses to porcine reproductive and respiratory syndrome virus. Clin. Vaccine Immunol. 2012, 19, 508–514. [Google Scholar] [CrossRef]
- Gorres, J.P.; Lager, K.M.; Kong, W.P.; Royals, M.; Todd, J.P.; Vincent, A.L.; Wei, C.J.; Loving, C.L.; Zanella, E.L.; Janke, B.; et al. DNA vaccination elicits protective immune responses against pandemic and classic swine influenza viruses in pigs. Clin. Vaccine Immunol. 2011, 18, 1987–1995. [Google Scholar] [CrossRef] [Green Version]
- Zuckermann, F.A.; Husmann, R.J.; Schwartz, R.; Brandt, J.; Mateu de Antonio, E.; Martin, S. Interleukin-12 enhances the virus-specific interferon gamma response of pigs to an inactivated pseudorabies virus vaccine. Vet. Immunol. Immunopathol. 1998, 63, 57–67. [Google Scholar] [CrossRef]
- Coudeville, L.; Bailleux, F.; Riche, B.; Megas, F.; Andre, P.; Ecochard, R. Relationship between haemagglutination-inhibiting antibody titres and clinical protection against influenza: Development and application of a bayesian random-effects model. BMC Med. Res. Methodol. 2010, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Tsang, T.K.; Cauchemez, S.; Perera, R.A.; Freeman, G.; Fang, V.J.; Ip, D.K.; Leung, G.M.; Malik Peiris, J.S.; Cowling, B.J. Association between antibody titers and protection against influenza virus infection within households. J. Infect. Dis. 2014, 210, 684–692. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.J.; Lapedes, A.S.; de Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.; Fouchier, R.A. Mapping the antigenic and genetic evolution of influenza virus. Science 2004, 305, 371–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westgeest, K.B.; Russell, C.A.; Lin, X.; Spronken, M.I.J.; Bestebroer, T.M.; Bahl, J.; van Beek, R.; Skepner, E.; Halpin, R.A.; de Jong, J.C.; et al. Genomewide analysis of reassortment and evolution of human influenza A (H3N2) viruses circulating between 1968 and 2011. J. Virol. 2013, 88, 2844–2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, T.M.; Allen, J.D.; Bebin-Blackwell, A.-G.; Carter, D.M.; Alefantis, T.; di Napoli, J.; Kleanthous, H.; Ross, T.M.; Schultz-Cherry, S. Computationally optimized broadly reactive hemagglutinin elicits hemagglutination inhibition antibodies against a panel of H3N2 influenza virus cocirculating variants. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, N.S.; Anderson, T.K.; Kitikoon, P.; Skepner, E.; Burke, D.F.; Vincent, A.L. Substitutions near the hemagglutinin receptor-binding site determine the antigenic evolution of influenza A H3N2 viruses in U.S. swine. J. Virol. 2014, 88, 4752–4763. [Google Scholar] [CrossRef] [Green Version]
- Lam, J.H.; Baumgarth, N. The multifaceted B cell response to influenza virus. J. Immunol. 2019, 202, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.Y.; Ha do, L.A.; Simmons, C.; de Jong, M.D.; Chau, N.V.; Schumacher, R.; Peng, Y.C.; McMichael, A.J.; Farrar, J.J.; Smith, G.L.; et al. Memory T cells established by seasonal human influenza A infection cross-react with avian influenza A (H5N1) in healthy individuals. J. Clin. Investig. 2008, 118, 3478–3490. [Google Scholar] [CrossRef]
- Keynan, Y.; Card, C.M.; Ball, B.T.; Li, Y.; Plummer, F.A.; Fowke, K.R. Cellular immune responses to recurring influenza strains have limited boosting ability and limited cross-reactivity to other strains. Clin. Microbiol. Infect. 2010, 16, 1179–1186. [Google Scholar] [CrossRef]
- Ellebedy, A. Immunizing the immune: Can we overcome influenza’s most formidable challenge? Vaccines 2018, 6, 68. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.Z.; Martin, J.M.; Gross, F.L.; Jefferson, S.; Cole, K.S.; Archibald, C.A.; Nowalk, M.P.; Susick, M.; Moehling, K.; Spencer, S.; et al. Neutralizing antibody responses to antigenically drifted influenza A (H3N2) viruses among children and adolescents following 2014–2015 inactivated and live attenuated influenza vaccination. Clin. Vaccine Immunol. 2016, 23, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.F.; Huang, Y.; Kaur, K.; Popova, L.I.; Ho, I.Y.; Pauli, N.T.; Dunand, C.J.H.; Taylor, W.M.; Lim, S.; Huang, M.; et al. Immune history profoundly affects broadly protective B cell responses to influenza. Sci. Transl. Med. 2015, 7, ra192–ra316. [Google Scholar] [CrossRef] [Green Version]
- Abreu, R.B.; Kirchenbaum, G.A.; Clutter, E.F.; Sautto, G.A.; Ross, T.M. Preexisting subtype immunodominance shapes memory B cell recall response to influenza vaccination. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, C.S.; Gramer, M.R.; Barbé, F.; Van Doorsselaere, J.; Van Reeth, K. Efficacy of commercial swine influenza vaccines against challenge with a recent European H1N1 field isolate. Vet. Microbiol. 2010, 144, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Kohler, H. Novel vaccine concept based on back-boost effect in viral infection. Vaccine 2015, 33, 3274–3275. [Google Scholar] [CrossRef] [PubMed]
- Chepkwony, S.; Van Reeth, K. Hemagglutination Inhibition antibodies against novel North American swine influenza viruses, A/swine/Missouri/A01476459/2012 and A/swine/Missouri/A01840724/2015 after heterologous prime-boost vaccination with A/swine/Gent/172/2008, followed by A/swine/Pennsylvania/A01076777/2010. 2018; unpublished data. [Google Scholar]
- Francis, M.E.; McNeil, M.; Dawe, N.J.; Foley, M.K.; King, M.L.; Ross, T.M.; Kelvin, A.A. Historical H1N1 influenza virus imprinting increases vaccine protection by influencing the activity and sustained production of antibodies elicited at vaccination in ferrets. Vaccines 2019, 7, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, T.M.; Levine, M.; Fitzgerald, T.; Luke, C.; Sangster, M.Y.; Jin, H.; Topham, D.; Katz, J.; Treanor, J.; Subbarao, K. Live attenuated H7N7 influenza vaccine primes for a vigorous antibody response to inactivated H7N7 influenza vaccine. Vaccine 2014, 32, 6798–6804. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Krammer, F.; Albrecht, R.A. A live-attenuated prime, inactivated boost vaccination strategy with chimeric hemagglutinin-based universal influenza virus vaccines provides protection in ferrets: A confirmatory study. Vaccines 2018, 6, 47. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.D.; Jang, H.; di Napoli, J.; Kleanthous, H.; Ross, T.M. Elicitation of protective antibodies against 20 years of future H3N2 cocirculating influenza virus variants in ferrets preimmune to historical H3N2 influenza viruses. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Brandtzaeg, P. Induction of secretory immunity and memory at mucosal surfaces. Vaccine 2007, 25, 5467–5484. [Google Scholar] [CrossRef]
- Sandbulte, M.R.; Roth, J.A. Methods for analysis of cell-mediated immunity in domestic animal species. J. Am. Vet. Med. Assoc. 2004, 225, 522–530. [Google Scholar] [CrossRef]
- Vincent, A.L.; Ciacci-Zanella, J.R.; Lorusso, A.; Gauger, P.C.; Zanella, E.L.; Kehrli, M.E., Jr.; Janke, B.H.; Lager, K.M. Efficacy of inactivated swine influenza virus vaccines against the 2009 A/H1N1 influenza virus in pigs. Vaccine 2010, 28, 2782–2787. [Google Scholar] [CrossRef]
- Qiu, Y.; de Hert, K.; Van Reeth, K. Cross-protection against European swine influenza viruses in the context of infection immunity against the 2009 pandemic H1N1 virus: Studies in the pig model of influenza. Vet. Res. 2015, 46, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; van der Meulen, K.; Van Reeth, K. Prior infection of pigs with a recent human H3N2 influenza virus confers minimal cross-protection against a European swine H3N2 virus. Influenza Other Respir. Viruses 2013, 7, 1260–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, F.D.; Harwood, N.E. The who, how and where of antigen presentation to B cells. Nat. Rev. Immunol. 2009, 9, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Pape, K.A.; Catron, D.M.; Itano, A.A.; Jenkins, M.K. The humoral immune response is initiated in lymph nodes by B cells that acquire soluble antigen directly in the follicles. Immunity 2007, 26, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Van Riet, E.; Ainai, A.; Suzuki, T.; Hasegawa, H. Mucosal IgA responses in influenza virus infections; thoughts for vaccine design. Vaccine 2012, 30, 5893–5900. [Google Scholar] [CrossRef]
- Carter, M.J.; Mitchell, R.M.; Meyer Sauteur, P.M.; Kelly, D.F.; Trück, J. The antibody-secreting cell response to infection: Kinetics and clinical applications. Front. Immunol. 2017, 8, 630. [Google Scholar] [CrossRef]
- Jegaskanda, S.; Mason, R.D.; Andrews, S.F.; Wheatley, A.K.; Zhang, R.; Reynoso, G.V.; Ambrozak, D.R.; Santos, C.P.; Luke, C.J.; Matsuoka, Y.; et al. Intranasal live influenza vaccine priming elicits localized B cell responses in mediastinal lymph nodes. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percentage Amino Acid (aa) Identities (Lower Left) and Number of Different aa at Antigenic Sites, out of a Total of 40 (Upper Right) | ||||
|---|---|---|---|---|
| NC95 | G08 | PA10 | MO15 | |
| NC95 | - | 18 | 12 | 14 |
| G08 | 83.0 | - | 20 | 19 |
| PA10 | 91.5 | 81.5 | - | 16 |
| MO15 | 87.2 | 79.6 | 84.2 | - |
| Group | H3N2 Influenza Strains Used for: | |||
|---|---|---|---|---|
| Infection (Week 0) | Primary Vaccination (Week 7) | Booster Vaccination (Week 11) | Challenge (Week 17) | |
| Naïve challenge control | PBS | PBS | PBS | MO15 |
| Infection-immune challenge control | NC95 | PBS | PBS | |
| Infection-immune vaccinated (Homologous prime-boost) | NC95 | G08 | G08 | |
| NC95 | PA10 | PA10 | ||
| Infection-immune vaccinated (Heterologous prime-boost) | NC95 | G08 | PA10 | |
| NC95 | PA10 | G08 | ||
| Geometric Mean HI Antibody Titers Per Group at: (Number of Pigs with Titer ≥40) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Week 9 | Week 13 | ||||||||||
| Virus Strains | Inf.: 1°: 2°: | NC95 PBS PBS | NC95 G08 G08 | NC95 PA10 PA10 | NC95 G08 PA10 | NC95 PA10 G08 | NC95 PBS PBS | NC95 G08 G08 | NC95 PA10 PA10 | NC95 G08 PA10 | NC95 PA10 G08 |
| EU swine | |||||||||||
| G84 | 5 (0) | 160 (5) | 53 (3) | 184 (5) | 46 (4) | 5 (0) | 1470 (5) | 160 (4) | 279 (5) | 368 (5) | |
| ENG87 | 5 (0) | 15 (0) | 10 (0) | 17 (1) | 9 (0) | 5 (0) | 160 (5) | 30 (3) | 46 (5) | 23 (2) | |
| G08 | 5 (0) | 243 (5) | 139 (5) | 279 (5) | 80 (5) | 5 (0) | 2941 (5) | 485 (5) | 970 (5) | 1114 (5) | |
| N.A swine | |||||||||||
| TX98 | 13 (0) | 844 (5) | 640 (5) | 422 (5) | 1689 (5) | 17 (1) | 368 (5) | 1470 (5) | 243 (5) | 1940 (5) | |
| MN99 | 10 (0) | 485 (5) | 485 (5) | 368 (5) | 844 (5) | 10 (0) | 211 (5) | 735 (5) | 121 (5) | 1280 (5) | |
| ONT05 | 6 (0) | 106 (5) | 243 (5) | 80 (5) | 485 (5) | 5 (0) | 40 (3) | 845 (5) | 46 (3) | 485 (5) | |
| PA10 | 5 (0) | 121 (5) | 485 (5) | 70 (5) | 1114 (5) | 5 (0) | 80 (4) | 4457 (5) | 320 (5) | 1470 (5) | |
| IN11 | 5 (0) | 80 (4) | 184 (5) | 61 (4) | 279 (5) | 5 (0) | 121 (5) | 3880 (5) | 243 (4) | 1940 (5) | |
| IA11 | 5 (0) | 368 (5) | 640 (5) | 368 (5) | 1470 (5) | 6 (0) | 243 (5) | 3378 (5) | 422 (5) | 1689 (5) | |
| MO12 | 5 (0) | 121 (5) | 160 (5) | 160 (5) | 368 (5) | 5 (0) | 121 (5) | 211 (5) | 160 (5) | 279 (5) | |
| MO15 | 5 (0) | 160 (5) | 320 (5) | 243 (5) | 485 (5) | 5 (0) | 106 (5) | 640 (5) | 139 (5) | 422 (5) | |
| Human | |||||||||||
| VIC75 | 5 (0) | 23 (2) | 13 (1) | 20 (1) | 20 (1) | 5 (0) | 121 (5) | 35 (4) | 48 (3) | 70 (3) | |
| ENG88 | 5 (0) | 243 (5) | 139 (5) | 160 (5) | 320 (5) | 5 (0) | 106 (5) | 184 (5) | 160 (5) | 160 (5) | |
| NC95 | 80 (5) | 1689 (5) | 2560 (5) | 2229 (5) | 4457 (5) | 70 (5) | 1114 (5) | 5120 (5) | 2560 (5) | 5881 (5) | |
| WI05 | 6 (0) | 9 (0) | 6 (0) | 6 (0) | 7 (0) | 5 (0) | 5 (0) | 10 (1) | 5 (0) | 9 (0) | |
| PER09 | 6 (0) | 8 (0) | 11 (1) | 8 (0) | 10 (1) | 5 (0) | 7 (0) | 23 (2) | 7 (0) | 10 (1) | |
| VIC11 | 5 (0) | 5 (0) | 5 (0) | 5 (0) | 5 (0) | 5 (0) | 6 (0) | 7 (0) | 6 (0) | 12 (1) | |
| HK14 | 5 (0) | 6 (0) | 7 (0) | 5 (0) | 6 (0) | 5 (0) | 5 (0) | 35 (3) | 5 (0) | 7 (0) | |
| No. of strains (n/18) against which a titer ≥40 was achieved per group | 1 | 12 | 12 | 12 | 12 | 1 | 14 | 12 | 14 | 13 | |
| Geomean NI Titers Per Group at: | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Week 9 | Week 13 | ||||||||||
| Virus Strains | Inf.: 1°: 2°: | NC95 PBS PBS | NC95 G08 G08 | NC95 PA10 PA10 | NC95 G08 PA10 | NC95 PA10 G08 | NC95 PBS PBS | NC95 G08 G08 | NC95 PA10 PA10 | NC95 G08 PA10 | NC95 PA10 G08 |
| EU swine | |||||||||||
| G08 | 7 | 557 | 320 | 844 | 485 | 15 | 5881 | 1689 | 1940 | 2941 | |
| G12 * | 5 | 557 | 121 | 106 | 184 | 10 | 139 | 735 | 184 | 368 | |
| N.A swine | |||||||||||
| TX98 | 23 | 485 | 844 | 422 | 1470 | 23 | 279 | 1470 | 368 | 1114 | |
| PA10 | 7 | 640 | 2560 | 485 | 4457 | 11 | 485 | 17,829 | 1470 | 10,240 | |
| IN11 | 8 | 640 | 1689 | 640 | 2229 | 35 | 1689 | 35,658 | 3880 | 23,525 | |
| IA11 | 26 | 114 | 2229 | 1280 | 4457 | 23 | 557 | 5881 | 970 | 4457 | |
| MO15 | 8 | 243 | 485 | 243 | 735 | 9 | 211 | 1114 | 320 | 320 | |
| Human | |||||||||||
| NC95 | 139 | 2941 | 3378 | 2941 | 6756 | 106 | 1689 | 5120 | 2560 | 6756 | |
| PER09 | 5 | 11 | 10 | 6 | 30 | 5 | 13 | 80 | 15 | 40 | |
| VIC11 | 8 | 40 | 46 | 23 | 80 | 7 | 20 | 70 | 35 | 121 | |
| Vaccine Group | Geomean HI Antibody Titers | Mean Lung Virus Titers (log10 TCID50/g) |
|---|---|---|
| PBS-PBS-PBS | 5.0 | 6.14 |
| NC95-PBS-PBS | 5.0 | 3.44 |
| NC95-G08-G08 | 26.4 | 1.25 |
| NC95-PA10-PA10 | 242.5 | 0.85 |
| NC95-G08-PA10 | 40.0 | 1.82 |
| NC95-PA10-G08 | 160.0 | 0.85 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chepkwony, S.; Parys, A.; Vandoorn, E.; Chiers, K.; Van Reeth, K. Efficacy of Heterologous Prime-Boost Vaccination with H3N2 Influenza Viruses in Pre-Immune Individuals: Studies in the Pig Model. Viruses 2020, 12, 968. https://0-doi-org.brum.beds.ac.uk/10.3390/v12090968
Chepkwony S, Parys A, Vandoorn E, Chiers K, Van Reeth K. Efficacy of Heterologous Prime-Boost Vaccination with H3N2 Influenza Viruses in Pre-Immune Individuals: Studies in the Pig Model. Viruses. 2020; 12(9):968. https://0-doi-org.brum.beds.ac.uk/10.3390/v12090968
Chicago/Turabian StyleChepkwony, Sharon, Anna Parys, Elien Vandoorn, Koen Chiers, and Kristien Van Reeth. 2020. "Efficacy of Heterologous Prime-Boost Vaccination with H3N2 Influenza Viruses in Pre-Immune Individuals: Studies in the Pig Model" Viruses 12, no. 9: 968. https://0-doi-org.brum.beds.ac.uk/10.3390/v12090968