
Seroprevalence of Infection with Feline Morbilliviruses Is Associated with FLUTD and Increased Blood Creatinine Concentrations in Domestic Cats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Collection
2.2. Virus Stock Preparation
2.3. Immunofluorescence Assay
2.4. Blood Creatinine Determination
2.5. Statistical Analysis
3. Results
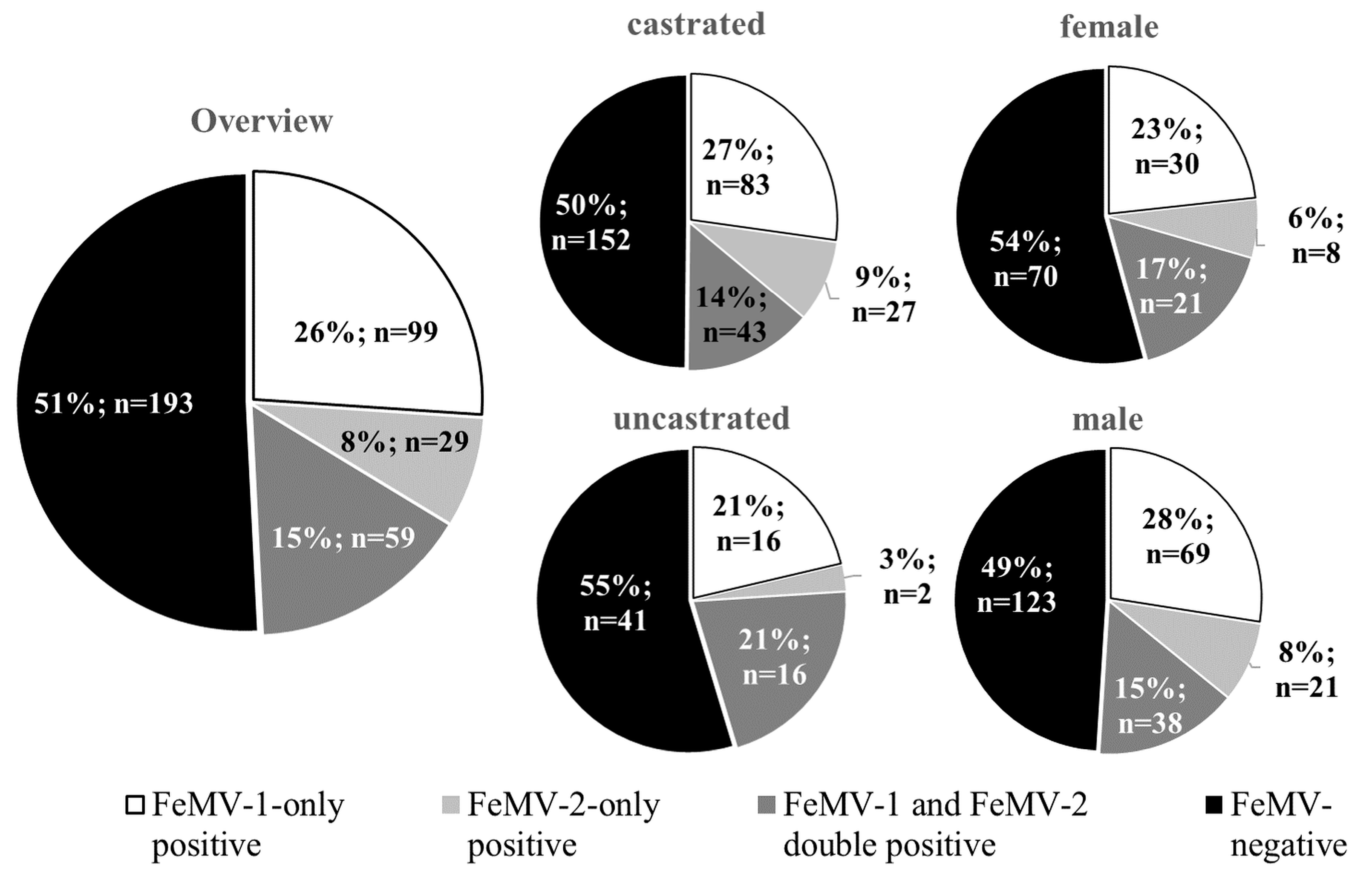
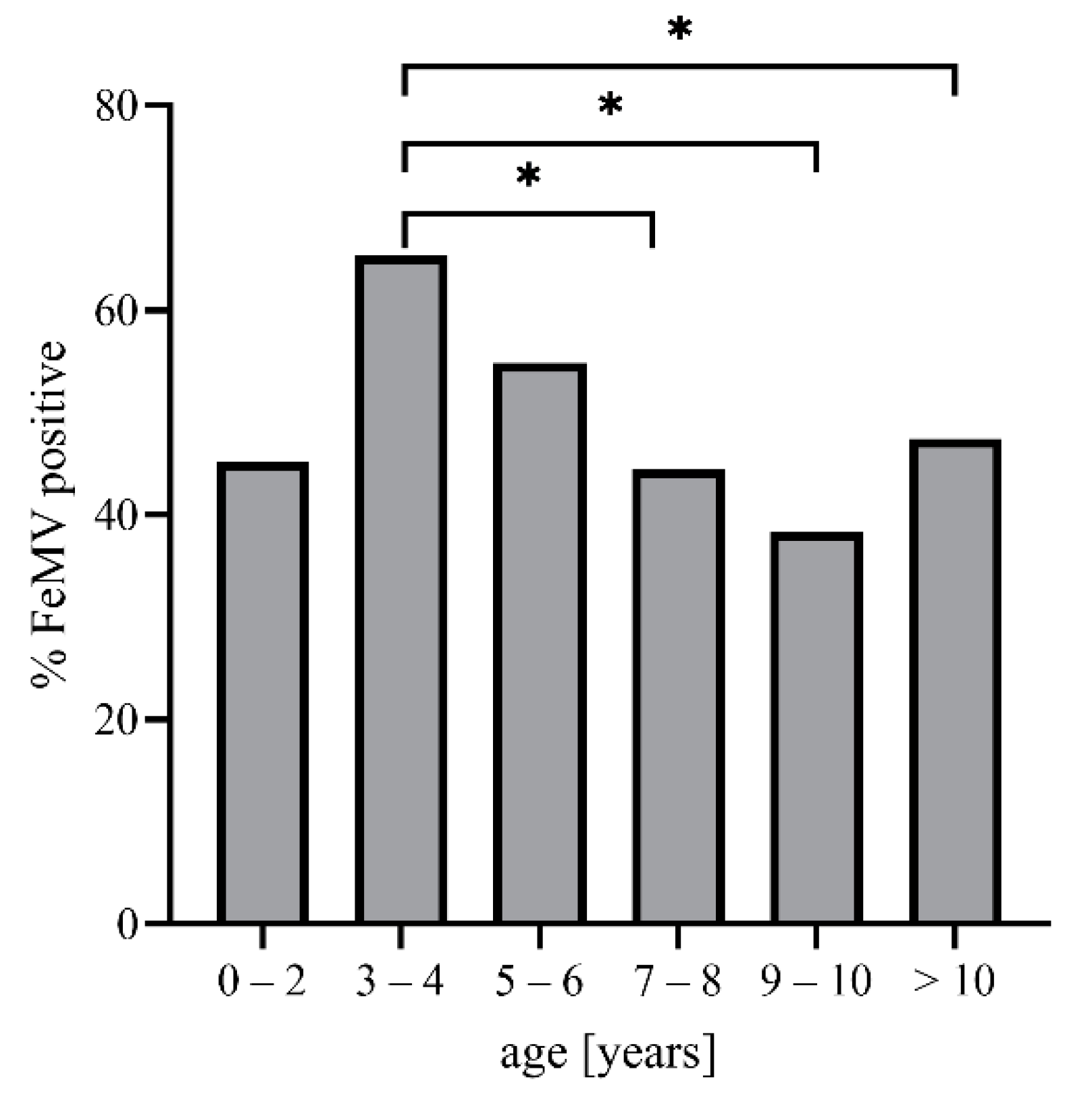
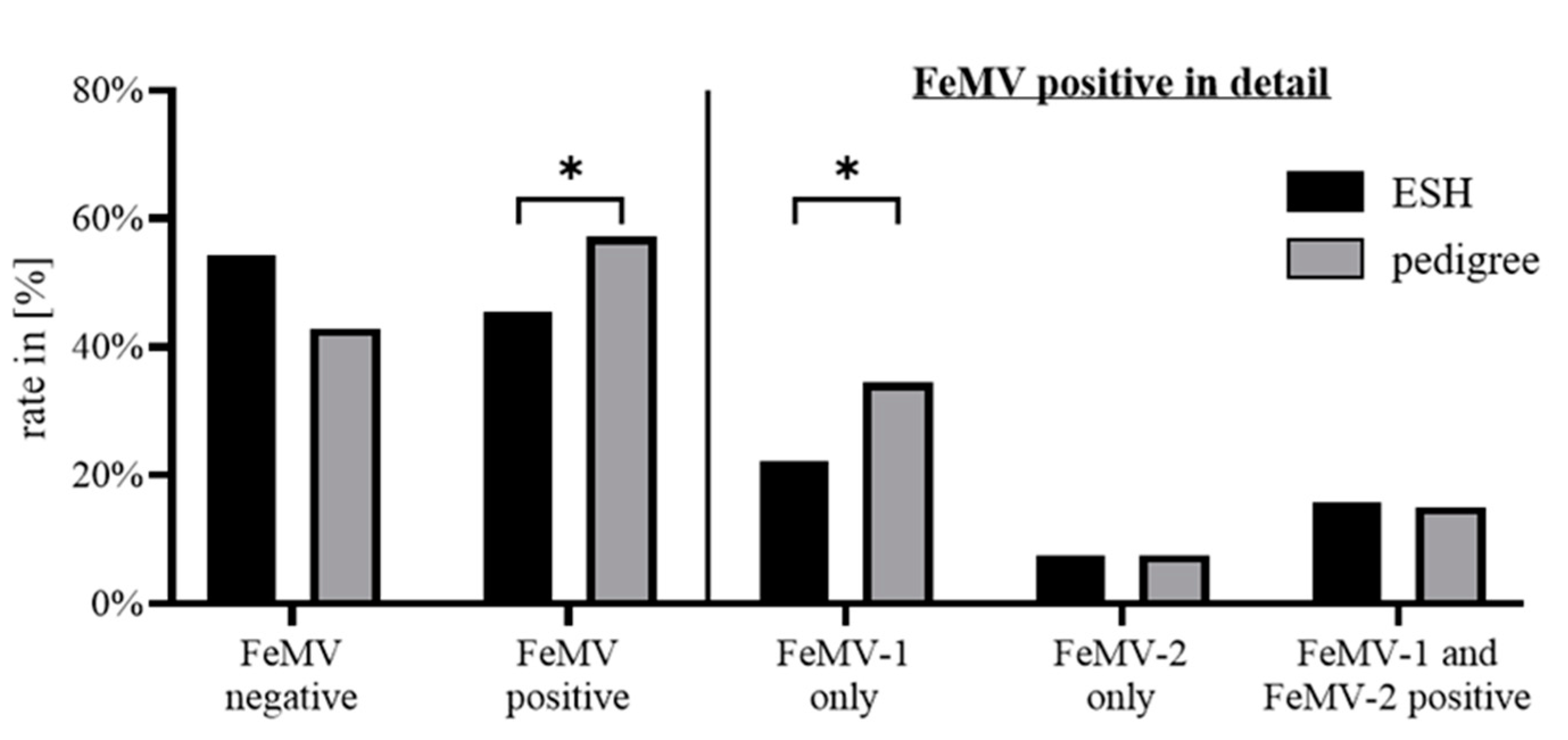
3.1. Study Population
3.2. FeMV Seroprevalence
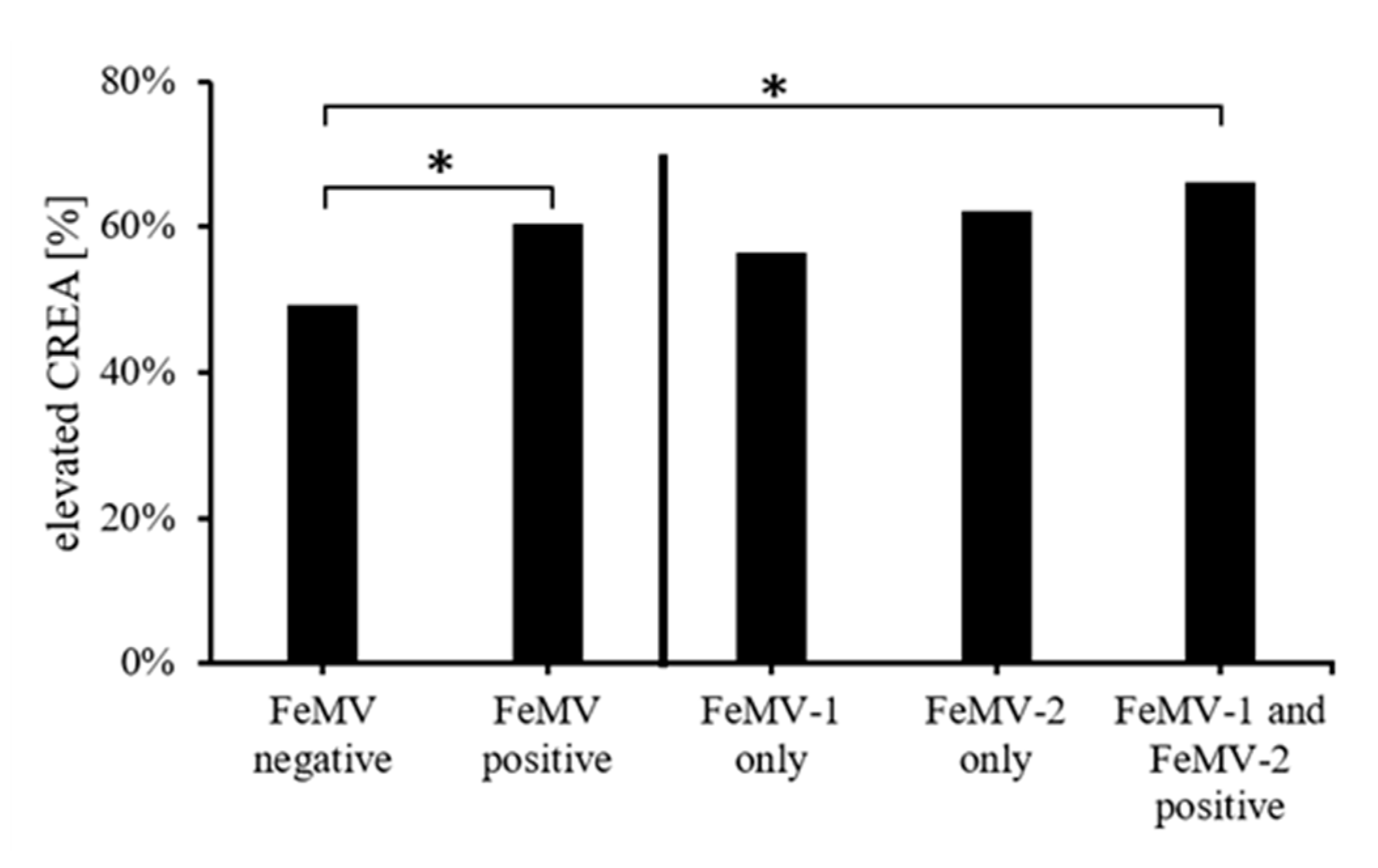
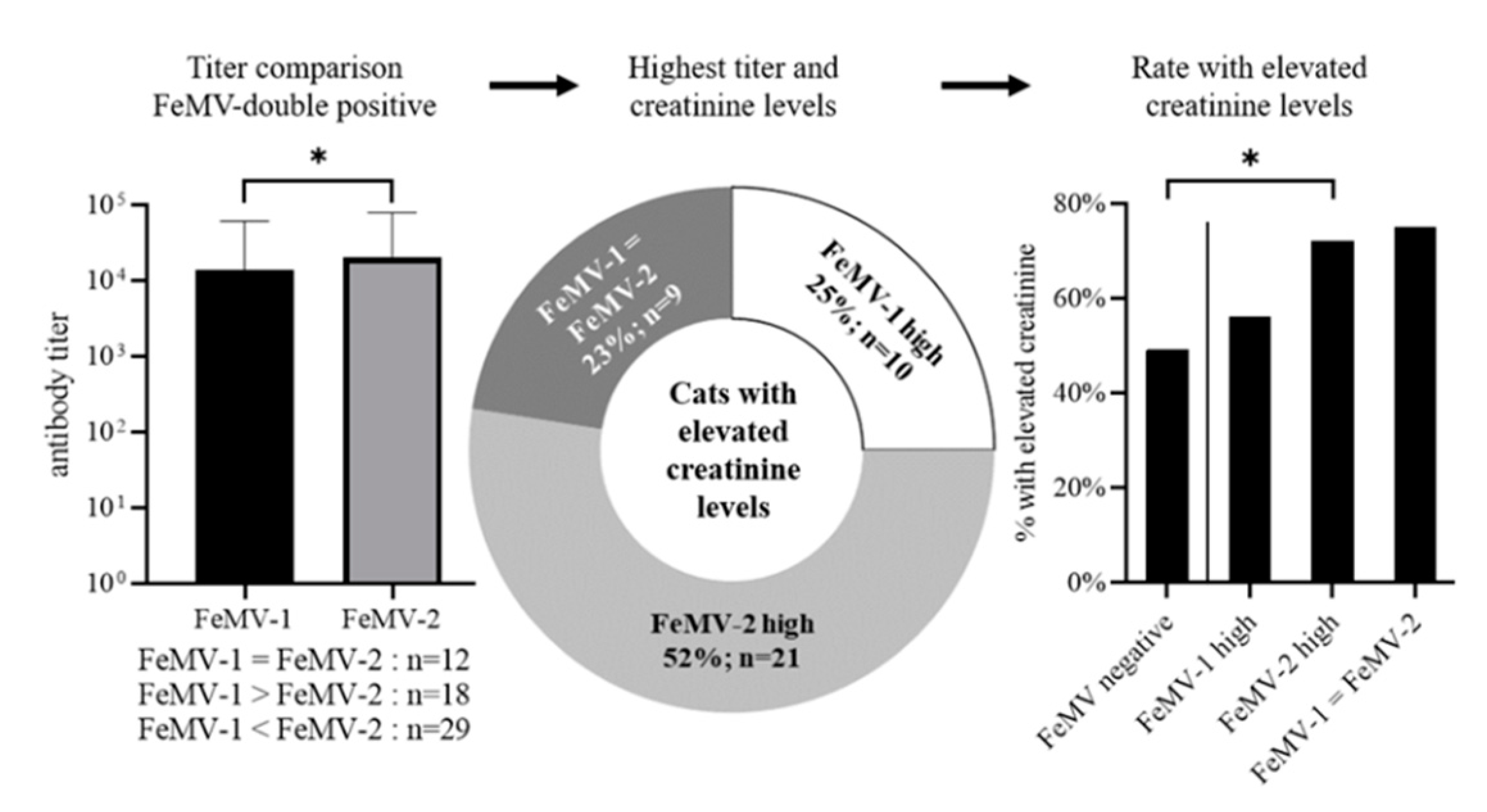
3.3. Correlation of Blood Creatinine Values with FeMV-Antibody Responses
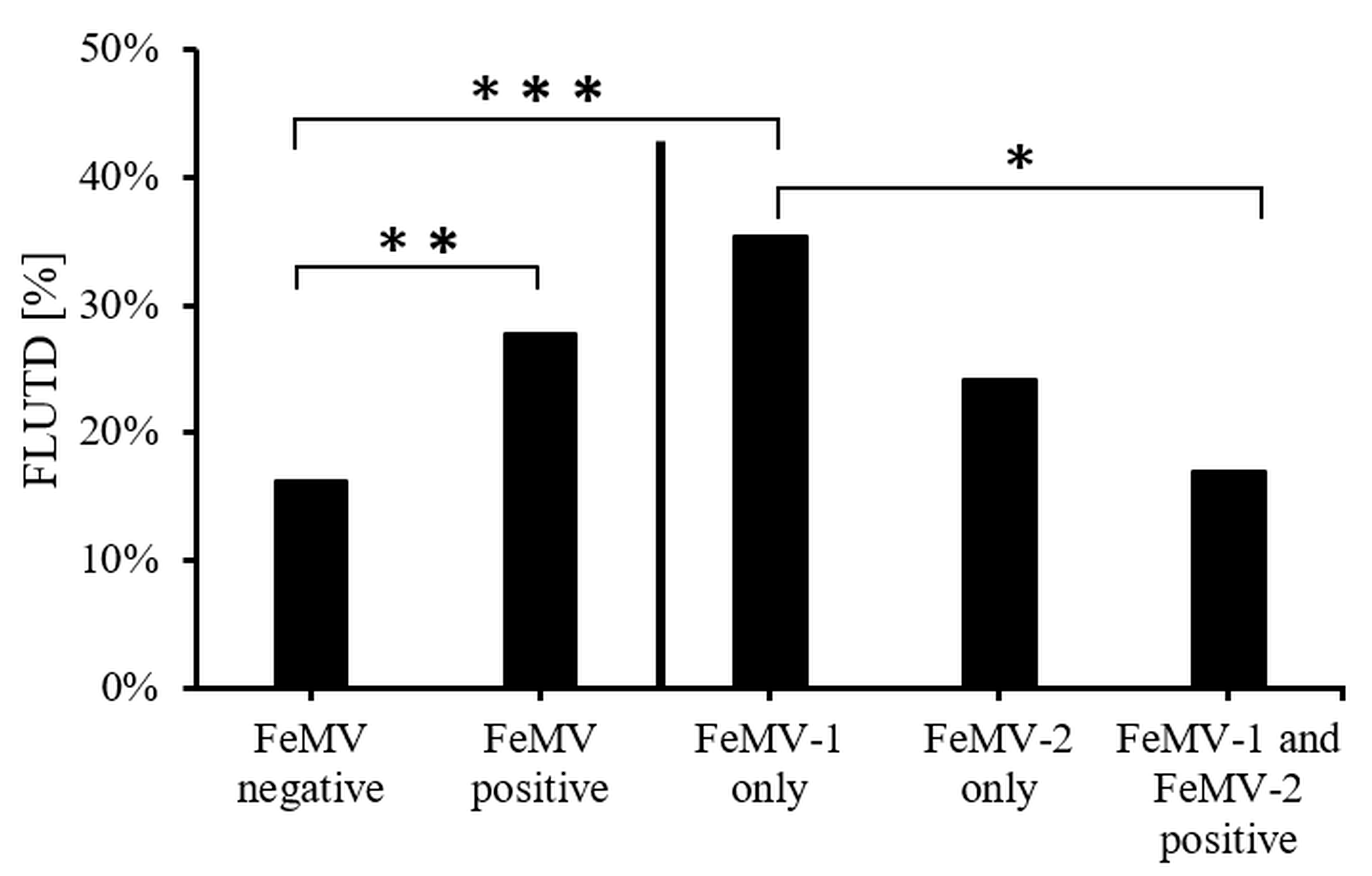
3.4. Association of FeMV-Antibody Response with Clinical Diagnoses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statista. Households with Pets by Type in Germany 2019|Statista. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/552974/households-with-pets-by-type-in-germany/ (accessed on 14 January 2021).
- Statista. Pets: Number by Type Germany 2019|Statista. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/552971/pets-number-by-type-germany/ (accessed on 14 January 2021).
- Marino, C.L.; Lascelles, B.D.X.; Vaden, S.L.; Gruen, M.E.; Marks, S.L. Prevalence and classification of chronic kidney disease in cats randomly selected from four age groups and in cats recruited for degenerative joint disease studies. J. Feline Med. Surg. 2014, 16, 465–472. [Google Scholar] [CrossRef] [Green Version]
- DiBartola, S.P.; Rutgers, H.C.; Zack, P.M.; Tarr, M.J. Clinicopathologic findings associated with chronic renal disease in cats: 74 cases (1973-1984). J. Am. Veter. Med. Assoc. 1987, 190, 1196–1202. [Google Scholar]
- Chakrabarti, S.; Syme, H.M.; Brown, C.A.; Elliott, J. Histomorphometry of Feline Chronic Kidney Disease and Correlation with Markers of Renal Dysfunction. Veter. Pathol. 2012, 50, 147–155. [Google Scholar] [CrossRef]
- Chen, H.; Dunaevich, A.; Apfelbaum, N.; Kuzi, S.; Mazaki-Tovi, M.; Aroch, I.; Segev, G. Acute on chronic kidney disease in cats: Etiology, clinical and clinicopathologic findings, prognostic markers, and outcome. J. Veter. Intern. Med. 2020, 34, 1496–1506. [Google Scholar] [CrossRef] [PubMed]
- Bartges, J.W. Chronic Kidney Disease in Dogs and Cats. Veter. Clin. North Am. Small Anim. Pr. 2012, 42, 669–692. [Google Scholar] [CrossRef]
- E Lees, G. Early diagnosis of renal disease and renal failure. Veter. Clin. North Am. Small Anim. Pr. 2004, 34, 867–885. [Google Scholar] [CrossRef]
- Pittari, J.; Rodan, I.; Beekman, G.; Gunn-Moore, D.; Polzin, D.; Taboada, J.; Tuzio, H.; Zoran, D. American Association of Feline Practitioners: Senior Care Guidelines. J. Feline Med. Surg. 2009, 11, 763–778. [Google Scholar] [CrossRef] [Green Version]
- Elliott, J.; Rawlings, J.M.; Markwell, P.J.; Barber, P.J. Survival of cats with naturally occurring chronic renal failure: Effect of dietary management. J. Small Anim. Pr. 2000, 41, 235–242. [Google Scholar] [CrossRef]
- Hartmann, K.; Pennisi, M.G.; Dorsch, R. Infectious Agents in Feline Chronic Kidney Disease. Adv. Small Anim. Care 2020, 1, 189–206. [Google Scholar] [CrossRef]
- Forrester, S.D.; Roudebush, P. Evidence-Based Management of Feline Lower Urinary Tract Disease. Veter. Clin. North Am. Small Anim. Pr. 2007, 37, 533–558. [Google Scholar] [CrossRef]
- Hostutler, R.A.; Chew, D.J.; DiBartola, S.P. Recent concepts in feline lower urinary tract disease. Veter. Clin. North Am. Small Anim. Pr. 2005, 35, 147–170. [Google Scholar] [CrossRef]
- Buffington, C.A.T. External and internal influences on disease risk in cats. J. Am. Veter. Med Assoc. 2002, 220, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.C.; Kruger, J.M.; Venta, P.J.; Vilnis, A.; Maas, K.A.; Dulin, J.A.; Maes, R.K. Genetic characterization of 2 novel feline caliciviruses isolated from cats with idiopathic lower urinary tract disease. J. Vet. Intern. Med. 2002, 16, 293–302. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Wong, B.H.L.; Fan, R.Y.Y.; Wong, A.Y.P.; Zhang, A.J.X.; Wu, Y.; Choi, G.K.Y.; Li, K.S.M.; Hui, J.; et al. Feline morbillivirus, a previously undescribed paramyxovirus associated with tubulointerstitial nephritis in domestic cats. Proc. Natl. Acad. Sci. USA 2012, 109, 5435–5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutummaporn, K.; Suzuki, K.; Machida, N.; Mizutani, T.; Park, E.-S.; Morikawa, S.; Furuya, T.; Shigeru, M. Association of feline morbillivirus infection with defined pathological changes in cat kidney tissues. Veter. Microbiol. 2019, 228, 12–19. [Google Scholar] [CrossRef]
- McCallum, K.E.; Stubbs, S.; Hope, N.; Mickleburgh, I.; Dight, D.; Tiley, L.; Williams, T.L. Detection and seroprevalence of morbillivirus and other paramyxoviruses in geriatric cats with and without evidence of azotemic chronic kidney disease. J. Veter. Intern. Med. 2018, 32, 1100–1108. [Google Scholar] [CrossRef]
- Isa, N.H.M.; Selvarajah, G.T.; Khor, K.H.; Tan, S.W.; Manoraj, H.; Omar, N.H.; Omar, A.R.; Mustaffa-Kamal, F. Molecular detection and characterisation of feline morbillivirus in domestic cats in Malaysia. Veter. Microbiol. 2019, 236, 108382. [Google Scholar] [CrossRef]
- Furuya, T.; Sassa, Y.; Omatsu, T.; Nagai, M.; Fukushima, R.; Shibutani, M.; Yamaguchi, T.; Uematsu, Y.; Shirota, K.; Mizutani, T. Existence of feline morbillivirus infection in Japanese cat populations. Arch. Virol. 2014, 159, 371–373. [Google Scholar] [CrossRef]
- Sieg, M.; Heenemann, K.; Rückner, A.; A Burgener, I.; Oechtering, G.; Vahlenkamp, T.W. Discovery of new feline paramyxoviruses in domestic cats with chronic kidney disease. Virus Genes 2015, 51, 294–297. [Google Scholar] [CrossRef]
- Lorusso, A.; Di Tommaso, M.; Di Felice, E.; Zaccaria, G.; Luciani, A.; Marcacci, M.; Aste, G.; Boari, A.; Savini, G. First report of feline morbillivirus in Europe. Vet. Ital. 2015, 51, 235–237. [Google Scholar] [PubMed]
- De Luca, E.; Crisi, P.E.; Di Domenico, M.; Malatesta, D.; Vincifori, G.; Di Tommaso, M.; Di Guardo, G.; Di Francesco, G.; Petrini, A.; Savini, G.; et al. A real-time RT-PCR assay for molecular identification and quantitation of feline morbillivirus RNA from biological specimens. J. Virol. Methods 2018, 258, 24–28. [Google Scholar] [CrossRef]
- De Luca, E.; Crisi, P.E.; Marcacci, M.; Malatesta, D.; Di Sabatino, D.; Cito, F.; D’Alterio, N.; Puglia, I.; Berjaoui, S.; Colaianni, M.L.; et al. Epidemiology, pathological aspects and genome heterogeneity of feline morbillivirus in Italy. Veter. Microbiol. 2020, 240, 108484. [Google Scholar] [CrossRef]
- Muratore, E.; Cerutti, F.; Colombino, E.; Biasibetti, E.; Caruso, C.; Brovida, C.; Cavana, P.; Poncino, L.; Caputo, M.P.; Peletto, S.; et al. Feline morbillivirus in northwestern Italy: First detection of genotype 1-B. J. Feline Med. Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Donato, G.; De Luca, E.; Crisi, P.E.; Pizzurro, F.; Masucci, M.; Marcacci, M.; Cito, F.; Di Sabatino, D.; Boari, A.; D’Alterio, N.; et al. Isolation and genome sequences of two Feline Morbillivirus genotype 1 strains from Italy. Vet. Ital. 2019, 55, 179–182. [Google Scholar] [PubMed]
- Sharp, C.R.; Nambulli, S.; Acciardo, A.S.; Rennick, L.J.; Drexler, J.F.; Rima, B.K.; Williams, T.; Duprex, W.P. Chronic Infection of Domestic Cats with Feline Morbillivirus, United States. Emerg. Infect. Dis. 2016, 22, 760–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darold, G.M.; Alfieri, A.A.; Muraro, L.S.; Amude, A.M.; Zanatta, R.; Yamauchi, K.C.I.; Alfieri, A.F.; Lunardi, M. First report of feline morbillivirus in South America. Arch. Virol. 2017, 162, 469–475. [Google Scholar] [CrossRef]
- Darold, G.M.; Alfieri, A.A.; Araújo, J.P.; Da Cruz, T.F.; Bertti, K.M.D.L.B.; Da Silva, G.C.P.; Amude, A.M.; Muraro, L.S.; Lavorente, F.L.P.; Lunardi, M. High genetic diversity of paramyxoviruses infecting domestic cats in Western Brazil. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Balbo, L.C.; Fritzen, J.T.T.; Lorenzetti, E.; Medeiros, T.N.S.; Jardim, A.M.; Alfieri, A.A.; Alfieri, A.F. Molecular characterization of Feline paramyxovirus and Feline morbillivirus in cats from Brazil. Braz. J. Microbiol. 2021, 1–5. [Google Scholar] [CrossRef]
- Yilmaz, H.; Tekelioglu, B.K.; Gurel, A.; E Bamac, O.; Ozturk, G.Y.; Cizmecigil, U.Y.; Altan, E.; Aydin, O.; Yilmaz, A.; Berriatua, E.; et al. Frequency, clinicopathological features and phylogenetic analysis of feline morbillivirus in cats in Istanbul, Turkey. J. Feline Med. Surg. 2017, 19, 1206–1214. [Google Scholar] [CrossRef]
- Ou, J.; Ye, S.; Xu, H.; Zhao, J.; Ren, Z.; Lu, G.; Li, S. First report of feline morbillivirus in mainland China. Arch. Virol. 2020, 165, 1837–1841. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Nakagawa, S.; Yoshikawa, R.; Kuwahara, C.; Hagiwara, H.; Asai, K.-I.; Kawakami, K.; Yamamoto, Y.; Ogawa, M.; Miyazawa, T. Genetic diversity of feline morbilliviruses isolated in Japan. J. Gen. Virol. 2014, 95, 1464–1468. [Google Scholar] [CrossRef]
- Park, E.-S.; Suzuki, M.; Kimura, M.; Mizutani, H.; Saito, R.; Kubota, N.; Hasuike, Y.; Okajima, J.; Kasai, H.; Sato, Y.; et al. Epidemiological and pathological study of feline morbillivirus infection in domestic cats in Japan. BMC Veter. Res. 2016, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sieg, M.; Busch, J.; Eschke, M.; Böttcher, D.; Heenemann, K.; Vahlenkamp, A.; Reinert, A.; Seeger, J.; Heilmann, R.; Scheffler, K.; et al. A New Genotype of Feline Morbillivirus Infects Primary Cells of the Lung, Kidney, Brain and Peripheral Blood. Viruses 2019, 11, 146. [Google Scholar] [CrossRef] [Green Version]
- Arikawa, K.; Wachi, A.; Imura, Y.; Sutummaporn, K.; Kai, C.; Park, E.-S.; Morikawa, S.; Uematsu, Y.; Yamaguchi, T.; Furuya, T. Development of an ELISA for serological detection of feline morbillivirus infection. Arch. Virol. 2017, 162, 2421–2425. [Google Scholar] [CrossRef]
- Busch, J.; Sacristán, I.; Cevidanes, A.; Millán, J.; Vahlenkamp, T.W.; Napolitano, C.; Sieg, M. High seroprevalence of feline morbilliviruses in free-roaming domestic cats in Chile. Arch. Virol. 2020. [Google Scholar] [CrossRef]
- Sieg, M.; Vahlenkamp, A.; Baums, C.G.; Vahlenkamp, T.W. First Complete Genome Sequence of a Feline Morbillivirus Isolate from Germany. Genome Announc. 2018, 6, e00244-18. [Google Scholar] [CrossRef] [Green Version]
- Studer, N.; Lutz, H.; Saegerman, C.; Gönczi, E.; Meli, M.L.; Boo, G.; Hartmann, K.; Hosie, M.J.; Moestl, K.; Tasker, S.; et al. Pan-European Study on the Prevalence of the Feline Leukaemia Virus Infection–Reported by the European Advisory Board on Cat Diseases (ABCD Europe). Viruses 2019, 11, 993. [Google Scholar] [CrossRef] [Green Version]
- Klein-Richers, U.; Hartmann, K.; Hofmann-Lehmann, R.; Unterer, S.; Bergmann, M.; Rieger, A.; Leutenegger, C.; Pantchev, N.; Balzer, J.; Felten, S. Prevalence of Feline Coronavirus Shedding in German Catteries and Associated Risk Factors. Viruses 2020, 12, 1000. [Google Scholar] [CrossRef]
- Crisi, P.E.; Dondi, F.; De Luca, E.; Di Tommaso, M.; Vasylyeva, K.; Ferlizza, E.; Savini, G.; Luciani, A.; Malatesta, D.; Lorusso, A.; et al. Early Renal Involvement in Cats with Natural Feline Morbillivirus Infection. Animals 2020, 10, 828. [Google Scholar] [CrossRef]
- Stranieri, A.; Lauzi, S.; Dallari, A.; Gelain, M.E.; Bonsembiante, F.; Ferro, S.; Paltrinieri, S. Feline morbillivirus in Northern Italy: Prevalence in urine and kidneys with and without renal disease. Veter. Microbiol. 2019, 233, 133–139. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busch, J.; Heilmann, R.M.; Vahlenkamp, T.W.; Sieg, M. Seroprevalence of Infection with Feline Morbilliviruses Is Associated with FLUTD and Increased Blood Creatinine Concentrations in Domestic Cats. Viruses 2021, 13, 578. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040578
Busch J, Heilmann RM, Vahlenkamp TW, Sieg M. Seroprevalence of Infection with Feline Morbilliviruses Is Associated with FLUTD and Increased Blood Creatinine Concentrations in Domestic Cats. Viruses. 2021; 13(4):578. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040578
Chicago/Turabian StyleBusch, Johannes, Romy M. Heilmann, Thomas W. Vahlenkamp, and Michael Sieg. 2021. "Seroprevalence of Infection with Feline Morbilliviruses Is Associated with FLUTD and Increased Blood Creatinine Concentrations in Domestic Cats" Viruses 13, no. 4: 578. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040578