
Virome Diversity among Mosquito Populations in a Sub-Urban Region of Marseille, France

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
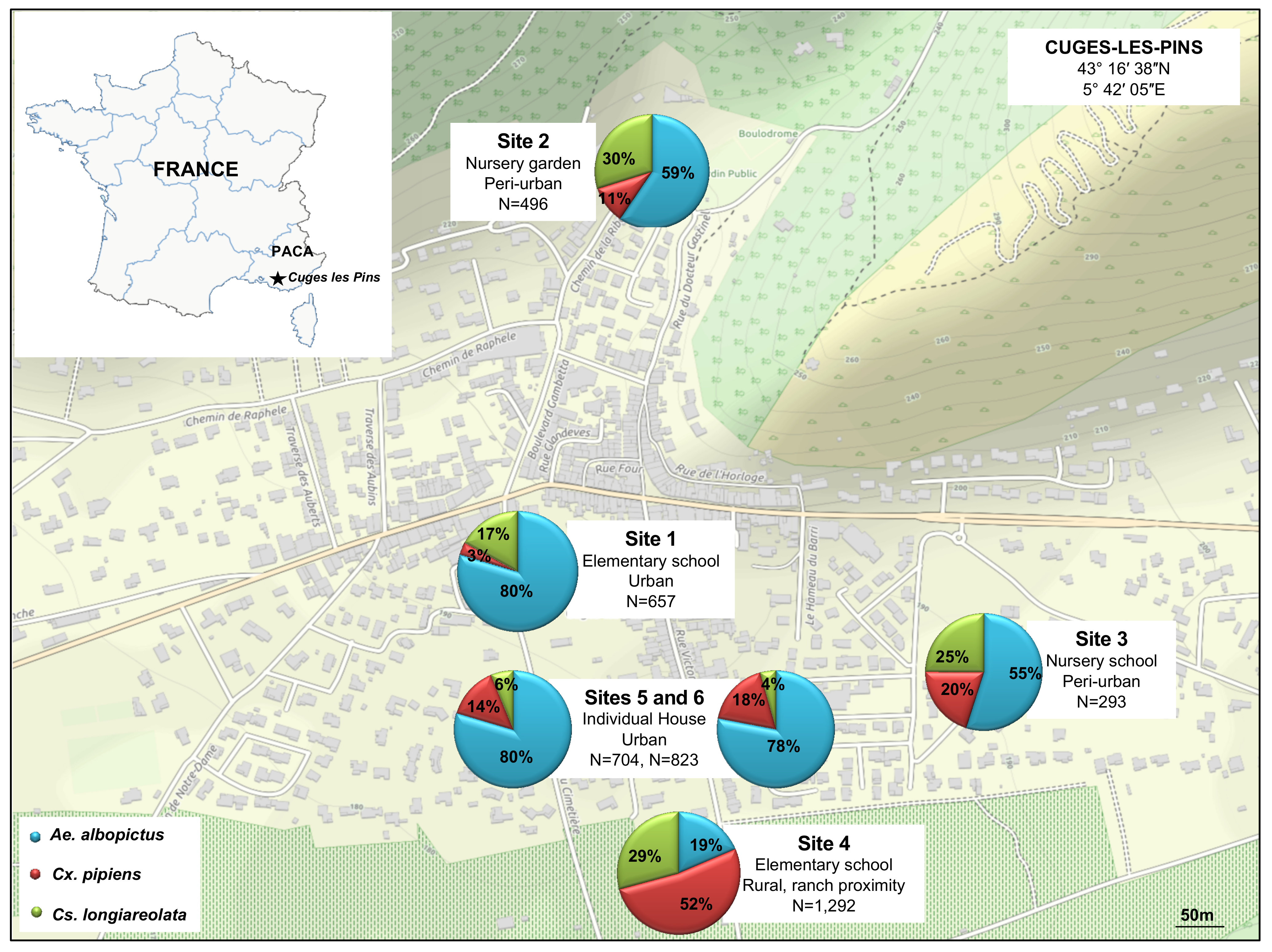
2.1. Study Area and Mosquito Collection
2.2. Morphological Mosquito Identification
2.3. MALDI-TOF MS
2.4. Virome Preparation
2.5. Transmission Electron Microscopy (TEM)
2.6. Bioinformatic Analyses of the Viromes
2.7. Phylogenetic Analyses
2.8. Isolation on Vero and C6/36 Cells
3. Results
3.1. Mosquito Collection and Identification
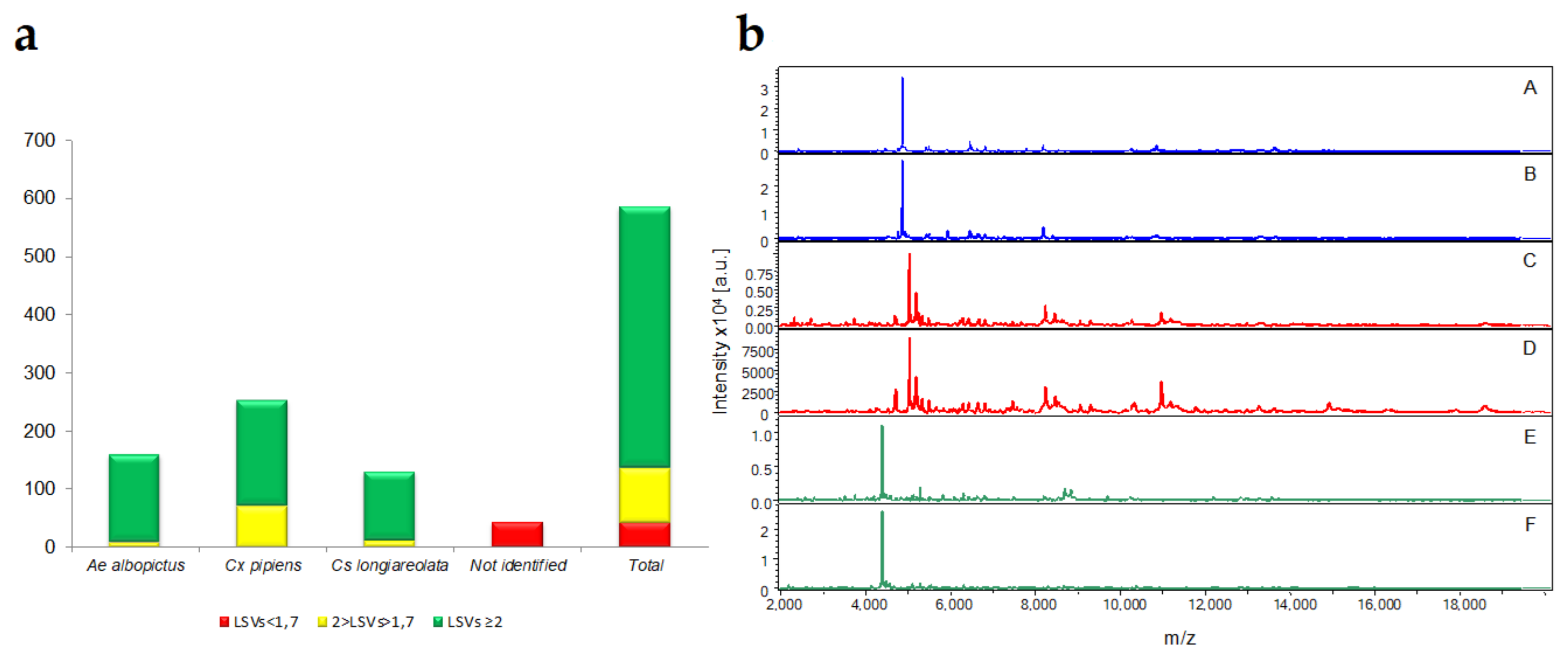
3.2. MALDI-TOF MS Analysis and Blind Tests
3.3. TEM
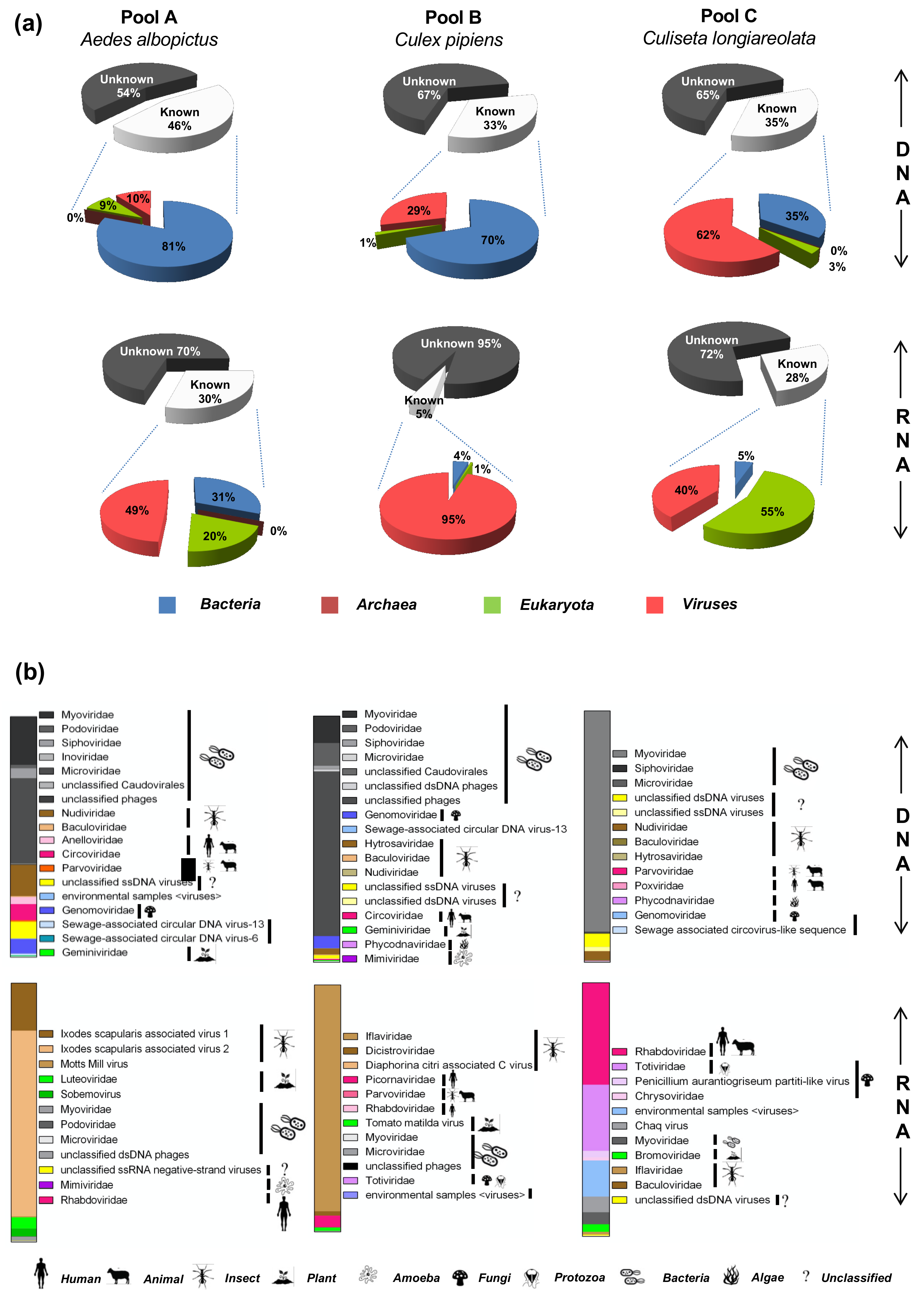
3.4. Virome Composition Based on the Taxonomic Annotation of the Sequencing Reads
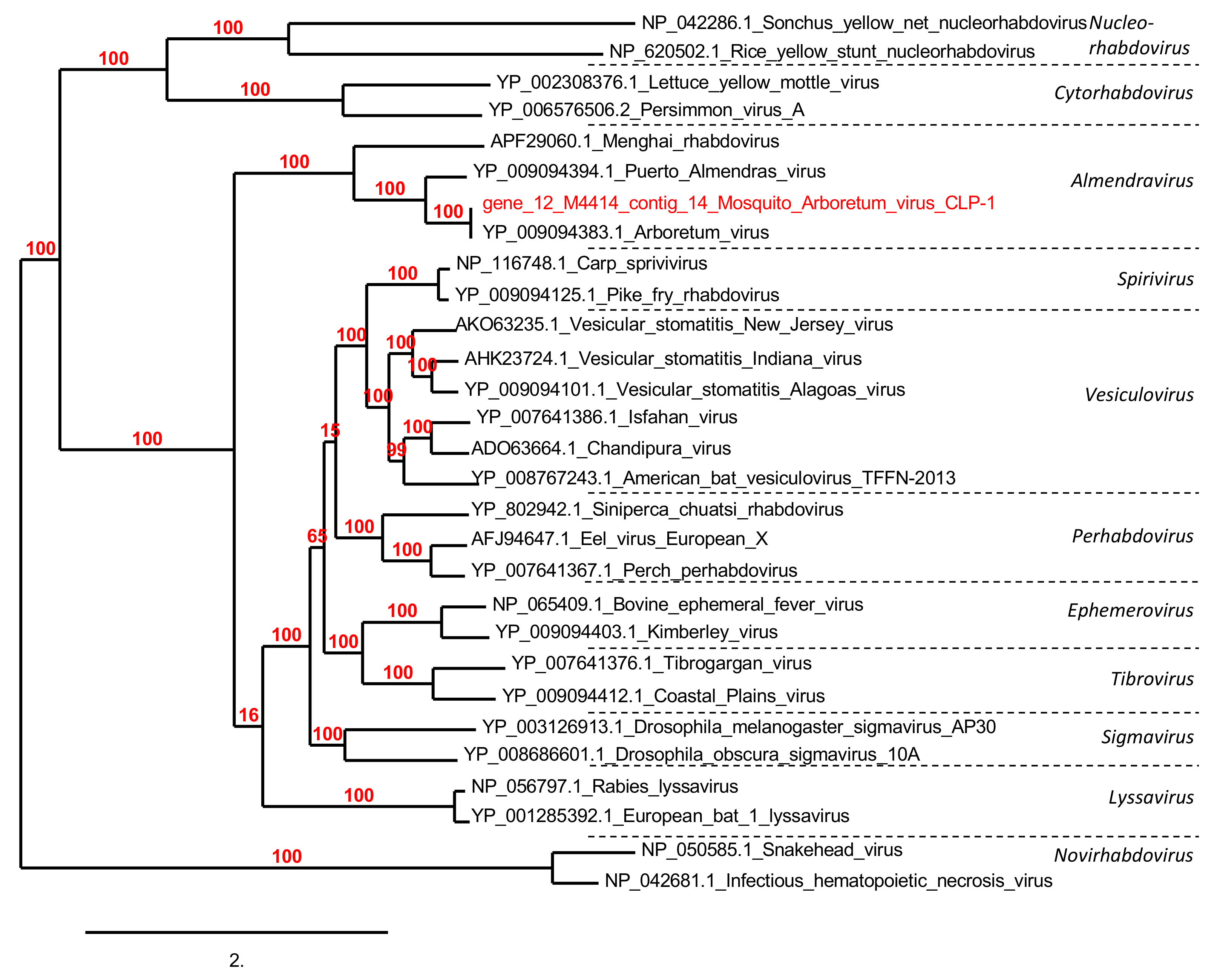
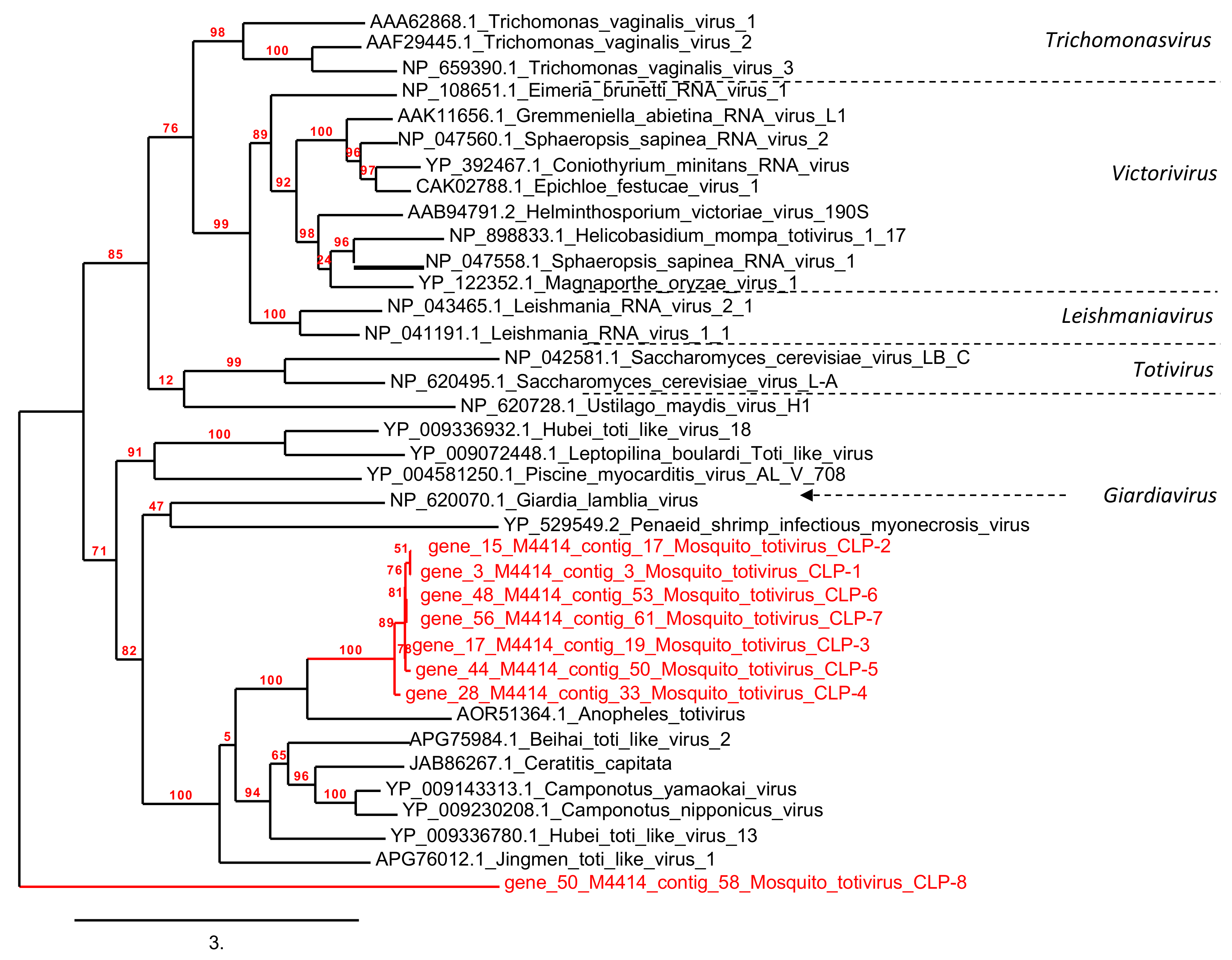
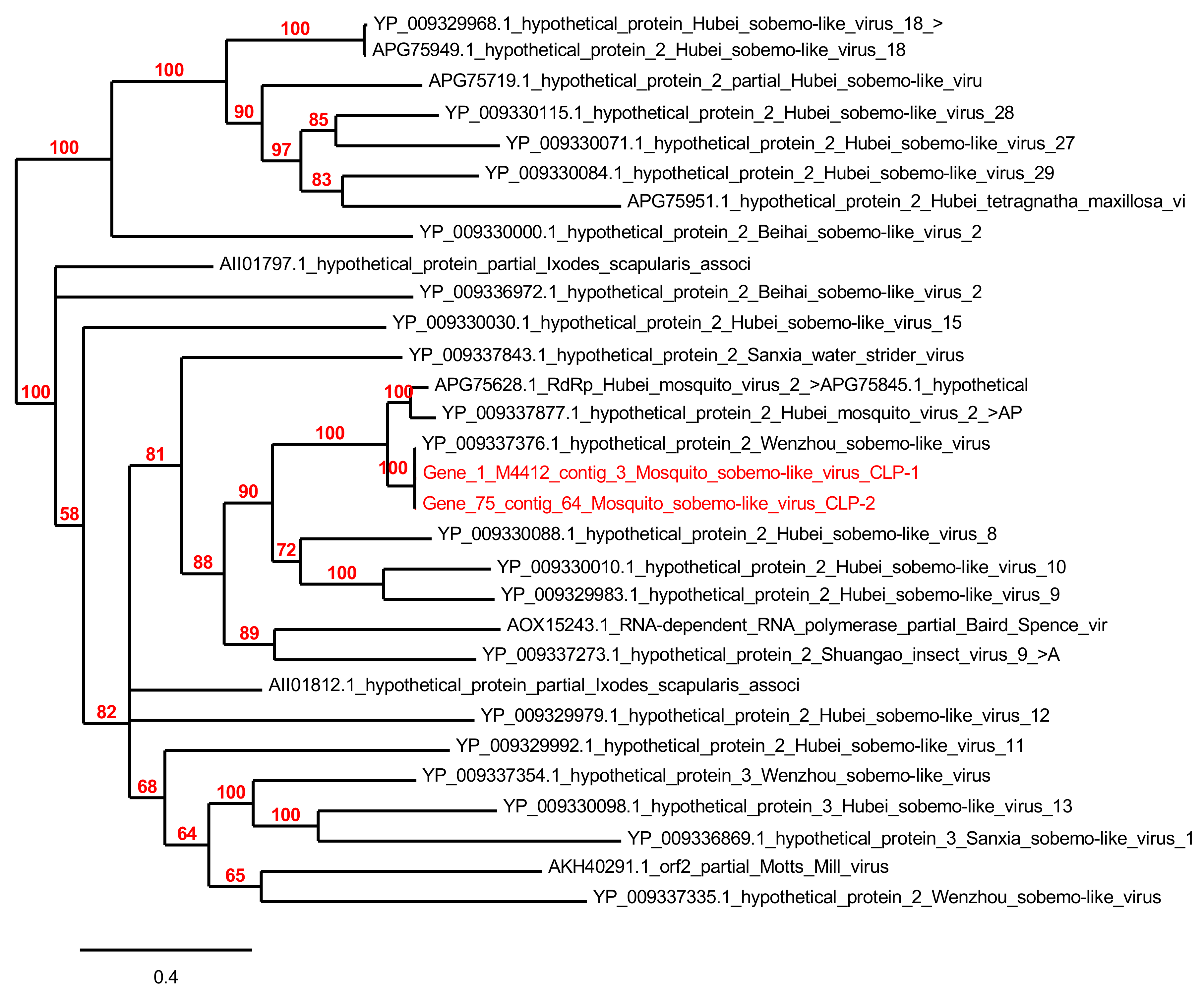
3.5. Phylogenetic Analyses
3.5.1. Viruses Associated with Animals
3.5.2. Protozoan and Fungus Viruses
3.5.3. Arthropod Viruses
3.5.4. Plant Viruses
3.5.5. Amoeba and Algae Viruses (Giant Viruses)
3.6. Virus Isolation on Vero and C6/36 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilkerson, R.C.; Linton, Y.-M.; Fonseca, D.M.; Schultz, T.R.; Price, D.C.; Strickman, D.A. Making Mosquito Taxonomy Useful: A Stable Classification of Tribe Aedini that Balances Utility with Current Knowledge of Evolutionary Relationships. PLoS ONE 2015, 10, e0133602. [Google Scholar] [CrossRef]
- Tolle, M.A. Mosquito-borne Diseases. Curr. Probl. Pediatr. Adolesc. Health Care 2009, 39, 97–140. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieme, C.; Bechah, Y.; Socolovschi, C.; Audoly, G.; Berenger, J.-M.; Faye, O.; Raoult, D.; Parola, P. Transmission potential of Rickettsia felis infection by Anopheles gambiae mosquitoes. Proc. Natl. Acad. Sci. USA 2015, 112, 8088–8093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.-P.; Tian, J.-H.; Lin, X.-D.; Ni, X.-B.; Chen, X.-P.; Liao, Y.; Yang, S.-Y.; Dumler, J.S.; Holmes, E.C.; Zhang, Y.-Z. Extensive genetic diversity of Rickettsiales bacteria in multiple mosquito species. Sci. Rep. 2016, 6, 38770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blagrove, M.S.C.; Caminade, C.; Waldmann, E.; Sutton, E.R.; Wardeh, M.; Baylis, M. Co-occurrence of viruses and mosquitoes at the vectors’ optimal climate range: An underestimated risk to temperate regions? PLOS Negl. Trop. Dis. 2017, 11, e0005604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Delaunay, P.; Jeannin, C.; Schaffner, F.; Marty, P. News on the presence of the tiger mosquito Aedes albopictus in metropolitan France. Arch. Pediatr. 2009, 16, S66–S71. [Google Scholar] [CrossRef]
- Delaunay, P.; Hubiche, T.; Blanc, V.; Perrin, Y.; Marty, P.; Del Giudice, P. Aedes albopictus in metropolitan France. Ann Dermatol. Venereol. 2012, 139, 396–401. [Google Scholar] [CrossRef]
- Succo, T.; Leparc-Goffart, I.; Ferre, J.B.; Roiz, D.; Broche, B.; Maquart, M.; Noel, H.; Catelinois, O.; Entezam, F.; Caire, D.; et al. Autochthonous dengue outbreak in Nimes, South of France, July to September 2015. Eurosurveillance 2016, 21, 30240. [Google Scholar] [CrossRef]
- Cadot, L.; Segondy, M.; Foulongne, V. Laboratory surveillance of arboviral infections in a southern France region colonized by Aedes albopictus. Epidemiol. Infect. 2016, 145, 710–714. [Google Scholar] [CrossRef] [Green Version]
- InVS. Chikungunya, Dengue et Zika—Données de la Surveillance Renforcée en France Métropolitaine en 2019. Available online: https://www.santepubliquefrance.fr/maladies-et-traumatismes/maladies-a-transmission-vectorielle/chikungunya/articles/donnees-en-france-metropolitaine/chikungunya-dengue-et-zika-donnees-de-la-surveillance-renforcee-en-france-metropolitaine-en-2019 (accessed on 30 March 2021).
- Calba, C.; Guerbois-Galla, M.; Franke, F.; Jeannin, C.; Auzet-Caillaud, M.; Grard, G.; Pigaglio, L.; Decoppet, A.; Weicherding, J.; Savaill, M.-C.; et al. Preliminary report of an autochthonous chikungunya outbreak in France, July to September 2017. Eurosurveillance 2017, 22, 17-00647. [Google Scholar] [CrossRef] [Green Version]
- Eldin, C.; Mailhe, M.; Zandotti, C.; Grard, G.; Galla, M.; Parola, P.; Brouqui, P.; Lagier, J.-C. West Nile virus outbreak in the South of France: Implications for travel medicine. Travel Med. Infect. Dis. 2019, 28, 100–101. [Google Scholar] [CrossRef]
- ECDC. Zika Virus Disease in Var Department, France. 2019. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/RRA-Zika-France-16-Oct-2019-corrected.pdf (accessed on 30 March 2021).
- ECDC. Epidemiological Update: Second Case of Locally Acquired Zika Virus Disease in Hyères, France. 2019. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-second-case-locally-acquired-zika-virus-disease-hyeres-france (accessed on 30 March 2021).
- Temmam, S.; Davoust, B.; Berenger, J.-M.; Raoult, D.; Desnues, C. Viral Metagenomics on Animals as a Tool for the Detection of Zoonoses Prior to Human Infection? Int. J. Mol. Sci. 2014, 15, 10377–10397. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, A.; Nitsche, A.; Kohl, C. Viral Metagenomics on Blood-Feeding Arthropods as a Tool for Human Disease Surveillance. Int. J. Mol. Sci. 2016, 17, 1743. [Google Scholar] [CrossRef]
- Ng, T.F.F.; Willner, D.L.; Lim, Y.W.; Schmieder, R.; Chau, B.; Nilsson, C.; Anthony, S.; Ruan, Y.; Rohwer, F.; Breitbart, M. Broad Surveys of DNA Viral Diversity Obtained through Viral Metagenomics of Mosquitoes. PLoS ONE 2011, 6, e20579. [Google Scholar] [CrossRef] [Green Version]
- Cholleti, H.; Hayer, J.; Abilio, A.P.; Mulandane, F.C.; Verner-Carlsson, J.; Falk, K.I.; Fafetine, J.M.; Berg, M.; Blomström, A.-L. Discovery of Novel Viruses in Mosquitoes from the Zambezi Valley of Mozambique. PLoS ONE 2016, 11, e0162751. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Neville, P.; Nicholson, J.; Eden, J.-S.; Imrie, A.; Holmes, E.C. High-Resolution Metatranscriptomics Reveals the Ecological Dynamics of Mosquito-Associated RNA Viruses in Western Australia. J. Virol. 2017, 91, e00680-17. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Beller, L.; Deboutte, W.; Yinda, K.C.; Delang, L.; Vega-Rúa, A.; Failloux, A.-B.; Matthijnssens, J. Stable distinct core eukaryotic viromes in different mosquito species from Guadeloupe, using single mosquito viral metagenomics. Microbiome 2019, 7, 1–20. [Google Scholar] [CrossRef]
- Tianxian, L.; Shuang, T.; Zhang, D.; Tang, S.; Zhang, Z.; Kou, Z.; Fan, Z.; Bente, D.; Zeng, C.; Li, T. Metagenomic Profile of the Viral Communities in Rhipicephalus spp. Ticks from Yunnan, China. PLoS ONE 2015, 10, e0121609. [Google Scholar] [CrossRef] [Green Version]
- Temmam, S.; Monteil-Bouchard, S.; Robert, C.; Baudoin, J.-P.; Sambou, M.; Aubadie-Ladrix, M.; Labas, N.; Raoult, D.; Mediannikov, O.; Desnues, C. Characterization of Viral Communities of Biting Midges and Identification of Novel Thogotovirus Species and Rhabdovirus Genus. Viruses 2016, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nat. Cell Biol. 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Atoni, E.; Zhao, L.; Karungu, S.; Obanda, V.; Agwanda, B.; Xia, H.; Yuan, Z. The discovery and global distribution of novel mosquito-associated viruses in the last decade (2007–2017). Rev. Med. Virol. 2019, 29, e2079. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.A.; Bielefeldt-Ohmann, H.; McLean, B.J.; O’Brien, C.A.; Colmant, A.M.; Piyasena, T.B.; Harrison, J.J.; Newton, N.D.; Barnard, R.T.; Prow, N.A.; et al. Commensal Viruses of Mosquitoes: Host Restriction, Transmission, and Interaction with Arboviral Pathogens. Evol. Bioinform. 2016, 12, EBO-S40740. [Google Scholar] [CrossRef] [PubMed]
- Yssouf, A.; Socolovschi, C.; Flaudrops, C.; Ndiath, M.O.; Sougoufara, S.; Dehecq, J.-S.; Lacour, G.; Berenger, J.-M.; Sokhna, C.S.; Raoult, D.; et al. Matrix-Assisted Laser Desorption Ionization—Time of Flight Mass Spectrometry: An Emerging Tool for the Rapid Identification of Mosquito Vectors. PLoS ONE 2013, 8, e72380. [Google Scholar] [CrossRef] [Green Version]
- Nebbak, A.; Koumare, S.; Willcox, A.C.; Berenger, J.-M.; Raoult, D.; Almeras, L.; Parola, P. Field application of MALDI-TOF MS on mosquito larvae identification. Parasitology 2017, 145, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Nebbak, A.; Willcox, A.C.; Koumare, S.; Berenger, J.-M.; Raoult, D.; Parola, P.; Fontaine, A.; Briolant, S.; Almeras, L. Longitudinal monitoring of environmental factors at Culicidae larval habitats in urban areas and their association with various mosquito species using an innovative strategy. Pest Manag. Sci. 2019, 75, 923–934. [Google Scholar] [CrossRef] [Green Version]
- Schaffner, F.; Angel, G.; Geoffroy, B.; Hervy, J.P.; Rhaiem, A.; Brunhes, J. The Mosquitoes of Europe; Institut de Recherche pour le Developpement: Marseille, France, 2001.
- Nebbak, A.; Willcox, A.C.; Bitam, I.; Raoult, D.; Parola, P.; Almeras, L. Standardization of sample homogenization for mosquito identification using an innovative proteomic tool based on protein profiling. Proteomics 2016, 16, 3148–3160. [Google Scholar] [CrossRef]
- Tandina, F.; Niaré, S.; Laroche, M.; Koné, A.K.; Diarra, A.Z.; Ongoiba, A.; Berenger, J.M.; Doumbo, O.K.; Raoult, D.; Parola, P. Using MALDI-TOF MS to identify mosquitoes collected in Mali and their blood meals. Parasitology 2018, 145, 1170–1182. [Google Scholar] [CrossRef] [Green Version]
- Temmam, S.; Monteil-Bouchard, S.; Robert, C.; Pascalis, H.; Michelle, C.; Jardot, P.; Charrel, R.; Raoult, D.; Desnues, C. Host-Associated Metagenomics: A Guide to Generating Infectious RNA Viromes. PLoS ONE 2015, 10, e0139810. [Google Scholar] [CrossRef] [Green Version]
- Froussard, P. A random-PCR method (rPCR) to construct whole cDNA library from low amounts of RNA. Nucleic Acids Res. 1992, 20, 2900. [Google Scholar] [CrossRef] [Green Version]
- Temmam, S.; Monteil-Bouchard, S.; Sambou, M.; Aubadie-Ladrix, M.; Azza, S.; Decloquement, P.; Khalil, J.Y.B.; Baudoin, J.-P.; Jardot, P.; Robert, C.; et al. Faustovirus-Like Asfarvirus in Hematophagous Biting Midges and Their Vertebrate Hosts. Front. Microbiol. 2015, 6, 1406. [Google Scholar] [CrossRef] [Green Version]
- Sime-Ngando, T.; Mignot, J.P.; Amblard, C.; Bourdier, G.; Desvilettes, C.; Quiblier-Lloberas, C. Characterization of planktonic virus-like particles in a French mountain lake: Methodological aspects and preliminary results. Ann. Limnol. 1996, 32, 259–263. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Besemer, J.; Borodovsky, M. Heuristic approach to deriving models for gene finding. Nucleic Acids Res. 1999, 27, 3911–3920. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Contreras, M.A.; Eastwood, G.; Guzman, H.; Popov, V.; Savit, C.; Uribe, S.; Kramer, L.D.; Wood, T.G.; Widen, S.G.; Fish, D.; et al. Almendravirus: A Proposed New Genus of Rhabdoviruses Isolated from Mosquitoes in Tropical Regions of the Americas. Am. J. Trop. Med. Hyg. 2016, 96, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Vasilakis, N.; Castro-Llanos, F.; Widen, S.G.; Aguilar, P.V.; Guzman, H.; Guevara, C.; Fernandez, R.; Auguste, A.J.; Wood, T.G.; Popov, V.L.; et al. Arboretum and Puerto Almendras viruses: Two novel rhabdoviruses isolated from mosquitoes in Peru. J. Gen. Virol. 2014, 95, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Fauver, J.R.; Grubaugh, N.D.; Krajacich, B.J.; Weger-Lucarelli, J.; Lakin, S.M.; Fakoli, L.S.; Bolay, F.K.; Diclaro, J.W.; Dabiré, K.R.; Foy, B.D.; et al. West African Anopheles gambiae mosquitoes harbor a taxonomically diverse virome including new insect-specific flaviviruses, mononegaviruses, and totiviruses. Virology 2016, 498, 288–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Ruche, G.; Souarès, Y.; Armengaud, A.; Peloux-Petiot, F.; Delaunay, P.; Desprès, P.; Lenglet, A.; Jourdain, F.; Leparc-Goffart, I.; Charlet, F.; et al. First two autochthonous dengue virus infections in metropolitan France, September 2010. Eurosurveillance 2010, 15, 19676. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Gallian, P.; de Lamballerie, X.; Charrel, R. First cases of autochthonous dengue fever and chikungunya fever in France: From bad dream to reality! Clin. Microbiol. Infect. 2010, 16, 1702–1704. [Google Scholar] [CrossRef] [Green Version]
- Delisle, E.; Rousseau, C.; Broche, B.; Leparc-Goffart, I.; L’Ambert, G.; Cochet, A.; Prat, C.; Foulongne, V.; Ferré, J.B.; Catelinois, O.; et al. Chikungunya outbreak in Montpellier, France, September to October 2014. Eurosurveillance 2015, 20, 21108. [Google Scholar] [CrossRef] [Green Version]
- Laroche, M.; Bérenger, J.-M.; Delaunay, P.; Charrel, R.; Pradines, B.; Berger, F.; Ranque, S.; Bitam, I.; Davoust, B.; Raoult, D.; et al. Medical Entomology: A Reemerging Field of Research to Better Understand Vector-Borne Infectious Diseases. Clin. Infect. Dis. 2017, 65, S30–S38. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Xia, H.; Wang, Y.; Shi, C.; Atoni, E.; Zhao, L.; Yuan, Z. Comparative Metagenomic Profiling of Viromes Associated with Four Common Mosquito Species in China. Virol. Sin. 2018, 33, 59–66. [Google Scholar] [CrossRef]
- Davis, B.M.; Rall, G.F.; Schnell, M.J. Everything You Always Wanted to Know About Rabies Virus (But Were Afraid to Ask). Annu. Rev. Virol. 2015, 2, 451–471. [Google Scholar] [CrossRef]
- Ergünay, K.; Brinkmann, A.; Litzba, N.; Günay, F.; Kar, S.; Öter, K.; Örsten, S.; Sarıkaya, Y.; Alten, B.; Nitsche, A.; et al. A novel rhabdovirus, related to Merida virus, in field-collected mosquitoes from Anatolia and Thrace. Arch. Virol. 2017, 162, 1903–1911. [Google Scholar] [CrossRef]
- Shahhosseini, N.; Lühken, R.; Jöst, H.; Jansen, S.; Börstler, J.; Rieger, T.; Krüger, A.; Yadouleton, A.; Campos, R.D.M.; Cirne-Santos, C.C.; et al. Detection and characterization of a novel rhabdovirus in Aedes cantans mosquitoes and evidence for a mosquito-associated new genus in the family Rhabdoviridae. Infect. Genet. Evol. 2017, 55, 260–268. [Google Scholar] [CrossRef]
- Xu, C.L.; Cantalupo, P.G.; Saenz-Robles, M.T.; Baldwin, A.; Fitzpatrick, D.; Norris, D.E.; Jackson, E.; Pipas, J.M. Draft Genome Sequence of a Novel Rhabdovirus Isolated from Deinocerites Mosquitoes. Genome Announc. 2018, 6, e01438-17. [Google Scholar] [CrossRef] [Green Version]
- Delwart, E.; Li, L. Rapidly expanding genetic diversity and host range of the Circoviridae viral family and other Rep encoding small circular ssDNA genomes. Virus Res. 2012, 164, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Garigliany, M.-M.; Börstler, J.; Jöst, H.; Badusche, M.; Desmecht, D.; Schmidt-Chanasit, J.; Cadar, D. Characterization of a novel circo-like virus in Aedes vexans mosquitoes from Germany: Evidence for a new genus within the family Circoviridae. J. Gen. Virol. 2015, 96, 915–920. [Google Scholar] [CrossRef]
- Okamoto, H.; Takahashi, M.; Nishizawa, T.; Tawara, A.; Fukai, K.; Muramatsu, U.; Naito, Y.; Yoshikawa, A. Genomic characterization of TT viruses (TTVs) in pigs, cats and dogs and their relatedness with species-specific TTVs in primates and tupaias. J. Gen. Virol. 2002, 83, 1291–1297. [Google Scholar] [CrossRef]
- Shi, C.; Liu, Y.; Hu, X.; Xiong, J.; Zhang, B.; Yuan, Z. A Metagenomic Survey of Viral Abundance and Diversity in Mosquitoes from Hubei Province. PLoS ONE 2015, 10, e0129845. [Google Scholar] [CrossRef]
- Haller, S.L.; Peng, C.; McFadden, G.; Rothenburg, S. Poxviruses and the evolution of host range and virulence. Infect. Genet. Evol. 2014, 21, 15–40. [Google Scholar] [CrossRef] [Green Version]
- Minke, J.M.; Siger, L.; Karaca, K.; Austgen, L.; Gordy, P.; Bowen, R.; Renshaw, R.W.; Loosmore, S.; Audonnet, J.C.; Nordgren, B. Recombinant canarypoxvirus vaccine carrying the prM/E genes of West Nile virus protects horses against a West Nile virus-mosquito challenge. Emerg. Control Zoonotic Viral Enceph. 2004, 221–230. [Google Scholar] [CrossRef]
- Karaca, K.; Bowen, R.; Austgen, L.; Teehee, M.; Siger, L.; Grosenbaugh, D.; Loosemore, L.; Audonnet, J.-C.; Nordgren, R.; Minke, J. Recombinant canarypox vectored West Nile virus (WNV) vaccine protects dogs and cats against a mosquito WNV challenge. Vaccine 2005, 23, 3808–3813. [Google Scholar] [CrossRef]
- Siger, L.; Bowen, R.A.; Karaca, K.; Murray, M.J.; Gordy, P.W.; Loosmore, S.M.; Audonnet, J.-C.F.; Nordgren, R.M.; Minke, J.M. Assessment of the efficacy of a single dose of a recombinant vaccine against West Nile virus in response to natural challenge with West Nile virus-infected mosquitoes in horses. Am. J. Veter Res. 2004, 65, 1459–1462. [Google Scholar] [CrossRef]
- Siger, L.; Bowen, R.; Karaca, K.; Murray, M.; Jagannatha, S.; Echols, B.; Nordgren, R.; Minke, J.M. Evaluation of the efficacy provided by a Recombinant Canarypox-Vectored Equine West Nile Virus vaccine against an experimental West Nile Virus intrathecal challenge in horses. Veter Ther. Res. Appl. Veter Med. 2006, 7, 249–256. [Google Scholar]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of lumpy skin disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugman, V.A.; Hernández-Triana, L.M.; Prosser, S.W.J.; Weland, C.; Westcott, D.G.; Fooks, A.R.; Johnson, N. Molecular species identification, host preference and detection of myxoma virus in the Anopheles maculipennis complex (Diptera: Culicidae) in southern England, UK. Parasites Vectors 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Droit, L.; Tesh, R.B.; Popov, V.L.; Little, N.S.; Upton, C.; Virgin, H.W.; Wang, D. The Genome of Yoka Poxvirus. J. Virol. 2011, 85, 10230–10238. [Google Scholar] [CrossRef] [Green Version]
- Lakis, N.S.; Li, Y.; Abraham, J.L.; Upton, C.; Blair, D.C.; Smith, S.; Zhao, H.; Damon, I.K. Novel Poxvirus Infection in an Immune Suppressed Patient. Clin. Infect. Dis. 2015, 61, 1543–1548. [Google Scholar] [CrossRef]
- Zhai, Y.; Attoui, H.; Jaafar, F.M.; Wang, H.-Q.; Cao, Y.-X.; Fan, S.-P.; Sun, Y.-X.; Liu, L.-D.; Mertens, P.P.C.; Meng, W.-S.; et al. Isolation and full-length sequence analysis of Armigeres subalbatus totivirus, the first totivirus isolate from mosquitoes representing a proposed novel genus (Artivirus) of the family Totiviridae. J. Gen. Virol. 2010, 91, 2836–2845. [Google Scholar] [CrossRef]
- Isawa, H.; Kuwata, R.; Hoshino, K.; Tsuda, Y.; Sakai, K.; Watanabe, S.; Nishimura, M.; Satho, T.; Kataoka, M.; Nagata, N.; et al. Identification and molecular characterization of a new nonsegmented double-stranded RNA virus isolated from Culex mosquitoes in Japan. Virus Res. 2011, 155, 147–155. [Google Scholar] [CrossRef]
- Colmant, A.M.G.; Etebari, K.; Webb, C.E.; Ritchie, S.A.; Jansen, C.C.; Hurk, A.F.V.D.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Asgari, S.; Hall, R.A. Discovery of new orbiviruses and totivirus from Anopheles mosquitoes in Eastern Australia. Arch. Virol. 2017, 162, 3529–3534. [Google Scholar] [CrossRef]
- Krupovic, M.; Ghabrial, S.A.; Jiang, D.; Varsani, A. Genomoviridae: A new family of widespread single-stranded DNA viruses. Arch. Virol. 2016, 161, 2633–2643. [Google Scholar] [CrossRef] [Green Version]
- Coffey, L.L.; Page, B.L.; Greninger, A.L.; Herring, B.L.; Russell, R.C.; Doggett, S.L.; Haniotis, J.; Wang, C.; Deng, X.; Delwart, E.L. Enhanced arbovirus surveillance with deep sequencing: Identification of novel rhabdoviruses and bunyaviruses in Australian mosquitoes. Virology 2014, 448, 146–158. [Google Scholar] [CrossRef]
- Bolling, B.G.; Weaver, S.C.; Tesh, R.B.; Vasilakis, N. Insect-Specific Virus Discovery: Significance for the Arbovirus Community. Viruses 2015, 7, 4911–4928. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; Van Oers, M.M.; Mowery, J.D.; Bauchan, G.R.; et al. ICTV Virus Taxonomy Profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef]
- Atoni, E.; Wang, Y.; Karungu, S.; Waruhiu, C.; Zohaib, A.; Obanda, V.; Agwanda, B.; Mutua, M.; Xia, H.; Yuan, Z. Metagenomic Virome Analysis of Culex Mosquitoes from Kenya and China. Viruses 2018, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Abd-Alla, A.M.M.; Vlak, J.M.; Bergoin, M.; Maruniak, J.E.; Parker, A.; Burand, J.P.; Jehle, J.A.; Boucias, D.G. Hytrosaviridae: A proposal for classification and nomenclature of a new insect virus family. Arch. Virol. 2009, 154, 909–918. [Google Scholar] [CrossRef]
- Kariithi, H.; Meki, I.K.; Boucias, D.G.; Abd-Alla, A.M. Hytrosaviruses: Current status and perspective. Curr. Opin. Insect Sci. 2017, 22, 71–78. [Google Scholar] [CrossRef]
- Aherfi, S.; Colson, P.; La Scola, B.; Raoult, D. Giant Viruses of Amoebas: An Update. Front. Microbiol. 2016, 7, 349. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pool A Aedes albopictus | Pool B Culex pipiens | Pool C Culiseta longiareolata | |
|---|---|---|---|
| Site 1 | 20 | 8 | 20 |
| Site 2 | 20 | 18 | 20 |
| Site 3 | 20 | 18 | 16 |
| Site 4 | 20 | 31 | 24 |
| Site 5 | 8 | 11 | 12 |
| Site 6 | 12 | 14 | 8 |
| Total | 100 | 100 | 100 |
| Pool A Ae. albopictus | Pool B Cx. pipiens | Pool C Cs. longiareolata | ||||
|---|---|---|---|---|---|---|
| DNA | RNA | DNA | RNA | DNA | RNA | |
| Raw reads (R1) | 2,120,986 | 1,453,981 | 1,547,071 | 1,806,087 | 2,581,494 | 1,405,593 |
| Trimmed reads (R1) | 2,066,242 | 1,410,512 | 1,493,456 | 1,752,390 | 2,490,631 | 1,316,476 |
| Assigned reads (R1) | 972,296 | 430,138 | 503,057 | 87,106 | 901,150 | 391,087 |
| Contigs | 13,261 | 3789 | 4873 | 342 | 9440 | 1501 |
| Average contig length (bp) | 794 | 530 | 1083 | 570 | 768 | 669 |
| ORFs | 15,141 | 3674 | 9071 | 407 | 9202 | 1653 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nebbak, A.; Monteil-Bouchard, S.; Berenger, J.-M.; Almeras, L.; Parola, P.; Desnues, C. Virome Diversity among Mosquito Populations in a Sub-Urban Region of Marseille, France. Viruses 2021, 13, 768. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050768
Nebbak A, Monteil-Bouchard S, Berenger J-M, Almeras L, Parola P, Desnues C. Virome Diversity among Mosquito Populations in a Sub-Urban Region of Marseille, France. Viruses. 2021; 13(5):768. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050768
Chicago/Turabian StyleNebbak, Amira, Sonia Monteil-Bouchard, Jean-Michel Berenger, Lionel Almeras, Philippe Parola, and Christelle Desnues. 2021. "Virome Diversity among Mosquito Populations in a Sub-Urban Region of Marseille, France" Viruses 13, no. 5: 768. https://0-doi-org.brum.beds.ac.uk/10.3390/v13050768