Cytokine-Targeted Therapeutics for KSHV-Associated Disease


Department of Biomedical and Pharmaceutical Sciences, Chapman University School of Pharmacy, Irvine, CA 92618, USA
*
Author to whom correspondence should be addressed.
Viruses 2020, 12(10), 1097; https://0-doi-org.brum.beds.ac.uk/10.3390/v12101097
Submission received: 1 September 2020
/
Revised: 23 September 2020
/
Accepted: 25 September 2020
/
Published: 28 September 2020
(This article belongs to the Special Issue New Advances in Kaposi's Sarcoma-Associated Herpesvirus Research)
Abstract
:Kaposi’s sarcoma-associated herpesvirus (KSHV) also known as human herpesvirus 8 (HHV-8), is linked to several human malignancies including Kaposi sarcoma (KS), primary effusion lymphoma (PEL), multicentric Castleman’s disease (MCD) and recently KSHV inflammatory cytokine syndrome (KICS). As with other diseases that have a significant inflammatory component, current therapy for KSHV-associated disease is associated with significant off-target effects. However, recent advances in our understanding of the pathogenesis of KSHV have produced new insight into the use of cytokines as potential therapeutic targets. Better understanding of the role of cytokines during KSHV infection and tumorigenesis may lead to new preventive or therapeutic strategies to limit KSHV spread and improve clinical outcomes. The cytokines that appear to be promising candidates as KSHV antiviral therapies include interleukins 6, 10, and 12 as well as interferons and tumor necrosis factor-family cytokines. This review explores our current understanding of the roles that cytokines play in promoting KSHV infection and tumorigenesis, and summarizes the current use of cytokines as therapeutic targets in KSHV-associated diseases.
1. Introduction
In 1994, Kaposi’s sarcoma-associated herpesvirus (KSHV) was first identified in a Kaposi’s sarcoma (KS) lesion by Chang and Moore [1]. KS is a highly proliferative tumor derived from lymphatic endothelial cells [2]. KSHV infection is also linked to two B cell lymphoproliferative disorders—primary effusion lymphoma (PEL) and multicentric Castleman’s disease (MCD) [3,4]. In addition, recent studies have demonstrated a third KSHV-associated disease. KSHV-associated inflammatory cytokine syndrome (KICS) is characterized by high levels of viral interleukin-6 (vIL6) or human IL-6 (hIL-6) [5]. Cytokine production has been shown to contribute to KS pathogenesis [6,7,8]. In fact, high levels of IL-6, TNF-α and IL-10 were identified in sera of KS patients [9] and KS tumors have shown elevated level of IL-6, IL-10 and IFN-γ [10]. Furthermore, in vitro studies have reported that high production of proinflammatory cytokines, such as IL-6, TNF-α, CCL-2 and, CXCL-10, may contribute to the progression of KS [11,12,13,14,15,16,17,18]. One of the mechanisms that may trigger KSHV reactivation, is the secretion of inflammatory cytokines, through activation of Toll-like receptors 7 and 8 (TLR7/8), oncostatin M (OSM), hepatocyte growth factor (HGF) and secretion of interferon-γ (IFN-γ) when stimulated by co-infection with other viruses [19,20]. Previous reports have also shown that KSHV infection induces proinflammatory cytokines including IL-1a, IL-1b, which are implicated in the pathogenesis of KSHV-associated malignancies [21,22].
Cytokines are soluble messengers controlling immune responses, and are involved in myriad biological process including inflammation, cell proliferation and cell migration. Manipulation of cytokines may have therapeutic benefit in diseases ranging from cancer to autoimmune manifestations and infectious disease. Multiple recombinant cytokines have been approved by the Food and Drug Administration (FDA) including IL-2 for the treatment of advanced renal cell carcinoma (RCC) [23] and IFN-α for the treatment of hairy cell leukemia [24] and Acquired immunodeficiency syndrome (AIDS)-related Kaposi’s sarcoma [25]. Development of cytokine-based therapies that target immunomodulatory mechanisms may be promising treatments for KSHV-associated diseases. Indeed, tocilizumab, the first humanized IL-6 receptor blocking antibody, has been approved for treatment of MCD in Japan. As knowledge of KSHV-associated diseases has grown, the research and the clinical trials have expanded to include cytokine targeted therapeutics (Table 1).
2. IL-6
Interleukin-6 is an inflammatory cytokine with pleiotropic effects produced by many cells in the body, such as epithelial cells, muscle cells, hematopoietic cells and stromal cells. It plays a role in various biological processes, including cancers and immune system diseases [26,27]. In the early stages of inflammation, monocytes, and macrophages secret IL-6 in response to activation of Toll-like receptors. Upregulation and persistent IL-6 stimulation are observed in in several inflammatory and autoimmune diseases [28]. The KSHV K2 gene [29] encodes a protein, viral IL-6 (vIL-6), that shares 24.8% amino acid similarity with human interleukin-6 (hIL-6) [29,30]. Several studies have uncovered roles of vIL-6 in KSHV-associated diseases, reviewed in [31]. Recently, a new in silico study presented the significant similarity between vIL6 and hIL6 folding and used this analysis to describe the similarity of the functions or interactions of both proteins, and defined four amino acid to be a new target to inhibit the pathological role of vIL6 [32]. A recent pilot study in 25 children with human immunodeficiency viruses (HIV)-associated KS in Malawi has demonstrated associations between KSHV viral load and hIL-6 levels in the blood, which may contribute to disease progression. [33]. Expression of vIL-6 protein has been detected in serum from patients with KS, PEL and MCD [34], and high levels of plasma hIL-6 drive progression of MCD and correlate with poor prognosis in PEL [35]. Moreover, experiments performed in AIDS/KS-derived cells demonstrated that hIL-6 is required for tumor cell proliferation [11]. vIL-6 can also serve as autocrine growth factor for PEL cell lines [36], and induces vascular endothelial growth factor (VEGF) in vitro and in vivo which plays a major role in PEL pathogenesis [37]. In addition, a study has reported that Tocilizumab, inhibited ascites formation in the BCBL-1 intraperitoneal xenograft mouse model of PEL [38], suggesting that hIL-6 is a viable therapeutic target in PEL. Patients with KICS, show elevated levels of hIL- 6 and vIL-6 [5]. KICS is one of the most aggressive clinical manifestations linked to KSHV infection, and it is associated with cytokine storm including high levels of hIL-6 [5], due to the overproduction of hIL-6 with high KSHV viral load it is most likely to contribute to the clinical features of the disease. Moreover, there is significant crosstalk between vIL-6 and hIL-6. vIL-6 can induce endogenous hIL-6 secretion in patients with MCD [39]. In an MCD mouse model, vIL-6 transgenic mice that have serum levels of vIL-6 comparable to KSHV-infected patients only develop the MCD-like phenotype if endogenous IL-6 is also present [40]. In 23 HIV/MCD patients, a strong correlation was found between the HHV8 viral load and plasma hIL-6 [41]. Due to the contribution of hIL-6 in the biology of KSHV and the elevated serum levels of hIL-6 which may contribute to the clinical features of MCD, inhibiting the activity of hIL6 is a promising therapeutic strategy for MCD which already has some data to support it. Current anti-IL-6 therapeutics include tocilizumab, an anti–interleukin-6 receptor antibody, and siltuximab, anti–hIL-6 monoclonal antibody. Both of these therapeutics are currently being used in clinical trials for therapeutic efficacy in MCD, and have shown promise when used either preventively or therapeutically [41]. One study has reported the efficacy of a rat anti-mouse IL-6R antibody, MR16-1, in a murine model of MCD in which transgenic mice carry a human IL-6 cDNA fused with a murine major histocompatibility class-I promoter (H-2Ld). Treatment of these mice with MR16-1 improved most of the MCD-like symptoms in compared with controls [42]. In humans, a clinical study evaluating the safety and efficacy of tocilizumab in 28 MCD patients for 16 weeks, showed that tocilizumab therapy alleviated the inflammatory symptoms and biochemical abnormalities associated with MCD with sustained improvement over one year [43]. Another retrospective study has reported that eleven out of twelve MCD patents treated with tocilizumab had partial remission and three achieved complete remission [44]. An additional Phase II clinical trial with tocilizumab, is currently in progress to determine if it will be an effective therapy for KSHV-MCD. (NCT01441063). In this study, 18 out of 23 patients (78%) had clinical benefits response and 12 patients (52%) showed objective tumor response. All 11 patients treated with the highest dose of 12 mg/kg had clinical benefit response and eight patients (73%) showed objective tumor response. A recent open label, single center pilot study has evaluated tocilizumab (8 mg/kg) administered intravenously on Day One of 14-day cycles for a maximum of six cycles both alone and in combination with zidovudine (AZT) and valganciclovir (VGC) in 8 KSHV-MCD patients with HIV. All patients in the study were on antiretroviral therapy (ART). One patient had complete clinical remission and four patients had partial responses on tocilizumab alone within the first two cycles of treatment. Of three patients who had AZT/VGC in combination with tocilizumab, two had partial responses and one had a complete remission response within four weeks of treatment [45]. The initial data for siltuximab activity comes from a Phase I, open-label, dose-finding study reported by Van Rhee and colleagues in 23 patients with symptomatic, multicentric or unresectable unicentric Castleman disease [46]. Moreover, a randomized, double-blind, placebo-controlled trial in 79 MCD patients treated with siltuximab showed a major benefit, 18 (34%) of 53 patients in the siltuximab group had durable tumor and symptomatic responses while no response was seen in 26 patients in the placebo group [47]. Further studies and clinical research are needed to better understand the role of elevated hIL-6 in MCD in order to develop this paradigm into an effective treatment strategy.
3. IL-10
Interleukin 10 is a potent anti-inflammatory cytokine produced by several immune cells, including dendritic cells, macrophages, B and T cells. Originally, IL-10 was labeled as cytokine synthesis inhibitory factor (CSIF) due to its activity as an inhibitor factor to the production of proinflammatory IFN-γ and TNFα cytokines by T helper l (Th1) cells. IL-10 has critical roles in the immune system primarily associated with regulating and suppressing the induction of proinflammatory cytokines to protect the host from inflammatory tissue damage. [48,49]. Several herpesviruses including cytomegalovirus (CMV), equine herpesvirus type 2 (EHV-2) and Epstein-Barr virus (EBV) encode viral homologs of IL-10, which contribute to immunosuppressive activity and suppress inflammatory responses [50,51,52]. The viral IL-10 homologs show immunomodulatory activities such as inhibition of DC maturation in both infected and bystander cells [53]. Many studies have suggested that anti-inflammatory cytokines including IL-10, are associated with the KSHV infection and/or AIDS KS development [54,55,56]. KSHV replication and transcription activator protein (K-RTA), activates the promoter of the human IL-10 gene [57]. K-RTA is an immediate early gene and is critical for the induction of lytic replication and the spread of KSHV infection [58,59,60]. Interestingly, studies have reported the induction of IL-10 secretion by macrophages during the KSHV infection or expression of the viral miRNAs miR-K12-3 and miR-K12-7 through targeting C/EBPβ p20 (LIP), a negative transcription regulator of IL-10 [61]. STAT3 activation resulting from dendritic cells exposure to UV-inactivated KSHV was correlated to IL-10 production [61]. More recently, studies have shown an elevated level of serum IL-10 in visceral AIDS-KS patients compared to HIV-positive individuals and classic KS patients [62]. In addition, IL-10 is expressed by PEL cells in vitro and in vivo and serves as autocrine growth factor for PEL-derived BCBL-1 and BC-1 cells [36]. Similarly, studies have demonstrated the involvement of elevated levels of cellular cytokines including IL-10 in the pathogenesis of KSHV-associated diseases, including PEL and MCD in patients with HIV/AIDS [36,54,63,64]. These studies suggest that IL-10 may have an important role in the development and progression of KSHV-associated diseases. A recent case study reported an elevated level of IL-10 and KSHV load in a KICS patient who, remarkably, not have elevated levels of hIL-6 [65]. These data indicate the powerful role of IL-10 in the pathogenesis of KSHV and the potential benefit of IL-10 as a biomarker for KICS patients. Neutralization of IL-10 been studied as therapeutic approach in viral infection [49,66]. Notably, IL-10 blockade restores IFN-γ production on HIV-1-Specific CD4 T Cell [67] and restores function of HIV specific and HCV specific T cells in vitro [68,69]. Thus, manipulation of IL-10 may hold promise as an immunotherapeutic strategy for KSHV-associated disease.
4. Interferons
Interferons (IFNs) are cytokines that play an important role in the immune system with potent antiviral activity. IFNs have a critical defense role against intracellular pathogens. In general, IFNs are classified into three types, Type I, Type II and Type III. Type I IFNs include IFN-α and IFN-β, which are released by virus infected cells. Type II IFNs include IFN-γ, which is secreted by natural killer cells, CD4+ T helper (Th1) cells and CD8+ cytotoxic T cells [70,71]. Although each type of IFN signals through different receptors, there is overlap in the signal transducing responses such as Jak-STAT signaling. [72,73,74,75]. While all types of IFNs induce the expression of antiviral proteins, there are divergent responses in KS patients. Patients who showed a response to IFN-α treatment with regression of KS lesions had an adequate number of CD4+ T cells compared to non-responders. [76,77,78]. KSHV infection upregulates the TLR3 pathway and its downstream target, interferon β (IFN-β) in monocytes [15]. In addition, KSHV infection induces IFN-α expression through TLR9 signaling in plasmacytoid dendritic cells [79]. Many KSHV proteins target the interferon response, and this inhibition may contribute to KSHV pathogenesis. Immediate-early lytic genes RTA and ORF45 inhibit the type I IFN pathway [80,81,82]. vIRFs are known to interact with cellular IRFs to block IFN production (reviewed in [83]). IRFs are best known for their rule in the regulation of IFN production and inflammatory responses downstream of pattern recognition receptors [84]. The latency-associated protein of KSHV, LANA inhibits the interferon pathway through binding to the IFN-β promoter [85]. Clinical trials for the treatment of KS patients with IFN-γ were halted due to unforeseen progression of KS lesions [86,87]. However, several groups have reported that viral lytic gene expression in BCBL-1 or BC-1 cells is promoted by IFN-γ [19,88]. Interestingly, recent data have shown inhibitory effects in viral progeny production with the IFN-γ treatment in infected primary human lymphatic endothelial cells (LECs) as well as induced KSHV-producer cells (iSLK.219) [89]. Moreover, treatment of BCBL-1 cells with IFN-γ induced expression of antiviral proteins such as double-stranded RNA-activated protein kinase (PKR) [90]. A recent study has reported the association between interferon lambda (IFNL3/4) polymorphisms and susceptibility to KS in HIV-infected individuals among men who have sex with men [91]. To date, there are only two interferon therapeutics that have been approved for KSHV-associated diseases, recombinant IFN-α2a, and IFN- α2b. Complete or partial anti-tumor response were observed in 38% of AIDS-KS patients with treatment of recombinant IFN-α with anti-viral effects in patients with the highest CD4 counts [92]. When low dose of IFN-α with didanosine was used in AIDS-KS, there was a 40% tumor response rate and the median response duration was 110 weeks [93]. On the other hand, an animal model study has demonstrated that IFN-α, in combination with azidothymidine results in significantly increased mean survival time in KSHV infected PEL- engrafted non-obese diabetic/severe combined immunodeficient (NOD/SCID) mice as well as induction of apoptosis in PEL cells [94]. Due to its side effects, IFN-α is rarely used therapeutically. However, the pegylated formulations of IFN-α showed partial or complete tumor response in 8 of 10 AIDS-KS patients, all were on combination antiretroviral therapy (cART) at peg-IFN initiation [95]. Another case has reported that pegylated formulations of IFN-α were efficient against KS skin lesions in AIDS-KS patient who were on cART [96]. Taken together, this evidence suggests that combining pegylated IFN-α with antiviral therapy may display potent efficacy against KSHV-associated diseases.
5. TNFa
Tumor necrosis factor (TNF) plays a major role in host defense mechanisms and the immune response [97]. TNF cytokines were described based on their ability to kill tumor cells in vitro and to cause hemorrhagic necrosis of transplantable tumors in mice [98,99]. The TNF family includes TNF-α and TNF-β. The role of TNF-α in KSHV infection is complex. In vivo studies have correlated TNF-α levels to KSHV infection and the progression of KS [100]. In fact, KSHV stimulates TNF-α production, possibly creating an environment that favors KS disease [101]. Moreover, KS lesions contain elevated levels of TNF-α [102]. A recent study has reported that KSHV glycoprotein B promotes cell adhesion and inhibits cell migration through upregulating TNF-α [103]. On the other hand, another study has reported that TNF-α impaired the production of KSHV virions by 90%, while TNF-β showed moderate inhibition of lytic reactivation in induced iSLK.219 and infected LECs [89]. Elevated levels of TNF-α and NFκB are present in CD19 + cells and serum samples from HIV/AIDS, KSHV coinfected patients who have antibodies against lytic antigens [104]. NF-κB is constitutively activated in KSHV-infected PEL cell lines and activity NF-κB is essential for KSHV-infected lymphoma cells survival [105]. These results suggest that pharmacologic inhibition of NF-κB may be an effective treatment for KSHV associated diseases. The NFκB is a family of transcription factors that control cell survival and may be activated by the recognition of pathogen-associated molecular patterns (PAMPs), such as dsRNA, viral dsDNA and TNF-α. [106]. Activated NFκB regulates the production of a variety of genes involved in immune responses and inflammation signaling, such as TNF-α, IL-1β, IL-2, IL-6, IL-18, IL-12, MCP-1 [107]. Mutation or inappropriate activation of NFκB signaling pathways has been correlated with autoimmunity, chronic inflammation, and cancer [108,109,110].
Furthermore, many studies support an association between TNF blockade and infection or reactivation of KSHV. High incidence of lymphoproliferative disorders has been linked with therapeutic TNF-α blockade [111]. There are many cases published in the literature of KS following treatment with anti-TNF-α including infliximab and golimumab: a case of a rheumatoid arthritis patient who developed KS lesion after infliximab treatment [112] and a patient with giant cell arteritis who developed KS in double-blind trial with anti-TNF-α [113], and a case of psoriatic arthritis patients who developed KS lesions during the course of treatment with infliximab [114]. Further, there was a case with history of psoriasis and psoriatic arthritis who developed KS during the treatment with golimumab [115]. However, another example has reported absence of KSHV reactivation after infliximab therapy in rheumatoid arthritis patients [116]. In addition, thalidomide appeared to inhibit TNF-α production [117]. A recent study has reported the role of pomalidomide, a thalidomide-like drug, as an immunomodulator with antitumor activity via PD-L1 inhibition [118] and through the restoration of ICAM-1 and B7-2 during latent infection and prevention of down-regulation of MHC-I during lytic activation in PEL cells [119]. Phase II dose-escalation study has reported that thalidomide (100 mg/day for 12 months) improved the clinical manifestation in AIDS-KS, eight out of seventeen patients achieved partial response and two patients had stable conditions [120]. A phase I/II study has evaluated pomalidomide (5 mg/day for 21/28 days), a small molecule derivative of thalidomide, in 22 KS patients with and without HIV infection [121]. 16 patients achieved objective tumor response, 9 out of 15 HIV infected patients achieved objective response, and all of 7 HIV uninfected patients achieved objective response, with median progression-free survival of 16.6 months. Moreover, 10 of 17 patients with edema (59%) showed a decrease in tumor associated edema. Whereas eight patients showed 2-cm reduction of limb circumference and two patients showed substantial subjective improvement. Interestingly, on May 14, 2020, the FDA granted accelerated approval to pomalidomide and expanded the indication to include AIDS-KS and KS patients who are HIV negative. Further studies are needed to address the benefits of anti TNF-α agents alone or in combination and investigate the precise role in other KSHV-associated diseases.
6. IL-12
Interleukin 12 (IL-12) was initially described as a “cytotoxic lymphocyte maturation factor” from PMA-induced EBV-transformed B-cell lines. It is a pleiotropic cytokine that links between innate and adaptive immunity [122]. In addition, IL-12 stimulates the production of IFN-γ with anticancer activity [123]. IL-12 has also been implicated in KS pathogenesis. It has been shown that IL-12 down regulates ORF74, the constitutively active G protein-coupled receptor that is encoded by KSHV [124,125,126,127,128]. Thus, IL-12 seemed a good candidate to be used as immunotherapy in humans. However, a very narrow therapeutic index and side effects limited the therapeutic use of this cytokine. Despite those setbacks, IL-12 shows promising results in some cases. Little et al. used IL-12 in a phase 1 pilot study in 32 AIDS-KS patients who were on highly active antiretroviral therapy (HAART), of 24 patients who were treated with higher doses (300, 500, 625 ng/kg), 17 had a complete or partial KS tumor response (61%) [129]. IL-12 was used in a Phase II trial for the treatment of AIDS-KS patients receiving cART, in combination with pegylated liposomal doxorubicin for six, three weeks cycles, followed by (500 ng/kg) IL-12 for 36 weeks, 83.3% had complete or partial KS tumor response [130]. At present, there is an ongoing Phase I/II trial of NHS-IL12, an IL-12 immunocytokine, genetically engineered and composed of two IL-12 heterodimers fused to the NHS76 antibody which targets the therapeutic to necrotic cells, as both a monotherapy and in combination therapy in advanced KS (NCT04303117) [131]. These studies suggest that IL-12 can have potent anti-KS effects in the context of combination therapy. Randomized trials will be needed to further evaluate IL-12 activity in KSHV-associated disease.
7. Host and Viral Chemokines
The KSHV genome encodes for three chemokine homologs, vCCL1/vMIP-I, vCCL2/vMIP-II and vCCL3/vMIP-III, encoded by ORFs K6, K4, and K4.1 respectively [29,30,132,133]. All of the chemokines are expressed during the lytic cycle [134,135]. vCCL1 is most highly related to CCL18 (50% amino acid identity) and to human CCL3 (37.9% amino acid identity) [29]. vCCL1 was identified reproducibly as selective viral CCR8 agonist in three independent studies, [136,137,138], suggesting it may function to promote the migration of Th2 lymphocytes, monocytes and endothelial cells. Indeed, vCCL1 is a potent chemoattractant for CCR8+ vascular endothelial cells [139], which could promote dissemination of KSHV within endothelial cells as well as KS tumorigenesis. In addition, vCCL1 was found to play an antiapoptotic role in PEL cells via induction of VEGF-A [140]. vCCL2 binds to a broad spectrum of chemokine receptors, and has primarily inhibitory effects [141]. It can bind to chemokine receptors from all four structural subgroups. In particular, it interacts with CCR1, CCR2, CCR3, CCR5, CCR8, CCR10, CXCR4, CX3CR1, and XCR1 [138,142,143]. vCCL2 blocks the binding of host ligands to CX3CR1 and CCR5, thereby inhibiting the migration of naïve and activated NK cells [144]. In addition, vCCL1 and vCCL2 enhance the survival of endothelial cells and to promote virus productive replication through CCR8 signaling [145]. However, contradicting studies have reported that vCCL2 act as an agonist or as antagonist in different models [146,147,148]. Despite this controversy, vCCL2 may has potential clinical applications. For example, recombinant vCCL2 has been used in rat models of inflammation and it has promising therapeutic effects [146,149]. Moreover, it has potential activity to enhance graft survival after organ transplantation [150,151,152], and a synthetic peptide derived from the N-terminus of vCCL2 can inhibit HIV-1 replication in CXCR4+ T-cell lines [153,154]. vCCL3 is related to vCCL1 and vCCL2 (25–28% amino acid identity) and most highly homologous to human XCL1. vCCL3 is highly selective and potent agonist at the human lymphotactin receptor XCR1, suggesting that it functions in KSHV immune evasion by modifying the migration and function of antigen presenting cells [138]. Furthermore, a study has reported that vCCL3 is CCR4 agonist [155]. This finding also supports the characterization of vCCL3 as an immunomodulatory factor, suggesting that it may modulate the function of regulatory T cells and antigen presenting cells during infection.
KSHV also encodes a chemokine receptor homolog, called ORF74. ORF74 has 27% homology with CXCR2 which are expressed in KS lesions [124]. ORF74 displays a broad chemokine-binding repertoire; binding to CXCR2 ligands including CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL7, CXCL8 and CC chemokines CCL1 and CCL5 [156,157]. Remarkably, it has been reported that ORF74 is constitutively active and induces a variety of intracellular signaling pathways [157]. CXCR2 ligands generally function as ORF74 agonists [156,158], while non-CXCR2 ligands including CXCL10 and CXCL12 act as inverse agonists [125,156,159]. Studies have reported KSHV infection of human monocytes upregulates human CCL2, and CXCL10 transcripts [15], and production of CXCL1 and CCL5 in SiHa cell [160]. CCL2 is correlated with increased angiogenesis and migration of tumor cells, which may drive KS progression, and immune cell migration [161,162]. In endothelial cell models, it has been reported that KSHV promotes the production of CCL2 via induction of ATF4 [163]. Moreover, vIL-6 signaling in endothelial cells increases CCL2 secretion via induction of hypoxia-upregulated protein 1 [164]. Furthermore, CXCL12/CXCR4 and CXCR7 are upregulated in KS and correlates with the severity of KS lesions [165].
CXCL1 has been investigated as a therapeutic target and potential diagnostic biomarker in other cancers [166,167]. A recent study has tested an anti-CXCL1 monoclonal antibody to inhibit the growth of bladder and prostate tumors [168]. In addition, CCL2-targeted therapy has been studied intensively in the field of cancer, especially in combination therapies [169,170,171]. These findings indicate that chemokine-targeted therapeutics may represent useful therapeutic targets in viral cancers, and highlight the potential role of chemokines as biomarkers for KSHV diagnosis and progression, but further information is needed to understand the nuance of chemokine dysregulation in KSHV disease before it can be exploited as a therapeutic paradigm.
8. Summary and Future Considerations
Cytokine based therapies have great potential for treating a variety of diseases including cancers and infectious diseases. Our understanding of the complex roles of cytokines in KSHV infection and disease is advancing, but many questions remain. In this review, we have attempted to summarize the current state of knowledge in this niche and highlight areas where cytokines may represent therapeutic targets for KSHV-associated diseases (Figure 1). Clearly, understanding the role of the cytokines during the immune response and their mechanisms in the biology of KSHV infection and tumorigenesis will facilitate development of new targets and enhance the efficacy of exiting cytokine-targeted therapies. The success of specific cytokine-targeted therapeutics such as IL-6, IL-15, IL-2 and IFN-α in different malignancies, will give rise to novel strategies to consider in the future of KSHV therapy. In addition, many diseases KSHV have similar clinical presentations and are difficult to diagnose histologically, therefore cytokine profiles may be considered to be a useful tool as biomarkers to differentiate these conditions. Genetic association studies of IFN lambda polymorphisms in KS, suggest a potential role of this cytokine in disease, and it will be interesting to determine if this class of interferons has therapeutic potential in KSHV-associated malignancy. Current research in the field of immunobiology of cytokines has resulted in promising ongoing clinical trials for cytokine-targeted therapy in KSHV-associated disease. Continued research in this field will provide additional insights into the major role of cytokines in the biology and pathogenesis of KSHV infection and KSHV-related diseases.
Author Contributions
Conceptualization, N.A. and J.T.; writing—original draft preparation, N.A.; writing—review and editing, J.T. and N.A.; supervision, J.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by NIH National Cancer Institute, grant number R01CA239590.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ablashi, D.V.; Chatlynne, L.G.; Whitman, J.E.; Cesarman, E. Spectrum of Kaposi’s sarcoma-associated herpesvirus, or human herpesvirus 8, diseases. Clin. Microbiol. Rev. 2002, 15, 439–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesarman, E.; Chang, Y.; Moore, P.S.; Said, J.W.; Knowles, D.M. Kaposi’s sarcoma–associated herpesvirus-like DNA sequences in AIDS-related body-cavity–based lymphomas. N. Engl. J. Med. 1995, 332, 1186–1191. [Google Scholar] [CrossRef] [PubMed]
- Soulier, J.; Grollet, L.; Oksenhendler, E.; Cacoub, P.; Cazals-Hatem, D.; Babinet, P.; d’Agay, M.; Clauvel, J.-P.; Raphael, M.; Degos, L. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman’s disease [see comments]. Blood 1995, 86, 1276–1280. [Google Scholar] [CrossRef] [Green Version]
- Uldrick, T.S.; Wang, V.; O’Mahony, D.; Aleman, K.; Wyvill, K.M.; Marshall, V.; Steinberg, S.M.; Pittaluga, S.; Maric, I.; Whitby, D. An interleukin-6-related systemic inflammatory syndrome in patients co-infected with Kaposi sarcoma-associated herpesvirus and HIV but without Multicentric Castleman disease. Clin. Infect. Dis. 2010, 51, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Levy, J.; Ziegler, J. Acquired immunodeficiency syndrome is an opportunistic infection and Kaposi’s sarcoma results from secondary immune stimulation. Lancet 1983, 322, 78–81. [Google Scholar] [CrossRef]
- Lukac, D.M.; Renne, R.; Kirshner, J.R.; Ganem, D. Reactivation of Kaposi’s sarcoma-associated herpesvirus infection from latency by expression of the ORF 50 transactivator, a homolog of the EBV R protein. Virology 1998, 252, 304–312. [Google Scholar] [CrossRef] [Green Version]
- Roth, W.K.; Brandstetter, H.; Sturzl, M. Cellular and molecular features of HIV-associated Kaposi’s sarcoma. Aids 1992, 6, 895–914. [Google Scholar] [CrossRef]
- Ramos, J.C.; Sin, S.H.; Staudt, M.R.; Roy, D.; Vahrson, W.; Dezube, B.J.; Harrington, W.; Dittmer, D.P. Nuclear factor kappa B pathway associated biomarkers in AIDS defining malignancies. Int. J. Cancer 2012, 130, 2728–2733. [Google Scholar] [CrossRef] [Green Version]
- Stürzl, M.; Brandstetter, H.; Zietz, C.; Eisenburg, B.; Raivich, G.; Gearing, D.P.; Brockmeyer, N.H.; Hofschneider, P. Identification of interleukin-1 and platelet-derived growth factor-B as major mitogens for the spindle cells of Kaposi’s sarcoma: A combined in vitro and in vivo analysis. Oncogene 1995, 10, 2007. [Google Scholar]
- Miles, S.A.; Rezai, A.R.; Salazar-Gonzalez, J.F.; Vander Meyden, M.; Stevens, R.H.; Logan, D.M.; Mitsuyasu, R.T.; Taga, T.; Hirano, T.; Kishimoto, T. AIDS Kaposi sarcoma-derived cells produce and respond to interleukin 6. Proc. Natl. Acad. Sci. USA 1990, 87, 4068–4072. [Google Scholar] [CrossRef] [Green Version]
- Sciacca, F.L.; Stürzl, M.; Bussolino, F.; Sironi, M.; Brandstetter, H.; Zietz, C.; Zhou, D.; Matteucci, C.; Peri, G.; Sozzani, S. Expression of adhesion molecules, platelet-activating factor, and chemokines by Kaposi’s sarcoma cells. J. Immunol. 1994, 153, 4816–4825. [Google Scholar] [PubMed]
- Ensoli, B.; Stürzl, M.; Monini, P. Cytokine-mediated growth promotion of Kaposi’s sarcoma and primary effusion lymphoma. Semin. Cancer Biol. 2000, 10, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, M.M.; Pietrek, M.; Dittrich-Breiholz, O.; Kracht, M.; Schulz, T.P. Modulation of host gene expression by the K15 protein of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2007, 81, 42–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.; Damania, B. Upregulation of the TLR3 pathway by Kaposi’s sarcoma-associated herpesvirus during primary infection. J. Virol. 2008, 82, 5440–5449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couty, J.-P.; Lupu-Meiri, M.; Oron, Y.; Gershengorn, M.C. Kaposi’s sarcoma-associated herpesvirus-G protein-coupled receptor-expressing endothelial cells exhibit reduced migration and stimulated chemotaxis by chemokine inverse agonists. J. Pharmacol. Exp. Ther. 2009, 329, 1142–1147. [Google Scholar] [CrossRef]
- Mesri, E.A.; Cesarman, E.; Boshoff, C. Kaposi’s sarcoma and its associated herpesvirus. Nat. Rev. Cancer 2010, 10, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Tso, F.Y.; Kossenkov, A.V.; Lidenge, S.J.; Ngalamika, O.; Ngowi, J.R.; Mwaiselage, J.; Wickramasinghe, J.; Kwon, E.H.; West, J.T.; Lieberman, P.M. RNA-Seq of Kaposi’s sarcoma reveals alterations in glucose and lipid metabolism. PLoS Pathog. 2018, 14, e1006844. [Google Scholar] [CrossRef] [Green Version]
- Mercader, M.; Taddeo, B.; Panella, J.R.; Chandran, B.; Nickoloff, B.J.; Foreman, K.E. Induction of HHV-8 lytic cycle replication by inflammatory cytokines produced by HIV-1-infected T cells. Am. J. Pathol. 2000, 156, 1961–1971. [Google Scholar] [CrossRef] [Green Version]
- Gregory, S.M.; West, J.A.; Dillon, P.J.; Hilscher, C.; Dittmer, D.P.; Damania, B. Toll-like receptor signaling controls reactivation of KSHV from latency. Proc. Natl. Acad. Sci. USA 2009, 106, 11725–11730. [Google Scholar] [CrossRef] [Green Version]
- Riva, G.; Barozzi, P.; Torelli, G.; Luppi, M. Immunological and inflammatory features of Kaposi’s sarcoma and other Kaposi’s sarcoma-associated herpesvirus/human herpesvirus 8-associated neoplasias. Aids Rev. 2010, 12, 40–51. [Google Scholar] [PubMed]
- Ensoli, B.; Stürzl, M. Kaposi’s sarcoma: A result of the interplay among inflammatory cytokines, angiogenic factors and viral agents. Cytokine Growth Factor Rev. 1998, 9, 63–83. [Google Scholar] [CrossRef]
- Fyfe, G.; Fisher, R.I.; Rosenberg, S.A.; Sznol, M.; Parkinson, D.R.; Louie, A.C. Results of treatment of 255 patients with metastatic renal cell carcinoma who received high-dose recombinant interleukin-2 therapy. J. Clin. Oncol. 1995, 13, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Golomb, H.M.; Jacobs, A.; Fefer, A.; Ozer, H.; Thompson, J.; Portlock, C.; Ratain, M.; Golde, D.; Vardiman, J.; Burke, J. Alpha-2 interferon therapy of hairy-cell leukemia: A multicenter study of 64 patients. J. Clin. Oncol. 1986, 4, 900–905. [Google Scholar] [CrossRef]
- Groopman, J.E.; Gottlieb, M.S.; Goodman, J.; Mitsuyasu, R.T.; Conant, M.A.; Prince, H.; Fahey, J.L.; Derezin, M.; Weinstein, W.M.; Casavante, C. Recombinant alpha-2 interferon therapy for Kaposi’s sarcoma associated with the acquired immunodeficiency syndrome. Ann. Intern. Med. 1984, 100, 671–676. [Google Scholar] [CrossRef]
- Trikha, M.; Corringham, T.; Klein, B.; Rossi, J.-F. Targeted anti-interleukin-6 monoclonal antibody therapy for cancer: A review of the rationale and clinical evidence. Clin. Cancer Res. 2003, 9, 4653–4665. [Google Scholar]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Srirangan, S.; Choy, E.H. The role of interleukin 6 in the pathophysiology of rheumatoid arthritis. Ther. Adv. Musculoskelet. Dis. 2010, 2, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Moore, P.S.; Boshoff, C.; Weiss, R.A.; Chang, Y. Molecular mimicry of human cytokine and cytokine response pathway genes by KSHV. Science 1996, 274, 1739–1744. [Google Scholar] [CrossRef]
- Nicholas, J.; Ruvolo, V.R.; Burns, W.H.; Sandford, G.; Wan, X.; Ciufo, D.; Hendrickson, S.B.; Guo, H.-G.; Hayward, G.S.; Reixz, M.S. Kaposi’s sarcoma-associated human herpesvirus-8 encodes homologues of macrophage inflammatory protein-1 and interleukin-6. Nat. Med. 1997, 3, 287–292. [Google Scholar] [CrossRef]
- Sakakibara, S.; Tosato, G. Viral Interleukin-6: Role in Kaposi’s Sarcoma-Associated Herpesvirus–Associated Malignancies. J. Interferon Cytokine Res. 2011, 31, 791–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghani, B.; Hashempour, T.; Hasanshahi, Z. Interaction of human herpesvirus 8 viral interleukin-6 with human interleukin-6 receptor using in silico approach: The potential role in HHV-8 pathogenesis. Curr. Proteom. 2020, 17, 107–116. [Google Scholar] [CrossRef]
- El-Mallawany, N.K.; Mehta, P.S.; Kamiyango, W.; Villiera, J.; Peckham-Gregory, E.C.; Kampani, C.; Krysiak, R.; Sanders, M.K.; Caro-Vegas, C.; Eason, A.B. KSHV viral load and Interleukin-6 in HIV-associated pediatric Kaposi sarcoma—Exploring the role of lytic activation in driving the unique clinical features seen in endemic regions. Int. J. Cancer 2019, 144, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Yarchoan, R.; Wyvill, K.; Okamoto, S.-I.; Little, R.F.; Tosato, G. Detection of viral interleukin-6 in Kaposi sarcoma–associated herpesvirus–linked disorders. Blood J. Am. Soc. Hematol. 2001, 97, 2173–2176. [Google Scholar] [CrossRef]
- Lurain, K.; Polizzotto, M.N.; Aleman, K.; Bhutani, M.; Wyvill, K.M.; Gonçalves, P.H.; Ramaswami, R.; Marshall, V.A.; Miley, W.; Steinberg, S.M. Viral, immunologic, and clinical features of primary effusion lymphoma. Blood J. Am. Soc. Hematol. 2019, 133, 1753–1761. [Google Scholar]
- Jones, K.D.; Aoki, Y.; Chang, Y.; Moore, P.S.; Yarchoan, R.; Tosato, G. Involvement of interleukin-10 (IL-10) and viral IL-6 in the spontaneous growth of Kaposi’s sarcoma herpesvirus-associated infected primary effusion lymphoma cells. Blood J. Am. Soc. Hematol. 1999, 94, 2871–2879. [Google Scholar]
- Aoki, Y.; Jaffe, E.S.; Chang, Y.; Jones, K.; Teruya-Feldstein, J.; Moore, P.S.; Tosato, G. Angiogenesis and Hematopoiesis Induced by Kaposi’s Sarcoma-Associated Herpesvirus-Encoded Interleukin-6: Presented in part at the 40th Annual American Society of Hematology Meeting, December 7, 1998 (Miami Beach, FL). Blood J. Am. Soc. Hematol. 1999, 93, 4034–4043. [Google Scholar]
- Goto, H.; Kudo, E.; Kariya, R.; Taura, M.; Katano, H.; Okada, S. Targeting VEGF and interleukin-6 for controlling malignant effusion of primary effusion lymphoma. J. Cancer Res. Clin. Oncol. 2015, 141, 465–474. [Google Scholar] [CrossRef]
- Mori, Y.; Nishimoto, N.; Ohno, M.; Inagi, R.; Dhepakson, P.; Amou, K.; Yoshizaki, K.; Yamanishi, K. Human herpesvirus 8-encoded interleukin-6 homologue (viral IL-6) induces endogenous human IL-6 secretion. J. Med. Virol. 2000, 61, 332–335. [Google Scholar] [CrossRef]
- Suthaus, J.; Stuhlmann-Laeisz, C.; Tompkins, V.S.; Rosean, T.R.; Klapper, W.; Tosato, G.; Janz, S.; Scheller, J.; Rose-John, S. HHV-8–encoded viral IL-6 collaborates with mouse IL-6 in the development of multicentric Castleman disease in mice. Blood 2012, 119, 5173–5181. [Google Scholar] [CrossRef]
- Oksenhendler, E.; Carcelain, G.; Aoki, Y.; Boulanger, E.; Maillard, A.; Clauvel, J.-P.; Agbalika, F.L. High levels of human herpesvirus 8 viral load, human interleukin-6, interleukin-10, and C reactive protein correlate with exacerbation of multicentric castleman disease in HIV-infected patients. Blood J. Am. Soc. Hematol. 2000, 96, 2069–2073. [Google Scholar]
- Katsume, A.; Saito, H.; Yamada, Y.; Yorozu, K.; Ueda, O.; Akamatsu, K.-I.; Nishimoto, N.; Kishimoto, T.; Yoshizaki, K.; Ohsugi, Y. Anti-interleukin 6 (IL-6) receptor antibody suppresses Castleman’s disease like symptoms emerged in IL-6 transgenic mice. Cytokine 2002, 20, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, N.; Kanakura, Y.; Aozasa, K.; Johkoh, T.; Nakamura, M.; Nakano, S.; Nakano, N.; Ikeda, Y.; Sasaki, T.; Nishioka, K. Humanized anti–interleukin-6 receptor antibody treatment of multicentric Castleman disease. Blood 2005, 106, 2627–2632. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, H.; Kadowaki, N.; Nishikori, M.; Kitawaki, T.; Kondo, T.; Ishikawa, T.; Yoshifuji, H.; Yamakawa, N.; Imura, T.; Mimori, T. Clinical features and treatment of multicentric castleman’s disease: A retrospective study of 21 Japanese patients at a single institute. J. Clin. Exp. Hematop. 2013, 53, 69–77. [Google Scholar] [CrossRef]
- Ramaswami, R.; Lurain, K.; Peer, C.J.; Serquiña, A.; Wang, V.; Widell, A.; Goncalves, P.; Steinberg, S.M.; Marshall, V.; George, J. Tocilizumab in Patients with Symptomatic Kaposi sarcoma Herpesvirus (KSHV)-associated Multicentric Castleman disease. Blood J. 2020, 135, 2316–2319. [Google Scholar] [CrossRef]
- Van Rhee, F.; Fayad, L.; Voorhees, P.; Furman, R.; Lonial, S.; Borghaei, H.; Sokol, L.; Crawford, J.; Cornfeld, M.; Qi, M. Siltuximab, a novel anti–interleukin-6 monoclonal antibody, for Castleman’s disease. J. Clin. Oncol. 2010, 28, 3701–3708. [Google Scholar] [CrossRef]
- Van Rhee, F.; Wong, R.S.; Munshi, N.; Rossi, J.-F.; Ke, X.-Y.; Fosså, A.; Simpson, D.; Capra, M.; Liu, T.; Hsieh, R.K. Siltuximab for multicentric Castleman’s disease: A randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2014, 15, 966–974. [Google Scholar] [CrossRef]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Brooks, D.G.; Trifilo, M.J.; Edelmann, K.H.; Teyton, L.; McGavern, D.B.; Oldstone, M.B. Interleukin-10 determines viral clearance or persistence in vivo. Nat. Med. 2006, 12, 1301–1309. [Google Scholar] [CrossRef]
- Kotenko, S.V.; Saccani, S.; Izotova, L.S.; Mirochnitchenko, O.V.; Pestka, S.V. Human cytomegalovirus harbors its own unique IL-10 homolog (cmvIL-10). Proc. Natl. Acad. Sci. USA 2000, 97, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Rode, H.-J.; Bugert, J.J.; Handermann, M.; Schnitzler, P.; Kehm, R.; Janssen, W.; Delius, H.; Darai, G. Molecular characterization and determination of the coding capacity of the genome of equine herpesvirus type 2 between the genome coordinates 0.235 and 0.258 (theEcoRI DNA fragment N; 4.2 kbp). Virus Genes 1994, 9, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Haig, D.M.; Fleming, S. Immunomodulation by virulence proteins of the parapoxvirus orf virus. Vet. Immunol. Immunopathol. 1999, 72, 81–86. [Google Scholar] [CrossRef]
- Raftery, M.J.; Wieland, D.; Gronewald, S.; Kraus, A.A.; Giese, T.; Schönrich, G. Shaping phenotype, function, and survival of dendritic cells by cytomegalovirus-encoded IL-10. J. Immunol. 2004, 173, 3383–3391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Kearney, P.; Plaisance, K.; Parsons, C.H. Pivotal advance: Kaposi’s sarcoma-associated herpesvirus (KSHV)-encoded microRNA specifically induce IL-6 and IL-10 secretion by macrophages and monocytes. J. Leukoc. Biol. 2010, 87, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Machado, P.R.L.; Farias, K.J.S.; Genre, J.; Oliveira, C.J.F.; Guedes, P.M.M.; da FonsecA, B.A.L. Disseminated Kaposi’s sarcoma in patients with HIV infection correlates to high serum levels of IL-10. Viral Immunol. 2014, 27, 356–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lidenge, S.J.; Tso, F.Y.; Ngalamika, O.; Ngowi, J.R.; Mortazavi, Y.; Kwon, E.H.; Shea, D.M.; Minhas, V.; Mwaiselage, V.; Wood, C. Similar Immunological Profiles Between African Endemic and Human Immunodeficiency Virus Type 1–Associated Epidemic Kaposi Sarcoma (KS) Patients Reveal the Primary Role of KS-Associated Herpesvirus in KS Pathogenesis. J. Infect. Dis. 2019, 219, 1318–1328. [Google Scholar] [CrossRef]
- Miyazawa, M.; Noguchi, K.; Kujirai, M.; Katayama, K.; Yamagoe, S.; Sugimoto, Y. IL-10 promoter transactivation by the viral K-RTA protein involves the host-cell transcription factors, specificity proteins 1 and 3. J. Biol. Chem. 2018, 293, 662–676. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Lin, S.-F.; Gradoville, L.; Yuan, Y.; Zhu, F.; Miller, G. A viral gene that activates lytic cycle expression of Kaposi’s sarcoma-associated herpesvirus. Proc. Natl. Acad. Sci. USA 1998, 95, 10866–10871. [Google Scholar] [CrossRef] [Green Version]
- Lukac, D.M.; Kirshner, J.R.; Ganem, D. Transcriptional activation by the product of open reading frame 50 of Kaposi’s sarcoma-associated herpesvirus is required for lytic viral reactivation in B cells. J. Virol. 1999, 73, 9348–9361. [Google Scholar] [CrossRef] [Green Version]
- Staudt, M.; Dittmer, D. The Rta/Orf50 transactivator proteins of the gamma-herpesviridae. In Kaposi Sarcoma Herpesvirus: New Perspectives; Springer: Berlin/Heidelberg, Germany, 2007; pp. 71–100. [Google Scholar]
- Santarelli, R.; Gonnella, R.; Di Giovenale, G.; Cuomo, L.; Capobianchi, A.; Granato, M.; Gentile, G.; Faggioni, A.; Cirone, M. STAT3 activation by KSHV correlates with IL-10, IL-6 and IL-23 release and an autophagic block in dendritic cells. Sci. Rep. 2014, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lopes, T.R.R.; Gonçales, J.P.; Júnior, J.V.J.S.; de Lorena, V.M.B.; Toscano, A.L.C.C.; Akamatsu, S.M.; Salles, A.C.; Tozetto-Mendoza, T.R.; de Morais, V.M.S.; Coêlho, M.R.C.D. Association of IL-6, IL-10 and CXCL10 serum concentrations with visceral Kaposi’s sarcoma in people living with HIV/AIDS. Hum. Immunol. 2020, 81, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Polizzotto, M.N.; Uldrick, T.S.; Hu, D.; Yarchoan, R. Clinical manifestations of Kaposi sarcoma herpesvirus lytic activation: Multicentric Castleman disease (KSHV–MCD) and the KSHV inflammatory cytokine syndrome. Front. Microbiol. 2012, 3, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasperini, P.; Sakakibara, S.; Tosato, G. Contribution of viral and cellular cytokines to Kaposi’s sarcoma-associated herpesvirus pathogenesis. J. Leukoc. Biol. 2008, 84, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Caro-Vegas, C.; Sellers, S.; Host, K.M.; Seltzer, J.; Landis, J.; Fischer II, W.A.; Damania, B.; Dittmer, D.P. Runaway Kaposi Sarcoma-associated herpesvirus replication correlates with systemic IL-10 levels. Virology 2020, 539, 18–25. [Google Scholar] [CrossRef]
- Ejrnaes, M.; von Herrath, M.G.; Christen, U. Cure of Chronic Viral Infection and Virus-Induced Type 1 Diabetes by Neutralizing Antibodies. Clin. Dev. Immunol. 2006, 13, 810853. [Google Scholar] [CrossRef]
- Porichis, F.; Hart, M.G.; Zupkosky, J.; Barblu, L.; Kwon, D.S.; McMullen, A.; Brennan, T.; Ahmed, R.; Freeman, G.J.; Kavanagh, D.G.; et al. Differential Impact of PD-1 and/or Interleukin-10 Blockade on HIV-1-Specific CD4 T Cell and Antigen-Presenting Cell Functions. J. Virol. 2014, 88, 2508–2518. [Google Scholar] [CrossRef] [Green Version]
- Wilson, E.B.; Brooks, D.G. The role of IL-10 in regulating immunity to persistent viral infections. Curr. Top. Microbiol. Immunol. 2011, 350, 39–65. [Google Scholar]
- Rigopoulou, E.I.; Abbott, W.G.H.; Haigh, P.; Naoumov, N.V. Blocking of interleukin-10 receptor--a novel approach to stimulate T-helper cell type 1 responses to hepatitis C virus. Clin. Immunol. 2005, 117, 57–64. [Google Scholar] [CrossRef]
- Biron, C.A. Initial and innate responses to viral infections—pattern setting in immunity or disease. Curr. Opin. Microbiol. 1999, 2, 374–381. [Google Scholar] [CrossRef]
- Biron, C.A.; Nguyen, K.B.; Pien, G.C.; Cousens, L.P.; Salazar-Mather, T.P. Natural killer cells in antiviral defense: Function and regulation by innate cytokines. Annu. Rev. Immunol. 1999, 17, 189–220. [Google Scholar] [CrossRef]
- Doly, J.; Civas, A.; Navarro, S.; Uze, G. Type I interferons: Expression and signalization. Cell. Mol. Life Sci. CMLS 1998, 54, 1109–1121. [Google Scholar] [CrossRef]
- Schindler, C. Cytokines and JAK–STAT signaling. Exp. Cell Res. 1999, 253, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.W. Series introduction: JAK-STAT signaling in human disease. J. Clin. Investig. 2002, 109, 1133–1137. [Google Scholar] [CrossRef]
- Schindler, C.; Brutsaert, S. Interferons as a paradigm for cytokine signal transduction. Cell. Mol. Life Sci. CMLS 1999, 55, 1509–1522. [Google Scholar] [CrossRef]
- Fischl, M.A.; Finkelstein, D.M.; He, W.; Powderly, W.G.; Triozzi, P.L.; Steigbigel, R.T. A phase II study of recombinant human interferon-α2a and zidovudine in patients with AIDS-related Kaposi’s sarcoma. JAIDS J. Acquir. Immune Defic. Syndr. 1996, 11, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Krown, S.E. Interferon and other biologic agents for the treatment of Kaposi’s sarcoma. Hematol./Oncol. Clin. N. Am. 1991, 5, 311–322. [Google Scholar] [CrossRef]
- Sawyer, L.A.; Metcalf, J.A.; Zoon, K.C.; Boone, E.J.; Kovacs, J.A.; Lane, H.C.; Quinnan Jr, G.V. Effects of interferon-α in patients with aids-associated Kaposi’s sarcoma are related to blood interferon levels and dose. Cytokine 1990, 2, 247–252. [Google Scholar] [CrossRef]
- West, J.A.; Gregory, S.M.; Sivaraman, V.; Su, L.; Damania, B. Activation of plasmacytoid dendritic cells by Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2011, 85, 895–904. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Hayward, G.S. The ubiquitin E3 ligase RAUL negatively regulates type i interferon through ubiquitination of the transcription factors IRF7 and IRF3. Immunity 2010, 33, 863–877. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wang, S.E.; Hayward, G.S. The KSHV immediate-early transcription factor RTA encodes ubiquitin E3 ligase activity that targets IRF7 for proteosome-mediated degradation. Immunity 2005, 22, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.X.; King, S.M.; Smith, E.L.; Levy, D.E.; Yuan, Y. A Kaposi’s sarcoma-associated herpesviral protein inhibits virus-mediated induction of type I interferon by blocking IRF-7 phosphorylation and nuclear accumulation. Proc. Natl. Acad. Sci. USA 2002, 99, 5573–5578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damania, B.; Jacobs, S.R. The viral interferon regulatory factors of KSHV: Immunosuppressors or oncogenes? Front. Immunol. 2011, 2, 19. [Google Scholar]
- Honda, K.; Taniguchi, T. IRFs: Master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol. 2006, 6, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, N.; Flamand, L. Kaposi sarcoma-associated herpesvirus latency-associated nuclear antigen inhibits interferon (IFN) β expression by competing with IFN regulatory factor-3 for binding to IFNB promoter. J. Biol. Chem. 2010, 285, 7208–7221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganser, A.; Brücher, W.; Brodt, H.; Busch, W.; Brandhorst, I.; Helm, E.; Hoelzer, D. Treatment of AIDS-Related Kaposi’s Sarcoma with Recombinant γ-Interferon. Oncol. Res. Treat. 1986, 9, 163–166. [Google Scholar] [CrossRef]
- Krigel, R.L.; Odajnyk, C.M.; Laubenstein, L.J.; Ostreicher, R.; Wernz, J.; Vilcek, J.; Rubinstein, P.; Friedman-Kien, A.E. Therapeutic trial of interferon-gamma in patients with epidemic Kaposi’s sarcoma. J. Biol. Response Modif. 1985, 4, 358–364. [Google Scholar]
- Chang, J.; Renne, R.; Dittmer, D.; Ganem, D. Inflammatory cytokines and the reactivation of Kaposi’s sarcoma-associated herpesvirus lytic replication. Virology 2000, 266, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Park, M.-K.; Cho, H.; Roh, S.W.; Kim, S.-J.; Myoung, J. Cell Type-Specific Interferon-γ-mediated Antagonism of KSHV Lytic Replication. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pozharskaya, V.P.; Weakland, L.L.; Offermann, M.K. Inhibition of infectious human herpesvirus 8 production by gamma interferon and alpha interferon in BCBL-1 cells. J. Gen. Virol. 2004, 85, 2779–2787. [Google Scholar] [CrossRef]
- Bibert, S.; Wójtowicz, A.; Taffé, P.; Tarr, P.E.; Bernasconi, E.; Furrer, H.; Günthard, H.F.; Hoffmann, M.; Kaiser, L.; Osthoff, M. Interferon lambda 3/4 polymorphisms are associated with AIDS-related Kaposi’s sarcoma. Aids 2018, 32, 2759–2765. [Google Scholar] [CrossRef] [Green Version]
- Lane, H.C.; Feinberg, J.; Davey, V.; Deyton, L.; Baseler, M.; Manischewitz, J.; Masur, H.; Kovacs, J.; Herpin, B.; Walker, R. Anti-retroviral effects of interferon-α in AIDS-associated Kaposi’s sarcoma. Lancet 1988, 332, 1218–1222. [Google Scholar] [CrossRef]
- Krown, S.E.; Li, P.; Von Roenn, J.H.; Paredes, J.; Huang, J.; Testa, M.A. Efficacy of low-dose interferon with antiretroviral therapy in Kaposi’s sarcoma: A randomized phase II AIDS clinical trials group study. J. Interferon Cytokine Res. 2002, 22, 295–303. [Google Scholar] [CrossRef]
- Wu, W.; Rochford, R.; Toomey, L.; Harrington Jr, W.; Feuer, G. Inhibition of HHV-8/KSHV infected primary effusion lymphomas in NOD/SCID mice by azidothymidine and interferon-α. Leuk. Res. 2005, 29, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Rokx, C.; van der Ende, M.E.; Verbon, A.; Rijnders, B.J. Peginterferon alfa-2a for AIDS-associated Kaposi sarcoma: Experience with 10 patients. Clin. Infect. Dis. 2013, 57, 1497–1499. [Google Scholar] [CrossRef] [Green Version]
- Rouanet, I.; Lechiche, C.; Doncesco, R.; Mauboussin, J.M.; Sotto, A. Interferon therapy for Kaposi sarcoma associated with acquired immunodeficiency syndrome: Still a valid treatment option? AIDS Patient Care STDs 2013, 27, 537–538. [Google Scholar] [CrossRef]
- Kollias, G.; Douni, E.; Kassiotis, G.; Kontoyiannis, D. The function of tumour necrosis factor and receptors in models of multi-organ inflammation, rheumatoid arthritis, multiple sclerosis and inflammatory bowel disease. Ann. Rheum. Dis. 1999, 58 (Suppl. 1), I32–I39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Old, L.J. Tumor necrosis factor. Science 1985, 230, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Bazzoni, F.; Beutler, B. The tumor necrosis factor ligand and receptor families. N. Engl. J. Med. 1996, 334, 1717–1725. [Google Scholar] [CrossRef]
- Aboulafia, D.; Miles, S.A.; Saks, S.R.; Mitsuyasu, R.T. Intravenous recombinant tumor necrosis factor in the treatment of AIDS-related Kaposi’s sarcoma. JAIDS J. Acquir. Immune Defic. Syndr. 1989, 2, 54–58. [Google Scholar]
- Murakami-Mori, K.; Mori, S.; Bonavida, B.; Nakamura, S. Implication of TNF receptor-I-mediated extracellular signal-regulated kinases 1 and 2 (ERK1/2) activation in growth of AIDS-associated Kaposi’s sarcoma cells: A possible role of a novel death domain protein MADD in TNF-α-induced ERK1/2 activation in Kaposi’s sarcoma cells. J. Immunol. 1999, 162, 3672–3679. [Google Scholar]
- Oxholm, A.; Oxholm, P.; Permin, H.; Bendtzen, K. Epidermal tumour necrosis factor α and interleukin 6-like activities in AIDS-related Kaposi’s sarcoma: An immunohistological study. Apmis 1989, 97, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.A.; UAbdel-Raouf, M.; Akula, S.M. Membrane-Associated Kaposi Sarcoma-Associated Herpesvirus Glycoprotein B Promotes Cell Adhesion and Inhibits Migration of Cells via Upregulating IL-1β and TNF-α. Intervirology 2017, 60, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Gonçales, J.P.; Lopes, T.R.R.; de Lorena, V.M.B.; Aciole, M.R.; Júnior, J.V.J.S.; Tozetto-Mendoza, T.R.; Braz-Silva, P.H.; Coêlho, M.R.C.D. Association of NFκB and related-cytokines with the viral load and development of antibodies against HHV-8 in people living with HIV/AIDS. Med Microbiol. Immunol. 2020, 209, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.A.; Schattner, E.J.; Cesarman, E. Inhibition of NF-κB induces apoptosis of KSHV-infected primary effusion lymphoma cells. Blood J. Am. Soc. Hematol. 2000, 96, 2537–2542. [Google Scholar]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S. NF-ĸB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Oeckinghaus, A.; Ghosh, S. The NF-κB family of transcription factors and its regulation. Cold Spring Harbor Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Basseres, D.; Baldwin, A. Nuclear factor-κ B and inhibitor of κ B kinase pathways in oncogenic initiation and progression. Oncogene 2006, 25, 6817–6830. [Google Scholar] [CrossRef] [Green Version]
- Napetschnig, J.; Wu, H. Molecular basis of NF-κB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef] [Green Version]
- Courtois, G.; Gilmore, T. Mutations in the NF-κ B signaling pathway: Implications for human disease. Oncogene 2006, 25, 6831–6843. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.L.; Greene, M.H.; Gershon, S.K.; Edwards, E.T.; Braun, M.M. Tumor necrosis factor antagonist therapy and lymphoma development: Twenty-six cases reported to the Food and Drug Administration. Arthritis Rheum. 2002, 46, 3151–3158. [Google Scholar] [CrossRef]
- Cohen, C.; Horster, S.; Sander, C.; Bogner, J. Kaposi’s sarcoma associated with tumour necrosis factor α neutralising therapy. Ann. Rheum. Dis. 2003, 62, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuttikat, A.; Joshi, A.; Saeed, I.; Chakravarty, K. Kaposi sarcoma in a patient with giant cell arteritis. Dermatol. Online J. 2006, 12, 16. [Google Scholar] [PubMed]
- Ursini, F.; Naty, S.; Mazzei, V.; Spagnolo, F.; Grembiale, R.D. Kaposi’s sarcoma in a psoriatic arthritis patient treated with infliximab. Int. Immunopharmacol. (Print) 2010, 10, 827–828. [Google Scholar] [CrossRef]
- Vural, S.; Gündoğdu, M.; Akay, B.N.; Korkmaz, P.; Şanli, H.; Heper, A.O.; Kundakci, N. Aggressive Kaposi’s Sarcoma Associated With Golimumab Therapy. Arch. Rheumatol. 2018, 33, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Torre-Cisneros, J.; Del Castillo, J.M.; Caston, J.; Castro, M.; Perez, V.; Collantes, E. Infliximab does not activate replication of lymphotropic herpesviruses in patients with refractory rheumatoid arthritis. Rheumatology 2005, 44, 1132–1135. [Google Scholar] [CrossRef] [Green Version]
- Klausner, J.D.; VFreedman, H.; Kaplan, G. Thalidomide as an anti-TNF-α inhibitor: Implications for clinical use. Clin. Immunol. Immunopathol. 1996, 81, 219–223. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Sun, Y.; Torphy, R.J.; He, J.; Yanaga, K.; Edil, B.H.; Schulick, R.D.; Zhu, Y. Pomalidomide inhibits PD-L1 induction to promote antitumor immunity. Cancer Res. 2018, 78, 6655–6665. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Mishra, S.; Anagho, H.A.; Aisabor, A.I.; Shrestha, P.; Wang, V.; Takamatsu, Y.; Maeda, K.; Mitsuya, H.; Zeldis, J.B. Restoration of immune surface molecules in Kaposi sarcoma-associated herpes virus infected cells by lenalidomide and pomalidomide. Oncotarget 2017, 8, 50342. [Google Scholar] [CrossRef] [Green Version]
- Little, R.F.; Wyvill, K.M.; Pluda, J.M.; Welles, L.; Marshall, V.; Figg, W.D.; Newcomb, F.M.; Tosato, G.; Feigal, E.; Steinberg, S.M. Activity of thalidomide in AIDS-related Kaposi’s sarcoma. J. Clin. Oncol. 2000, 18, 2593–2602. [Google Scholar] [CrossRef]
- Polizzotto, M.N.; Uldrick, T.S.; Wyvill, K.M.; Aleman, K.; Peer, C.J.; Bevans, M.; Sereti, I.; Maldarelli, F.; Whitby, D.; Marshall, V. Pomalidomide for symptomatic Kaposi’s sarcoma in people with and without HIV infection: A phase I/II study. J. Clin. Oncol. 2016, 34, 4125. [Google Scholar] [CrossRef]
- Stern, A.S.; Podlaski, F.J.; Hulmes, J.D.; Pan, Y.; Quinn, P.M.; Wolitzky, A.; Familletti, P.C.; Stremlo, D.L.; Truitt, T.; Chizzonite, R. Purification to homogeneity and partial characterization of cytotoxic lymphocyte maturation factor from human B-lymphoblastoid cells. Proc. Natl. Acad. Sci. USA 1990, 87, 6808–6812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakóbisiak, M.; Lasek, W.; Gołąb, J. Natural mechanisms protecting against cancer. Immunol. Lett. 2003, 90, 103–122. [Google Scholar] [CrossRef]
- Cesarman, E.; Nador, R.G.; Bai, F.; Bohenzky, R.A.; Russo, J.J.; Moore, P.S.; Chang, Y.; Knowles, D.M. Kaposi’s sarcoma-associated herpesvirus contains G protein-coupled receptor and cyclin D homologs which are expressed in Kaposi’s sarcoma and malignant lymphoma. J. Virol. 1996, 70, 8218–8223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geras-Raaka, E.; Varma, A.; Ho, H.; Clark-Lewis, I.; Gershengorn, M.C. Human Interferon-γ–inducible Protein 10 (IP-10) Inhibits Constitutive Signaling of Kaposi’s Sarcoma–associated Herpesvirus G Protein–coupled Receptor. J. Exp. Med. 1998, 188, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodhi, A.; Montaner, S.; Patel, V.; Zohar, M.; Bais, C.; Mesri, E.A.; Gutkind, J.S. The Kaposi’s sarcoma-associated herpes virus G protein-coupled receptor up-regulates vascular endothelial growth factor expression and secretion through mitogen-activated protein kinase and p38 pathways acting on hypoxia-inducible factor 1α. Cancer Res. 2000, 60, 4873–4880. [Google Scholar] [PubMed]
- Bais, C.; Santomasso, B.; Coso, O.; Arvanitakis, L.; Raaka, E.G.; Gutkind, J.S.; Asch, A.S.; Cesarman, E.; Gerhengorn, M.C.; Mesri, E.A. G-protein-coupled receptor of Kaposi’s sarcoma-associated herpesvirus is a viral oncogene and angiogenesis activator. Nature 1998, 391, 86–89. [Google Scholar] [CrossRef]
- Nakamura, S.; Murakami-Mori, K.; Rao, N.; Weich, H.A.; Rajeev, B. Vascular endothelial growth factor is a potent angiogenic factor in AIDS-associated Kaposi’s sarcoma-derived spindle cells. J. Immunol. 1997, 158, 4992–5001. [Google Scholar]
- Little, R.F.; Pluda, J.M.; Wyvill, K.M.; Rodriguez-Chavez, I.R.; Tosato, G.; Catanzaro, A.T.; Steinberg, S.M.; Yarchoan, R. Activity of subcutaneous interleukin-12 in AIDS-related Kaposi sarcoma. Blood 2006, 107, 4650–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, R.F.; Aleman, K.; Kumar, P.; Wyvill, K.M.; Pluda, J.M.; Read-Connole, E.; Wang, V.; Pittaluga, S.; Catanzaro, A.T.; Steinberg, S.M. Phase 2 study of pegylated liposomal doxorubicin in combination with interleukin-12 for AIDS-related Kaposi sarcoma. Blood J. Am. Soc. Hematol. 2007, 110, 4165–4171. [Google Scholar] [CrossRef] [Green Version]
- Sharifi, J.; Khawli, L.A.; Hu, P.; King, S.; Epstein, A.L. Characterization of a phage display-derived human monoclonal antibody (NHS76) counterpart to chimeric TNT-1 directed against necrotic regions of solid tumors. Hybrid. Hybridomics 2001, 20, 305–312. [Google Scholar] [CrossRef]
- Neipel, F.; Albrecht, J.-C.; Fleckenstein, B. Cell-homologous genes in the Kaposi’s sarcoma-associated rhadinovirus human herpesvirus 8: Determinants of its pathogenicity? J. Virol. 1997, 71, 4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, J.J.; Bohenzky, R.A.; Chien, M.-C.; Chen, J.; Yan, M.; Maddalena, D.; Parry, J.P.; Peruzzi, D.; Edelman, I.S.; Chang, Y. Nucleotide sequence of the Kaposi sarcoma-associated herpesvirus (HHV8). Proc. Natl. Acad. Sci. USA 1996, 93, 14862–14867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenner, R.G.; Albà, M.M.; Boshoff, C.; Kellam, P. Kaposi’s sarcoma-associated herpesvirus latent and lytic gene expression as revealed by DNA arrays. J. Virol. 2001, 75, 891–902. [Google Scholar] [CrossRef] [Green Version]
- Paulose-Murphy, M.; Ha, N.-K.; Xiang, C.; Chen, Y.; Gillim, L.; Yarchoan, R.; Meltzer, P.; Bittner, M.; Trent, J.; Zeichner, S. Transcription program of human herpesvirus 8 (Kaposi’s sarcoma-associated herpesvirus). J. Virol. 2001, 75, 4843–4853. [Google Scholar] [CrossRef] [Green Version]
- Dairaghi, D.J.; Fan, R.A.; McMaster, B.E.; Hanley, M.R.; Schall, T.J. HHV8-encoded vMIP-I selectively engages chemokine receptor CCR8 Agonist and antagonist profiles of viral chemokines. J. Biol. Chem. 1999, 274, 21569–21574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endres, M.J.; Garlisi, C.G.; Xiao, H.; Shan, L.; Hedrick, J.A. The Kaposi’s sarcoma–related herpesvirus (KSHV)-encoded chemokine vMIP-I is a specific agonist for the CC chemokine receptor (CCR) 8. J. Exp. Med. 1999, 189, 1993–1998. [Google Scholar] [CrossRef] [Green Version]
- Lüttichau, H.R.; Johnsen, A.H.; Jurlander, J.; Rosenkilde, M.M.; Schwartz, T.W. Kaposi sarcoma-associated herpes virus targets the lymphotactin receptor with both a broad spectrum antagonist vCCL2 and a highly selective and potent agonist vCCL3. J. Biol. Chem. 2007, 282, 17794–17805. [Google Scholar] [CrossRef] [Green Version]
- Haque, N.S.; Fallon, J.T.; Taubman, M.B.; Harpel, P.C. The chemokine receptor CCR8 mediates human endothelial cell chemotaxis induced by I-309 and Kaposi sarcoma herpesvirus-encoded vMIP-I and by lipoprotein (a)-stimulated endothelial cell conditioned medium. Blood J. Am. Soc. Hematol. 2001, 97, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Okruzhnov, Y.; Li, H.; Nicholas, J. Human herpesvirus 8 (HHV-8)-encoded cytokines induce expression of and autocrine signaling by vascular endothelial growth factor (VEGF) in HHV-8-infected primary-effusion lymphoma cell lines and mediate VEGF-independent antiapoptotic effects. J. Virol. 2001, 75, 10933–10940. [Google Scholar] [CrossRef] [Green Version]
- Szpakowska, M.; Chevigné, A. vCCL2/vMIP-II, the viral master KEYmokine. J. Leukoc. Biol. 2016, 99, 893–900. [Google Scholar] [CrossRef]
- Kledal, T.N.; Rosenkilde, M.M.; Coulin, F.; Simmons, G.; Johnsen, A.H.; Alouani, S.; Power, C.A.; Lüttichau, H.R.; Gerstoft, J.; Clapham, P.R. A broad-spectrum chemokine antagonist encoded by Kaposi’s sarcoma-associated herpesvirus. Science 1997, 277, 1656–1659. [Google Scholar] [CrossRef] [PubMed]
- Lüttichau, H.R.; Lewis, I.C.; Gerstoft, J.; Schwartz, T. The herpesvirus 8-encoded chemokine vMIP-II, but not the poxvirus-encoded chemokine MC148, inhibits the CCR10 receptor. Eur. J. Immunol. 2001, 31, 1217–1220. [Google Scholar] [CrossRef]
- Yamin, R.; Kaynan, N.S.; Glasner, A.; Vitenshtein, A.; Tsukerman, P.; Bauman, Y.; Ophir, Y.; Elias, S.; Bar-On, Y.; Gur, C. The viral KSHV chemokine vMIP-II inhibits the migration of Naive and activated human NK cells by antagonizing two distinct chemokine receptors. PLoS Pathog. 2013, 9, e1003568. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.B.; Nicholas, J. Autocrine and paracrine promotion of cell survival and virus replication by human herpesvirus 8 chemokines. J. Virol. 2008, 82, 6501–6513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Bacon, K.B.; Li, L.; Garcia, G.E.; Xia, Y.; Lo, D.; Thompson, D.A.; Siani, M.A.; Yamamoto, T.; Harrison, J.K. In vivo inhibition of CC and CX3C chemokine–induced leukocyte infiltration and attenuation of glomerulonephritis in Wistar-Kyoto (WKY) rats by vMIP-II. J. Exp. Med. 1998, 188, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Sozzani, S.; Luini, W.; Bianchi, G.; Allavena, P.; Wells, T.; Napolitano, M.; Bernardini, G.; Vecchi, A.; D’Ambrosio, D.; Mazzeo, D. The viral chemokine macrophage inflammatory protein-II is a selective Th2 chemoattractant. Blood J. Am. Soc. Hematol. 1998, 92, 4036–4039. [Google Scholar]
- Nakano, K.; Isegawa, Y.; Zou, P.; Tadagaki, K.; Inagi, R.; Yamanishi, K. Kaposi’s sarcoma-associated herpesvirus (KSHV)-encoded vMIP-I and vMIP-II induce signal transduction and chemotaxis in monocytic cells. Arch. Virol. 2003, 148, 871–890. [Google Scholar] [CrossRef]
- Ghirnikar, R.; Lee, Y.; Eng, L. Chemokine antagonist infusion attenuates cellular infiltration following spinal cord contusion injury in rat. J. Neurosci. Res. 2000, 59, 63–73. [Google Scholar] [CrossRef]
- Cherqui, S.; Kingdon, K.M.; Thorpe, C.; Kurian, S.M.; Salomon, D.R. Lentiviral gene delivery of vMIP-II to transplanted endothelial cells and endothelial progenitors is proangiogenic in vivo. Mol. Ther. 2007, 15, 1264–1272. [Google Scholar] [CrossRef]
- DeBruyne, L.; Li, K.; Bishop, D.; Bromberg, J. Gene transfer of virally encoded chemokine antagonists vMIP-II and MC148 prolongs cardiac allograft survival and inhibits donor-specific immunity. Gene Ther. 2000, 7, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Pillai, R.G.; Beutelspacher, S.C.; Larkin, D.F.P.; George, A.J. Expression of the chemokine antagonist vMIP II using a non-viral vector can prolong corneal allograft survival. Transplantation 2008, 85, 1640–1647. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Fan, X.; Zhou, N.; Hiraoka, M.; Luo, J.; Kaji, H.; Huang, Z. Structure−function study and anti-HIV activity of synthetic peptide analogues derived from viral chemokine vMIP-II. Biochemistry 2000, 39, 13545–13550. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Luo, Z.; Luo, J.; Hall, J.W.; Huang, Z. A novel peptide antagonist of CXCR4 derived from the N-terminus of viral chemokine vMIP-II. Biochemistry 2000, 39, 3782–3787. [Google Scholar] [CrossRef] [PubMed]
- Stine, J.T.; Wood, C.; Hill, M.; Epp, A.; Raport, C.J.; Schweickart, V.L.; Endo, Y.; Sasaki, T.; Simmons, G.; Boshoff, C. KSHV-encoded CC chemokine vMIP-III is a CCR4 agonist, stimulates angiogenesis, and selectively chemoattracts TH2 cells. Blood J. Am. Soc. Hematol. 2000, 95, 1151–1157. [Google Scholar] [CrossRef]
- Rosenkilde, M.M.; Kledal, T.N.; Bräuner-Osborne, H.; Schwartz, T.W. Agonists and inverse agonists for the herpesvirus 8-encoded constitutively active seven-transmembrane oncogene product, ORF-74. J. Biol. Chem. 1999, 274, 956–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvanitakis, L.; Geras-Raaka, E.; Varma, A.; Gershengorn, M.C.; Cesarman, E. Human herpesvirus KSHV encodes a constitutively active G-protein-coupled receptor linked to cell proliferation. Nature 1997, 385, 347–350. [Google Scholar] [CrossRef]
- De Munnik, S.M.; Kooistra, A.J.; Van Offenbeek, J.; Nijmeijer, S.; de Graaf, C.; Smit, M.J.; Leurs, R.; Vischer, H.F. The viral G protein-coupled receptor ORF74 hijacks β-arrestins for endocytic trafficking in response to human chemokines. PLoS ONE 2015, 10, e0124486. [Google Scholar] [CrossRef] [Green Version]
- Gershengorn, M.C.; Geras-Raaka, E.; Varma, A.; Clark-Lewis, I. Chemokines activate Kaposi’s sarcoma-associated herpesvirus G protein-coupled receptor in mammalian cells in culture. J. Clin. Investig. 1998, 102, 1469–1472. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Cao, Y.; Jiang, W.; Zabaleta, J.; Liu, Z.; Qiao, J.; Qin, Z. KSHV co-infection down-regulates HPV16 E6 and E7 from cervical cancer cells. Oncotarget 2017, 8, 35792. [Google Scholar] [CrossRef]
- Qian, B.-Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Wolf, M.J.; Hoos, A.; Bauer, J.; Boettcher, S.; Knust, M.; Weber, A.; Simonavicius, N.; Schneider, C.; Lang, M.; Stürzl, M. Endothelial CCR2 signaling induced by colon carcinoma cells enables extravasation via the JAK2-Stat5 and p38MAPK pathway. Cancer Cell 2012, 22, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caselli, E.; Benedetti, S.; Grigolato, J.; Caruso, A.; Di Luca, D. Activating transcription factor 4 (ATF4) is upregulated by human herpesvirus 8 infection, increases virus replication and promotes proangiogenic properties. Arch. Virol. 2012, 157, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Giffin, L.; Yan, F.; Major, M.B.; Damania, B. Modulation of Kaposi’s sarcoma-associated herpesvirus interleukin-6 function by hypoxia-upregulated protein 1. J. Virol. 2014, 88, 9429–9441. [Google Scholar] [CrossRef] [Green Version]
- Desnoyer, A.; Dupin, N.; Assoumou, L.; Carlotti, A.; Gaudin, F.; Deback, C.; Peytavin, G.; Marcelin, A.; Boué, F.; Balabanian, K. Expression pattern of the CXCL 12/CXCR 4–CXCR 7 trio in Kaposi sarcoma skin lesions. Br. J. Dermatol. 2016, 175, 1251–1262. [Google Scholar] [CrossRef]
- Miyake, M.; Lawton, A.; Goodison, S.; Urquidi, V.; Rosser, C.J. Chemokine (CXC motif) ligand 1 (CXCL1) protein expression is increased in high-grade prostate cancer. Pathol. Res. Pract. 2014, 210, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Lawton, A.; Goodison, S.; Urquidi, V.; Gomes-Giacoia, E.; Zhang, G.; Ross, S.; Kim, J.; Rosser, C.J. Chemokine (CXC) ligand 1 (CXCL1) protein expression is increased in aggressive bladder cancers. BMC Cancer 2013, 13, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, M.; Furuya, H.; Onishi, S.; Hokutan, K.; Anai, S.; Chan, O.; Shi, S.; Fujimoto, K.; Goodison, S.; Cai, W. Monoclonal antibody against CXCL1 (HL2401) as a novel agent in suppressing IL6 expression and tumoral growth. Theranostics 2019, 9, 853. [Google Scholar] [CrossRef]
- Kalbasi, A.; Komar, C.; Tooker, G.M.; Liu, M.; Lee, J.W.; Gladney, W.L.; Ben-Josef, E.; Beatty, G.L. Tumor-derived CCL2 mediates resistance to radiotherapy in pancreatic ductal adenocarcinoma. Clin. Cancer Res. 2017, 23, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Bonapace, L.; Coissieux, M.-M.; Wyckoff, J.; Mertz, K.D.; Varga, Z.; Junt, T.; Bentires-Alj, M. Cessation of CCL2 inhibition accelerates breast cancer metastasis by promoting angiogenesis. Nature 2014, 515, 130–133. [Google Scholar] [CrossRef]
- Binder, P.S.; Cullinan, D.; Nywening, T.; Wilkinson-Ryan, I.; Belt, B.; Goedegebuure, P.; Powell, M.A.; Mutch, D.G.; Hawkins, W. CCR2 blockade alters the tumor microenvironment immune infiltrate and enhances anti-tumor activity in ovarian cancer. Gynecol. Oncol. 2017, 145, 36. [Google Scholar] [CrossRef]
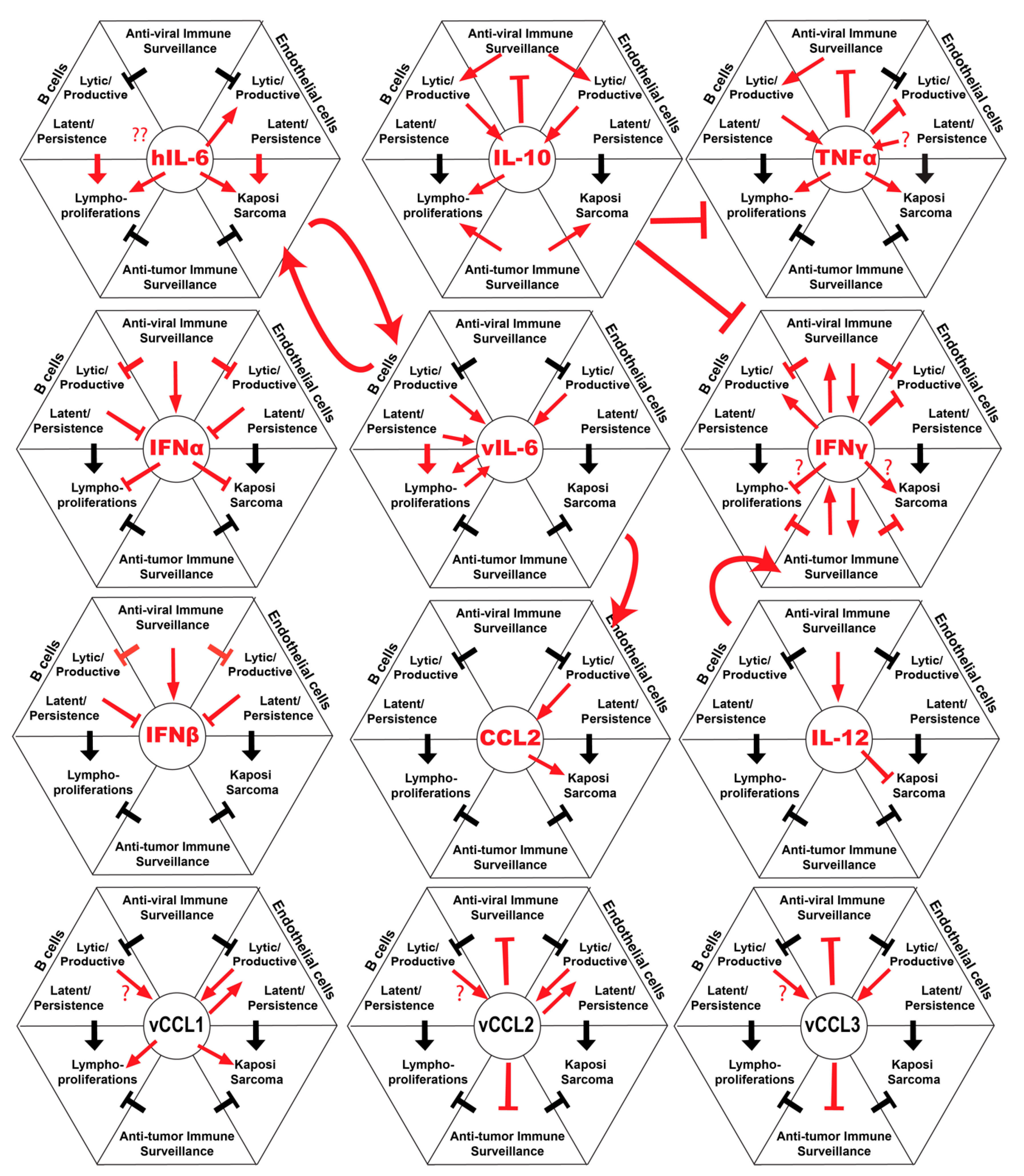
Figure 1.
Summary of host and viral cytokines and their interactions with Kaposi’s sarcoma-associated herpesvirus (KSHV) infection, immunity and tumor progression.
Figure 1.
Summary of host and viral cytokines and their interactions with Kaposi’s sarcoma-associated herpesvirus (KSHV) infection, immunity and tumor progression.


{kind=link}
Table 1.
Summary of the clinical trials referenced in this review.
| Cytokine | KSHV Disease | Drug | Dose | Study Type | Summary of the Result |
|---|---|---|---|---|---|
| IL-6 | MCD | Tocilizumab | 8 mg/kg MRA biweekly | Open-label trial in 28 patients for 16 weeks [43] | Alleviated the inflammatory symptoms and biochemical abnormalities |
| 8 mg/kg every 2 weeks | Ongoing Phase II clinical trial (NCT01441063). | ||||
| Tocilizumab Zidovudine Valganciclovir | 8 mg/kg IV on day 1 of 14-day cycles for a maximum of 6 cycles 600mg every 6 h 900mg every 12 h | Open label, single center pilot study [45] | One patient had complete clinical benefit response and four patients had partial clinical benefit responses on tocilizumab alone. Three patients who had AZT/VGC in combination with tocilizumab, two had partial clinical benefit responses and one had complete clinical benefit response | ||
| Siltuximab | Cohorts 1 to 5 enrolled in sequential order. 2-h infusion at 3 mg/kg every 2 weeks, 6 mg/kg every 2 weeks, 12 mg/kg every 3 weeks, 6 mg/kg weekly, and 12 mg/kg every 2 weeks, respectively. Cohort 6, 1-h infusion at 12 mg/kg every 3 weeks. Cohort 7, 9 mg/kg every 3 weeks. | Phase I, open-label, dose-finding and seven-cohort study in 23 patients [46] | 18 out of 23 patients had clinical benefits response and 12 patients showed objective tumor response. 11 patients treated with the highest dose of 12 mg/kg had clinical benefit response and 8 patients showed objective tumor response | ||
| 11 mg/kg intravenous infusion every 3 weeks | Randomized, double-blind, placebo-controlled trial in 79 patients [47]. | 18 of 53 had tumor and symptomatic responses | |||
| IFNa | AIDS-KS | IFNa | Starting dose 35×106 Reduced to 5–10×106 | Open therapeutic trial [92] | Anti-viral effects Anti-tumor response in 38% of patients |
| AIDS-KS | IFNa with didanosine | 1 million (low dose) or 10 million (intermediate dose) IU per day by subcutaneous (s.c.) injection. | randomized phase II clinical trials [93] | 40% tumor response | |
| TNFa | AIDS-KS | Thalidomide | 100 mg/day for 12 months | Phase II dose-escalation study [120] | Improved the clinical manifestation 8 out of 17 patients achieved partial response and 2 patients had stable conditions |
| AIDS-KS and KS | Pomalidomide | 5 mg/day for 21/28 days | phase I/II study in 22 KS patients with and without HIV infection. [121] | 16 patients. Achieved objective tumor response and 9 out of 15 HIV infected patients. Achieved objective response. | |
| IL-12 | AIDS-KS | L12 | 100, 300, 500 and 625 ng/kg | Phase 1 pilot study [129] | 17 had a complete or partial KS tumor response (61%) with three highest does. |
| AIDS-KS | IL12 alone and in combination with pegylated liposomal | 300 ng/kg subcutaneously twice weekly for six 3-week cycles, followed by 500 ng/kg subcutaneous IL-12 twice weekly | Phase II clinical trial [130] | 83.3% complete or partial KS tumor response. | |
| Advanced KS | NHS-IL12 | Ongoing Phase I/II clinical trial (NCT04303117) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alomari, N.; Totonchy, J. Cytokine-Targeted Therapeutics for KSHV-Associated Disease. Viruses 2020, 12, 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101097
AMA Style
Alomari N, Totonchy J. Cytokine-Targeted Therapeutics for KSHV-Associated Disease. Viruses. 2020; 12(10):1097. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101097
Chicago/Turabian StyleAlomari, Nedaa, and Jennifer Totonchy. 2020. "Cytokine-Targeted Therapeutics for KSHV-Associated Disease" Viruses 12, no. 10: 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101097
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.