
Antiviral Active Compounds Derived from Natural Sources against Herpes Simplex Viruses

 , ,
, ,
Abstract
:1. Introduction
2. Antiviral Active Ingredients from Natural Sources
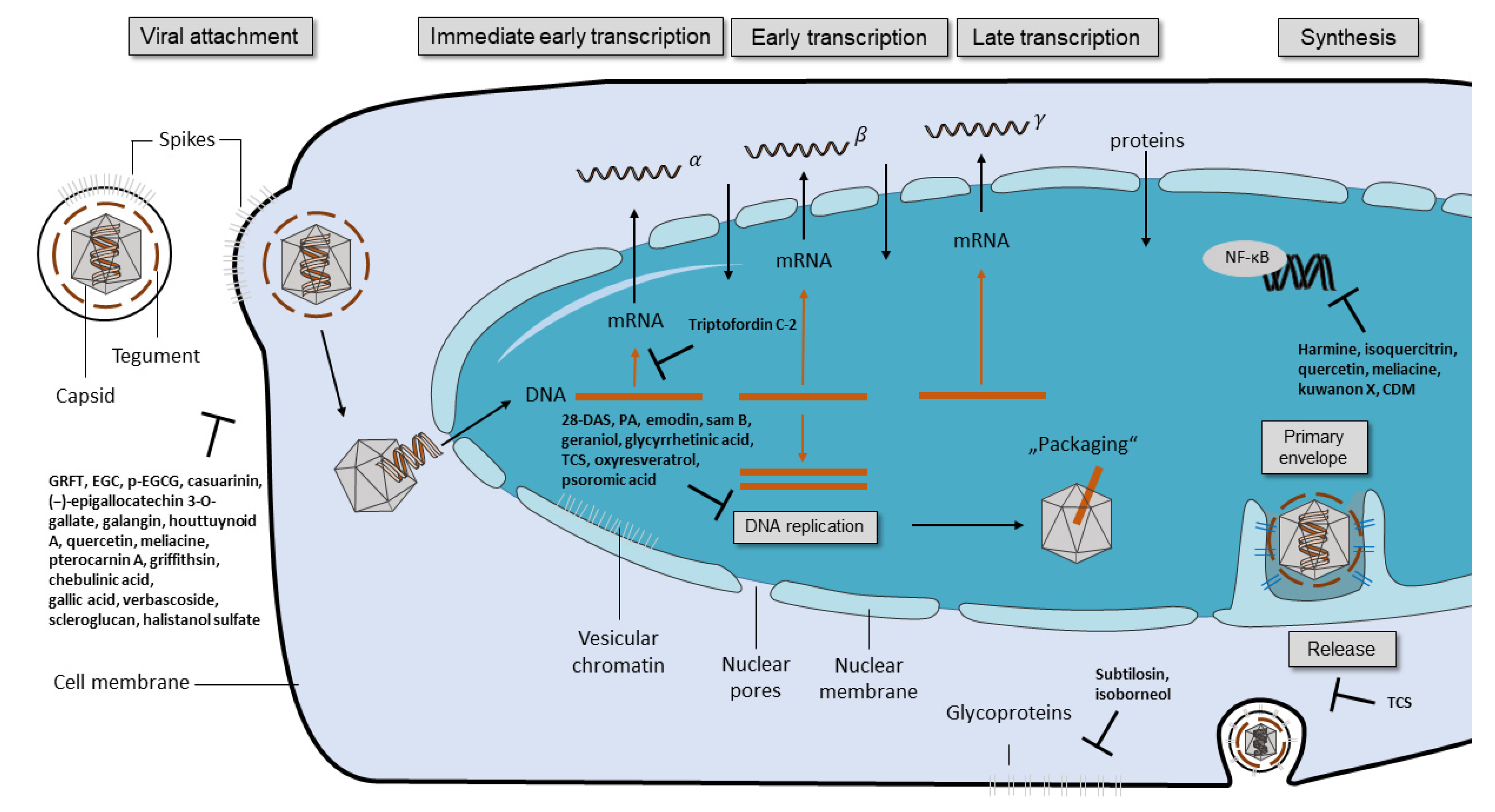
2.1. Inhibition of Viral Replication
2.2. Compounds Targeting Viral Glycoproteins
2.3. Compounds Suppressing NF-κB Activity
2.4. Compounds Affecting Viral Replication by Other Mechanisms
2.5. Efficacy of Natural Compounds In Vivo
3. Summary and Additional Comments
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pebody, R.; Andrews, N.; Brown, D.; Gopal, R.; Melker, H.; François, G.; Gatcheva, N.; Hellenbrand, W.; Jokinen, S.; Klavs, I.; et al. The seroepidemiology of herpes simplex virus type 1 and 2 in Europe. Sex. Transm. Infect. 2004, 80, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Berrington, W.R.; Jerome, K.R.; Cook, L.; Wald, A.; Corey, L.; Casper, C. Clinical correlates of herpes simplex virus viremia among hospitalized adults. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2009, 49, 1295–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimberlin, D.W.; Lin, C.Y.; Jacobs, R.F.; Powell, D.A.; Corey, L.; Gruber, W.C.; Rathore, M.; Bradley, J.S.; Diaz, P.S.; Kumar, M.; et al. Safety and efficacy of high-dose intravenous acyclovir in the management of neonatal herpes simplex virus infections. Pediatrics 2001, 108, 230–238. [Google Scholar] [CrossRef]
- Akinyi, B.; Odhiambo, C.; Otieno, F.; Inzaule, S.; Oswago, S.; Kerubo, E.; Ndivo, R.; Zeh, C. Prevalence, incidence and correlates of HSV-2 infection in an HIV incidence adolescent and adult cohort study in western Kenya. PLoS ONE 2017, 12, e0178907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.-C.; Feng, H.; Lin, Y.-C.; Guo, X.-R. New strategies against drug resistance to herpes simplex virus. Int. J. Oral Sci. 2016, 8, 1–6. [Google Scholar] [CrossRef]
- Baeten, J.M.; Strick, L.B.; Lucchetti, A.; Whittington, W.L.; Sanchez, J.; Coombs, R.W.; Magaret, A.; Wald, A.; Corey, L.; Celum, C. Herpes simplex virus (HSV)-suppressive therapy decreases plasma and genital HIV-1 levels in HSV-2/HIV-1 coinfected women: A randomized, placebo-controlled, cross-over trial. J. Infect. Dis. 2008, 198, 1804–1808. [Google Scholar] [CrossRef] [Green Version]
- Delany, S.; Mlaba, N.; Clayton, T.; Akpomiemie, G.; Capovilla, A.; Legoff, J.; Belec, L.; Stevens, W.; Rees, H.; Mayaud, P. Impact of aciclovir on genital and plasma HIV-1 RNA in HSV-2/HIV-1 co-infected women: A randomized placebo-controlled trial in South Africa. AIDS 2009, 23, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Nagot, N.; Ouédraogo, A.; Foulongne, V.; Konaté, I.; Weiss, H.A.; Vergne, L.; Defer, M.C.; Djagbaré, D.; Sanon, A.; Andonaba, J.B.; et al. Reduction of HIV-1 RNA levels with therapy to suppress herpes simplex virus. N. Engl. J. Med. 2007, 356, 790–799. [Google Scholar] [CrossRef] [Green Version]
- McMahon, M.A.; Siliciano, J.D.; Lai, J.; Liu, J.O.; Stivers, J.T.; Siliciano, R.F.; Kohli, R.M. The antiherpetic drug acyclovir inhibits HIV replication and selects the V75I reverse transcriptase multidrug resistance mutation. J. Biol. Chem. 2008, 283, 31289–31293. [Google Scholar] [CrossRef] [Green Version]
- Matamoros, T.; Kim, B.; Menéndez-Arias, L. Mechanistic insights into the role of Val75 of HIV-1 reverse transcriptase in misinsertion and mispair extension fidelity of DNA synthesis. J. Mol. Biol. 2008, 375, 1234–1248. [Google Scholar] [CrossRef] [Green Version]
- Farooq, A.V.; Shukla, D. Herpes simplex epithelial and stromal keratitis: An epidemiologic update. Surv. Ophthalmol. 2012, 57, 448–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stránská, R.; Schuurman, R.; Nienhuis, E.; Goedegebuure, I.W.; Polman, M.; Weel, J.F.; Wertheim-Van Dillen, P.M.; Berkhout, R.J.; van Loon, A.M. Survey of acyclovir-resistant herpes simplex virus in the Netherlands: Prevalence and characterization. J. Clin. Virol. 2005, 32, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Sauerbrei, A.; Bohn, K.; Heim, A.; Hofmann, J.; Weissbrich, B.; Schnitzler, P.; Hoffmann, D.; Zell, R.; Jahn, G.; Wutzler, P.H.; et al. Novel resistance-associated mutations of thymidine kinase and DNA polymerase genes of herpes simplex virus type 1 and type 2. Antivir. Ther. 2011, 16, 1297–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blot, N.; Schneider, P.; Young, P.; Janvresse, C.; Dehesdin, D.; Tron, P.; Vannier, J.P. Treatment of an acyclovir and foscarnet-resistant herpes simplex virus infection with cidofovir in a child after an unrelated bone marrow transplant. Bone Marrow Transpl. 2000, 26, 903–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Su, A.; Fu, Y.; Wang, X.; Lv, X.; Xu, W.; Xu, S.; Wang, H.; Wu, Z. Harmine blocks herpes simplex virus infection through downregulating cellular NF-κB and MAPK pathways induced by oxidative stress. Antivir. Res. 2015, 123, 27–38. [Google Scholar] [CrossRef]
- Hutterer, C.; Milbradt, J.; Hamilton, S.; Zaja, M.; Leban, J.; Henry, C.; Vitt, D.; Steingruber, M.; Sonntag, E.; Zeitträger, I.; et al. Inhibitors of dual-specificity tyrosine phosphorylation-regulated kinases (DYRK) exert a strong anti-herpesviral activity. Antivir. Res. 2017, 143, 113–121. [Google Scholar] [CrossRef]
- Xiong, H.R.; Luo, J.; Hou, W.; Xiao, H.; Yang, Z.Q. The effect of emodin, an anthraquinone derivative extracted from the roots of Rheum tanguticum, against herpes simplex virus in vitro and in vivo. J. Ethnopharmacol. 2011, 133, 718–723. [Google Scholar] [CrossRef]
- Lyu, S.Y.; Rhim, J.Y.; Park, W.B. Antiherpetic activities of flavonoids against herpes simplex virus type 1 (HSV-1) and type 2 (HSV-2) in vitro. Arch. Pharm. Res. 2005, 28, 1293–1301. [Google Scholar] [CrossRef]
- Isaacs, C.E.; Wen, G.Y.; Xu, W.; Jia, J.H.; Rohan, L.; Corbo, C.; Di Maggio, V.; Jenkins, E.C., Jr.; Hillier, S. Epigallocatechin gallate inactivates clinical isolates of herpes simplex virus. Antimicrob. Agents Chemother. 2008, 52, 962–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, A.; Adams, S.D.; Lee, L.H.; Murray, S.R.; Hsu, S.D.; Hammond, J.R.; Dickinson, D.; Chen, P.; Chu, T.C. Inhibition of herpes simplex virus type 1 with the modified green tea polyphenol palmitoyl-epigallocatechin gallate. Food Chem. Toxicol. 2013, 52, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.C.; Lin, L.C.; Tsai, W.J.; Chou, C.J.; Kung, S.H.; Ho, Y.H. Samarangenin B from Limonium sinense suppresses herpes simplex virus type 1 replication in Vero cells by regulation of viral macromolecular synthesis. Antimicrob. Agents Chemother. 2002, 46, 2854–2864. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.C.; Kuo, Y.C.; Chou, C.J. Anti-herpes simplex virus type-1 flavonoids and a new flavanone from the root of Limonium sinense. Planta Med. 2000, 66, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.Y.; Lin, C.C.; Lin, T.C. Antiherpes simplex virus type 2 activity of casuarinin from the bark of Terminalia arjuna Linn. Antivir. Res. 2002, 55, 447–455. [Google Scholar] [CrossRef]
- Luo, Z.; Kuang, X.P.; Zhou, Q.Q.; Yan, C.Y.; Li, W.; Gong, H.B.; Kurihara, H.; Li, W.X.; Li, Y.F.; He, R.R. Inhibitory effects of baicalein against herpes simplex virus type 1. Acta Pharm. Sin. B 2020, 10, 2323–2338. [Google Scholar] [CrossRef]
- Isaacs, C.E.; Xu, W.; Merz, G.; Hillier, S.; Rohan, L.; Wen, G.Y. Digallate dimers of (-)-epigallocatechin gallate inactivate herpes simplex virus. Antimicrob. Agents Chemother. 2011, 55, 5646–5653. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Liu, L.; Wu, H.; Chen, S.; Zhu, Q.; Gao, H.; Yu, X.; Wang, Y.; Su, W.; Yao, X.; et al. Anti-herpes simplex virus type 1 activity of Houttuynoid A, a flavonoid from Houttuynia cordata Thunb. Antivir. Res. 2017, 144, 273–280. [Google Scholar] [CrossRef]
- Hung, P.Y.; Ho, B.C.; Lee, S.Y.; Chang, S.Y.; Kao, C.L.; Lee, S.S.; Lee, C.N. Houttuynia cordata targets the beginning stage of herpes simplex virus infection. PLoS ONE 2015, 10, e0115475. [Google Scholar] [CrossRef] [Green Version]
- Chiang, L.C.; Chiang, W.; Liu, M.C.; Lin, C.C. In vitro antiviral activities of Caesalpinia pulcherrima and its related flavonoids. J. Antimicrob. Chemother. 2003, 52, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.Y.; Lin, T.C.; Yang, C.M.; Wang, K.C.; Lin, C.C. Mechanism of action of the suppression of herpes simplex virus type 2 replication by pterocarnin A. Microbes Infect. 2004, 6, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Bag, P.; Ojha, D.; Mukherjee, H.; Halder, U.C.; Mondal, S.; Chandra, N.S.; Nandi, S.; Sharon, A.; Sarkar, M.C.; Chakrabarti, S.; et al. An Indole Alkaloid from a Tribal Folklore Inhibits Immediate Early Event in HSV-2 Infected Cells with Therapeutic Efficacy in Vaginally Infected Mice. PLoS ONE 2013, 8, e77937. [Google Scholar] [CrossRef]
- Bag, P.; Ojha, D.; Mukherjee, H.; Halder, U.C.; Mondal, S.; Biswas, A.; Sharon, A.; Van Kaer, L.; Chakrabarty, S.; Das, G.; et al. A dihydro-pyrido-indole potently inhibits HSV-1 infection by interfering the viral immediate early transcriptional events. Antivir. Res. 2014, 105, 126–134. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Berchová-Bímová, K.; Šudomová, M.; Malaník, M.; Šmejkal, K.; Rengasamy, K.R.R. In Vitro Study of Multi-Therapeutic Properties of Thymus bovei Benth. Essential Oil and Its Main Component for Promoting Their Use in Clinical Practice. J. Clin. Med. 2018, 7, 283. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Yokomizo, K.; Okawa, M.; Tsuchihashi, R.; Kinjo, J.; Nohara, T.; Uyeda, M. Anti-herpes virus type 1 activity of oleanane-type triterpenoids. Biol. Pharm. Bull. 2005, 28, 1779–1781. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Chen, X.; Li, Q.; Li, P.; Zhao, G.; Xu, M.; Xie, P. Inhibition of intercellular adhesion in herpex simplex virus infection by glycyrrhizin. Cell Biochem. Biophys. 2012, 62, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.T.; Chen, T.Y.; Chung, C.Y.; Noyce, R.S.; Grindley, T.B.; McCormick, C.; Lin, T.C.; Wang, G.H.; Lin, C.C.; Richardson, C.D. Hydrolyzable tannins (chebulagic acid and punicalagin) target viral glycoprotein-glycosaminoglycan interactions to inhibit herpes simplex virus 1 entry and cell-to-cell spread. J. Virol. 2011, 85, 4386–4398. [Google Scholar] [CrossRef] [Green Version]
- Kesharwani, A.; Polachira, S.K.; Nair, R.; Agarwal, A.; Mishra, N.N.; Gupta, S.K. Anti-HSV-2 activity of Terminalia chebula Retz extract and its constituents, chebulagic and chebulinic acids. BMC Complement. Altern. Med. 2017, 17, 110. [Google Scholar] [CrossRef] [Green Version]
- Kratz, J.M.; Andrighetti-Fröhner, C.R.; Kolling, D.J.; Leal, P.C.; Cirne-Santos, C.C.; Yunes, R.A.; Nunes, R.J.; Trybala, E.; Bergström, T.; Frugulhetti, I.C.; et al. Anti-HSV-1 and anti-HIV-1 activity of gallic acid and pentyl gallate. Mem. Inst. Oswaldo Cruz. 2008, 103, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Kratz, J.M.; Andrighetti-Fröhner, C.R.; Leal, P.C.; Nunes, R.J.; Yunes, R.A.; Trybala, E.; Bergström, T.; Barardi, C.R.; Simões, C.M. Evaluation of anti-HSV-2 activity of gallic acid and pentyl gallate. Biol. Pharm. Bull. 2008, 31, 903–907. [Google Scholar] [CrossRef] [Green Version]
- El-Toumy, S.; Salib, J.; El-Kashak, W.; Marty, C.; Bedoux, G.; Bourgougnon, N. Antiviral effect of polyphenol rich plant extracts on herpes simplex virus type 1. Food Sci. Hum. Wellness 2018, 7, 91–101. [Google Scholar] [CrossRef]
- Kutluay, S.B.; Doroghazi, J.; Roemer, M.E.; Triezenberg, S.J. Curcumin inhibits herpes simplex virus immediate-early gene expression by a mechanism independent of p300/CBP histone acetyltransferase activity. Virology 2008, 373, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Martins, F.O.; Esteves, P.F.; Mendes, G.S.; Barbi, N.S.; Menezes, F.S.; Romanos, M.T. Verbascoside isolated from Lepechinia speciosa has inhibitory activity against HSV-1 and HSV-2 in vitro. Nat. Prod. Commun. 2009, 4, 1693–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa Cardozo, F.T.G.; Camelini, C.M.; Mascarello, A.; José Rossi, M.; José Nunes, R.; Monte Barardi, C.R.; de Mendonça, M.M.; Simões, C.M.O. Antiherpetic activity of a sulfated polysaccharide from Agaricus brasiliensis mycelia. Antivir. Res. 2011, 92, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.; Pisani, S.; Pietropaolo, V.; Seganti, L.; Nicoletti, R.; Degener, A.; Orsi, N. Antiviral effect of a polysaccharide from Sclerotium glucanicum towards herpes simplex virus type 1 infection. Planta Med. 1996, 62, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, Á.; Habtemariam, S.L.; Abdel Moneim, A.E.; Melón, S.; Dalton, K.P.; Parra, F. A spiroketal-enol ether derivative from Tanacetum vulgare selectively inhibits HSV-1 and HSV-2 glycoprotein accumulation in Vero cells. Antivir. Res. 2015, 119, 8–18. [Google Scholar] [CrossRef]
- da Rosa Guimarães, T.; Quiroz, C.G.; Borges, C.R.; de Oliveira, S.Q.; de Almeida, M.T.; Bianco, É.M.; Moritz, M.I.; Carraro, J.L.; Palermo, J.A.; Cabrera, G.; et al. Anti HSV-1 activity of halistanol sulfate and halistanol sulfate C isolated from Brazilian marine sponge Petromica citrina (Demospongiae). Mar. Drugs 2013, 11, 4176–4192. [Google Scholar] [CrossRef]
- Ma, F.; Shen, W.; Zhang, X.; Li, M.; Wang, Y.; Zou, Y.; Li, Y.; Wang, H. Anti-HSV Activity of Kuwanon X from Mulberry Leaves with Genes Expression Inhibitory and HSV-1 Induced NF-κB Deactivated Properties. Biol. Pharm. Bull. 2016, 39, 1667–1674. [Google Scholar] [CrossRef] [Green Version]
- Chuanasa, T.; Phromjai, J.; Lipipun, V.; Likhitwitayawuid, K.; Suzuki, M.; Pramyothin, P.; Hattori, M.; Shiraki, K. Anti-herpes simplex virus (HSV-1) activity of oxyresveratrol derived from Thai medicinal plant: Mechanism of action and therapeutic efficacy on cutaneous HSV-1 infection in mice. Antivir. Res. 2008, 80, 62–70. [Google Scholar] [CrossRef]
- Likhitwitayawuid, K.; Sritularak, B.; Benchanak, K.; Lipipun, V.; Mathew, J.; Schinazi, R.F. Phenolics with antiviral activity from Millettia erythrocalyx and Artocarpus lakoocha. Nat. Prod. Res. 2005, 19, 177–182. [Google Scholar] [CrossRef]
- Krawczyk, E.; Luczak, M.; Kniotek, M.; Nowaczyk, M. Cytotoxic, antiviral (in-vitro and in-vivo), immunomodulatory activity and influence on mitotic divisions of three taxol derivatives: 10-deacetyl-baccatin III, methyl (N-benzoyl-(2′R,3′S)-3′-phenylisoserinate) and N-benzoyl-(2′R,3′S)-3′-phenylisoserine. J. Pharm. Pharm. 2005, 57, 791–797. [Google Scholar] [CrossRef]
- Kim, M.; Kim, S.K.; Park, B.N.; Lee, K.H.; Min, G.H.; Seoh, J.Y.; Park, C.G.; Hwang, E.S.; Cha, C.Y.; Kook, Y.H. Antiviral effects of 28-deacetylsendanin on herpes simplex virus-1 replication. Antivir. Res. 1999, 43, 103–112. [Google Scholar] [CrossRef]
- Armaka, M.; Papanikolaou, E.; Sivropoulou, A.; Arsenakis, M. Antiviral properties of isoborneol, a potent inhibitor of herpes simplex virus type 1. Antivir. Res. 1999, 43, 79–92. [Google Scholar] [CrossRef]
- Barquero, A.A.; Michelini, F.M.; Alché, L.E. 1-Cinnamoyl-3,11-dihydroxymeliacarpin is a natural bioactive compound with antiviral and nuclear factor-kappaB modulating properties. Biochem. Biophys. Res. Commun. 2006, 344, 955–962. [Google Scholar] [CrossRef]
- Bueno, C.A.; Barquero, A.A.; Di Cónsoli, H.; Maier, M.S.; Alché, L.E. A natural tetranortriterpenoid with immunomodulating properties as a potential anti-HSV agent. Virus Res. 2009, 141, 47–54. [Google Scholar] [CrossRef]
- Alché, L.E.; Assad Ferek, G.; Meo, M.; Coto, C.E.; Maier, M.S. An Antiviral Meliacarpin from Leaves of Melia azedarach L. Z. Für. Nat. C 2003, 58, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Hayashi, T.; Ujita, K.; Takaishi, Y. Characterization of antiviral activity of a sesquiterpene, triptofordin C-2. J. Antimicrob. Chemother. 1996, 37, 759–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, S.T.S.; Šudomová, M.; Berchová-Bímová, K.; Šmejkal, K.; Echeverría, J. Psoromic Acid, a Lichen-Derived Molecule, Inhibits the Replication of HSV-1 and HSV-2, and Inactivates HSV-1 DNA Polymerase: Shedding Light on Antiherpetic Properties. Molecules 2019, 24, 2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nixon, B.; Stefanidou, M.; Mesquita, P.M.; Fakioglu, E.; Segarra, T.; Rohan, L.; Halford, W.; Palmer, K.E.; Herold, B.C. Griffithsin protects mice from genital herpes by preventing cell-to-cell spread. J. Virol. 2013, 87, 6257–6269. [Google Scholar] [CrossRef] [Green Version]
- Torres, N.I.; Noll, K.S.; Xu, S.; Li, J.; Huang, Q.; Sinko, P.J.; Wachsman, M.B.; Chikindas, M.L. Safety, formulation, and in vitro antiviral activity of the antimicrobial peptide subtilosin against herpes simplex virus type 1. Probiotics Antimicrob. Proteins 2013, 5, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Quintana, V.M.; Torres, N.I.; Wachsman, M.B.; Sinko, P.J.; Castilla, V.; Chikindas, M. Antiherpes simplex virus type 2 activity of the antimicrobial peptide subtilosin. J. Appl. Microbiol. 2014, 117, 1253–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Chan, H.; Wang, Y.Y.; Ouyang, D.Y.; Zheng, Y.T.; Tam, S.C. Trichosanthin suppresses the elevation of p38 MAPK, and Bcl-2 induced by HSV-1 infection in Vero cells. Life Sci. 2006, 79, 1287–1292. [Google Scholar] [CrossRef]
- He, D.X.; Tam, S.C. Trichosanthin affects HSV-1 replication in Hep-2 cells. Biochem. Biophys. Res. Commun. 2010, 402, 670–675. [Google Scholar] [CrossRef]
- Zheng, Y.T.; Chan, W.L.; Chan, P.; Huang, H.; Tam, S.C. Enhancement of the anti-herpetic effect of trichosanthin by acyclovir and interferon. FEBS Lett. 2001, 496, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Spear, P.G. Herpes simplex virus: Receptors and ligands for cell entry. Cell Microbiol. 2004, 6, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Däumer, M.P.; Schneider, B.; Giesen, D.M.; Aziz, S.; Kaiser, R.; Kupfer, B.; Schneweis, K.E.; Schneider-Mergener, J.; Reineke, U.; Matz, B.; et al. Characterisation of the epitope for a herpes simplex virus glycoprotein B-specific monoclonal antibody with high protective capacity. Med. Microbiol. Immunol. 2011, 200, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Hanson, J.; McLean, T.I.; Olgiate, J.; Hilton, M.; Miller, W.E.; Bachenheimer, S.L. Herpes Simplex Virus Type 1 Induction of Persistent NF-κB Nuclear Translocation Increases the Efficiency of Virus Replication. Virology 1998, 247, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Goodkin, M.L.; Ting, A.T.; Blaho, J.A. NF-κB Is Required for Apoptosis Prevention during Herpes Simplex Virus Type 1 Infection. J. Virol. 2003, 77, 7261–7280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, L.C.; Cheng, H.Y.; Liu, M.C.; Chiang, W.; Lin, C.C. In vitro anti-herpes simplex viruses and anti-adenoviruses activity of twelve traditionally used medicinal plants in Taiwan. Biol. Pharm. Bull. 2003, 26, 1600–1604. [Google Scholar] [CrossRef] [Green Version]
- Vitali, D.; Bagri, P.; Wessels, J.M.; Arora, M.; Ganugula, R.; Parikh, A.; Mandur, T.; Felker, A.; Garg, S.; Kumar, M.N.V.R.; et al. Curcumin Can Decrease Tissue Inflammation and the Severity of HSV-2 Infection in the Female Reproductive Mucosa. Int. J. Mol. Sci. 2020, 21, 337. [Google Scholar] [CrossRef] [Green Version]
- Cardozo, F.T.G.S.; Larsen, I.V.; Carballo, E.V.; Jose, G.; Stern, R.A.; Brummel, R.C.; Camelini, C.M.; Rossi, M.J.; Simões, C.M.O.; Brandt, C.R. In VivoAnti-Herpes Simplex Virus Activity of a Sulfated Derivative of Agaricus brasiliensis Mycelial Polysaccharide. Antimicrob. Agents Chemother. 2013, 57, 2541–2549. [Google Scholar] [CrossRef] [Green Version]
- Petrera, E.; Coto, C.E. Therapeutic effect of meliacine, an antiviral derived from Melia azedarach L.; in mice genital herpetic infection. Phytother. Res. 2009, 23, 1771–1777. [Google Scholar] [CrossRef]
- Alché, L.E.; Berra, A.; Veloso, M.J.; Coto, C.E. Treatment with meliacine, a plant derived antiviral, prevents the development of herpetic stromal keratitis in mice. J. Med. Virol. 2000, 61, 474–480. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
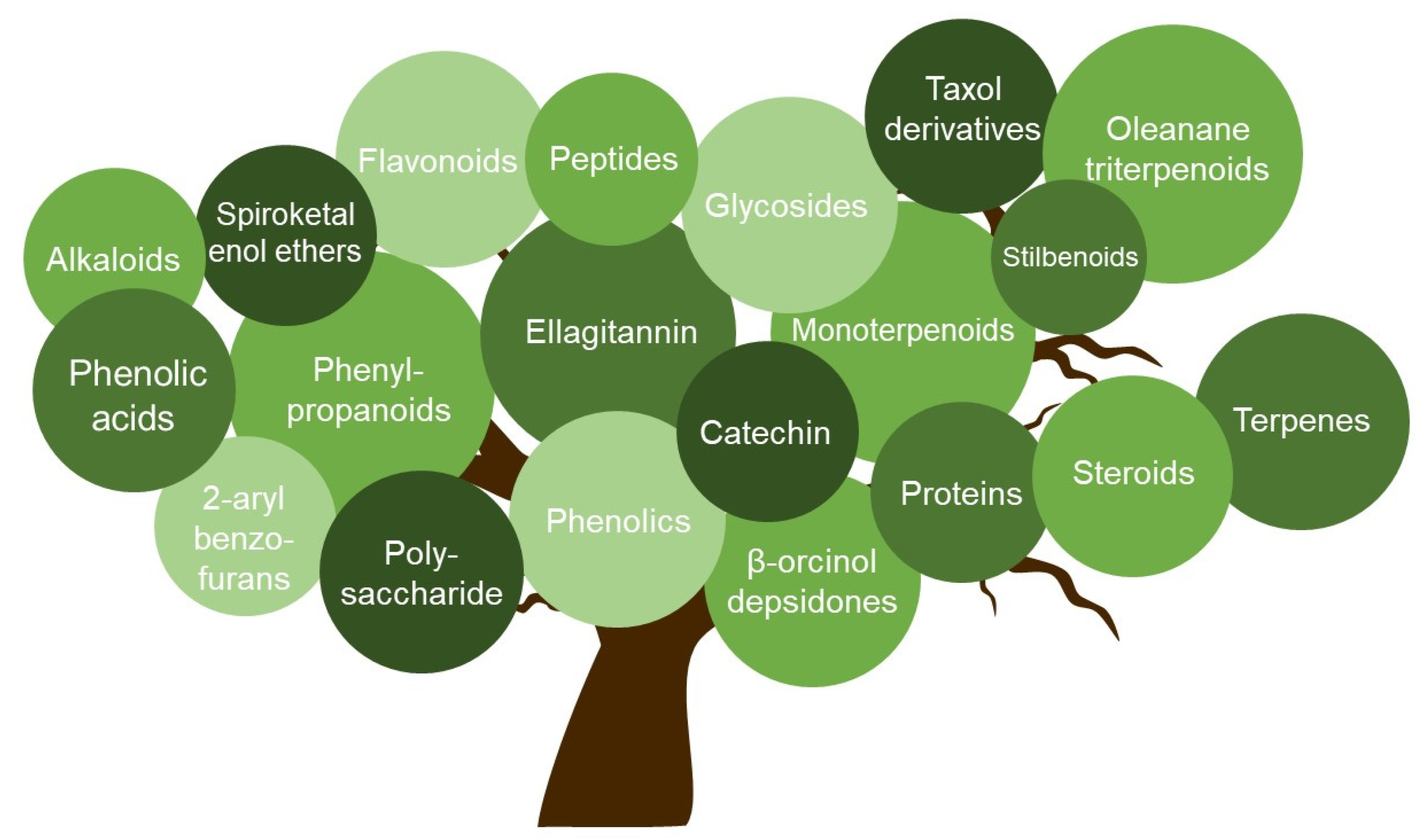
| No. | Group | Compound | Plant/Other | Assay Employed; Cell Line | CC50 | HSV-1 EC50/IC50; SI | HSV-2 EC50/IC50; SI | Virus | Mechanism of Action/Target Structure | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
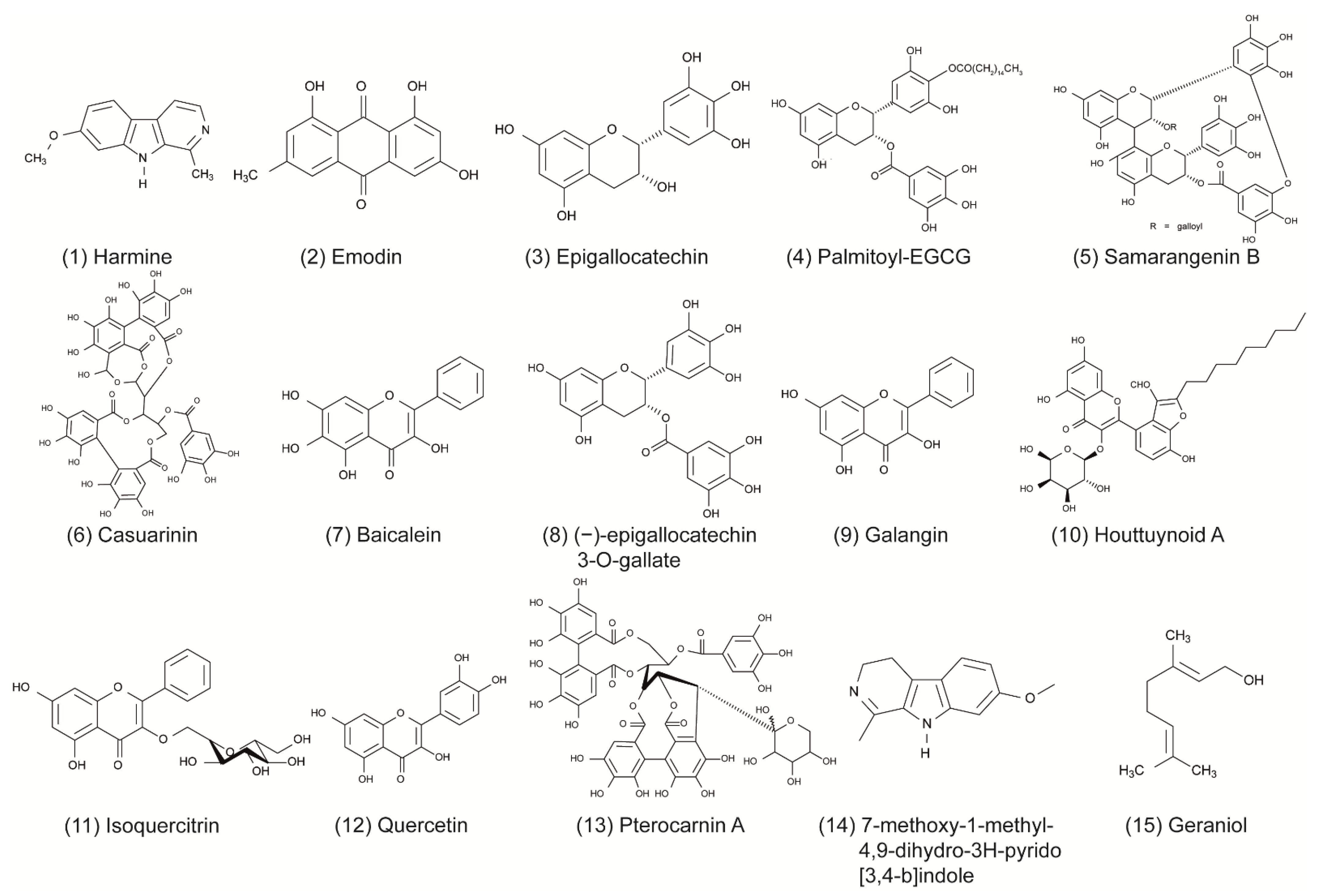
| 1 | Alkaloid | Harmine | Peganum harmala | CPE; Hec-1-A cells | >300 µM | 4.56 µM | 1.47 µM | HSV-1 F; HSV-2 G | Tyrosine phosphorylation-regulated kinase inhibitor; downregulation of cellular NF-κB and MAPK pathways. | [15,16] |
| 2 | Anthraquinone | Emodin | Rheum tanguticum | CPE; HEp-2 cells | >1000 µg/mL (>3.7 mM) | n/A | n/A | HSV-1 F; HSV-2 (333) | Inhibition of viral replication. | [17] |
| 3 | Catechin | Epigallocatechin (EGC) | Thea sinensis L. | CPE; Vero cells | 500 µM | 4 μM; 125 | 63 μM; 7.9 | HSV-1 KOS; HSV-2 G | Glycoproteins B and D. | [18,19] |
| 4 | Palmitoyl-EGCG (p-EGCG) | Plaque assay; Vero cells | >75 µM | <50 µM | n/A | HSV-1 UL46 | Glycoprotein D expression is reduced. | [20] | ||
| 5 | Samarangenin B (Sam B) | Limonium sinense L. | >100 μM | 11.4 μM | n/A | HSV-1 KOS | Suppresses expression of ICP0 and ICP4 genes and viral DNA-polymerase (ß transcripts). | [21,22] | ||
| 6 | Ellagitannin | Casuarinin | Terminalia arjuna Linn | Plaque assay; Vero cells | 89 ± 1 µM | n/A | 1.5 μM; 59 | HSV-2 (196) | Inhibition of viral attachment and penetration. | [23] |
| 7 | Flavonoid | Baicalein | Scutellaria baicalensis | Plaque assay; Vero cells | >200 µM | n/A | 12.4 µM; >16.1 | HSV-1 F | Inactivation of free viral particles and downregulating cellular NF-κB. | [24] |
| HaCat cells | >200 µM | n/A | 20.1 µM; >9.95 | |||||||
| 8 | (−)-epigallocatechin 3-O-gallate (EGCG) | Camellia sinensis | Titer reduction; Vero cells | n/A | 102.0/104.4 reduction at 100 μM | 104.0–5.0/104.0 reduction at 100 μM | HSV-1 F; HSV-2 (333) | Glycoproteins B and D. | [25] | |
| 9 | Galangin | Helichrysum aureonitens | CPE; Vero cells | 1000 µM | 2.5 μM; 400 | n/A | HSV-1 KOS | Inhibition of of viral adsorption. | [18] | |
| 10 | Houttuynoid A | Houttuynia cordata | Plaque assay; Vero cells | 166.36 ± 9.27 μM | 23.50 ± 1.82 μM | HSV-1 F | Blocking viral attachment. | [26] | ||
| 11 | Isoquercitrin | Houttuynia cordata | >100 µg/mL (215.34 µM) | 0.42 µg/mL (0.9 µM); >512.8 | Downregulation of cellular NF-κB. | [27] | ||||
| 12 | Quercetin | Caesalpinia pulcherrima | XTT; BCC-1/KMC cells | 496.9 µg/mL (16.44 mM) | 22.6 ± 4.2 µg/mL (74.78 ± 13.89 µM); 22.0 | 86.7 ± 7.4 µg/mL (286.86 ± 24.48); 5.7 | HSV-1 KOS; HSV-2 (196) | Downregulation of cellular NF-κB and blocks viral entry (gD cellular binding site). | [27,28] | |
| 13 | Glycosides | Pterocarnin A | Pterocarya stenoptera | XTT; Vero cells | 31.7 ± 1.6 μM | n/A | 5.4 ± 0.3 μM; 5.9 | HSV-2 (196) | Inhibition of of viral attachment. | [29] |
| 14 | Harmaline | 7-methoxy-1-methyl-4,9-dihydro-3H-pyrido[3,4-b]indole (HM) | Ophiorrhiza nicobarica Balkr | Plague assay; Vero cells | 30 µg/mL (120.14 µM) | 1.1 ± 0.1 µg/mL (4.41 ± 0.4 µM); 27.27 | 1.5 ± 0.1 µg/mL (6.01 ± 0.4 µM); 20 | HSV-1 F; HSV-2 G | Suppression of viral immediate early gene expression. | [30,31] |
| 15 | Monoterpenoid | Geraniol | Thymus bovei | Titer reduction; Vero cells | >210 µg/mL (1361.42 µM) | n/A | 1.92 ± 0.84 µg/mL (12.45 ± 5.45 µM); >109.38 | HSV-2 | In silico: Interacts with HSV-2 protease. | [32] |
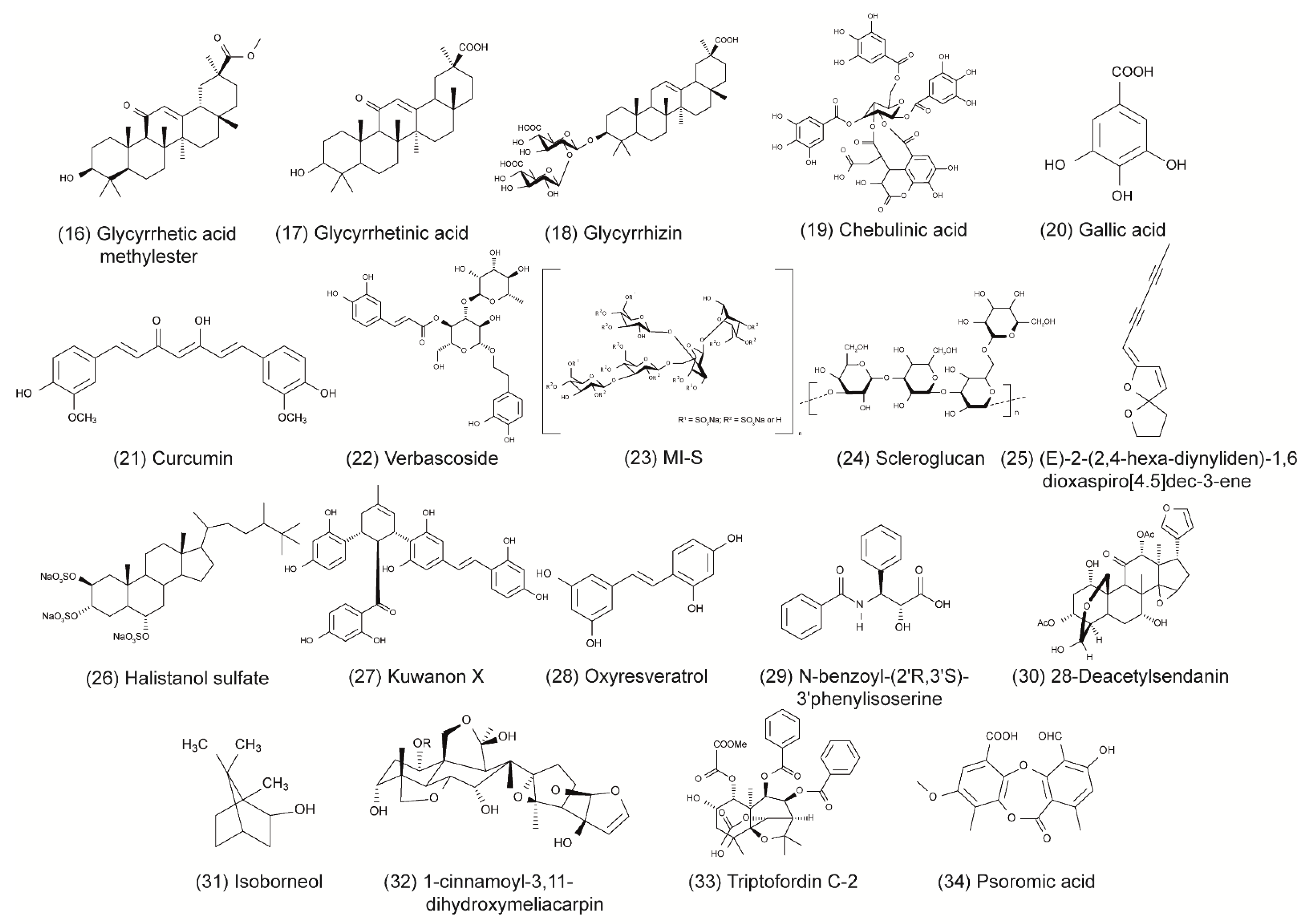
| 16 | Oleanane triterpenoid | Glycyrrhetinic acid methylester | Glycyrrhiza glabra | Plaque assay; Vero cells | >207 µM | 8.1 ± 0.2 µM/mL; >26 | n/A | HSV-1 KOS | Inhibition of HSV-1 replication. | [33] |
| 17 | Glycyrrhetinic acid | 84.0 ± 2.8 µM | 21.7 ± 0.6 µM; 3.9 | Induces the autophagy activator Beclin 1 → blocks HSV replication. | [33] | |||||
| 18 | Glycyrrhizin | >608 µM | 225 ± 24.1 µM/mL; >2.7 | Reduces adhesion force between CCECs and PMNs. | [33,34] | |||||
| 19 | Phenolic acid | Chebulinic acid | Terminalia chebula | Plaque assay; Vero cells | >200 µg/mL | 17.02 ± 2.82 µM; 18.62 | 0.06 ± 0.002 μg/mL | HSV-1 KOS; HSV-2 G | Prevention of HSV-1 glycoprotein-mediated cell fusion events and attachment of HSV-2. | [35,36] |
| 20 | Gallic acid | Galla | Plaque assay; Vero cells and GMK AH1 | 668.7 ± 54.5 μM | 57.1 ± 2.3 μM; 11.72 | 33.56; 64.35 μM (during; after infection) | HSV-1 KOS; HSV-2 (333) | Inhibition of ICP2, gC, gD, and VP5 expression (effects on viral attachment). | [37,38] | |
| 21 | Phenolic | Curcumin | The curry spice turmeric | CPE; Vero cells | 49.8 ± 0.4 µg/mL (135.18 ± 1.09 µM) | n/A | n/A | HSV-1 (17) | P300/CBP histone acetyltransferase. | [39,40] |
| 22 | Phenylpropanoid | Verbascoside | Lepechinia speciosa | Plaque assay; Vero cells | >200 µg/mL (320.21 µM) | 58 µg/mL (92.86 µM); >3.4 | 8.9 µg/mL (14.25 µM); >22.4 | HSV-1; HSV-2 (clinical isolates) | HSV-1: prevention of viral adsorption, intracellular viral inhibition; HSV-2: inhibition of attachment and penetration. | [41] |
| 23 | Polysaccharide | MI-S | Agaricus brasiliensis | Plaque assay; Vero cells | 2415.29 ± 389.21 µg/mL (134.18 ± 21.61 µM) | 1.24 ± 0.05 µg/mL (0.07 µM), 1948; 5.50 ± 0.58 µg/mL (0.31 ± 0.03 µM), 439 (during; after infection) | 0.39 ± 0.17 µg/mL (0.02 ± 0.01 µM), 6193; 4.30 ± 0.36 µg/mL (0.24 ± 0.02 µM), 562 (during; after infection) | HSV-1 KOS; HSV-2 333 | Inhibition of attachment, penetration and cell-to-cell spread. | [42] |
| 24 | Scleroglucan | Scierotium glucanicum | CPE; Vero cells | 400 µg/mL (559.83 µM) | 5 µg/mL (7 µM); 80 | n/A | HSV-1 F | Glycoproteins of HSV-1 (inhibits adsorption step). | [43] | |
| 25 | Spiroketal-enol ether derivative | (E)-2-(2,4-hexa-diynyliden)-1,6dioxaspiro[4.5] dec-3-ene | Tanacetum vulgare | Time-of-addition assay; Vero cells | >30 µg/mL (>149.83 µM) | 0.146 ± 0.013 µg/mL (0.73 ± 0.06 µM); >205 | 0.127 ± 0.009 µg/mL (0.63 ± 0.04 µM); >236 | HSV-1 (AY240815.1); HSV-2 (HM011430.1) | Suppression of viral RNA synthesis. | [44] |
| 26 | Steroid | Halistanol sulfate | Petromica citrina | Plaque assay; Vero cells | 13.83 ± 3.75 µg/mL (20.08 ± 5.44 µM) | 5.63 ± 1.3 µg/mL (8.17 ± 1.89 µM); 2.46 | n/A | HSV-1 KOS | Inhibition of attachment and penetration. Impairs HSV-1 gD and ICP27 levels. | [45] |
| 27 | Stilbenoid and 2-arylbenzofuran | Kuwanon X | Moru salba L. | Plaque assay; Vero cells | 80.3 ± 3.2 µg/mL (128.15 ± 5.12 µM) | 2.2 ± 0.1 µg/mL (3.5 ± 0.16 µM); 37 | 2.5 ± 0.3 µg/mL (3.99 ± 0.48 µM); 32 | HSV-1 (15577); HSV-2 (333) | Downregulation of cellular NF-κB and viral RNA/DNA synthesis. | [46] |
| 28 | Oxyresveratrol | Artocarpus lakoocha | >100 µM | 63.5 μM | 55.3 μM | HSV-1; HSV-2 | Inhibition of early and late replication. | [47,48] | ||
| 29 | Taxol derivative | n-benzoyl-(2’R,3’S)-3’phenylisoserine | Lactarius | CPE; Vero cells | >500 µg/mL | 21.7 µg/mL (76.06 µM); >23 | n/A | HSV-1 (McIntyre) | Inhibition of HSV-1 replication (possibly related to mitotic division). | [49] |
| 30 | Terpene | 28-Deacetylsendanin (28-DAS) | Melia azedarach | ELISA; Vero cells | >400 µg/mL (696.11 µM) | 1.46 µg/mL (2.54 µM) | n/A | HSV-1 (McIntyre) | Reduces activity of TK. | [50] |
| 31 | Isoborneol | Salvia fruticosa | Plaque assay; Vero cells | n/A | n/A | HSV-1 F | Affected TK-independent glycosylation process of viral glycoproteins B and D. | [51] | ||
| 32 | 1-cinnamoyl-3,11-dihydroxymeliacarpin (CDM) | Melia azedarach | Plaque assay; HCLE cells | >100 µM | 0.78 µM | HSV-1 KOS | Inhibition of glycoproteins B, gC, gD intracellular trafficking and downregulates cellular NF-κB. | [52,53,54] | ||
| 33 | Triptofordin C-2 | Tripterygium wilfordii | Plaque assay; HeLa cells | 89 ± 9.5 µg/mL (145.76 ± 15.56 µM) | 3.7 ± 0.90 µg/mL (6.06 ± 1.47 µM); 24 ± 3.2 | HSV-1 HF | Suppression of viral immediate early gene expression. | [55] | ||
| 34 | β-orcinol depsidone | Psoromic acid | Usnea | Plaque assay; Vero cells | >310 µM | 1.9 ± 0.42 µM; >163.2 | 2.7 ± 0.43 μM; 114.8 | HSV-1 KOS; HSV-2 (A234) | Inhibition of HSV replication: HSV-1 DNA-polymerase in vitro; HSV-2 DNA-polymerase in silico. | [56] |
| 35 | Peptide | Griffithsin | Griffithsia | Plaque assay; CaSki | no cytotoxic effect | n/A | 2.3 μg/mL (0.18 µM) | HSV-2 (333) | Inhibition of viral attachment (cell-to-cell spread). | [57] |
| 36 | Subtilosin | Bacillus amyloliquefaciens | Plaque assay; Vero cells | 314 µg/mL (92.3 µM) | 9.6 µg/mL (2.82 µM); 33 | 18.2 µg/mL (5.35 µM); 17.4 | HSV-1 F; HSV-2 G | Late stages of the viral replicative cycle and intracellular glycoprotein transport. | [58,59] | |
| 37 | Trichosanthin (TCS) | Trichosanthes kirilowii | ELISA; Vero cells | 416.5 ± 34.5 µg/mL (15.42 ± 1.28 µM) | 38.4 ± 17.5 µg/mL (1.42 ± 0.65 µM); 10.8 | n/A | HSV-1 F | Suppression of p38 MAPK protein and Bcl-2 gene activity, replication (E and L), DNA expression and viral release. | [60,61,62] |
| Group | Compound | Plant/Other | Animal | Virus | Way of Infection | Treatment; Dose | Survival under Treatment | Compared to Control/ACV | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Anthraquinone | Emodin | Rheum tanguticum | BALB/c mice | HSV-1F; HSV-2 (333) | Intracerebral | P.O.; 6.7 g/kg/day | HSV-1: 61.5% HSV-2: ≈70% | HSV-1: Untreated control: 0%/ACV: ≈20% (p < 0.01) HSV-2: Untreated control: 0%/ACV: ≈30% (p < 0.01) | [17] |
| Flavinoid | Baicalein | Scutellaria baicalensis | BALB/c mice | HSV-1 F | Intranasal | P.O.; 200 mg/kg/day | 75% | Untreated control: 33.3% (p < 0.05)/ACV: ≈90% | [24] |
| Harmaline | 7-methoxy-1-methyl-4,9-dihydro-3H-pyrido[3,4-b]indole | Ophiorrhiza nicobarica Balkr | BALB/c mice (female and male) | HSV-2 G | Genital | P.O.; 0.25 and 0.5 mg/kg 6 h after infection | 0.25 mg/kg: 45% 0.5 mg/kg: 70% | Untreated control: 5% (p < 0.05)/ACV: 80% | [30] |
| Peptide | Griffithsin (GRFT) | Griffithsia | BALB/c mice (female) | HSV-2 | Intravaginal | Intravaginal topically; 20 µL 0.1% GRFT gel pre-infection | ≈80% | Untreated control: ≈20% (p < 0.05) | [57] |
| Phenolics | Curcumin | The curry spice turmeric | BALB/c mice (female) | HSV-2 (333) | Intravaginal | Intravaginal topically; 100 µg 6 h pre-infection | 0% | 0% | [68] |
| Polysaccharids | MI-S | Agaricus brasiliensis | BALB/c mice (female) | HSV-2 (333) | Intravaginal | Intravaginal topically; 20 mg/mL 20 min pre-infection | 60% | Untreated control: 0% (p < 0.0001) | [69] |
| Stilbenoids and 2-arylbenzofurans | Oxyresveratrol | Artocarpus lakoocha | BALB/c mice (female) | HSV-1 (7401H) | Cutaneous | P.O.; 500 mg/kg 8 h pre-infection and 3x daily for 7 days after infection | 25% | Untreated control: 0% (n.s.)/ACV: 100% | [47] |
| Terpene | Meliacine (CDM) | Melia azedarach L. | BALB/c mice (female) | HSV-2 MS and G | Intravaginal | Intravaginal topically; 1 mg 2x daily for 5 days | HSV-2 MS: 20% HSV-2 G: 86% | Untreated control: HSV-2 MS: 0%; HSV-2 G: 42% | [70] |
| BALB/c mice (female and male) | HSV-1 (KOS) | Corneal | Corneal topically; 3x daily 1 day pre-infection and for 3 days after infection | Development of keratitis: 5% | Untreated control: 90% (p < 0.001) | [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van de Sand, L.; Bormann, M.; Schmitz, Y.; Heilingloh, C.S.; Witzke, O.; Krawczyk, A. Antiviral Active Compounds Derived from Natural Sources against Herpes Simplex Viruses. Viruses 2021, 13, 1386. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071386
van de Sand L, Bormann M, Schmitz Y, Heilingloh CS, Witzke O, Krawczyk A. Antiviral Active Compounds Derived from Natural Sources against Herpes Simplex Viruses. Viruses. 2021; 13(7):1386. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071386
Chicago/Turabian Stylevan de Sand, Lukas, Maren Bormann, Yasmin Schmitz, Christiane Silke Heilingloh, Oliver Witzke, and Adalbert Krawczyk. 2021. "Antiviral Active Compounds Derived from Natural Sources against Herpes Simplex Viruses" Viruses 13, no. 7: 1386. https://0-doi-org.brum.beds.ac.uk/10.3390/v13071386