
A Versatile qPCR for Diagnosis of Leporid Gammaherpesvirus 5 Using Evagreen® or Taqman® Technologies

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Samples
2.2. DNA Extraction
2.3. Primer Design
2.4. Cloning
2.5. Evaluation of the PCR Component Variation
2.6. Field Samples Analysis
2.7. Detection Limit, Sensitivity, Specificity, Repeatability, and Reproducibility of the qPCR
2.8. Validation of 18S DNA as Internal Control
2.9. Clinical Samples Validation
3. Results
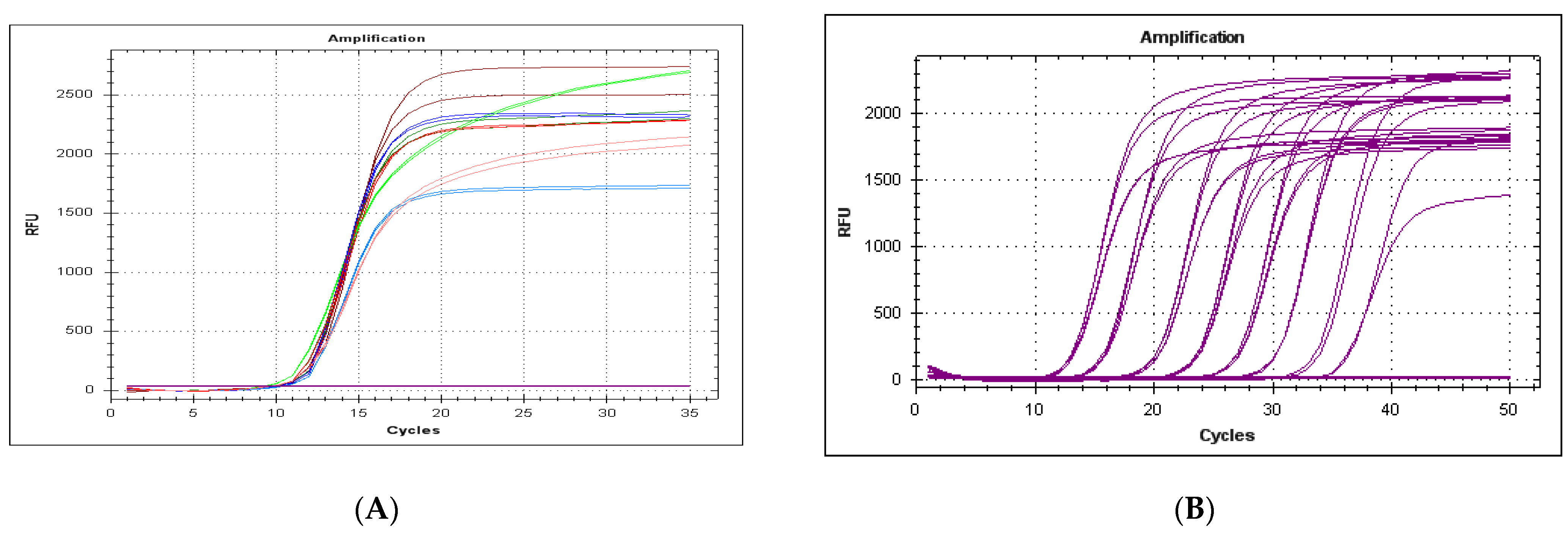
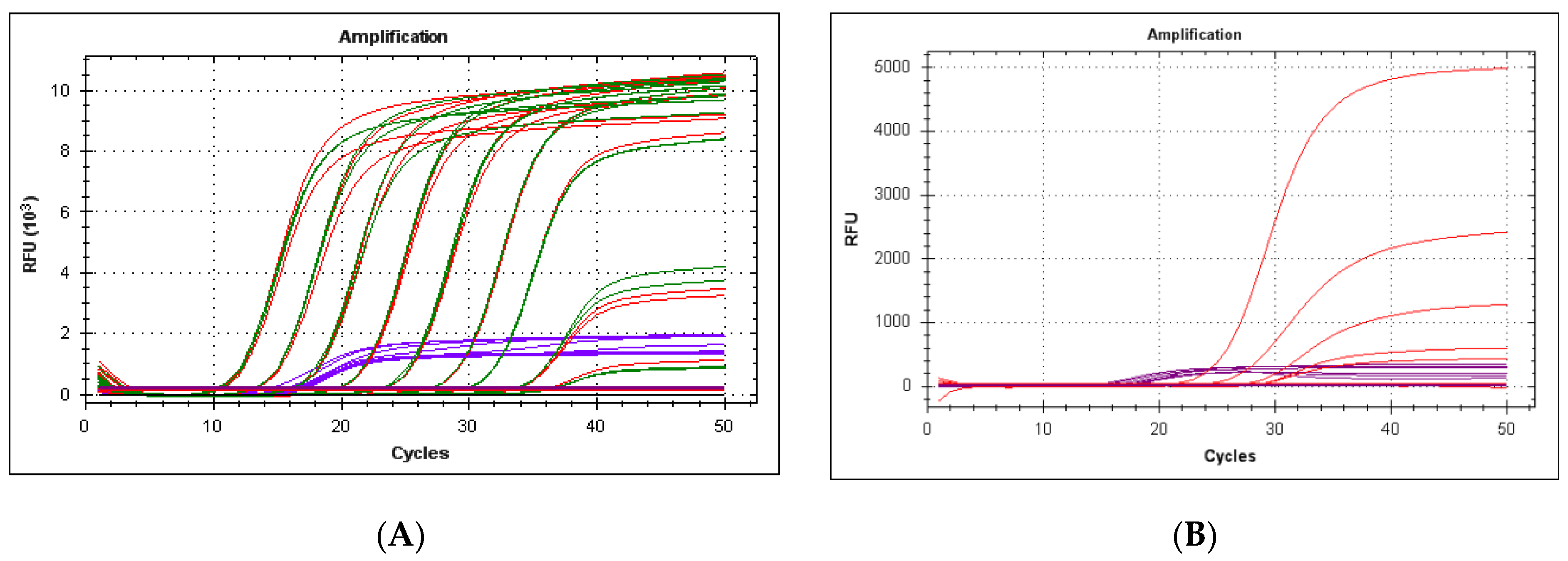
3.1. Specificity of the Method
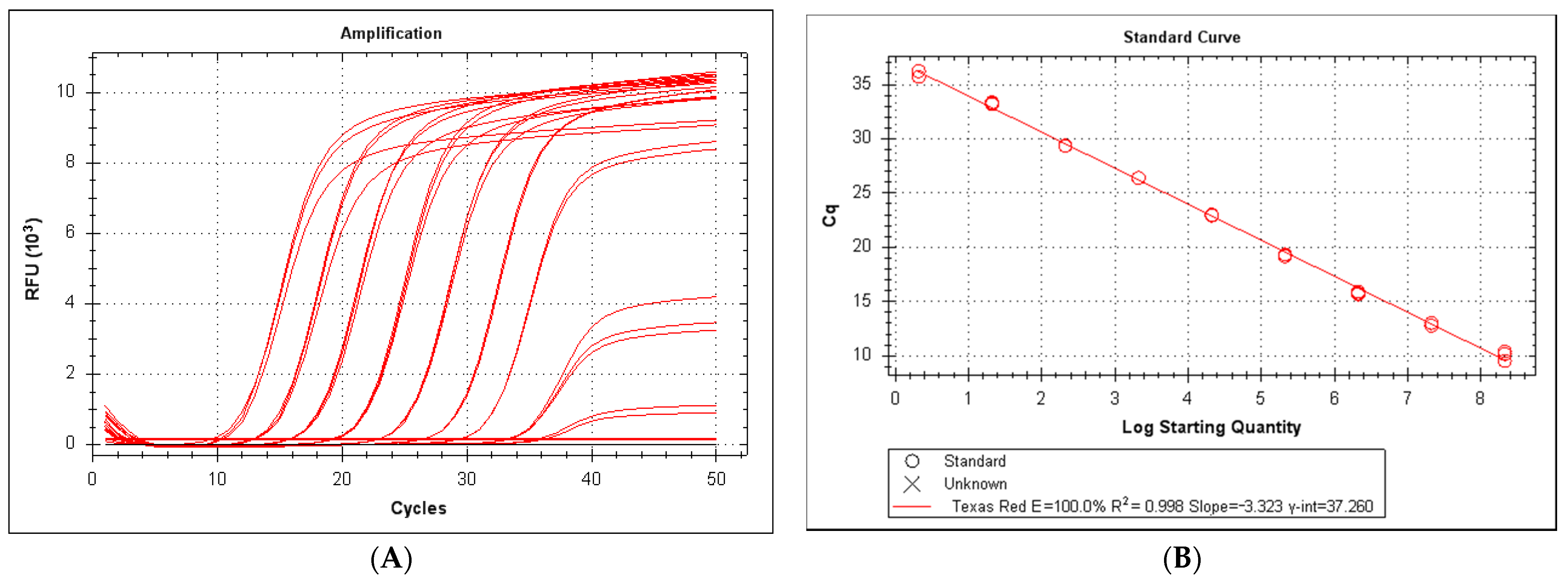
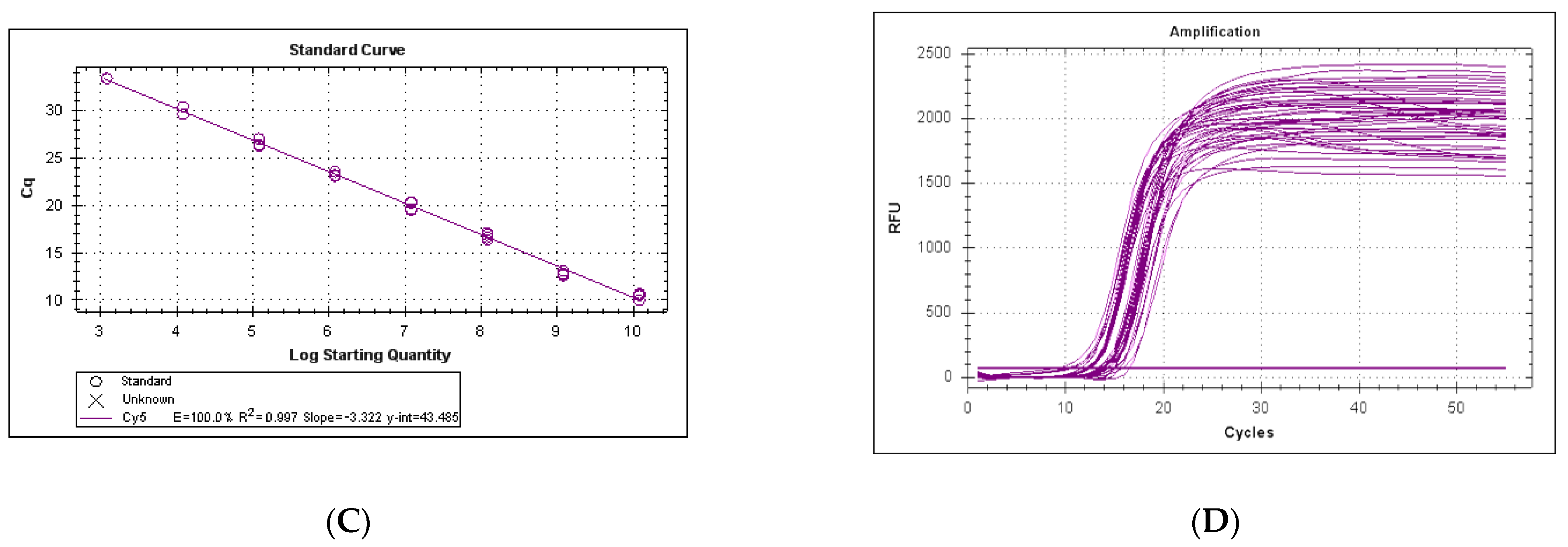
3.2. Efficiency and Sensitivity of the LeHV-5 Taqman System
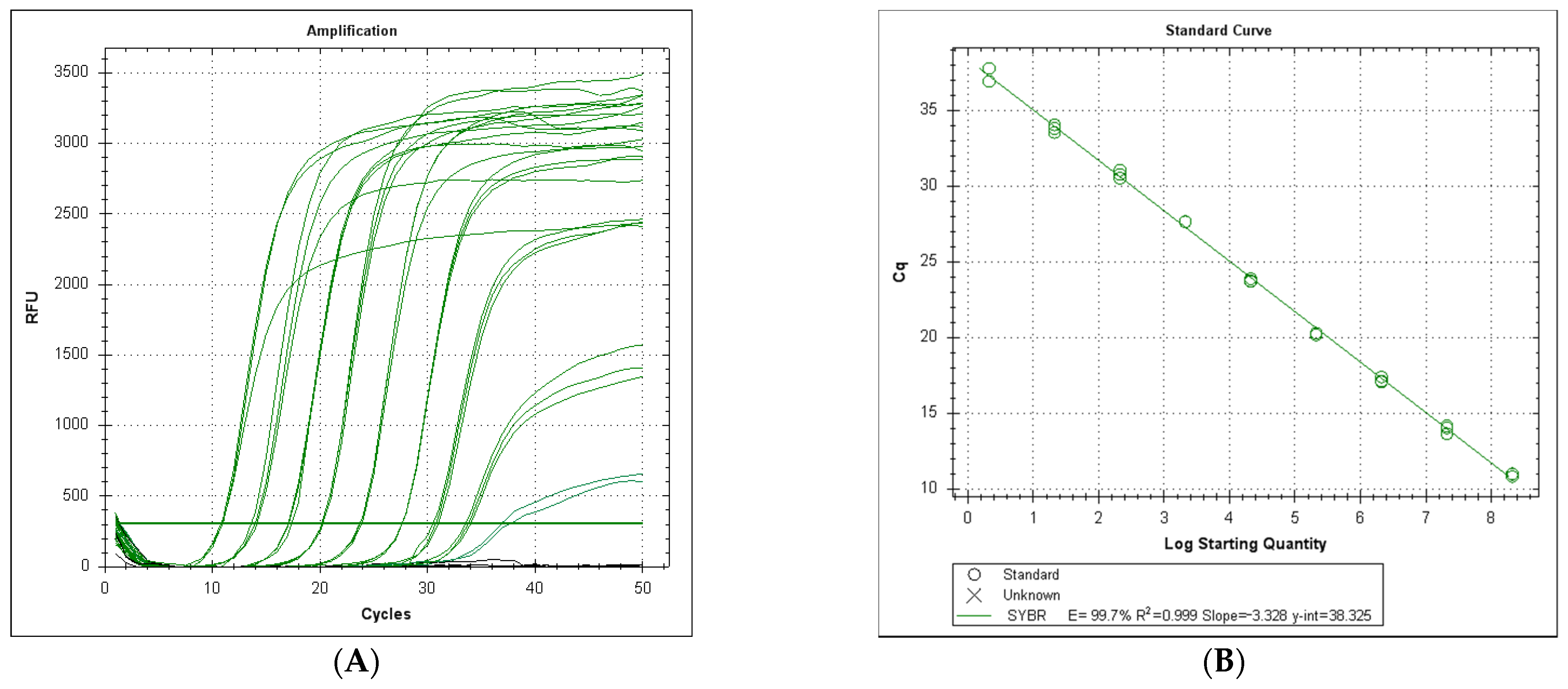
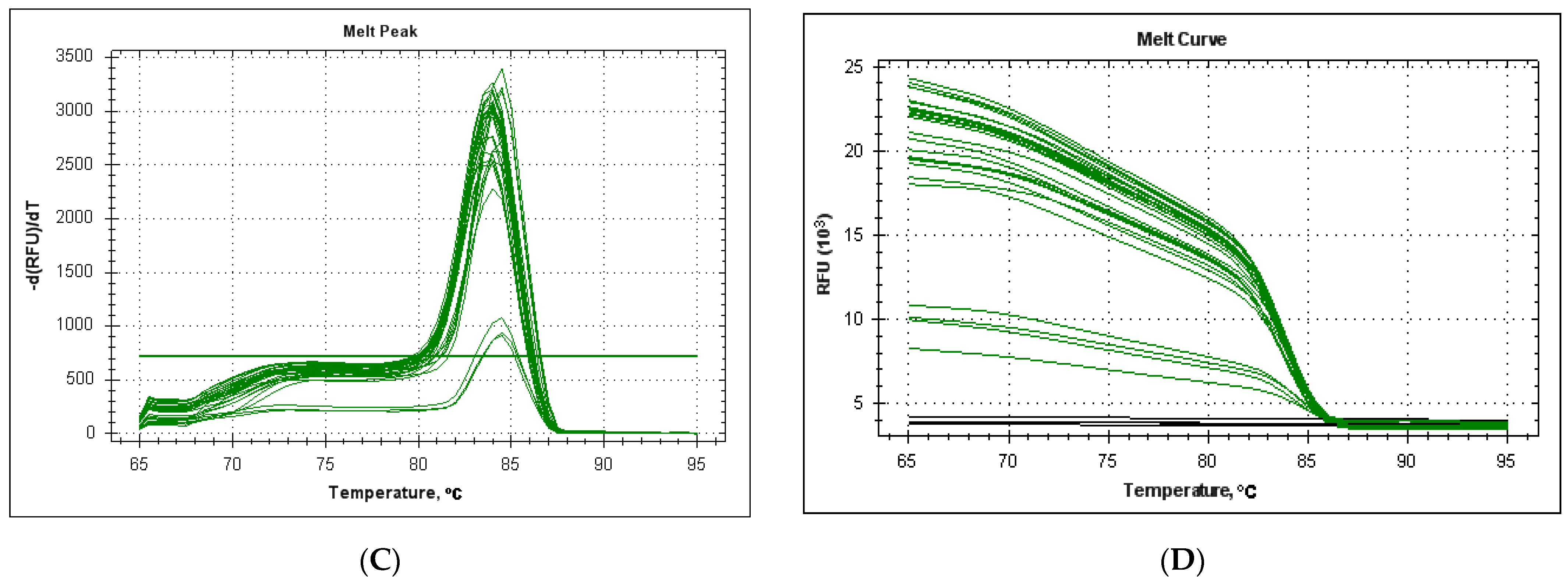
3.3. Efficiency and Sensitivity of the LeHV-5 in the EvaGreen System
3.4. Behavior of 18S rDNA as an Internal Control
3.5. Intra and Inter-Assay Reproducibility in the LeHV-5 Taqman and EvaGreen Systems
3.6. Performance in the Diagnosis of Clinical Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Declaration
Conflicts of Interest
References
- Duarte, M.D.; Carvalho, C.; Abade dos Santos, F.A.; Monteiro, J.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Tavares Santos, P.; Melo, P. The Health and Future of the Six Hare Species in Europe: A Closer Look at the Iberian Hare. In Lagomorphs; IntechOpen: London, UK, 2020; in press. [Google Scholar]
- Soriguer, R.; Carro, F. Lepus granatensis. The IUCN Red List of Threatened Species 2019. Available online: https://0-dx-doi-org.brum.beds.ac.uk/10.2305/IUCN.UK.2019-1.RLTS.T41306A2953195.en (accessed on 13 April 2021).
- Duarte, M.D.; Carvalho, C.L.; Abade dos Santos, F.A.; Gomes, J.; Alves, P.C.; Esteves, P.J.; Abrantes, J.; Lopes, A.M.; Lopes, P.M.; Serronha, A.; et al. +Coelho: Avaliação Ecossanitária das Populações Naturais de Coelho Bravo Visando o Controlo da Doença Hemorrágica Viral. Master’s Thesis, University of Lisbon, Lisboa, Portugal, 2018. [Google Scholar]
- Carvalho, C.; Abade Dos Santos, F.A.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Duarte, M.D. First cases of myxomatosis in Iberian hares (Lepus granatensis) in Portugal. Vet. Rec. Case Rep. 2020, 8. [Google Scholar] [CrossRef]
- Bocanegra, I.G.; Camacho-Sillero, L.; Risalde, M.A.; Dalton, K.; Caballero-Gómez, J.; Aguero, M.; Zorrila, I.; Gómez-Guillamón, F. First outbreak of myxomatosis in Iberian hares (Lepus granatensis). Transbound. Emerg. Dis. 2019, 1–5. [Google Scholar] [CrossRef]
- Wibbelt, G.; Frolich, K. Infectious Diseases in European Brown Hare (lepus europaeus). Wildl. Biol. Pract. 2005, 1. [Google Scholar] [CrossRef]
- Barlow, A.; Lawrence, K.; Everest, D.; Dastjerdi, A.; Finnegan, C.; Steinbach, F. Confirmation of myxomatosis in a European brown hare in Great Britain. Vet. Rec. 2014, 175, 75–76. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.J. Myxomatosis in the common hare. Ir. Vet. J. 1955, 9, 268. [Google Scholar]
- Abade dos Santos, F.A.; Monteiro, M.; Pinto, A.; Carvalho, C.L.; Peleteiro, M.C.; Carvalho, P.; Mendonça, P.; Carvalho, T.; Duarte, M.D. First description of a herpesvirus infection in genus Lepus. PLoS ONE 2020, 14, 1–20. [Google Scholar] [CrossRef]
- Kuchipudi, S.V.; Tellabati, M.; Nelli, R.K.; White, G.A.; Perez, B.B.; Sebastian, S.; Slomka, M.J.; Brookes, S.M.; Brown, I.H.; Dunham, S.P.; et al. 18S rRNA is a reliable normalisation gene for real time PCR based on influenza virus infected cells. Virol. J. 2012, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devanter, D.R.V.A.N.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M.; Black, J. Detection and Analysis of Diverse Herpesviral Species by Consensus Primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaro, N.; Amorisco, F.; Desario, C.; Lorusso, E.; Camero, M.; Bellacicco, A.L.; Sciarretta, R.; Lucente, M.S.; Martella, V.; Buonavoglia, C. Development and validation of a real-time PCR assay for specific and sensitive detection of canid herpesvirus 1. J. Virol. Methods 2010, 169, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Hoffmann, B.; Kalthoff, D.; König, P.; Beer, M. Development and validation of a triplex real-time PCR assay for the rapid detection and differentiation of wild-type and glycoprotein E-deleted vaccine strains of Bovine herpesvirus type 1. J. Virol. Methods 2011, 174, 77–84. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Targeted | Primers (Name/Nucleotide Sequence (5′–3′)) | Probe (Name/Fluorophore/Sequence (5′–3′)/Quencher) |
|---|---|---|
| gB LeHV-5 | LeHV-5-gB Fw GGACTCAGTGAACATTCACCAAAGCC LeHV-5-gB Rv CACCCACGATAAAAAAGTGCTCTGCC | LeHV-5-gB_P [Texas Red] TGCTCTCCAACACGCAGCTCGAAACATGCC [BHQ1] |
| 18S rRNA | 18S Fw TATGGTTCCTTTGGTCGCTCGCTC 18S Rv TCTGATAAATGCACGCATCCCCCC | 18S Prb [CY5] AGCTAATACATGCCGACGGGCGCTGACC [BHQ2] |
| Fresh | RT, 7 Days | 37 °C, 7 Days | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Diluent | Liver | Lung | Heart | Liver | Lung | Heart | Liver | Lung | Heart |
| PBS | 12.29 | 17.70 | 15.40 | 25.05 | 15.52 | 16.19 | 23.29 | 15.18 | 16.06 |
| PBS, AB, AM | 12.35 | 17.73 | 17.78 | 25.27 | 15.49 | 16.06 | 23.31 | 15.33 | 16.10 |
| Copies/Reaction | Taqman System | |||||
| Repeatability (Quadruplicates) | Repeatability (Three Independent Assays) | |||||
| Mean Cq | SD | %CV | Mean Cq | SD | %CV | |
| 2.1 × 108 | 10.73 | 0.181 | 1.69 | 10.32 | 0.185 | 1.79 |
| 2.1 × 107 | 13.44 | 0.093 | 0.69 | 13.55 | 0.101 | 0.75 |
| 2.1 × 106 | 16.31 | 0.061 | 0.37 | 16.10 | 0.070 | 0.43 |
| 2.1 × 105 | 20.03 | 0.086 | 0.43 | 19.86 | 0.084 | 0.42 |
| 2.1 × 104 | 23.47 | 0.114 | 0.49 | 22.85 | 0.104 | 0.46 |
| 2.1 × 103 | 27.31 | 0.080 | 0.29 | 27.44 | 0.082 | 0.30 |
| 2.1 × 102 | 30.45 | 0.222 | 0.73 | 30.11 | 0.256 | 0.85 |
| 2.1 × 101 | 34.07 | 0.159 | 0.47 | 34.05 | 0.175 | 0.51 |
| 2.1 × 100 | 36.36 | 0.897 | 2.47 | 37.13 | 0.904 | 2.43 |
| EvaGreen SYSTEM | ||||||
| Copies/reaction | Repeatability (Quadruplicates) | Repeatability (Three Independent Assays) | ||||
| Mean Cq | SD | %CV | Mean Cq | SD | %CV | |
| 2.1 × 108 | 10.19 | 0.076 | 0.75 | 10.67 | 0.090 | 0.84 |
| 2.1 × 107 | 13.31 | 0.074 | 0.56 | 13.80 | 0.114 | 0.83 |
| 2.1 × 106 | 17.18 | 0.073 | 0.42 | 16.89 | 0.094 | 0.56 |
| 2.1 × 105 | 20.70 | 0.188 | 0.91 | 20.14 | 0.250 | 1.24 |
| 2.1 × 104 | 24.29 | 0.269 | 1.11 | 23.98 | 0.314 | 1.31 |
| 2.1 × 103 | 27.91 | 0.044 | 0.16 | 27.50 | 0.098 | 0.36 |
| 2.1 × 102 | 31.30 | 0.248 | 0.79 | 31.05 | 0.322 | 1.04 |
| 2.1 × 101 | 34.89 | 0.177 | 0.51 | 34.60 | 0.198 | 0.57 |
| 2.1 × 100 | 37.63 | 0.491 | 1.30 | 37.40 | 0.503 | 1.34 |
| Sample Code | Nested PCR * | Real Time (Cq) | Estimated Copies/mg Tissue * | |
|---|---|---|---|---|
| Amplification | Sequencing Analysis | |||
| 39189PT17 | N | - | NA | NA |
| 39190PT17 | P | LeHV-5 | 20.23 | 7.43 × 106 |
| 39191PT17 | N | - | NA | NA |
| 39192PT17 | P | LeHV-5 | 22.94 | 1.14 × 106 |
| 39344PT17 | P | LeHV-5 | 24.24 | 4.61 × 105 |
| 39351PT17 | N | - | NA | NA |
| 39355PT17 | N | - | NA | NA |
| 00751PT18 | N | - | NA | NA |
| 00807PT18 | N | - | NA | NA |
| 00813PT18 | P | LeHV-5 | 24.93 | 2.86 × 105 |
| 00814PT18 | P | LeHV-5 | 23.87 | 5.96 × 105 |
| 00815PT18 | P | LeHV-5 | 22.32 | 1.75 × 106 |
| 00816PT18 | P | LeHV-5 | 25.62 | 1.77 × 105 |
| 00817PT18 | P | LeHV-5 | 23.62 | 7.09 × 105 |
| 00818PT18 | N | - | NA | NA |
| 04971PT18 | N | - | NA | NA |
| 0492PT18 | N | - | NA | NA |
| 04483PT18 | N | - | NA | NA |
| 04985PT18 | N | - | NA | NA |
| 26935PT18 | N | - | NA | NA |
| 30901PT18 | N | - | NA | NA |
| 33014PT18 | D | unspecific | NA | NA |
| 33021PT18 | D | unspecific | NA | NA |
| 35866PT18 | D | unspecific | NA | NA |
| 35867PT18 | D | unspecific | NA | NA |
| 38457PT18 | N | - | NA | NA |
| 35869PT18 | P | LeHV-5 | 21.06 | 4.18 × 106 |
| 38455PT18 | P | LeHV-5 | 21.44 | 3.21 × 106 |
| 39375PT18 | D | LeHV-5 | 26.10 | 1.27 × 105 |
| 41434PT18 | D | unspecific | NA | NA |
| 41439PT18 | D | unspecific | NA | NA |
| 41440PT18 | D | unspecific | NA | NA |
| 00129FPT19 | D | unspecific | NA | NA |
| 03230PT19 | D | unspecific | NA | NA |
| 4033PT19 | P | LeHV-5 | 25.08 | 2.58 × 105 |
| 22059PT19 | P | LeHV-5 | 26.45 | 9.97 × 104 |
| 4037PT19 | P | LeHV-5 | 25.02 | 2.69 × 105 |
| 4064PT18 | P | LeHV-5 | 14.27 | 4.63 × 108 |
| 24980PT19 | D | unspecific | NA | NA |
| 29959PT18 | P | LeHV-5 | 25.19 | 2.39 × 105 |
| 1129PT18 | P | LeHV-5 | 23.65 | 6.94 × 105 |
| 29973PT18 | P | LeHV-5 | 25.71 | 1.67 × 105 |
| 30908PT18 | D | herpesvirus | 28.36 | 2.65 × 104 |
| 33023PT18 | P | LeHV-5 | 22.57 | 1.47 × 106 |
| 30903PT18 | P | LeHV-5 | 26.59 | 9.05× 104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abade dos Santos, F.A.; Carvalho, C.L.; Peleteiro, M.C.; Parra, F.; Duarte, M.D. A Versatile qPCR for Diagnosis of Leporid Gammaherpesvirus 5 Using Evagreen® or Taqman® Technologies. Viruses 2021, 13, 715. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040715
Abade dos Santos FA, Carvalho CL, Peleteiro MC, Parra F, Duarte MD. A Versatile qPCR for Diagnosis of Leporid Gammaherpesvirus 5 Using Evagreen® or Taqman® Technologies. Viruses. 2021; 13(4):715. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040715
Chicago/Turabian StyleAbade dos Santos, Fábio A., Carina L. Carvalho, Maria C. Peleteiro, Francisco Parra, and Margarida D. Duarte. 2021. "A Versatile qPCR for Diagnosis of Leporid Gammaherpesvirus 5 Using Evagreen® or Taqman® Technologies" Viruses 13, no. 4: 715. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040715