
Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes



Department of Food Science, University of Tennessee, Knoxville, TN 37996, USA
*
Author to whom correspondence should be addressed.
Viruses 2021, 13(4), 671; https://0-doi-org.brum.beds.ac.uk/10.3390/v13040671
Submission received: 1 January 2021
/
Revised: 26 March 2021
/
Accepted: 10 April 2021
/
Published: 14 April 2021
(This article belongs to the Special Issue Bacteriophages in Food Applications)
Abstract
:Listeria monocytogenes serotype 4b strains are the most prevalent clinical isolates and are widely found in food processing environments. Bacteriophages are natural viral predators of bacteria and are a promising biocontrol agent for L. monocytogenes. The aims of this study were to characterize phages that specifically infect serotype 4b strains and to assess their ability to inhibit the growth of serotype 4b strains. Out of 120 wild Listeria phages, nine phages were selected based on their strong lytic activity against the model serotype 4b strain F2365. These nine phages can be divided into two groups based on their morphological characteristics and host range. Comparison to previously characterized phage genomes revealed one of these groups qualifies to be defined as a novel species. Phages LP-020, LP-027, and LP-094 were selected as representatives of these two groups of phages for further characterization through one-step growth curve and inhibition of serotype 4b L. monocytogenes experiments. Listeria phages that target serotype 4b showed an inhibitory effect on the growth of F2365 and other serotype 4 strains and may be useful for biocontrol of L.monocytogenes in food processing environments.
1. Introduction
Listeria monocytogenes is a Gram-positive foodborne pathogen that infects humans and animals [1]. L. monocytogenes is widely isolated from soil, agriculture environments, and urban environments, and can tolerate high salt concentrations and a broad range of temperatures and pH levels [2,3,4,5]. Contamination of the food processing environment with L. monocytogenes can lead to consumers ingesting the pathogen, which can cause the potentially fatal invasive disease listeriosis [6]. The global burden of listeriosis cases has been estimated at 23,150 annually, with a mortality rate of 26% [7]. Within the United States alone, there is an estimated 2518 annual cases, with a 20% mortality rate [8,9], and economic losses caused by listeriosis were more than $3.1 billion in 2018 [10].
Bacteriophages are natural viral predators of bacteria, which infect and lyse specific host strains [11]. High specificity, self-replication capability, and tolerance of a wide range of temperatures and pHs [12] make bacteriophages a promising candidate for biocontrol of L. monocytogenes in the food processing environment [13]. Listeria phage biocontrol products have been approved for use by the United States Food and Drug Administration since 2006. These products are marketed to control L. monocytogenes contamination on food and in food processing plants. Currently used Listeria phages have been characterized as belonging to the genus Pecentumvirus [14]. Pecentumvirus phages have been shown to utilize rhamnose and N-acetylglucosamine of wall teichoic acids as binding receptors during the adsorption step of infection [15]. Presence or absence of these sugars corresponds to the various serotypes of L. monocytogenes [16]; thus, Listeria phages show some level of serotype specificity [17].
Based on cell surface antigenic determinants, L. monocytogenes can be divided into at least 13 serotypes. Serotype 4b strains account for most clinal isolates from humans, causing about 50% of illnesses; serotype 1/2a ranks second and is associated with 27% of cases [18,19,20,21]. Serotype 1/2a and 4b are the most frequently recovered from food and environmental samples [20,22,23,24]. Previous phage host-range studies employing efficiency of plaquing assays showed three Pecentumvirus phages, LP-048, LP-125, and A511, that effectively form plaques against serotype 1/2 strains and serotype 4b strain F2365 [25]; however, Homburgvirus LP-018, which also shows some potential use in biocontrol applications [26], was not able to infect this model strain [25]. To increase the diversity of characterized phages available for use against serotype 4 strains, we screened and characterized nine Listeria phages that show strong infectivity against the serotype 4b L. monocytogenes strain F2365.
2. Materials and Methods
2.1. Bacterial Strains and Bacteriophages
All bacterial strains in this study are listed in Table 1. L. monocytogenes MACK was used for phage titering and phage propagation of Listeria phages A511, LP-048, and LP-125. L. monocytogenes F2365 is the L. monocytogenes serotype 4b standard strain that was used for phage titering and phage propagation of the remaining Listeria phages (LP-020, LP-021, LP-024, LP-027, LP-053, LP-054, LP-057, LP-085, and LP-094). L. monocytogenes 10403S is a L. monocytogenes serotype 1/2a model strain. FSL D4-0014 and FSL D4-0119 are mutants of L. monocytogenes 10403S that lack N-acetyl glucosamine and rhamnose in their wall teichoic acid, respectively. All the strains were stored at −80 °C in Brain Heart Infusion (BHI) supplemented with 15% (w/v) glycerol and grown on 1.5% (w/v) BHI agar plates at 37 °C. Overnight cultures for each strain were inoculated with a single colony from a streak plate into BHI broth and grown at 37 °C in a shaking water bath at 160 RPM.
All Listeria phages in this study are listed in Table 2. Listeria phages LP-048 and LP-125 were well-studied phages that are able to infect serotype 1/2a strains. Listeria phage A511 is a broad range phage that is able to infect both serotype 1/2a strains and 4b strains [27,28]. The other Listeria phages (LP-020, LP-021, LP-024, LP-027, LP-053, LP-054, LP-057, LP-085, and LP-094) were included in the study due to their ability to show strong lytic activity against L. monocytogenes F2365. All phages were titered on lysogeny broth morpholino-propane sulfonic acid (LB-MOPS) agar supplemented with 0.1% glucose, 1 mM CaCl2, and 1 mM MgCl2 by 10 µL spot assay and were incubated at 25 °C overnight (16 ± 2 h). Phage stocks were prepared by liquid amplification. A culture of the host strain was grown to an OD600nm of 0.2, infected with the phage at a multiplicity of infection (MOI) of 0.1; after 3 h of incubation at 25 °C in a shaking water bath, the infected culture was filtered with a 0.45 µm SCFA sterile filter, and then centrifuged at 12,000× g at 4 °C for 2 h. The supernatant was then removed, and the pellet was resuspended in SM buffer (0.1% v/v gelatin, 0.05 M Tris-Cl pH 7.5, 0.58% w/v NaCl, 0.2% w/v MgSO4·7H2O) by static incubation at 4 °C for 24 h, then filtered with a 0.20 µm SCFA sterile filter and transferred to a sterile tube as new phage stock. All phage stocks were stored at 4 °C in SM buffer. Storage at 4 °C in liquid media with structurally similar phages has been demonstrated to maintain stable titers for months to years with minimal degradation of stock viability [29,30,31].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Listeria monocytogenes strains.
| Strain | Serotype | Reference or Original |
|---|---|---|
| 10403S | 1/2a | Bishop and Hinrichs, 1987 [32] |
| MACK | 1/2a | Hodgson, 2000 [33] |
| F2365 | 4b | Nelson, 2004 [34] |
| FSL J1-175 | 1/2b | Bergholz, 2010 [35] |
| FSL J1-208 | 4a | Roberts, 2006 [36] |
| FSL C1-115 | 3a | Fugett, 2006 [37] |
| FSL J1-094 | 1/2c | Fugett, 2006 [37] |
| FSL F2-695 | 4a | Roberts, 2006 [36] |
| FSL F2-501 | 4b | Roberts, 2006 [36] |
| FSL J2-071 | 4c | Roberts, 2006 [36] |
| FSL W1-110 | 4b | De Jesus and Whiting, 2003 [38] |
| FSL J1-158 | 4b | De Jesus and Whiting, 2003 [38] |
| FSL J1-169 | 3b | Fugett, 2006 [37] |
| FSL J1-049 | 3c | Fugett, 2006 [37] |
| FSL D4-0014 | 1/2a | Denes, 2015 [15] |
| FSL D4-0119 | 3 | Denes, 2015 [15] |
| FSL R9-0915 | 7 | Denes, 2015 [15] |
2.2. Transmission Electron Microscopy
A purified, high titer phage sample (~1 × 1010 PFU/mL) was prepared for transmission electron microscopy (TEM) as previously described with modifications [26]. One milliliter of each phage sample was washed using a 0.1 M ammonium acetate solution (pH 7) and centrifuged at 21,000× g with a microcentrifuge (Thermo Fischer Scientific, Waltham, MA, USA). One drop of the phage sample was deposited onto a 150–200 mesh carbon-coated Formvar film copper grid (Electron Microscopy Sciences, Hatfield, PA, USA) and stained using 1% phosphotungstic acid (PTA; pH 7.4). Samples were imaged using a JEOL 1400 Flash transmission electron microscope at 120 kV. Images were analyzed using Fiji 3 v.2.0.0-rc-69/1.52p.
2.3. DNA Extraction and Genomic Analysis
Phage DNA were extracted by the phenol-chloroform method as previously described [26].
Libraries were prepared using a Nextera kit (Illumina, San Diego, CA, USA) and sequenced with an Illumina NextSeq 550 using 150 bp paired-end read chemistry. LP-027 was additionally long-read sequenced; the library was prepared using a Rapid Barcoding Kit (SQK-RBK0004; Oxford Nanopore Technologies, Oxford, UK) and sequenced with a MinIon. Illumina reads were trimmed using Trimmomatic (v0.35) [42] and read quality statistics were generated using FastQC (v0.11.7). Reads were mapped to the L. monocytogenes propagation host strain (F2365) genome in order to filter out host contamination reads. Assemblies were generated using SPAdes (v3.12.0) [43] and a hybrid assembly was generated with Unicycler (v0.4.8-beta) [44] for LP-027 using both Illumina and Nanopore reads. For some genomes, reads were subsampled to obtain better assemblies. Final assemblies were re-oriented to start at the large terminase subunit and annotated using RASTtk [45], with the pipeline modified to run “annotate-proteins-phage” before “annotate-proteins-kmer-v2.” Assembly statistics were generated using Quast (v4.6.3) [46], BBMap (v38.08) [47], and SAMtools (v1.8) [48]. Average nucleotide identity (ANI) values between phages from this study and those described previously by Denes et al. [41] were calculated using BLAST with JSpeciesWS [49]. Genome similarity maps of the representative phages and most similar previously described phages were created using EasyFig (v2.2.2) [50] with similarity calculated using both BLASTn and tBLASTx (v2.11.0+) [51]. The phage lifestyles were classified using PHACTS [52] and were additionally assessed by manually inspecting the genomes for genes related to lifestyle (e.g., integrases) and the phage genomes were used as queries to BLAST against the nr/nt database limited to L. monocytogenes (txid: 1639). Sequencing data and assemblies are available on NCBI under BioProject PRJNA688926.
2.4. Efficiencies of Plaquing and Relative Phage Activity
All ten Listeria phages and sixteen L. monocytogenes strains (except MACK) listed in Table 1 were used to conduct efficiency of plaquing (EOP) and relative phage activity (RPA) assays as previously described [25,53]. In brief, bacterial lawns were prepared with the double agar overly method and allowed to solidify. All the phages were amplified from the original phage stock and diluted to 1 × 107 PFU/mL as a working stock. Serial dilutions were made from each working stock and spotted onto bacterial lawns. The EOP of each phage was determined from the highest dilution with countable plaques against the strain in question compared to the number of plaques against the phage propagation host strain. Similarly, the RPA of each phage was determined from the highest dilution with observable inhibitory activity against the strain in question compared to the phage propagation host strain. Inhibitory activity is defined as an observable inhibition of the growth of the bacterial lawn where the phage dilution was spotted either with or without the formation of any phage plaques. Three biological replicates were performed. EOP and RPA clustered heatmaps were generated using pheatmap in R [54].
2.5. One-Step Growth Curve
An exponential-phase culture of F2365 was infected with LP-020, LP-027, or LP-094 at a multiplicity of infection (MOI) of 0.1. The infected culture was incubated at 25 °C and 160 RPM for 3 h. To measure infected host cells and unabsorbed viable phages, two samples were taken at each time point. One sample was serially diluted and enumerated by the spot assay method immediately after collection, the other sample was treated with 5%(v/v) chloroform for 15 min and then serially diluted and enumerated by the spot assay method, and the plates were incubated at 25 °C for 12 h. Three biological replicates were performed.
2.6. Inhibition Growth Curve of Listeria monocytogenes F2365 by LP-020, LP-027, and LP-094
A measure of 2 mL of L. monocytogenes F2365 overnight culture was added into 100 mL LB-MOPS with 0.1% glucose, 1 mM CaCl2, and 1 mM MgCl2. The culture was incubated at 25 °C and 160 rpm until the OD600nm grew to ~0.1. The culture was then diluted 10-fold with fresh supplemented LB-MOPS. A measure of 7 mL of the diluted culture was added to twelve sterile 15 mL glass culture tubes. Each tube was infected with LP-020, LP-027, or LP-094 at MOI = 0.1, 1, 10 with SM buffer as a negative control and incubated at 25 °C and 160 RPM for 15 h. The OD600nm of each tube was measured every half an hour for 15 h on a Genesys 30 Visible spectrophotometer (Thermo-Fisher Scientific, Waltham, MA, USA). Three biological replicates were performed.
2.7. Inhibition Growth Curve of Listeria monocytogenes Cocktail by LP-020 and LP-094
Seven strains were used in this experiment: F2365 (4b), FSL J1-208 (4a), FSL F2-695 (4a), FSL F2-501 (4b), FSL J2-071 (4c), FSL W1-110 (4b), and FSL J1-148 (4b). Each strain was incubated in LB-MOPS supplemented with 0.1% glucose, 1 mM CaCl2, and 1 mM MgCl2 at 25 °C and 160 rpm until the OD600nm grew to ~0.1. A L. monocytogenes cocktail was prepared by transferring 1 mL of each strain into one 15 mL sterile tube and mixing by vortex mixer. The cocktail was then diluted 10-fold with fresh supplemented LB-MOPS. A total of 7 mL of the diluted cocktail was added to eight sterile 15 mL glass culture tubes. Each tube was infected with LP-020 or LP-094 with MOI = 0.1, 1, 10 or SM buffer as a negative control, grown at 25 °C, and shaken at 160 RPM for 15 h. The OD600nm of each tube was measured every half an hour for 15 h. Three biological replicates were performed.
3. Results and Discussion
3.1. Transmission Electron Microscopy Imaging of Wild Type Listeria Phages Revealed Two Distinct Morphologies
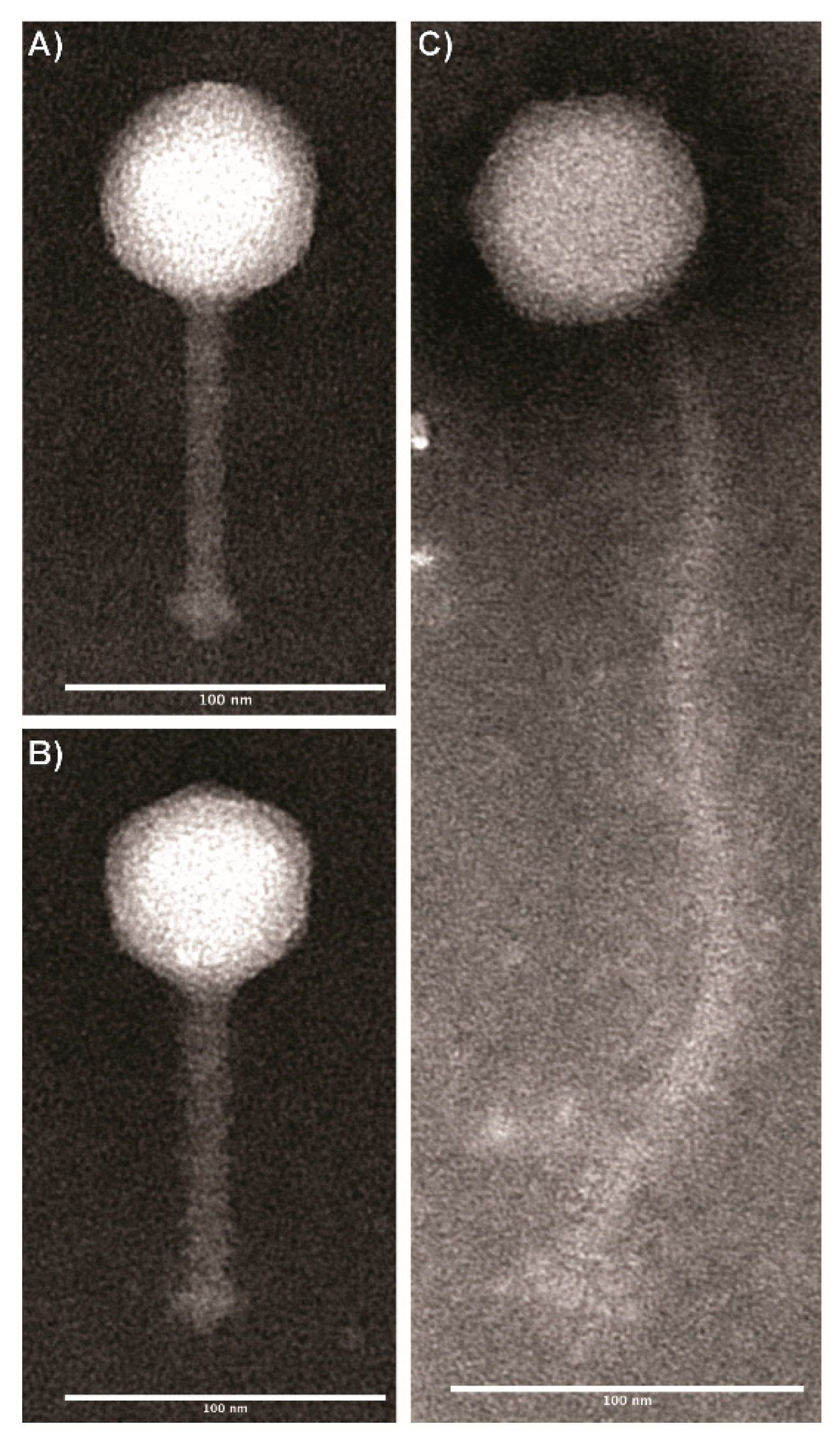
Based on morphological characteristics, all phages were classified within the Siphoviridae family (Figure 1). One group that includes two phages, LP-024 and LP-027, were found to have icosahedral capsids with flexible, elongated tails (Table 3). The second group also had icosahedral capsids; however, these phages were found to have short flexible tails (Table 3).
3.2. Genomic Analysis
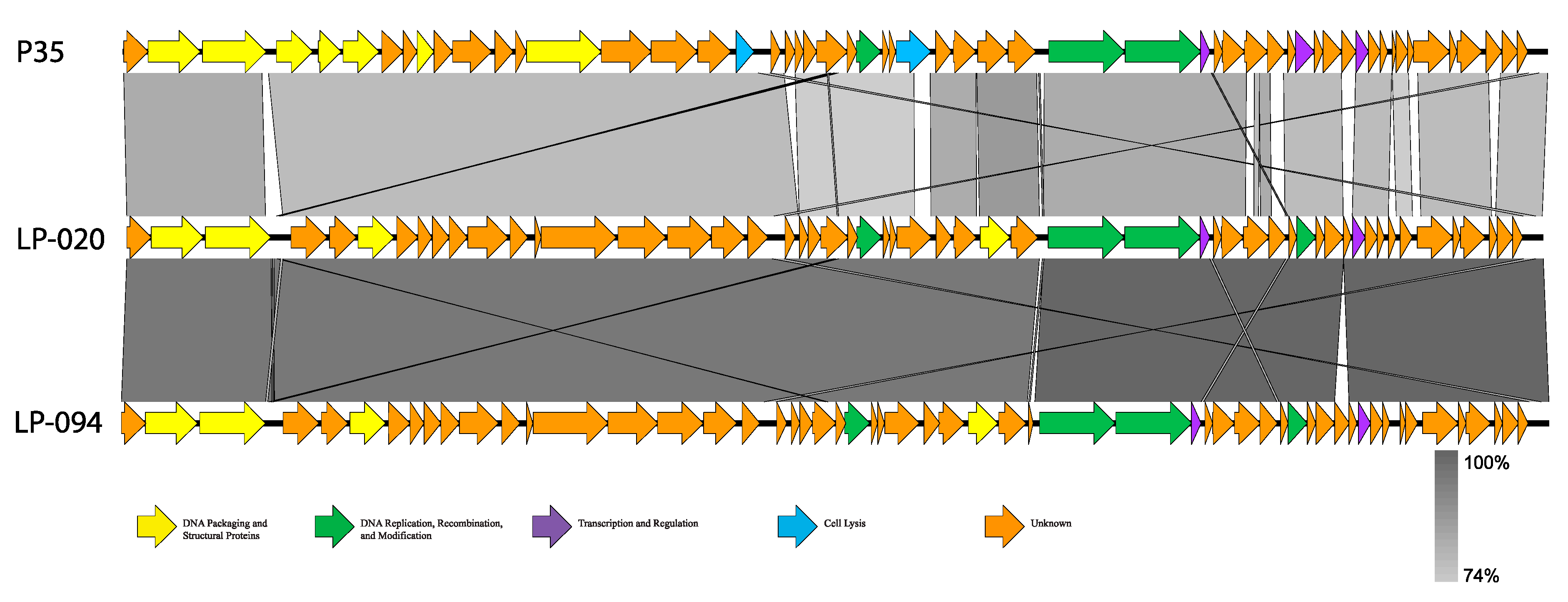
All phage reads assembled into complete, single-contig genomes; genome statistics are presented in Table 4. Based on genome statistics, two distinct groups were evident. LP-024 and LP-027 were 41.0–41.4 kb, with G+C contents of 36.5–36.6%, and each contained 74 coding sequences (CDS) and no RNAs. The other phage genomes were 35.6–36.0 kb, with G+C contents of 39.9–40.0%, and each contained 54–57 CDS and no RNAs. The two distinct groups were also supported by average nucleotide identity (ANI) values (Table 5 and Table 6) and amino acid similarity (Figure S1). LP-024 and LP-027 had an ANI of 100.00% over 95.51–96.63% of their genomes and were most similar to LP-030-3 (99.99–100.00% ANI over 95.92–96.63% of their genomes) (Table 5), a putative temperate phage. LP-030-3 was previously classified as an Orthocluster IV siphovirus [41], with a 41.2 kb genome containing 73 predicted genes. Electron micrographs show that LP-030-3 has a long and rigid tail [41]. Morphology and genome features of LP-030-3 are consistent with the LP-027-like phages and are likely putative temperate phage [41]. These two phages are likely the same genus and species as LP-030-3, as they are above the 50% and 95% cutoffs for genus and species delineation. LP-030-3 is currently listed as a “unclassified Siphoviridae”, with no genus classification, on NCBI (NCBI:txid1458852) and is not included in the most recent ICTV Master Species List 2019.v1. The other phages had an ANIs of 97.66–100.00%, over 90.63–99.24% of their genomes, and were most similar to P35 (79.51–80.36% ANI over only 83.03–87.67% of their genomes) (Table 6, Figure 2), a lytic phage [41]. P35 was previously classified as an Orthocluster II siphovirus [41], with a 35.8 kb genome containing 56 predicted genes. Electron micrographs show that P35 has a short tail [41,55]. Morphology and genome features of P35 are consistent with the LP-020-like phages and they are likely putative obligate lytic phages [41,55]. Given the >50% nucleotide similarity to P35, this second group of phages likely belongs to the same genus [56]. However, they clearly qualify as a novel species, as they are well below the 95% similarity cutoff to be considered the same species as P35. P35 is currently listed as a “unclassified Siphoviridae,” with no genus classification, on NCBI (NCBI:txid330398) and is not included in the most recent ICTV Master Species List 2019.v1.
LP-024 and LP-027 were confidently predicted as having a temperate lifestyle by PHACTS (Table S1) and both genomes contained a phage integrase gene. This was further confirmed through BLAST, which showed they have high similarity to L. monocytogenes genomes (up to 99.79% identity and 86% query coverage). LP-020 and LP-057 were non-confidently predicted as having a lytic lifestyle and LP-021, LP-053, LP-054, LP-085, and LP-094 were non-confidently predicted as having a temperate lifestyle (Table S1). However, none of these genomes contained an integrase gene, and BLAST showed they did not have high similarity to any L. monocytogenes genomes (only up to 5% query coverage), indicating that they likely have a lytic lifestyle.
3.3. Host Range Analysis
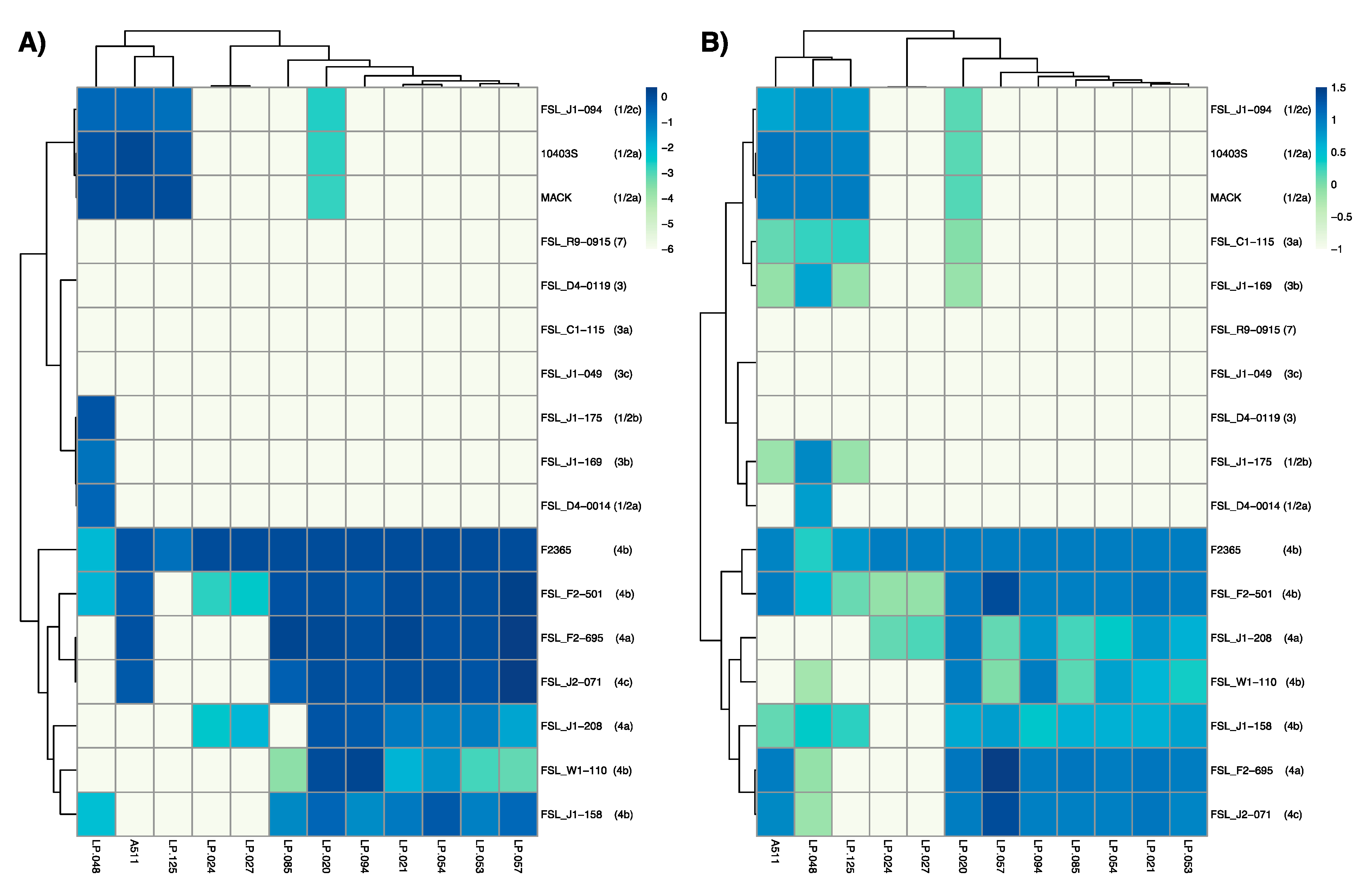
A511, which has been described as a broad host range phage [28], showed activity against 11 out of 17 L. monocytogenes (Figure 3A, Figure S2), but only formed visible plaques on seven strains (Figure 3B, Figure S2). Similarly, two previously characterized phages, LP-048 and LP-125, showed activity against a broad range of strains (thirteen and nine strains, respectively), compared to their ability to form plaques on the same strains (nine and four strains, respectively). All three of these Pecentumvirus phages were unable to infect all serotype 4b and 4a strains.
The phages selected for this study that showed strong activity against serotype 4b strain F2365 can be divided into two groups, and these groupings are consistent with those based on observed morphological characteristics. The first group, comprised of LP-024 and LP-027, showed a very narrow host range, forming plaques on F2364 (4b), FSL F2-501 (4b), and FSL J1-208 (4a). The second group, comprised of LP-020, LP-021, LP-053, LP-054, LP-057, LP-085, and LP-094, showed activity to all L. monocytogenes serotype 4 strains, and formed visible plaques on nearly all L. monocytogenes serotype 4 strains. Interestingly, LP-020 is the only phage in this group that was able to form plaques on non-serotype 4 strains.
FSL R9-0915 (7), FSL J1-049 (3c), and FSL D4-0119 (3) were the only three strains that were resistant to all phages in this study, although this is not surprising as these strains have consistently shown resistance to phage infection [17,25].
Based on the combination of their morphological characteristics and the results of their EOP and RPA, LP-020, LP-027, and LP-094 were selected as representative phages for further evaluation in this study.
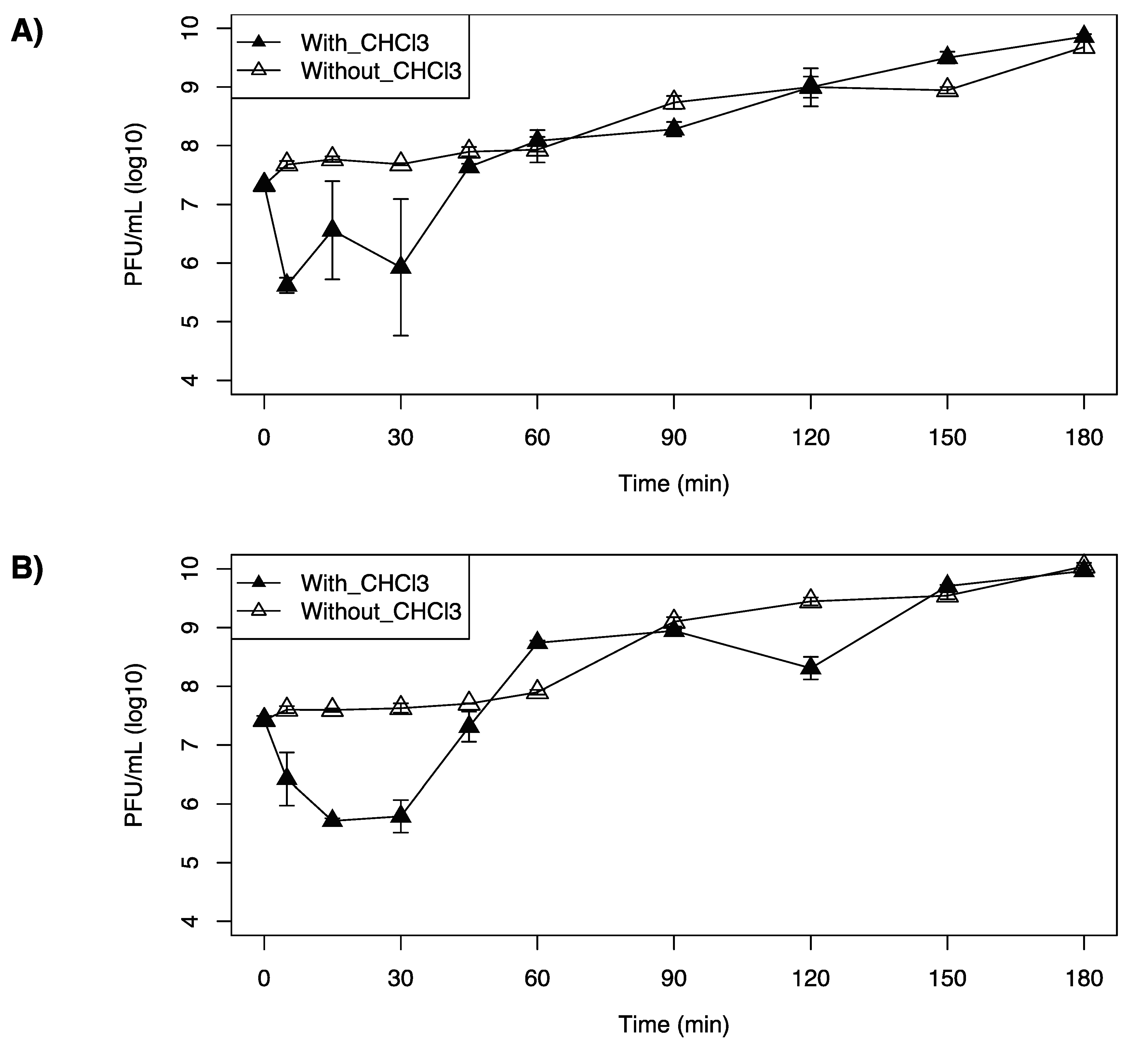
3.4. One-Step Growth Curves
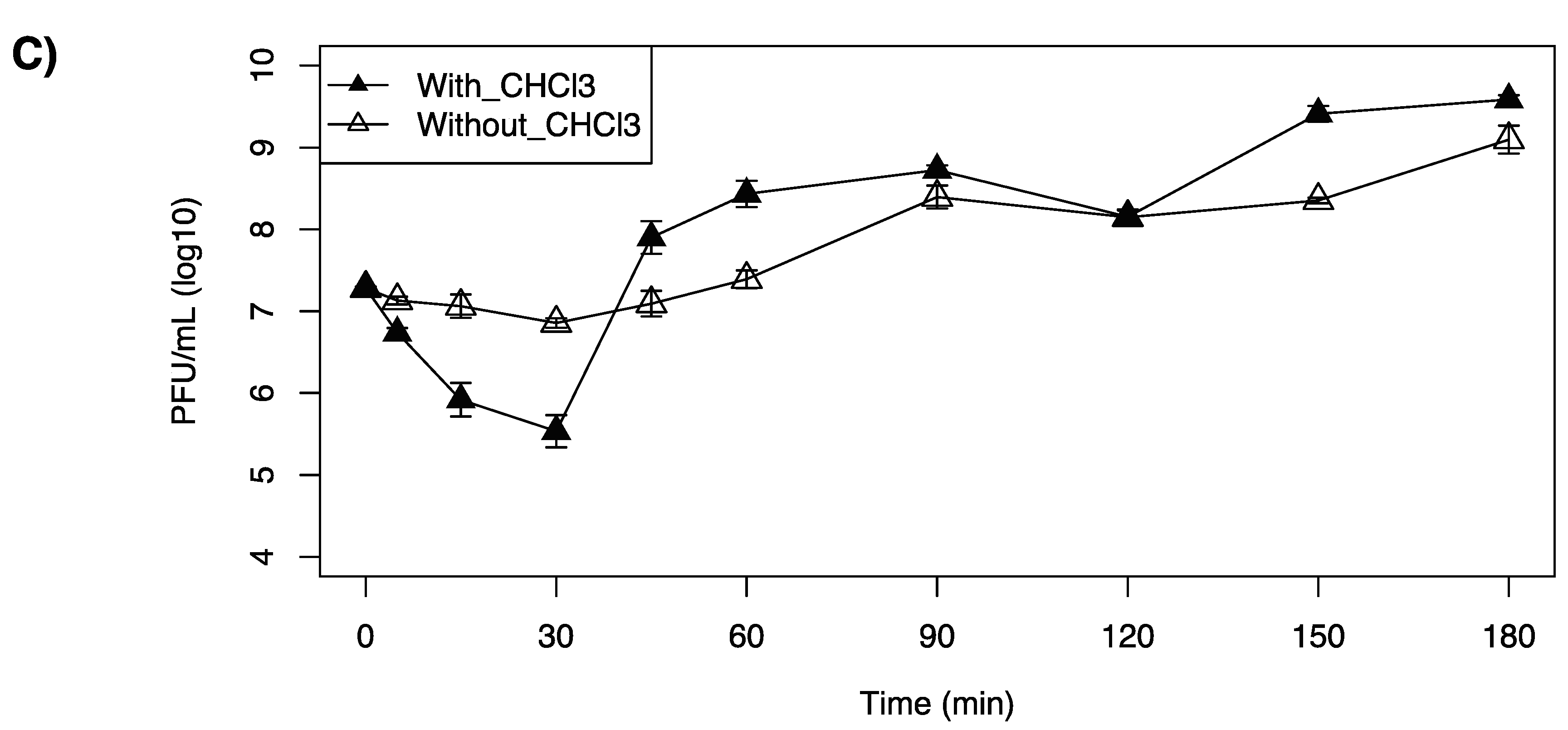
LP-020 was found to have a short adsorption time, in which 97.9% of LP-020 adsorbed to F2365 in 5 min. This is in contrast with LP-027 and LP-094, in which 30 min was required to attain adsorptions of 96.4% and 97.8%, respectively. One-step growth curve analysis of LP-020 showed a latent period of 60~90 min, an eclipse period of 5~15 min, and a burst size of ~9.7 (SE, 2.9) PFU/cell (Table 7, Figure 4A). One-step growth curve analysis of LP-027 showed a latent period of 45~60 min, an eclipse periods were 30~45 min, and a burst size of ~34.4 (SE, 5.8) PFU/cell (Table 7, Figure 4B). One-step growth curve analysis of LP-094 showed a latent period of 30~45 min, an eclipse period of 30~45 min, and a burst size of ~28.3 (SE, 4.1) PFU/cell (Table 7, Figure 4C).
However, the one-step growth curve of LP-020 showed a fluctuation in completed phage numbers between 5 min and 30 min (Figure 4A).
3.5. Inhibition Growth Curve of Listeria monocytogenes F2365 by LP-020, LP-027, and LP-094
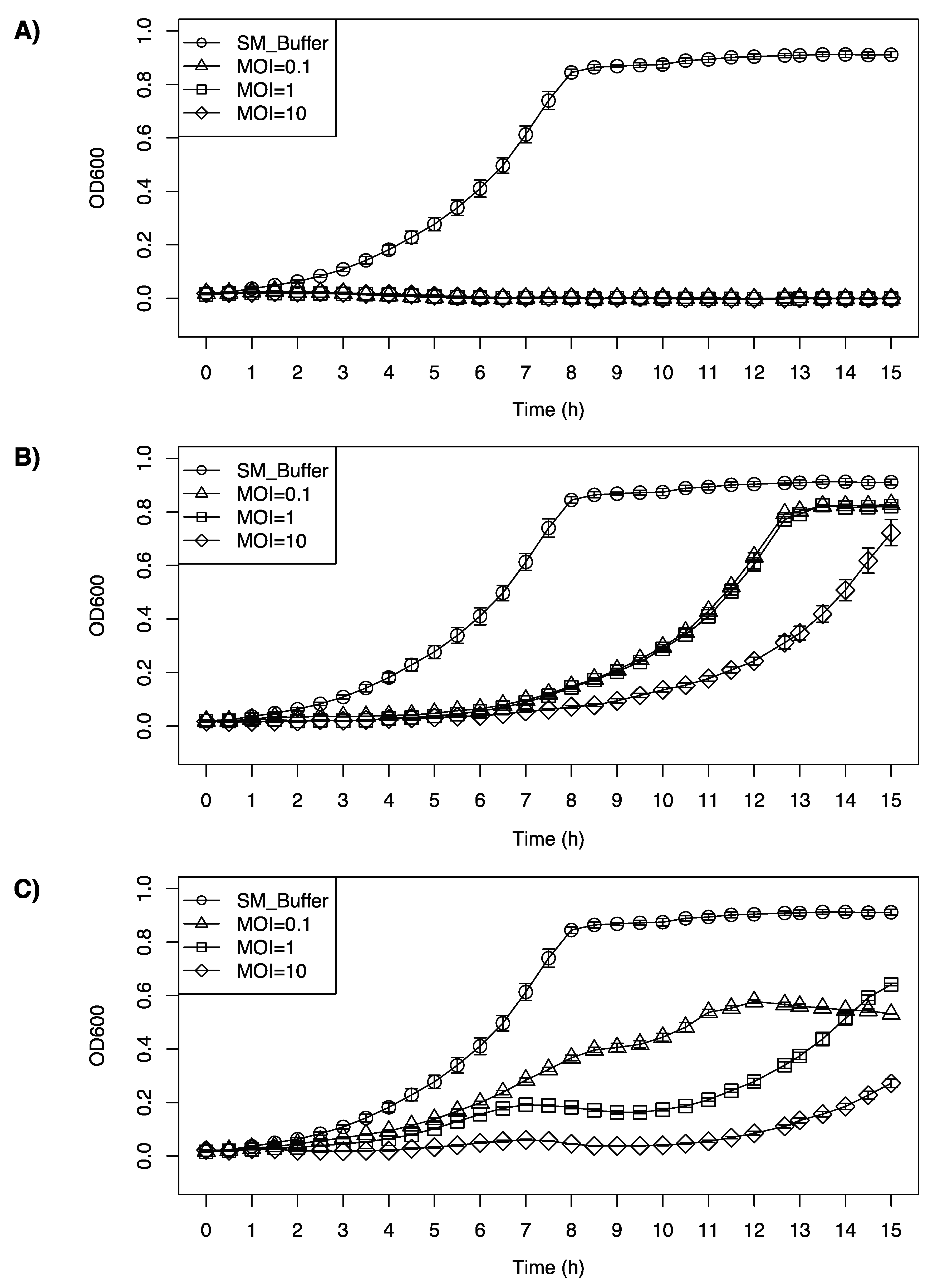
All the phages tested were able to inhibit the growth of F2365, at a range of different multiplicities of infection (MOIs). LP-020 showed high efficiency in inhibiting the growth of F2365. Even with the lowest concentration (MOI = 0.1), LP-020 can keep the OD600nm of F2365 under 0.03 for 15 h (Figure 5A). LP-027 could keep the OD600nm of F2365 under 0.1 for 7 h with MOI = 0.1 and MOI = 1, and for 9 h with MOI = 10 (Figure 5B). LP-094 could keep the OD600nm of the F2365 under 0.1 for 4 h and 4.5 h, with MOI = 0.1 and MOI = 1, respectively, and for 12 h with MOI = 10. Interestingly, after 12 h of incubation, the sample infected with LP-094 at MOI = 1 has a higher OD600nm than the sample infected with LP-094 at MOI = 0.1, possibly due to the lower MOI (0.1) providing an opportunity for host to continue during early infection, leading to greater levels of phage production during later infection (Figure 5C).
3.6. Inhibition Growth Curve of Listeria monocytogenes Cocktail by LP-020 and LP-094
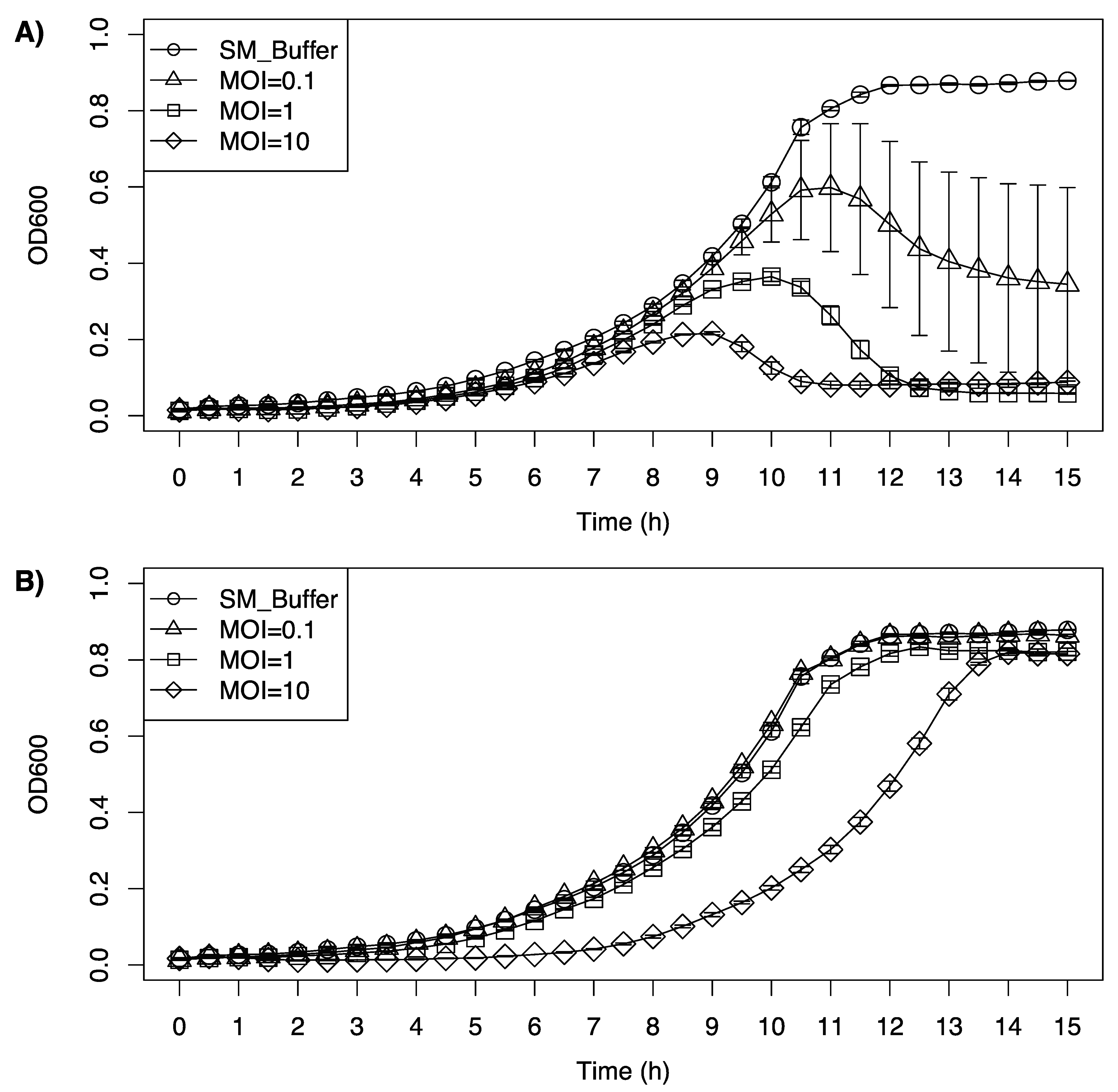
LP-020 was able to inhibit the growth of a cocktail of L. monocytogenes serotype 4 at all tested MOIs, while LP-094 only showed an inhibitory effect with the highest MOI (Figure 6). For the first 8 h after LP-020 incubation, the growth curve of the cocktail showed no substantial difference. The growth curve of the cocktail treated with different dosages of LP-020 reached its peak at 11 h, 10 h, and 9 h, respectively. After reaching their peak, the bacterial cocktails treated with LP-020 showed a reduction in OD600nm, with the 1 and 10 MOI infections reducing the OD600nm to 0.06 and 0.08, respectively. Interestingly, the low MOI treatment group (MOI = 0.1) showed a large variation in OD600nm, which may be due to the diversity of the host strains in the cocktail generating a more varied response to phage infection as the individual concentrations of each L. monocytogenes strain could vary from replicate to replicate due to complex growth interactions (Figure 6A). For the LP-094 infection, low MOI (MOI = 0.1, 1) showed no major influence of the growth curve of the cocktail. However, LP-094 can keep the OD600nm of the cocktail under 0.1 for 8 h with a high dosage (MOI = 10) (Figure 6B). Compared with LP-094, LP-020′s ability to reduce the OD600nm of a cocktail of serotype 4a, 4b, and 4c Listeria strains suggests it could be a promising phage for biocontrol applications in food processing environments.
4. Conclusions
In this study we describe two groups of Listeria phages that showed high levels of infectivity against serotype 4 strains of L. monocytogenes. One of these groups of phages, LP-020-like phages showed a level of nucleotide dissimilarity with previously sequenced phages that is well above the 5% cutoff for qualifying as a novel species. Inhibition assays of L. monocytogenes against a cocktail of serotype 4 strains (4a, 4b, and 4c) confirmed that EOP and RPA assays were predictive of a phage’s inhibitory potential. We identified LP-020 as a phage that may be particularly useful in biocontrol settings. It shows strong activity against all serotype 4a, 4b, and 4c strains tested here. Given that some types of phage resistance is specific to a single species or closely related group of phages, such as CRISPR-Cas mediated resistance, it is critical to consider using diverse cocktails of phages. LP-020 may serve together with Listeria phages from other groups, such as Pecentumviruses [11,57,58] and Homburgviruses [26,59,60], to increase both the diversity of Listeria phage cocktails and their infectivity against the problematic serotype 4 strains that are often associated with human illness. Further, the temperate phages described here, which are similar to LP-030-3, may be useful as biocontrol agents if their integrases are knocked out, which has previously been shown for the temperate Listeria phage PSA [61].
Supplementary Materials
Supplementary materials can be found at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/v13040671/s1.
Author Contributions
Conceptualization, Y.S. and T.G.D.; methodology, Y.S., L.K.H., D.W.B., T.L.P. and T.G.D.; formal analysis, L.K.H.; resources, T.G.D.; writing—original draft preparation, Y.S., T.L.P., L.K.H. and T.G.D.; writing—review and editing, all authors.; supervision, T.G.D.; funding acquisition, T.G.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Tennessee Fellowship for Graduate Excellence (awarded to T.L.P.), by the University of Tennessee Institute of Agriculture (experimental startup package for T.G.D.), and by multistate project S1077, “Enhancing Microbial Food Safety by Risk Analysis”. Additionally, this work was supported by Food Safety and Defense grant no. 2020-67017-30790/project accession no. 1021935 from the USDA National Institute of Food and Agriculture.
Data Availability Statement
Sequencing data and assemblies are available on NCBI under BioProject PRJNA688926.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Karthikeyan, R.; Gayathri, P.; Gunasekaran, P.; Jagannadham, M.V.; Rajendhran, J. Comprehensive proteomic analysis and pathogenic role of membrane vesicles of Listeria monocytogenes serotype 4b reveals proteins associated with virulence and their possible interaction with host. Int. J. Med. Microbiol. 2019, 309, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Vivant, A.L.; Garmyn, D.; Piveteau, P. Listeria monocytogenes, a down-to-earth pathogen. Front. Cell. Infect. Microbiol. 2013, 3, 87. [Google Scholar] [CrossRef] [Green Version]
- Sauders, B.D.; Overdevest, J.; Fortes, E.; Windham, K.; Schukken, Y.; Lembo, A.; Wiedmann, M. Diversity of Listeria Species in Urban and Natural Environments. Appl. Environ. Microbiol. 2012, 78, 4420–4433. [Google Scholar] [CrossRef] [Green Version]
- Fenlon, D. Wild birds and silage as reservoirs of Listeria in the agricultural environment. J. Appl. Bacteriol. 1985, 59, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Lawrence, M.L.; Ainsworth, A.J.; Austin, F.W. Comparative assessment of acid, alkali and salt tolerance in Listeria monocytogenes virulent and avirulent strains. FEMS Microbiol. Lett. 2005, 243, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Malakar, D.; Borah, P.; Das, L.; Kumar, N.S. A Comprehensive Review on Molecular Characteristics and Food-Borne Outbreaks of Listeria Monocytogenes. Sci. Technol. J. 2019, 7. [Google Scholar] [CrossRef]
- De Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7. [Google Scholar] [CrossRef]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607. [Google Scholar] [CrossRef] [PubMed]
- Cost Estimates of Foodborne Illnesses. 2021. Available online: https://www.ers.usda.gov/data-products/cost-estimates-of-foodborne-illnesses.aspx (accessed on 4 March 2021).
- Carlton, R.; Noordman, W.; Biswas, B.; de Meester, E.; Loessner, M. Bacteriophage P100 for control of Listeria monocytogenes in foods: Genome sequence, bioinformatic analyses, oral toxicity study, and application. Regul. Toxicol. Pharmacol. 2005, 43, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Jurczak-Kurek, A.; Gąsior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of bacteriophages: Morphological and biological properties of a large group of phages isolated from urban sewage. Sci. Rep. 2016, 6, 34338. [Google Scholar] [CrossRef]
- Sillankorva, S.M.; Oliveira, H.; Azeredo, J. Bacteriophages and Their Role in Food Safety. Int. J. Microbiol. 2012, 2012, 863945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibeu, A.; Agius, L.; Gao, A.; Sabour, P.M.; Kropinski, A.M.; Balamurugan, S. Efficacy of bacteriophage LISTEX™P100 combined with chemical antimicrobials in reducing Listeria monocytogenes in cooked turkey and roast beef. Int. J. Food Microbiol. 2013, 167, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Denes, T.; Bakker, H.C.D.; Tokman, J.I.; Guldimann, C.; Wiedmann, M. Selection and Characterization of Phage-Resistant Mutant Strains of Listeria monocytogenes Reveal Host Genes Linked to Phage Adsorption. Appl. Environ. Microbiol. 2015, 81, 4295–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumrall, E.T.; Keller, A.P.; Shen, Y.; Loessner, M.J. Structure and function of Listeria teichoic acids and their implications. Mol. Microbiol. 2020, 113, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Vongkamjan, K.; Switt, A.M.; Bakker, H.C.D.; Fortes, E.D.; Wiedmann, M. Silage Collected from Dairy Farms Harbors an Abundance of Listeriaphages with Considerable Host Range and Genome Size Diversity. Appl. Environ. Microbiol. 2012, 78, 8666–8675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasebe, R.; Nakao, R.; Ohnuma, A.; Yamasaki, T.; Sawa, H.; Takai, S.; Horiuchi, M. Listeria monocytogenes serotype 4b strains replicate in monocytes/macrophages more than the other serotypes. J. Vet. Med. Sci. 2017, 79, 962–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontello, M.; Guaita, A.; Sala, G.; Cipolla, M.; Gattuso, A.; Sonnessa, M.; Gianfranceschi, M.V. Listeria monocytogenes serotypes in human infections (Italy, 2000–2010). Ann. dell’Istituto Super. Sanità 2012, 48, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Aarnisalo, K.; Autio, T.; Sjöberg, A.M.; Lundén, J.; Korkeala, H.; Suihko, M.L. Typing of Listeria monocytogenes isolates originating from the food processing industry with automated ribotyping and pulsed-field gel electrophoresis. J. Food Prot. 2003, 66, 249–255. [Google Scholar] [CrossRef]
- Liu, D. Handbook of Listeria Monocytogenes; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Huang, Y.; Morvay, A.A.; Shi, X.; Suo, Y.; Shi, C.; Knøchel, S. Comparison of oxidative stress response and biofilm formation of Listeria monocytogenes serotypes 4b and 1/2a. Food Control 2018, 85, 416–422. [Google Scholar] [CrossRef]
- Nucera, D.; Lomonaco, S.; Bianchi, D.M.; Decastelli, L.; Grassi, M.A.; Bottero, M.T.; Civera, T. A five year surveillance report on PFGE types of Listeria monocytogenes isolated in Italy from food and food related environments. Int. J. Food Microbiol. 2010, 140, 271–276. [Google Scholar] [CrossRef]
- Leong, D.; Alvarez-Ordóñez, A.; Jordan, K. Monitoring occurrence and persistence of Listeria monocytogenes in foods and food processing environments in the Republic of Ireland. Front. Microbiol. 2014, 5, 436. [Google Scholar] [CrossRef]
- Peters, T.L.; Song, Y.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Mutant and Recombinant Phages Selected from In Vitro Coevolution Conditions Overcome Phage-Resistant Listeria monocytogenes. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Song, Y.; Peters, T.L.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Homburgvirus LP-018 Has a Unique Ability to Infect Phage-Resistant Listeria monocytogenes. Viruses 2019, 11, 1166. [Google Scholar] [CrossRef] [Green Version]
- Tokman, J.I.; Kent, D.J.; Wiedmann, M.; Denes, T. Temperature Significantly Affects the Plaquing and Adsorption Efficiencies of Listeria Phages. Front. Microbiol. 2016, 7, 631. [Google Scholar] [CrossRef]
- Kim, J.-W.; Kathariou, S. Temperature-Dependent Phage Resistance of Listeria monocytogenes Epidemic Clone II. Appl. Environ. Microbiol. 2009, 75, 2433–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, H.-W.; Tremblay, D.; Moineau, S. Long-Term Bacteriophage Preservation. 2004. Available online: https://www.researchgate.net/profile/Sylvain-Moineau/publication/285783875_Long-term_bacteriophage_preservation/links/56965e4b08ae34f3cf1dbb34/Long-term-bacteriophage-preservation.pdf (accessed on 12 April 2021).
- Clark, W.A. Comparison of several methods for preserving bacteriophages. Appl. Microbiol. 1962, 10, 466–471. [Google Scholar] [CrossRef]
- Cooper, C.; Denyer, S.; Maillard, J.-Y. Stability and purity of a bacteriophage cocktail preparation for nebulizer delivery. Lett. Appl. Microbiol. 2013, 58, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.; Hinrichs, D. Adoptive transfer of immunity to Listeria monocytogenes. The influence of in vitro stimulation on lymphocyte subset requirements. J. Immunol. 1987, 139, 2005–2009. [Google Scholar] [PubMed]
- Hodgson, D.A. Generalized transduction of serotype 1/2 and serotype 4b strains of Listeria monocytogenes. Mol. Microbiol. 2000, 35, 312–323. [Google Scholar] [CrossRef]
- Nelson, K.E.; Fouts, D.E.; Mongodin, E.F.; Ravel, J.; DeBoy, R.T.; Kolonay, J.F.; Rasko, D.A.; Angiuoli, S.V.; Gill, S.R.; Paulsen, I.T.; et al. Whole genome comparisons of serotype 4b and 1/2a strains of the food-borne pathogen Listeria monocytogenes reveal new insights into the core genome components of this species. Nucleic Acids Res. 2004, 32, 2386–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergholz, T.M.; Bakker, H.C.D.; Fortes, E.D.; Boor, K.J.; Wiedmann, M. Salt Stress Phenotypes in Listeria monocytogenes Vary by Genetic Lineage and Temperature. Foodborne Pathog. Dis. 2010, 7, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Nightingale, K.; Jeffers, G.; Fortes, E.; Kongo, J.M.; Wiedmann, M. Genetic and phenotypic characterization of Listeria monocytogenes lineage III. Microbiology 2006, 152, 685–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fugett, E.; Fortes, E.; Nnoka, C.; Wiedmann, M. International Life Sciences Institute North America Listeria monocytogenes strain collection: Development of standard Listeria monocytogenes strain sets for research and validation studies. J. Food Prot. 2006, 69, 2929–2938. [Google Scholar] [CrossRef]
- De Jesus, A.J.; Whiting, R.C. Thermal inactivation, growth, and survival studies of Listeria monocytogenes strains belonging to three distinct genotypic lineages. J. Food Prot. 2003, 66, 1611–1617. [Google Scholar] [CrossRef]
- Loessner, M.J.; Busse, M. Bacteriophage typing of Listeria species. Appl. Environ. Microbiol. 1990, 56, 1912–1918. [Google Scholar] [CrossRef] [Green Version]
- Klumpp, J.; Dorscht, J.; Lurz, R.; Bielmann, R.; Wieland, M.; Zimmer, M.; Calendar, R.; Loessner, M.J. The Terminally Redundant, Nonpermuted Genome of Listeria Bacteriophage A511: A Model for the SPO1-Like Myoviruses of Gram-Positive Bacteria. J. Bacteriol. 2008, 190, 5753–5765. [Google Scholar] [CrossRef] [Green Version]
- Denes, T.; Vongkamjan, K.; Ackermann, H.-W.; Switt, A.I.M.; Wiedmann, M.; Bakker, H.C.D. Comparative Genomic and Morphological Analyses of Listeria Phages Isolated from Farm Environments. Appl. Environ. Microbiol. 2014, 80, 4616–4625. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Bushnell, B. BBTools: A Suite of Fast, Multithreaded Bioinformatics Tools Designed for Analysis of DNA and RNA Sequence Data. Joint Genome Institute. 2018. Available online: https://jgi.doe.gov/data-and-tools/bbtools (accessed on 29 December 2020).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2015, 32, 929–931. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a computational approach to classifying the lifestyle of phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef] [Green Version]
- Trudelle, D.M.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Cross-resistance to phage infection in Listeria monocytogenes serotype 1/2a mutants. Food Microbiol. 2019, 84, 103239. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps (Version 1.0.12). 2019. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 1 October 2020).
- Dorscht, J.; Klumpp, J.; Bielmann, R.; Schmelcher, M.; Born, Y.; Zimmer, M.; Calendar, R.; Loessner, M.J. Comparative Genome Analysis of Listeria Bacteriophages Reveals Extensive Mosaicism, Programmed Translational Frameshifting, and a Novel Prophage Insertion Site. J. Bacteriol. 2009, 191, 7206–7215. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.M.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.D.; Klumpp, J.; Barrios, M.E.; Mbayed, V.A. Gene gain and loss and recombination shape evolution of Listeria bacteriophages of the genus Pecentumvirus. Genomics 2020, 113, 411–419. [Google Scholar] [CrossRef]
- Peters, T.L.; Hudson, L.K.; Song, Y.; Denes, T.G. Complete Genome Sequences of Two Listeria Phages of the Genus Pecentumvirus. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, L.K.; Peters, T.L.; Song, Y.; Denes, T.G. Complete Genome Sequences and Transmission Electron Micrographs of Listeria Phages of the Genus Homburgvirus. Microbiol. Resour. Announc. 2019, 8, e00825-19. [Google Scholar] [CrossRef] [Green Version]
- Schmuki, M.M.; Erne, D.; Loessner, M.J.; Klumpp, J. Bacteriophage P70: Unique Morphology and Unrelatedness to Other Listeria Bacteriophages. J. Virol. 2012, 86, 13099–13102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilcher, S.; Studer, P.; Muessner, C.; Klumpp, J.; Loessner, M.J. Cross-genus rebooting of custommade, synthetic bacteriophage genomes in L-form bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 567–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Transmission electron microscopy images of Listeria phages representing two morphologies: (A) LP-020 and (B) LP-094, characterized by an icosahedral capsid and a flexible, non-contractile tail; (C) LP-027, characterized by an icosahedral capsid and a long, flexible, non-contractile tail. Phages were stained with 1% phosphotungstic acid (pH &) and imaged at a final magnification of ×69,700–83,600. Images were analyzed using FIJI 3 (v2.0.0-rc-69/1.52p).
Figure 1.
Transmission electron microscopy images of Listeria phages representing two morphologies: (A) LP-020 and (B) LP-094, characterized by an icosahedral capsid and a flexible, non-contractile tail; (C) LP-027, characterized by an icosahedral capsid and a long, flexible, non-contractile tail. Phages were stained with 1% phosphotungstic acid (pH &) and imaged at a final magnification of ×69,700–83,600. Images were analyzed using FIJI 3 (v2.0.0-rc-69/1.52p).

Figure 2.
Linear BLASTn comparisons of representative P35-like Listeria phages. Genes are represented by arrows and are colored based on putative function (see key at bottom). The shaded region between genomes represents nucleotide similarity, with the darker gray representing higher similarity and lighter gray indicating lower similarity (see scale at bottom right).
Figure 2.
Linear BLASTn comparisons of representative P35-like Listeria phages. Genes are represented by arrows and are colored based on putative function (see key at bottom). The shaded region between genomes represents nucleotide similarity, with the darker gray representing higher similarity and lighter gray indicating lower similarity (see scale at bottom right).

Figure 3.
Host range analysis of Listeria phages against a panel of Listeria monocytogenes strains that represent different serotypes. Panel (A) represents efficiency of plaquing (EOP) results where values represent the log transformed efficiencies of plaquing of each phage against each bacterial strain compared to the phage propagation host strain. Panel (B) represents efficiency of activity (EOA) results where values represent the greatest dilution factor where phage activity was observed against each strain relative to the phage propagation host strain. Values are the mean of data from three biological replicates.
Figure 3.
Host range analysis of Listeria phages against a panel of Listeria monocytogenes strains that represent different serotypes. Panel (A) represents efficiency of plaquing (EOP) results where values represent the log transformed efficiencies of plaquing of each phage against each bacterial strain compared to the phage propagation host strain. Panel (B) represents efficiency of activity (EOA) results where values represent the greatest dilution factor where phage activity was observed against each strain relative to the phage propagation host strain. Values are the mean of data from three biological replicates.

Figure 4.
One-step growth curve of Listeria monocytogenes F2365 treated with (A) LP-20, (B) LP-0-27, or (C) LP-094 at a MOI = 0.1 at 25 °C. Filled triangles represent the phage titer in chloroform treated samples and unfilled triangles represent the phage titer in untreated samples. Data are mean values of three biological replicates and error bars represent standard error.
Figure 4.
One-step growth curve of Listeria monocytogenes F2365 treated with (A) LP-20, (B) LP-0-27, or (C) LP-094 at a MOI = 0.1 at 25 °C. Filled triangles represent the phage titer in chloroform treated samples and unfilled triangles represent the phage titer in untreated samples. Data are mean values of three biological replicates and error bars represent standard error.


Figure 5.
Inhibition growth curve of Listeria monocytogenes F2365 treated with (A) LP-020, (B) LP-027 or (C) LP-094 at different MOIs. Unfilled circles represent SM buffer control, unfilled triangles represent samples treated at an MOI of 0.1, unfilled squares represent samples treated at an MOI of 1, and diamonds represent samples treated at a MOI of 10. Data are mean values of three biological replicates and error bars represent the standard error.
Figure 5.
Inhibition growth curve of Listeria monocytogenes F2365 treated with (A) LP-020, (B) LP-027 or (C) LP-094 at different MOIs. Unfilled circles represent SM buffer control, unfilled triangles represent samples treated at an MOI of 0.1, unfilled squares represent samples treated at an MOI of 1, and diamonds represent samples treated at a MOI of 10. Data are mean values of three biological replicates and error bars represent the standard error.

Figure 6.
Inhibition growth curve of a cocktail of Listeria monocytogenes serotype 4 strains (F2365 (4b), FSL J1-208 (4a), FSL F2-695 (4a), FSL F2-501 (4b), FSL J2-071 (4c), FSL W1-110 (4b), and FSL J1-148 (4b)) treated with (A) LP-020 and (B) LP-094 at different MOIs. Unfilled circles represent SM buffer control, unfilled triangles represent samples treated at a MOI of 0.1, unfilled squares represent samples treated at a MOI of 1, and diamonds represent samples treated at a MOI of 10. Data are mean values of three biological replicates and error bars represent the standard error.
Figure 6.
Inhibition growth curve of a cocktail of Listeria monocytogenes serotype 4 strains (F2365 (4b), FSL J1-208 (4a), FSL F2-695 (4a), FSL F2-501 (4b), FSL J2-071 (4c), FSL W1-110 (4b), and FSL J1-148 (4b)) treated with (A) LP-020 and (B) LP-094 at different MOIs. Unfilled circles represent SM buffer control, unfilled triangles represent samples treated at a MOI of 0.1, unfilled squares represent samples treated at a MOI of 1, and diamonds represent samples treated at a MOI of 10. Data are mean values of three biological replicates and error bars represent the standard error.

Table 2.
Listeria monocytogenes phages.
| Phages | Description | Reference or Original |
|---|---|---|
| A511 | Loessner and Busse, 1990 [39]; Klumpp, Jochen, et al., 2008 [40] | |
| LP-020 | Vongkamjan et al., 2012 | |
| LP-021 | Vongkamjan et al., 2012 [17] | |
| LP-024 | Vongkamjan et al., 2012 [17] | |
| LP-027 | Vongkamjan et al., 2012 [17] | |
| LP-048 | P100-like phage | Vongkamjan et al., 2012; Denes et al., 2014 [17,41] |
| LP-053 | Vongkamjan et al., 2012 [17,41] | |
| LP-054 | Vongkamjan et al., 2012 [17] | |
| LP-057 | Vongkamjan et al., 2012 [17] | |
| LP-085 | Vongkamjan et al., 2012 [17] | |
| LP-094 | Vongkamjan et al., 2012 [17] | |
| LP-125 | P100-like phage | Vongkamjan et al., 2012; Denes et al., 2014 [17,41] |
Table 3.
Morphology of Listeria monocytogenes phages.
| Listeria monocytogenes Phages | Capsid Diameter (nm) | Tail Length (nm) | Tail Width (nm) |
|---|---|---|---|
| LP-020 | 73.40 ± 0.61 | 104.80 ± 1.45 | 11.54 ± 0.09 |
| LP-021 | 69.92 ± 4.62 | 102.45 ± 2.32 | 11.01 ± 0.61 |
| LP-024 | 66.10 ± 5.07 | 291.29 ± 18.56 | 8.25 ± 2.54 |
| LP-027 | 70.35 ± 2.40 | 295.38 ± 10.42 | 7.95 ± 2.09 |
| LP-053 | 68.03 ± 4.76 | 104.01 ± 3.00 | 11.88 ± 1.91 |
| LP-054 | 73.70 ± 0.50 | 100.71 ± 3.55 | 11.43 ± 0.33 |
| LP-057 | 73.60 ± 0.37 | 102.45 ± 1.62 | 13.46 ± 0.90 |
| LP-085 | 72.22 ± 0.62 | 104.02 ± 3.19 | 12.61 ± 0.77 |
| LP-094 | 71.75 ± 2.98 | 100.49 ± 2.87 | 12.81 ± 0.97 |
Table 4.
Assembly statistics for Listeria phages.
| Phage | BioSample ID | Length (bp) | Avg. Illumina Read Coverage (X) | G+C (%) | No. CDS | No. RNAs |
|---|---|---|---|---|---|---|
| LP-020 | SAMN17217625 | 35,609 | 326.6 | 40.0 | 54 | 0 |
| LP-021 | SAMN17217626 | 35,610 | 344.1 | 40.0 | 54 | 0 |
| LP-024 | SAMN17217627 | 40,964 | 153.7 | 36.5 | 74 | 0 |
| LP-027 | SAMN17217628 | 41,120 | 89.6 | 36.6 | 74 | 0 |
| LP-053 | SAMN17217629 | 35,951 | 143.5 | 40.0 | 57 | 0 |
| LP-054 | SAMN17217630 | 35,951 | 524.9 | 40.0 | 57 | 0 |
| LP-057 | SAMN17217631 | 35,608 | 1001.1 | 40.0 | 54 | 0 |
| LP-085 | SAMN17217632 | 35,951 | 650.4 | 39.9 | 57 | 0 |
| LP-094 | SAMN17217633 | 35,885 | 3397.6 | 40.0 | 56 | 0 |
Table 5.
JSpecies results for LP-030-03-like Listeria phages.
| Average Nucleotide Identity (ANI; %) [Aligned Nucleotides (%)] | ||||||
|---|---|---|---|---|---|---|
| Phage | LP-024 | LP-027 | LP-030-3 | A500 | A118 | A006 |
| LP-024 | 100.00 [96.63] | 100.00 [96.63] | 92.81 [63.99] | 86.34 [52.10] | 92.00 [6.82] | |
| LP-027 | 100.00 [95.51] | 100.00 [95.92] | 92.77 [63.41] | 86.10 [50.80] | 91.76 [5.70] | |
| LP-030-3 | 99.99 [96.21] | 100.00 [96.57] | 92.07 [62.93] | 84.54 [47.14] | 88.41 [6.80] | |
| A500 | 92.74 [68.96] | 92.74 [68.96] | 92.74 [69.01] | 84.52 [49.43] | 89.93 [6.42] | |
| A118 | 84.79 [53.25] | 84.78 [53.48] | 84.78 [53.51] | 84.00 [47.63] | 95.38 [23.01] | |
| A006 | 89.85 [7.30] | 89.54 [9.18] | 89.54 [9.18] | 84.93 [4.69] | 94.91 [27.18] | |
Table 6.
JSpecies results for P35-like Listeria phages.
| Average Nucleotide Identity (ANI; %) [Aligned Nucleotides (%)] | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Phage | LP-020 | LP-021 | LP-053 | LP-054 | LP-057 | LP-085 | LP-094 | P35 | P40 |
| LP-020 | 99.99 [97.36] | 97.68 [94.41] | 97.68 [94.41] | 98.71 [97.35] | 97.86 [94.41] | 97.66 [93.39] | 79.51 [85.61] | 61.76 [32.56] | |
| LP-021 | 99.99 [97.36] | 97.69 [94.41] | 97.69 [94.41] | 98.72 [97.34] | 97.87 [94.41] | 97.68 [93.39] | 79.52 [85.63] | 61.76 [32.55] | |
| LP-053 | 97.77 [90.63] | 97.78 [90.63] | 100.00 [99.24] | 99.15 [90.64] | 99.74 [99.24] | 99.98 [98.21] | 79.85 [87.67] | 61.47 [33.98] | |
| LP-054 | 97.77 [90.63] | 97.78 [90.63] | 100.00 [99.24] | 99.15 [90.64] | 99.74 [99.24] | 99.98 [98.21] | 79.85 [87.67] | 61.47 [33.98] | |
| LP-057 | 98.71 [97.34] | 98.72 [97.35] | 98.97 [94.42] | 98.97 [94.42] | 99.02 [94.42] | 98.96 [93.40] | 79.62 [85.79] | 61.80 [32.41] | |
| LP-085 | 97.97 [90.63] | 97.98 [90.63] | 99.74 [99.24] | 99.74 [99.24] | 99.22 [90.64] | 99.73 [98.21] | 79.87 [87.67] | 61.51 [33.94] | |
| LP-094 | 97.75 [91.58] | 97.76 [91.58] | 99.98 [98.24] | 99.98 [98.24] | 99.08 [91.59] | 99.73 [98.24] | 79.98 [86.27] | 61.54 [32.97] | |
| P35 | 79.85 [83.06] | 79.86 [83.07] | 80.35 [84.12] | 80.35 [84.12] | 80.02 [83.03] | 80.36 [84.12] | 80.33 [84.12] | 63.10 [23.90] | |
| P40 | 61.52 [25.48] | 61.81 [24.84] | 62.26 [22.30] | 62.26 [22.30] | 62.37 [22.14] | 62.36 [22.26] | 62.29 [22.30] | 62.80 [24.06] | |
Table 7.
Infection kinetics summary.
| Listeria monocytogenes Phages | LP-020 | LP-027 | LP-094 |
|---|---|---|---|
| Adsorption Time(min) | 5 | 30 | 30 |
| Adsorption Rate(%) | 97.9 ± 0.5 | 96.4 ± 2.3 | 97.8 ± 1.0 |
| Latent Period(min) | 60~90 | 45~60 | 30~45 |
| Eclipse Period(min) | 5~15 | 30~45 | 30~45 |
| Burst Size(PFU/cell) | 9.7 ± 2.9 | 34.4 ± 5.8 | 28.3 ± 4.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Song, Y.; Peters, T.L.; Bryan, D.W.; Hudson, L.K.; Denes, T.G. Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes. Viruses 2021, 13, 671. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040671
AMA Style
Song Y, Peters TL, Bryan DW, Hudson LK, Denes TG. Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes. Viruses. 2021; 13(4):671. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040671
Chicago/Turabian StyleSong, Yaxiong, Tracey L. Peters, Daniel W. Bryan, Lauren K. Hudson, and Thomas G. Denes. 2021. "Characterization of a Novel Group of Listeria Phages That Target Serotype 4b Listeria monocytogenes" Viruses 13, no. 4: 671. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040671
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.