Meta-Transcriptomic Discovery of a Divergent Circovirus and a Chaphamaparvovirus in Captive Reptiles with Proliferative Respiratory Syndrome

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. PCR Testing for Pneumotropic Viruses
2.3. Pathogen Discovery Using Meta-transcriptomics
2.4. PCR Testing for Chaphamaparvoviruses and Circoviruses
2.5. Phylogenetic Analysis
2.6. Mining the Sequence Read Archive (SRA)
3. Results
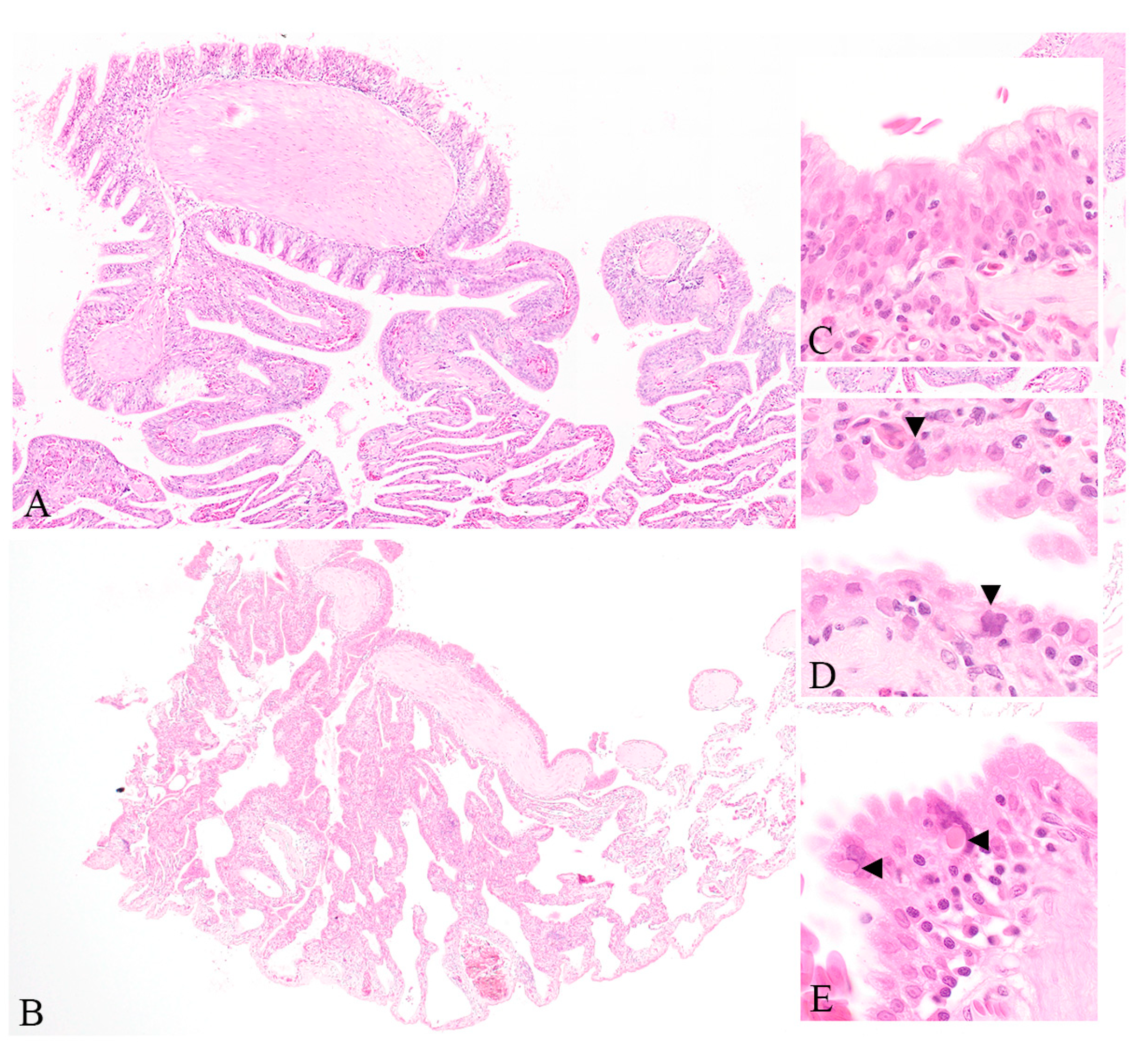
3.1. Clinical and Histopathological Findings of Diseased Captive Reptiles
3.2. PCR Testing for Pneumotropic Viruses
3.3. Metatranscriptomic Pathogen Discovery
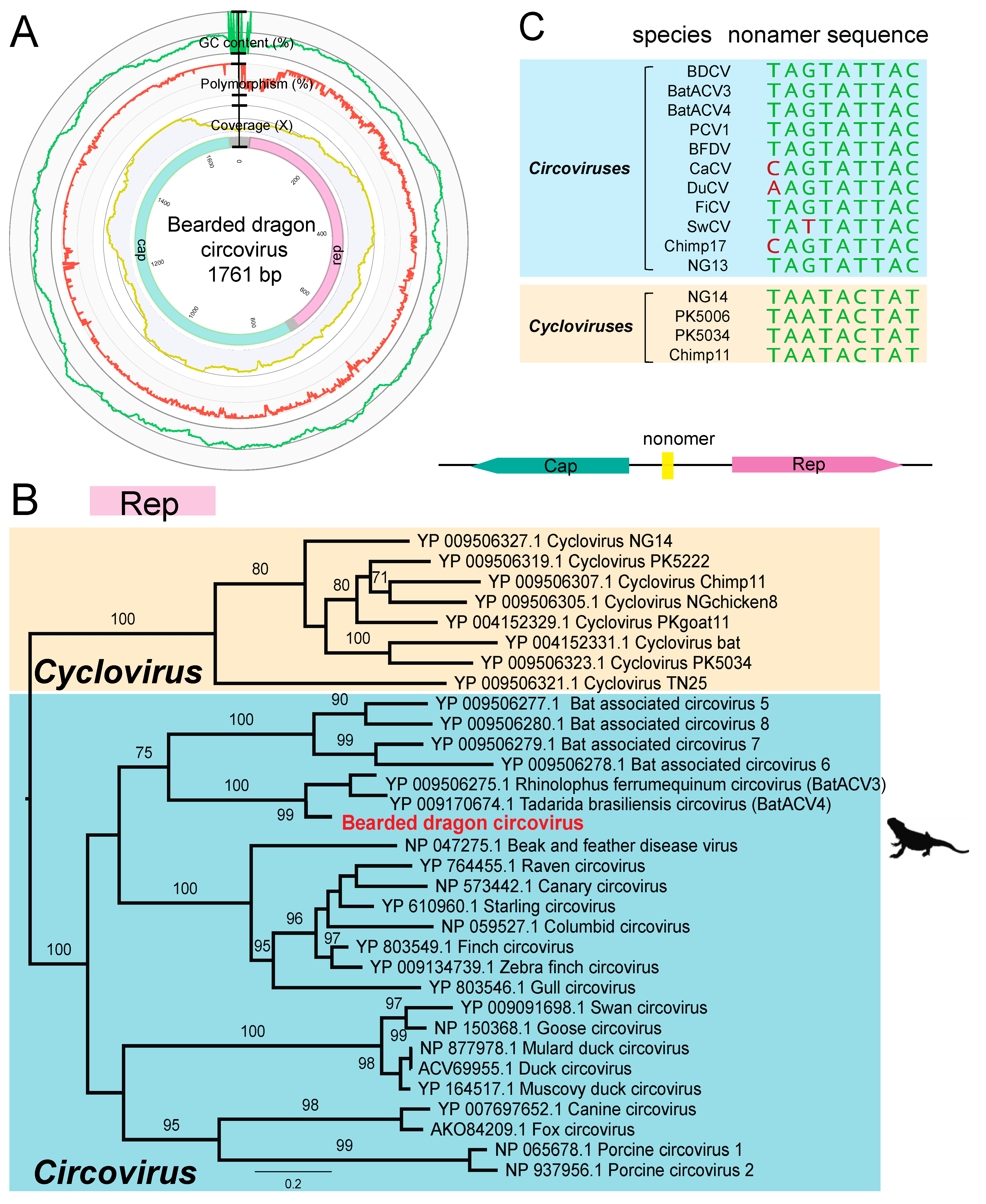
3.4. Genome Characterization of a Novel Bearded Dragon Circovirus
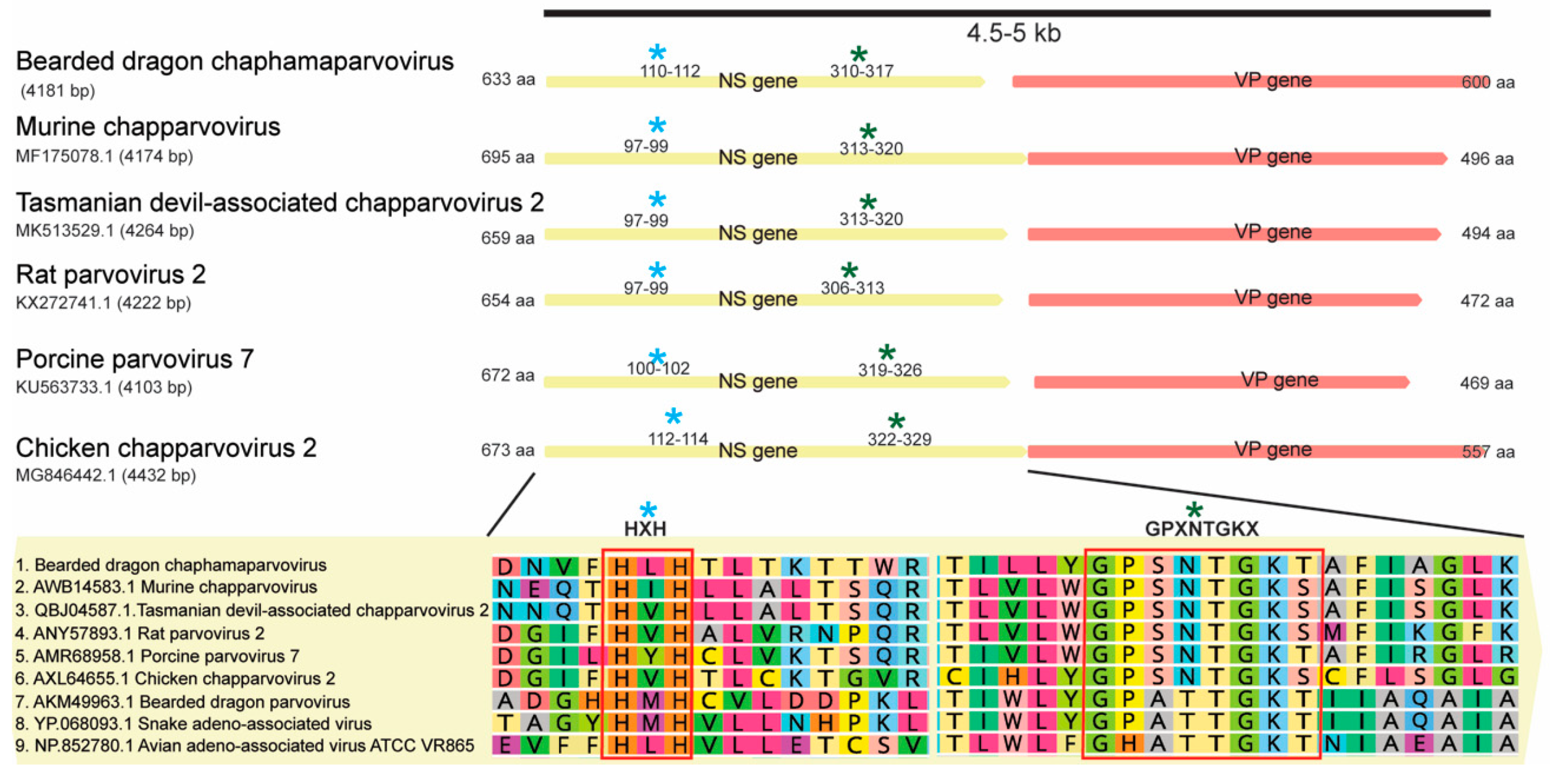
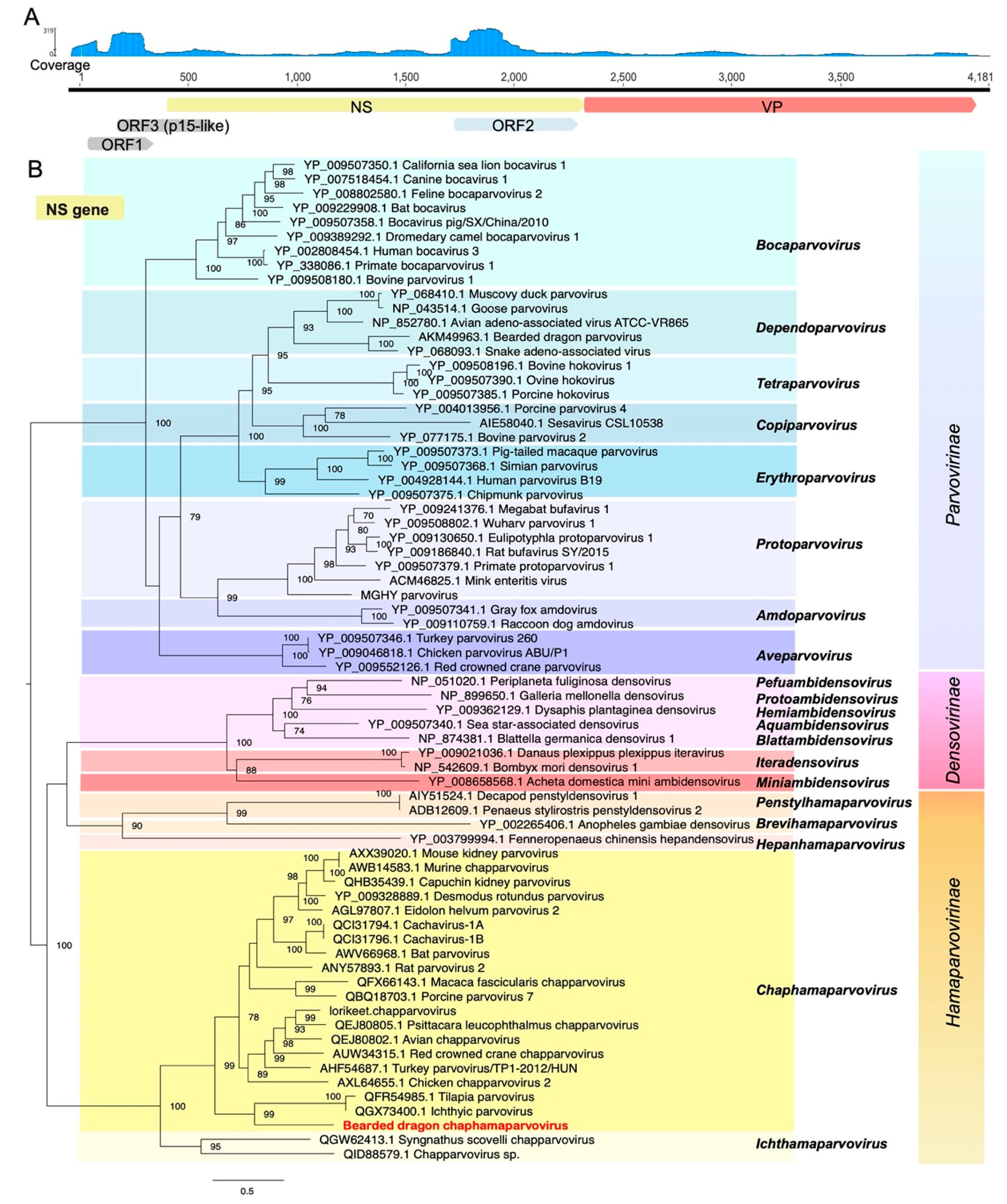
3.5. Identification of a Highly Divergent Bearded Dragon Chaphamaparvovirus
3.6. Prevalence of BDCV and BDchPV through PCR Screening
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tingley, R.; Macdonald, S.L.; Mitchell, N.J.; Woinarski, J.C.Z.; Meiri, S.; Bowles, P.; Cox, N.A.; Shea, G.M.; Böhm, M.; Chanson, J.; et al. Geographic and taxonomic patterns of extinction risk in Australian squamates. Biol. Conserv. 2019, 238, 108203. [Google Scholar] [CrossRef]
- Cogger, H.G. Reptiles and Amphibians of Australia, 7th ed.; CSIRO Publishing: Collingwood, VIC, Australia, 2014; 1033p. [Google Scholar]
- Deveson, I.W.; Holleley, C.E.; Blackburn, J.; Marshall Graves, J.A.; Mattick, J.S.; Waters, P.D.; Georges, A. Differential intron retention in Jumonji chromatin modifier genes is implicated in reptile temperature-dependent sex determination. Sci. Adv. 2017, 3, e1700731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holleley, C.E.; O’Meally, D.; Sarre, S.D.; Marshall Graves, J.A.; Ezaz, T.; Matsubara, K.; Azad, B.; Zhang, X.; Georges, A. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 2015, 523, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Stöhr, A.C.; Blahak, S.; Heckers, K.O.; Wiechert, J.; Behncke, H.; Mathes, K.A.; Günther, P.; Zwart, P.; Ball, I.; Rüschoff, B.; et al. Ranavirus infections associated with skin lesions in lizards. Vet. Res. 2013, 44, 84. [Google Scholar] [CrossRef] [Green Version]
- Tamukai, K.; Tokiwa, T.; Kobayashi, H.; Une, Y. Ranavirus in an outbreak of dermatophilosis in captive inland bearded dragons (Pogona vitticeps). Vet. Dermatol. 2016, 27, 99-e28. [Google Scholar] [CrossRef]
- Grosset, C.; Wellehan, J.F., Jr.; Owens, S.D.; McGraw, S.; Gaffney, P.M.; Foley, J.; Childress, A.L.; Yun, S.; Malm, K.; Groff, J.M.; et al. Intraerythrocytic iridovirus in central bearded dragons (Pogona vitticeps). J. Vet. Diagn. Investig. 2014, 26, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Papp, T.; Marschang, R.E. Detection and characterization of invertebrate iridoviruses found in reptiles and prey insects in Europe over the past two decades. Viruses 2019, 11, 600. [Google Scholar] [CrossRef] [Green Version]
- Benge, S.L.; Hyndman, T.H.; Funk, R.S.; Marschang, R.E.; Schneider, R.; Childress, A.L.; Wellehan, J.F. Identification of helodermatid adenovirus 2 in a captive central bearded dragon (Pogona Vitticeps), wild gila monsters (Heloderma Suspectum), and a death adder (Acanthophis Antarcticus). J. Zoo Wildl. Med. 2019, 50, 238–242. [Google Scholar]
- Infectious Diseases and Pathology of Reptiles: Color Atlas and Text, 2nd ed.; Jacobson, E.R.; Garner Michael, M. (Eds.) CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2020; 1030p. [Google Scholar]
- Pénzes, J.J.; Pham, H.T.; Benkő, M.; Tijssen, P.; Benk, M. Novel parvoviruses in reptiles and genome sequence of a lizard parvovirus shed light on Dependoparvovirus genus evolution. J. Gen. Virol. 2015, 96, 2769–2779. [Google Scholar] [CrossRef]
- Kim, D.Y.; Mitchell, M.A.; Bauer, R.W.; Poston, R.; Cho, D.Y. An outbreak of adenoviral infection in inland bearded dragons (Pogona vitticeps) coinfected with dependovirus and coccidial protozoa (Isospora sp.). J. Vet. Diagn. Investig. 2002, 14, 332–334. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, E.R.; Kopit, W.; Kennedy, F.A.; Funk, R.S. Coinfection of a bearded dragon, Pogona vitticeps, with adenovirus- and dependovirus-like Viruses. Vet. Pathol. 1996, 33, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Crossland, N.A.; DiGeronimo, P.M.; Sokolova, Y.; Childress, A.L.; Wellehan, J.F.X.; Nevarez, J.; Paulsen, D. Pneumonia in a captive central bearded dragon with concurrent detection of helodermatid adenovirus 2 and a novel mycoplasma species. Vet. Pathol. 2018, 55, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Abbas, M.D.; Ball, I.; Ruckova, Z.; Öfner, S.; Stöhr, A.C.; Marschang, R.E. Virological screening of bearded dragons (Pogona vitticeps) and the first detection of paramyxoviruses in this species. J. Herpetol. Med. Surg. 2012, 22, 86–90. [Google Scholar] [CrossRef]
- Capraro, A.; O’Meally, D.; Waters, S.A.; Patel, H.R.; Georges, A.; Waters, P.D. Waking the sleeping dragon: Gene expression profiling reveals adaptive strategies of the hibernating reptile Pogona vitticeps. BMC Genom. 2019, 20, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, K.K.; Aiello, C.M.; Bowen, L.; Lewison, R.L.; Esque, T.C.; Nussear, K.E.; Waters, S.C.; Hudson, P.J. Complex immune responses and molecular reactions to pathogens and disease in a desert reptile (Gopherus agassizii). Ecol. Evol. 2019, 9, 2516–2534. [Google Scholar] [CrossRef] [Green Version]
- Hyndman, T.H.; Johnson, R.S. Evidence for the vertical transmission of Sunshine virus. Vet. Microbiol. 2015, 175, 179–184. [Google Scholar] [CrossRef]
- Wellehan, J.F., Jr.; Childress, A.L.; Marschang, R.E.; Johnson, A.J.; Lamirande, E.W.; Roberts, J.F.; Vickers, M.L.; Gaskin, J.M.; Jacobson, E.R. Consensus nested PCR amplification and sequencing of diverse reptilian, avian, and mammalian orthoreoviruses. Vet. Microbiol. 2009, 133, 34–42. [Google Scholar] [CrossRef]
- Ahne, W.; Batts, W.N.; Kurath, G.; Winton, J.R. Comparative sequence analyses of sixteen reptilian paramyxoviruses. Virus Res. 1999, 63, 65–74. [Google Scholar] [CrossRef]
- Hyndman, T.H.; Marschang, R.E.; Wellehan, J.F.X., Jr.; Nicholls, P.K. Isolation and molecular identification of Sunshine virus, a novel paramyxovirus found in Australian snakes. Infect. Genet. Evol. 2012, 12, 1436–1446. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Lin, X.-D.; Chen, X.; Tian, J.-H.; Chen, L.-J.; Li, K.; Wang, W.; Eden, J.-S.; Shen, J.-J.; Liu, L.; et al. The evolutionary history of vertebrate RNA viruses. Nature 2018, 556, 197–202. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.-D.; Vasilakis, N.; Tian, J.-H.; Li, C.-X.; Chen, L.-J.; Eastwood, G.; Diao, X.-N.; Chen, M.-H.; Chen, X.; et al. Divergent viruses discovered in arthropods and vertebrates revise the evolutionary history of the Flaviviridae and related viruses. J. Virol. 2015, 90, 659–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protocol. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Mount, D.W. Using the Basic Local Alignment Search Tool (BLAST). CSH Protoc. 2007. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, B. BBMap Short-Read Aligner, And Other Bioinformatics Tools. 2016. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 28 May 2020).
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Fahsbender, E.; Altan, E.; Seguin, M.A.; Young, P.; Estrada, M.; Leutenegger, C.M.; Delwart, E. Chapparvovirus DNA found in 4% of dogs with diarrhea. Viruses 2019, 11, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Gordon, E.; Idle, A.; Altan, E.; Seguin, M.A.; Estrada, M.; Deng, X.; Delwart, E. Virome of a feline outbreak of diarrhea and vomiting includes bocaviruses and a novel chapparvovirus. Viruses 2020, 12, 506. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, Z.; Wang, Y.; Li, W.; Fu, X.; Lin, Y.; Shen, Q.; Wang, X.; Wang, H.; Zhang, W. A novel rodent Chapparvovirus in feces of wild rats. Virol. J. 2016, 13, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Wang, W.; Chan, J.F.; Wang, G.; Huang, Y.; Yi, Y.; Zhu, Z.; Peng, R.; Hu, X.; Wu, Y.; et al. Identification of a novel ichthyic parvovirus in marine species in Hainan Island, China. Front. Microbiol. 2019, 10, 2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Q.; Padula, M.P.; Pinello, N.; Williams, S.H.; O’Rourke, M.B.; Fumagalli, M.J.; Orkin, J.D.; Song, R.; Shaban, B.; Brenner, O.; et al. Murine and related chapparvoviruses are nephro-tropic and produce novel accessory proteins in infected kidneys. PLoS Pathog. 2020, 16, e1008262. [Google Scholar] [CrossRef] [Green Version]
- Souza, W.M.; Romeiro, M.F.; Fumagalli, M.J.; Modha, S.; de Araujo, J.; Queiroz, L.H.; Durigon, E.L.; Figueiredo, L.T.M.; Murcia, P.R.; Gifford, R.J. Chapparvoviruses occur in at least three vertebrate classes and have a broad biogeographic distribution. J. Gen. Virol. 2017, 98, 225–229. [Google Scholar] [CrossRef]
- Kapoor, A.; Dubovi, E.J.; Henriquez-Rivera, J.A.; Lipkin, W.I. Complete genome sequence of the first canine circovirus. J. Virol. 2012, 86, 7018. [Google Scholar] [CrossRef] [Green Version]
- Raidal, S.R.; Sarker, S.; Peters, A. Review of psittacine beak and feather disease and its effect on Australian endangered species. Aust. Vet. J. 2015, 93, 466–470. [Google Scholar] [CrossRef]
- Allan, G.; Krakowka, S.; Ellis, J.; Charreyre, C. Discovery and evolving history of two genetically related but phenotypically different viruses, porcine circoviruses 1 and 2. Virus Res. 2012, 164, 4–9. [Google Scholar] [CrossRef]
- Lian, H.; Liu, Y.; Li, N.; Wang, Y.; Zhang, S.; Hu, R. Novel circovirus from mink, China. Emerg. Infect. Dis. 2014, 20, 1548–1550. [Google Scholar] [CrossRef]
- Altan, E.; Kubiski, S.V.; Burchell, J.; Bicknese, E.; Deng, X.; Delwart, E. The first reptilian circovirus identified infects gut and liver tissues of black-headed pythons. Vet. Res. 2019, 50, 35. [Google Scholar] [CrossRef] [Green Version]
- Tarján, Z.L.; Pénzes, J.J.; Tóth, R.P.; Benkő, M. First detection of circovirus-like sequences in amphibians and novel putative circoviruses in fishes. Acta Vet. Hung. 2014, 62, 134–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marton, S.; Ihász, K.; Lengyel, G.; Farkas, S.L.; Dán, Á.; Paulus, P.; Banyai, K.; Fehér, E. Ubiquiter circovirus sequences raise challenges in laboratory diagnosis: The case of honey bee and bee mite, reptiles, and free living amoebae. Acta Microbiol. Immunol. Hung. 2015, 62, 57–73. [Google Scholar] [PubMed] [Green Version]
- Gilbert, C.; Meik, J.M.; Dashevsky, D.; Card, D.C.; Castoe, T.A.; Schaack, S. Endogenous hepadnaviruses, bornaviruses and circoviruses in snakes. Proc. Biol. Sci. 2014, 281, 20141122. [Google Scholar] [CrossRef] [PubMed]
- Dunham, S.; Daly, J.M. Canine parvovirus study. Vet Rec 2016, 179, 51. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Xie, Z.; Deng, X.; Xie, L.; Xie, Z.; Huang, L.; Fan, Q.; Luo, S.; Huang, J.; Zhang, Y.; et al. Genetic and phylogenetic analysis of a novel parvovirus isolated from chickens in Guangxi, China. Arch. Virol. 2016, 161, 3285–3289. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, P.; Wu, Y.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Yang, Q.; Wu, Y.; Zhao, X.-X.; et al. Growth characteristics of the novel goose parvovirus SD15 strain in vitro. BMC Vet. Res. 2019, 15, 63. [Google Scholar] [CrossRef]
- Palinski, R.M.; Mitra, N.; Hause, B.M. Discovery of a novel Parvovirinae virus, porcine parvovirus 7, by metagenomic sequencing of porcine rectal swabs. Virus Genes 2016, 52, 564–567. [Google Scholar] [CrossRef]
- Miłek, D.; Woźniak, A.; Stadejek, T. The detection and genetic diversity of novel porcine parvovirus 7 (PPV7) on Polish pig farms. Res. Vet. Sci. 2018, 120, 28–32. [Google Scholar] [CrossRef]
- Fahsbender, E.; Charlys da-Costa, A.; Elise Gill, D.; Augusto de Padua Milagres, F.; Brustulin, R.; Julio Costa Monteiro, F.; Octavio da Silva Rego, M.; Soares D’Athaide Ribeiro, E.; Cerdeira Sabino, E.; Delwart, E. Plasma virome of 781 Brazilians with unexplained symptoms of arbovirus infection include a novel parvovirus and densovirus. PLoS ONE 2020, 15, e0229993. [Google Scholar] [CrossRef] [Green Version]
- Farkas, S.L.; Zádori, Z.; Benkő, M.; Essbauer, S.; Harrach, B.; Tijssen, P. A parvovirus isolated from royal python (Python regius) is a member of the genus Dependovirus. J. Gen. Virol. 2004, 85, 555–561. [Google Scholar] [CrossRef]
- Wozniak, E.J.; DeNardo, D.F.; Brewer, A.; Wong, V.; Tarara, R.P. Identification of adenovirus- and dependovirus-like agents in an outbreak of fatal gastroenteritis in captive born California mountain kingsnakes, Lampropeltis zonata multicincta. J. Herpetol. Med. Surg. 2000, 10, 4–7. [Google Scholar] [CrossRef]
- Ahne, W.; Scheinert, P. Reptilian viruses: Isolation of parvovirus-like particles from corn snake Elapha guttata (Colubridae). J. Vet. Med. 1989, 36, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, E.R.; Gaskin, J.M.; Gardiner, C.H. Adenovirus-like infection in a boa constrictor. J. Am. Vet. Med. Assoc. 1985, 187, 1226–1227. [Google Scholar] [PubMed]
- Farkas, S.G.J. First Hungarian report of inclusion body hepatitis associated with adenoviruses and secondary parvovirus infection in an Indonesian pit-viper [Parias (Trimeresurus) hageni]. Magy. Állatorvosok Lapja 2008, 130, 755–761. [Google Scholar]
- Pénzes, J.J.; Benkő, M. Novel parvovirus from the worm lizard Trogonophis wiegmanni—First virus ever detected in amphisbaenian hosts. Acta Vet. Hung. 2014, 62, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Pénzes, J.J.; Söderlund-Venermo, M.; Canuti, M.; Eis-Hübinger, A.M.; Hughes, J.; Cotmore, S.F.; Harrach, B. Reorganizing the family Parvoviridae: A revised taxonomy independent of the canonical approach based on host association. Arch. Virol. 2020, 165, 2133–2146. [Google Scholar] [CrossRef]
- Pénzes, J.J.; de Souza, W.M.; Agbandje-McKenna, M.; Gifford, R.J. An ancient lineage of highly divergent parvoviruses infects both vertebrate and invertebrate hosts. Viruses 2019, 11, 525. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.A.; Bratanich, A.; Clark, E.G.; Allan, G.; Meehan, B.; Haines, D.M.; Harding, J.; West, K.H.; Krakowka, S.; Konoby, C.; et al. Coinfection by porcine circoviruses and porcine parvovirus in pigs with naturally acquired postweaning multisystemic wasting syndrome. J. Vet. Diagn. Invest. 2000, 12, 21–27. [Google Scholar] [CrossRef]
- Thaiwong, T.; Wise, A.G.; Maes, R.K.; Mullaney, T.; Kiupel, M. Canine circovirus 1 (CaCV-1) and canine parvovirus 2 (CPV-2): Recurrent dual infections in a papillon breeding colony. Vet. Pathol. 2016, 53, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.-S.; Eden, J.-S.; Hall, J.; Shi, M.; Rose, K.; Holmes, E.C. Meta-transcriptomic analysis of virus diversity in urban wild birds with paretic disease. J. Virol. 2020, 94, e00606-20. [Google Scholar] [CrossRef]
- Chong, R.; Shi, M.; Grueber, C.E.; Holmes, E.C.; Hogg, C.J.; Belov, K.; Barrs, V.R. Fecal viral diversity of captive and wild Tasmanian devils characterized using virion-enriched metagenomics and metatranscriptomics. J. Virol. 2019, 93, e00205-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Target | Sequence 5′-3′ | Site Start | Site End | Tm (°C) | GC (%) |
|---|---|---|---|---|---|---|
| BDCV_F1 | BDCV_cap | GTATTGTTTCAGAGATTCGCCG | 65 | 86 | 57.8 | 45.5 |
| BDCV_R1 | ATTGTAATGAAGGCGTTGTTCC | 1144 | 1165 | 57.6 | 40.9 | |
| BDCV_F2 | BDCV_cap | GCCGAAGTGTCTGGTAATACTA | 974 | 995 | 57.3 | 43.5 |
| BDCV_R2 | GATTGGTTAAGAGTCGGGATTTC | 440 | 462 | 57.6 | 40.9 | |
| BDCV_F3 | BDCV_rep | AATACCTGACCAGTCTGCCA | 1551 | 1570 | 57.3 | 50.0 |
| BDCV_R3 | CGATGGCGTAGGAACAGATG | 659 | 640 | 59.3 | 55.0 | |
| BDCV_F4 | BDCV_rep | GGTGAAGTCGATTGTTCCTTTG | 312 | 333 | 58.4 | 45.5 |
| BDCV_R4 | GTGGGTATTGTTTCAGAGATTCG | 1633 | 1611 | 58.9 | 43.5 | |
| BDchPV_F1 | BDchPV_NS | CTGAACAATATCTCTTCGTTGGAG | 217 | 240 | 57.4 | 41.7 |
| BDchPV_R1 | TCCCAGACACCCATATAACAATC | 1490 | 1512 | 57.7 | 43.5 | |
| BDchPV_F2 | BDchPV_NS | GCTCTCCTGAATATTGTGATATGTC | 954 | 978 | 57.4 | 40.0 |
| BDchPV_R2 | TTCCCTACCCACTTCTTTCTG | 2255 | 2275 | 56.9 | 47.6 | |
| BDchPV_F3 | BDchPV_VP | CTAGCTCAGACATCAGATCCAG | 2004 | 2025 | 57.5 | 50.0 |
| BDchPV_R3 | GTCGGTGGAGGATTGTCTATAG | 3346 | 3367 | 57.5 | 50.0 | |
| BDchPV_F4 | BDchPV_VP | AGCTTTTCCCAAATGACACTG | 2848 | 2868 | 57.0 | 42.9 |
| BDchPV_R4 | CAGAAGACACAACAGTAGGTAAAG | 4087 | 4110 | 57.2 | 41.7 |
| Name of Region | Sequence in the Circoviridae | Sequence in Bearded Dragon Circovirus | |
|---|---|---|---|
| RCR motif | RCR motif I | FT(L/I)NN | FTINN |
| RCR motif II | PHLQG | PHIQG | |
| RCR motif III | YC(S/x)K | YCSK | |
| SF3 helicase motif | Walker A | G(P/x)(P/x) GxGK(S/t) | GEPGSGKS |
| Walker B | uuDDF | ILDDF | |
| Walker C | uTSN | ITSN |
| Year | Case no. | Species | Clinical Sign | Organ | PCR Results | |
|---|---|---|---|---|---|---|
| BDCV | BDchPV | |||||
| 2014 | 10043.1 | CBD | Respiratory | Kidney | - | - |
| Lung | - | - | ||||
| 10043.2 | CBD | Respiratory | Liver | - | - | |
| 10043.3 | CBD | Respiratory | Kidney | + | - | |
| Lung | + | + | ||||
| 10043.4 | CBD | Respiratory | Liver | - | + | |
| 10043.5 | CBD | Respiratory | Liver | + | - | |
| 2020 | 13270.1 | CBD | Mild neurological | Brain | - | - |
| Liver | - | + | ||||
| 13270.2 | CBD | Normal | Brain | - | - | |
| Liver | - | - | ||||
| 13270.3 | CBD | Mild nonspecific illness | Brain | - | - | |
| Liver | + | + | ||||
| 13270.4 | CBD | Neurological | Brain | - | + | |
| Liver | + | + | ||||
| 13270.5 | CBD | Normal | Brain | - | - | |
| Liver | - | - | ||||
| 13270.6 | CBD | Neurological | Brain | n.a. | n.a. | |
| Liver | - | - | ||||
| 13270.7 | CBD | Neurological | Brain | n.a. | n.a. | |
| Liver | - | - | ||||
| 13270.8 | CBD | Neurological | Brain | n.a. | n.a. | |
| Liver | - | - | ||||
| 13270.9 | JL | Normal | Brain | - | - | |
| Liver | + | - | ||||
| 13270.10 | JL | Neurological | Brain | - | - | |
| Liver | - | - | ||||
| 13270.11 | JL | Neurological | Brain | - | - | |
| Liver | - | - | ||||
| 13270.12 | GED | Moribund | Brain | - | - | |
| Liver | - | - | ||||
| Total: BDCV positive cases: 5/17; BDchPV positive cases: 5/17 | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-S.; Li, C.-X.; Hall, J.; Eden, J.-S.; Hyndman, T.H.; Holmes, E.C.; Rose, K. Meta-Transcriptomic Discovery of a Divergent Circovirus and a Chaphamaparvovirus in Captive Reptiles with Proliferative Respiratory Syndrome. Viruses 2020, 12, 1073. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101073
Chang W-S, Li C-X, Hall J, Eden J-S, Hyndman TH, Holmes EC, Rose K. Meta-Transcriptomic Discovery of a Divergent Circovirus and a Chaphamaparvovirus in Captive Reptiles with Proliferative Respiratory Syndrome. Viruses. 2020; 12(10):1073. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101073
Chicago/Turabian StyleChang, Wei-Shan, Ci-Xiu Li, Jane Hall, John-Sebastian Eden, Timothy H. Hyndman, Edward C. Holmes, and Karrie Rose. 2020. "Meta-Transcriptomic Discovery of a Divergent Circovirus and a Chaphamaparvovirus in Captive Reptiles with Proliferative Respiratory Syndrome" Viruses 12, no. 10: 1073. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101073