
Evaluation of Merkel Cell Polyomavirus DNA in Tissue Samples from Italian Patients with Diagnosis of MCC

 and
and
Abstract
:1. Introduction
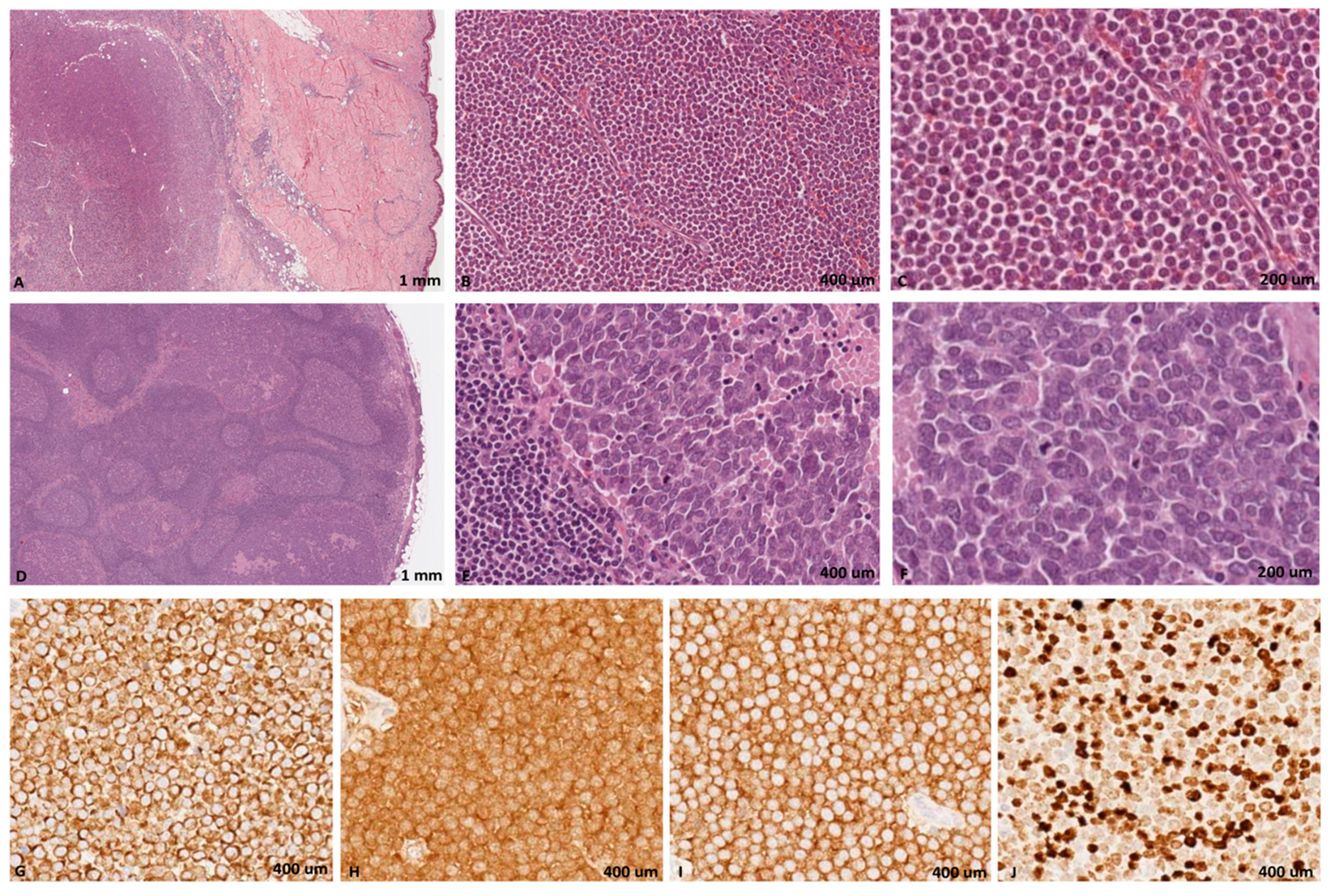
2. Materials and Methods
2.1. Clinical Specimens
2.2. DNA Extraction
2.3. Real-Time Polymerase Chain Reaction (qPCR)
2.4. MCPyV Nested PCR
2.5. MCPyV Phylogenetic Analysis
2.6. Statistical Analysis
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Approval
References
- Pietropaolo, V.; Prezioso, C.; Moens, U. Merkel Cell Polyomavirus and Merkel Cell Carcinoma. Cancers 2020, 12, 1774. [Google Scholar] [CrossRef]
- Liu, W.; You, J. Molecular Mechanisms of Merkel Cell Polyomavirus Transformation and Replication. Annu. Rev. Virol. 2020, 7, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Pastrana, D.V.; Tolstov, Y.L.; Becker, J.C.; Moore, P.S.; Chang, Y.; Buck, C.B. Quantitation of human seroresponsiveness to Merkel cell polyomavirus. PLoS Pathog. 2009, 5, e1000578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Moore, P.S. Merkel cell carcinoma: A virus-induced human cancer. Annu. Rev. Pathol. 2012, 7, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulitzer, M.P.; Amin, B.D.; Busam, K.J. Merkel cell carcinoma: Review. Adv. Anat. Pathol. 2009, 16, 135–144. [Google Scholar] [CrossRef]
- Harms, P.W.; Collie, A.M.; Hovelson, D.H.; Cani, A.K.; Verhaegen, M.E.; Patel, R.M.; Fullen, D.R.; Omata, K.; Dlugosz, A.A.; Tomlins, S.A.; et al. Next generation sequencing of Cytokeratin 20-negative Merkel cell carcinoma reveals ultraviolet-signature mutations and recurrent TP53 and RB1 inactivation. Mod. Pathol. 2016, 29, 240–248. [Google Scholar] [CrossRef]
- Harms, P.W.; Vats, P.; Verhaegen, M.E.; Robinson, D.R.; Wu, Y.M.; Dhanasekaran, S.M.; Palanisamy, N.; Siddiqui, J.; Cao, X.; Su, F.; et al. The Distinctive Mutational Spectra of Polyomavirus-Negative Merkel Cell Carcinoma. Cancer Res. 2015, 75, 3720–3727. [Google Scholar] [CrossRef] [Green Version]
- González-Vela, M.D.C.; Curiel-Olmo, S.; Derdak, S.; Beltran, S.; Santibañez, M.; Martínez, N.; Castillo-Trujillo, A.; Gut, M.; Sánchez-Pacheco, R.; Almaraz, C.; et al. Shared Oncogenic Pathways Implicated in Both Virus-Positive and UV-Induced Merkel Cell Carcinomas. J. Investig. Dermatol. 2017, 137, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Knepper, T.C.; Montesion, M.; Russell, J.S.; Sokol, E.S.; Frampton, G.M.; Miller, V.A.; Albacker, L.A.; McLeod, H.L.; Eroglu, Z.; Khushalani, N.I.; et al. The Genomic Landscape of Merkel Cell Carcinoma and Clinicogenomic Biomarkers of Response to Immune Checkpoint Inhibitor Therapy. Clin. Cancer Res. 2019, 25, 5961–5971. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.C.; Stang, A.; DeCaprio, J.A.; Cerroni, L.; Lebbé, C.; Veness, M.; Nghiem, P. Merkel cell carcinoma. Nat. Rev. Dis. Primers 2017, 3, 17077. [Google Scholar] [CrossRef] [PubMed]
- Pulitzer, M. Merkel Cell Carcinoma. Surg. Pathol. Clin. 2017, 10, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Sihto, H.; Kukko, H.; Koljonen, V.; Sankila, R.; Böhling, T.; Joensuu, H. Clinical factors associated with Merkel cell polyomavirus infection in Merkel cell carcinoma. J. Natl. Cancer Inst. 2009, 101, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Saiki, R.K.; Bugawan, T.L.; Horn, G.T.; Mullis, K.B.; Erlich, H.A. Analysis of enzymatically amplified beta-globin and HLA-DQ alpha DNA with allele-specific oligonucleotide probes. Nature 1986, 324, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Rodig, S.J.; Cheng, J.; Wardzala, J.; DoRosario, A.; Scanlon, J.J.; Laga, A.C.; Martinez-Fernandez, A.; Barletta, J.A.; Bellizzi, A.M.; Sadasivam, S.; et al. Improved detection suggests all Merkel cell carcinomas harbour Merkel polyomavirus. J. Clin. Investig. 2012, 122, 4645–4653. [Google Scholar] [CrossRef] [Green Version]
- Prezioso, C.; Obregon, F.; Ambroselli, D.; Petrolo, S.; Checconi, P.; Rodio, D.M.; Coppola, L.; Nardi, A.; Vito, C.; Sarmati, L.; et al. Merkel Cell Polyomavirus (MCPyV) in the Context of Immunosuppression: Genetic Analysis of Noncoding Control Region (NCCR) Variability among a HIV-1-Positive Population. Viruses 2020, 12, 507. [Google Scholar] [CrossRef]
- Prezioso, C.; Bianchi, M.; Obregon, F.; Ciotti, M.; Sarmati, L.; Andreoni, M.; Palamara, A.T.; Pascarella, S.; Moens, U.; Pietropaolo, V. Structural Analysis of Merkel Cell Polyomavirus (MCPyV) Viral Capsid Protein 1 (VP1) in HIV-1 Infected Individuals. Int. J. Mol. Sci. 2020, 21, 7998. [Google Scholar] [CrossRef]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Paik, J.Y.; Hall, G.; Clarkson, A.; Lee, L.; Toon, C.; Colebatch, A.; Chou, A.; Gill, A.J. Immunohistochemistry for Merkel cell polyomavirus is highly specific but not sensitive for the diagnosis of Merkel cell carcinoma in the Australian population. Hum. Pathol. 2011, 42, 1385–1390. [Google Scholar] [CrossRef]
- Andea, A.A.; Patel, R.; Ponnazhagan, S.; Isayeva, T.; Kumar, S.; Siegal, G.P. Detection of Merkel cell polyomavirus in formalin-fixed, paraffin-embedded tissue of Merkel cell carcinoma and correlation with prognosis. Rom. J. Morphol. Embryol. 2014, 55, 1057–1062. [Google Scholar]
- Stang, A.; Becker, J.C.; Nghiem, P.; Ferlay, J. The association between geographic location and incidence of Merkel cell carcinoma in comparison to melanoma: An international assessment. Eur. J. Cancer 2018, 94, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.J.; Zhang, Z.F.; Coit, D.G. Surgical management of Merkel cell carcinoma. Ann. Surg. 1999, 229, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Hashida, Y.; Higuchi, T.; Matsui, K.; Shibata, Y.; Nakajima, K.; Sano, S.; Daibata, M. Genetic variability of the noncoding control region of cutaneous Merkel cell polyomavirus: Identification of geographically related genotypes. J. Infect. Dis. 2018, 217, 1601–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, J.C.; Houben, R.; Ugurel, S.; Trefzer, U.; Pföhler, C.; Schrama, D. MC polyomavirus is frequently present in Merkel cell carcinoma of European patients. J. Investig. Dermatol. 2009, 129, 248–250. [Google Scholar] [CrossRef] [Green Version]
- Erovic, B.M.; Al Habeeb, A.; Harris, L.; Goldstein, D.P.; Ghazarian, D.; Irish, J.C. Significant overexpression of the Merkel cell polyomavirus (MCPyV) large T antigen in Merkel cell carcinoma. Head Neck 2013, 35, 184–189. [Google Scholar] [CrossRef]
- Haymerle, G.; Janik, S.; Fochtmann, A.; Pammer, J.; Schachner, H.; Nemec, L.; Mildner, M.; Houben, R.; Grasl, M.C.; Erovic, B.M. Expression of Merkelcell polyomavirus (MCPyV) large T-antigen in Merkel cell carcinoma lymph node metastases predicts poor outcome. PLoS ONE 2017, 12, e0180426. [Google Scholar] [CrossRef] [Green Version]
- Houben, R.; Grimm, J.; Willmes, C.; Weinkam, R.; Becker, J.C.; Schrama, D. Merkel cell carcinoma and Merkel cell polyomavirus: Evidence for hit-and-run oncogenesis. J. Investig. Dermatol. 2012, 132, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Csoboz, B.; Rasheed, K.; Sveinbjørnsson, B.; Moens, U. Merkel cell polyomavirus and non-Merkel cell carcinomas: Guilty or circumstantial evidence? APMIS 2020, 128, 104–120. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| MCCPL Case No. | Age (years) | Gender | Site | qPCR Results |
|---|---|---|---|---|
| MCCPL 1 | 77 | F | Skin | 35 × 10−1 copies |
| MCCPL 2 | 82 | M | forearm | 70 × 10−1 copies |
| MCCPL 3 | 93 | F | shoulder | 35 × 10−1 copies |
| MCCPL 4 | 93 | F | shoulder | 100 × 10−1 copies |
| MCCPL 5 | 96 | F | lower limb | 92 × 10−1 copies |
| MCCPL 6 | 80 | M | knee | 92 × 10−1 copies |
| MCCPL 7 | 76 | F | forearm | 95 × 10−1 copies |
| MCCPL 8 | 75 | M | shinbone | 75 × 10−1 copies |
| MCCPL 9 | 79 | M | H&N parotid | 95 × 10−1 copies |
| MCCPL 10 | 92 | F | H&N larynx | 33 × 10−1 copies |
| MCCPL 11 | 74 | F | H&N eyelid | 82 × 10−1 copies |
| MCCPL 12 | 79 | F | H&N superciliar skin | 18 × 10−1 copies |
| MCCPL 13 | 82 | M | H&N ear | 87 × 10−1 copies |
| MCCPL 14 | 79 | F | shoulder | NEGATIVE |
| MCCPL 15 | 76 | F | ellipse of skin and subcute arm | NEGATIVE |
| MCCPL 16 | 86 | F | skin | NEGATIVE |
| MCCPL 17 | 84 | F | arm | NEGATIVE |
| MCCML Case No. | Age (years) | Gender | Site | qPCR Results |
| MCCML 18 | 77 | F | Inguinal lymph node | NEGATIVE |
| MCCML 19 | 82 | M | back | NEGATIVE |
| MCCML 20 | 79 | M | lymph node | NEGATIVE |
| MCCML 21 | 79 | M | lower limb | NEGATIVE |
| MCCML 22 | 79 | M | hypocondrium | NEGATIVE |
| MCCML 23 | 74 | M | inguinal lymph node | NEGATIVE |
| MCCML 24 | 93 | M | inguinal lymph node | NEGATIVE |
| MCCML 25 | 93 | M | axillar lymphectomy | NEGATIVE |
| MCCML 26 | 85 | F | inguinal lymph node | NEGATIVE |
| MCC Case No. | VP1 Sequencing | NCCR Sequencing | Genotype |
|---|---|---|---|
| MCCPL 1 | 4192 TΔ | 5210–5211 GTTGA ins. | IIa-2 strain |
| MCCPL 2 | 4179 CΔ | 5220 T to C transition | not applicable |
| MCCPL 3 | no modification | no modification | not applicable |
| MCCPL 4 | 4204 T to C transition | no modification | not applicable |
| MCCPL 5 | no modification | no modification | not applicable |
| MCCPL 6 | no modification | 5104 G to T transversion | not applicable |
| MCCPL 7 | no modification | no modification | not applicable |
| MCCPL 8 | no modification | no modification | not applicable |
| MCCPL 9 | no modification | 5148 T to C transition; 5210–5211 GTTGA ins. | IIa-2 strain |
| MCCPL 10 | 4324 A to T transversions | no modification | not applicable |
| MCCPL 11 | no modification | no modification | not applicable |
| MCCPL 12 | no modification | no modification | not applicable |
| MCCPL 13 | no modification | 5176 A to T transversions | not applicable |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prezioso, C.; Carletti, R.; Obregon, F.; Piacentini, F.; Manicone, A.M.; Soda, G.; Moens, U.; Di Gioia, C.; Pietropaolo, V. Evaluation of Merkel Cell Polyomavirus DNA in Tissue Samples from Italian Patients with Diagnosis of MCC. Viruses 2021, 13, 61. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010061
Prezioso C, Carletti R, Obregon F, Piacentini F, Manicone AM, Soda G, Moens U, Di Gioia C, Pietropaolo V. Evaluation of Merkel Cell Polyomavirus DNA in Tissue Samples from Italian Patients with Diagnosis of MCC. Viruses. 2021; 13(1):61. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010061
Chicago/Turabian StylePrezioso, Carla, Raffaella Carletti, Francisco Obregon, Francesca Piacentini, Anna Maria Manicone, Giuseppe Soda, Ugo Moens, Cira Di Gioia, and Valeria Pietropaolo. 2021. "Evaluation of Merkel Cell Polyomavirus DNA in Tissue Samples from Italian Patients with Diagnosis of MCC" Viruses 13, no. 1: 61. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010061