
Peptide Inhibitor of Complement C1, RLS-0071, Reduces Zosteriform Spread of Herpes Simplex Virus Type 1 Skin Infection and Promotes Survival in Infected Mice

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Animals
2.2. Sequencing ACVR-HSV-1 17+
2.3. Plaque Assay
2.4. Zosteriform Infection Model and Treatments
2.5. Statistical Analyses
3. Results
3.1. RLS-0071 Has No In Vitro Antiviral Activity
3.2. Histidine Buffer and DMSO Are Neutral for HSV-1 Cutaneous Infection
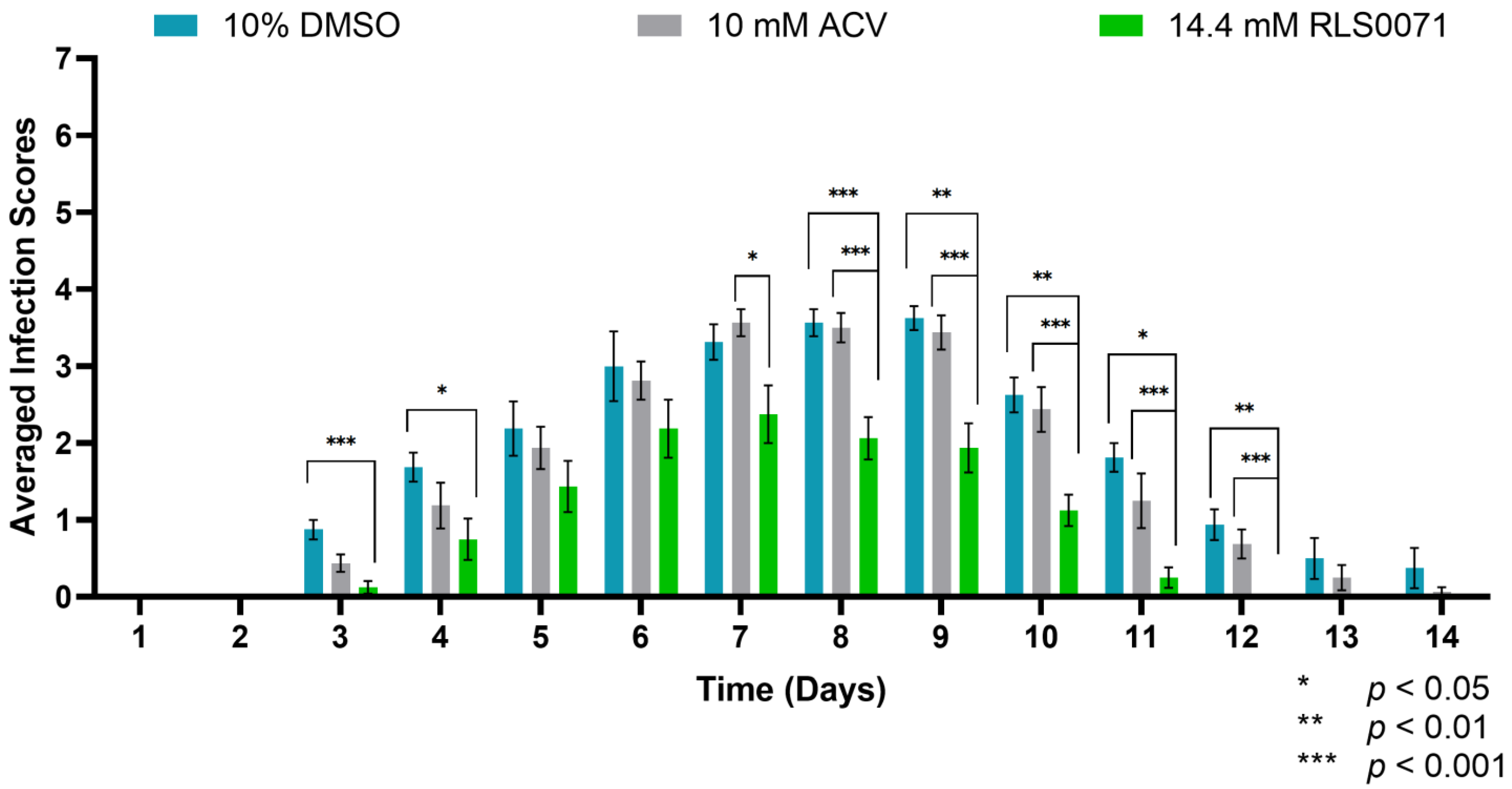
3.3. RLS-0071 Formulated in HEC Gel Protects BALB/cJ Mice Against GFP-HSV-1 Zosteriform Infection
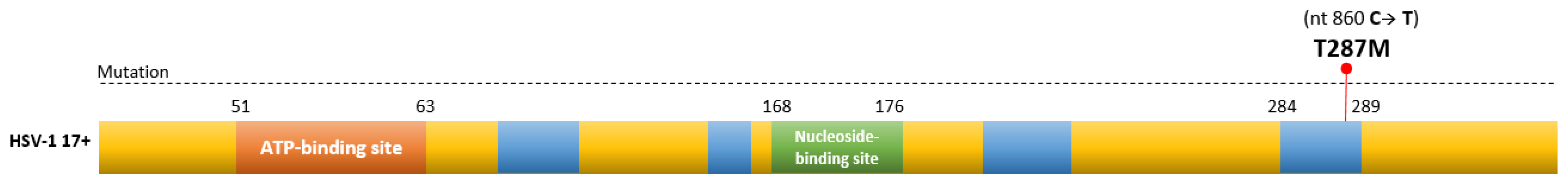
3.4. Sequencing of ACVR-HSV-1 Thymidine Kinase, UL23, Gene
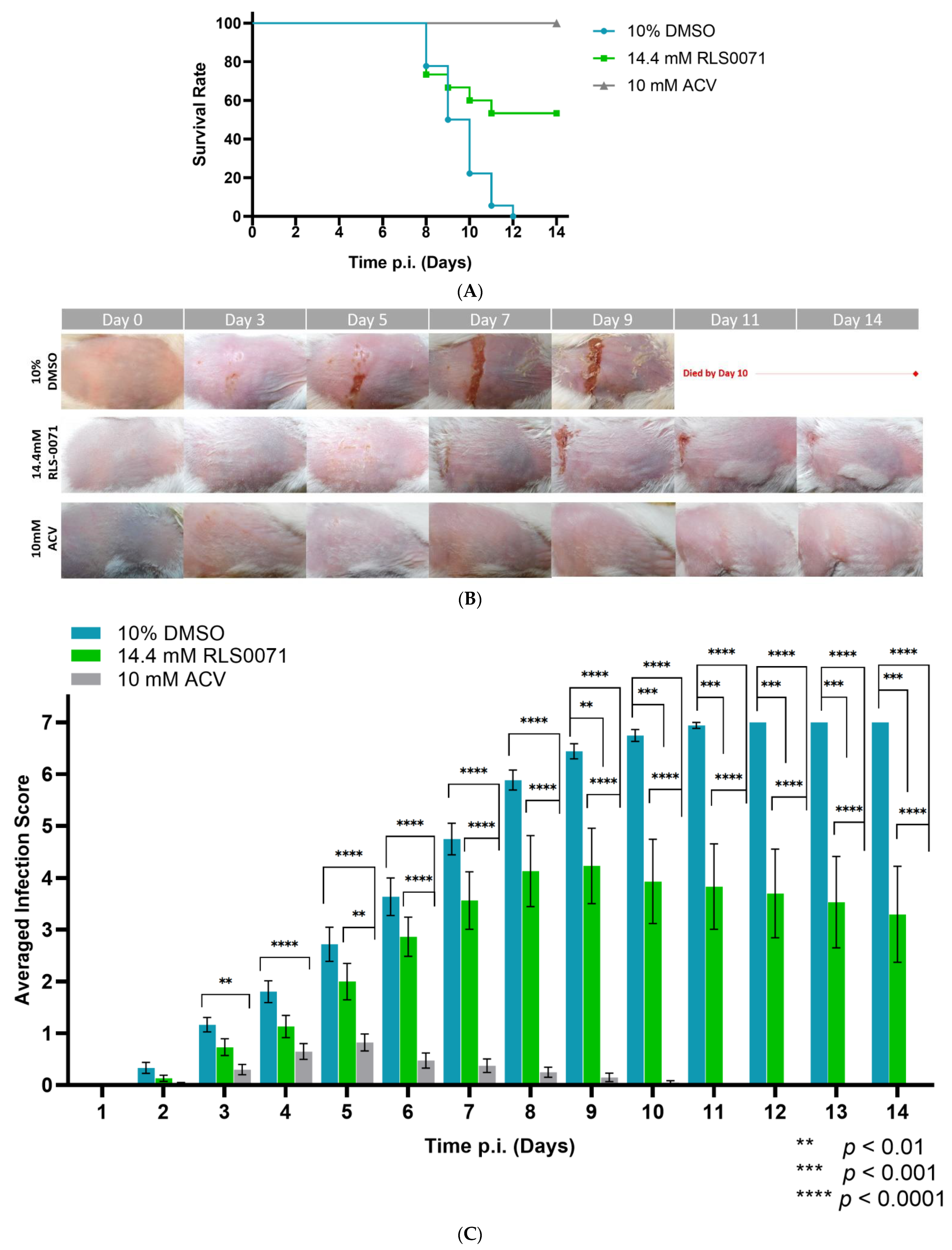
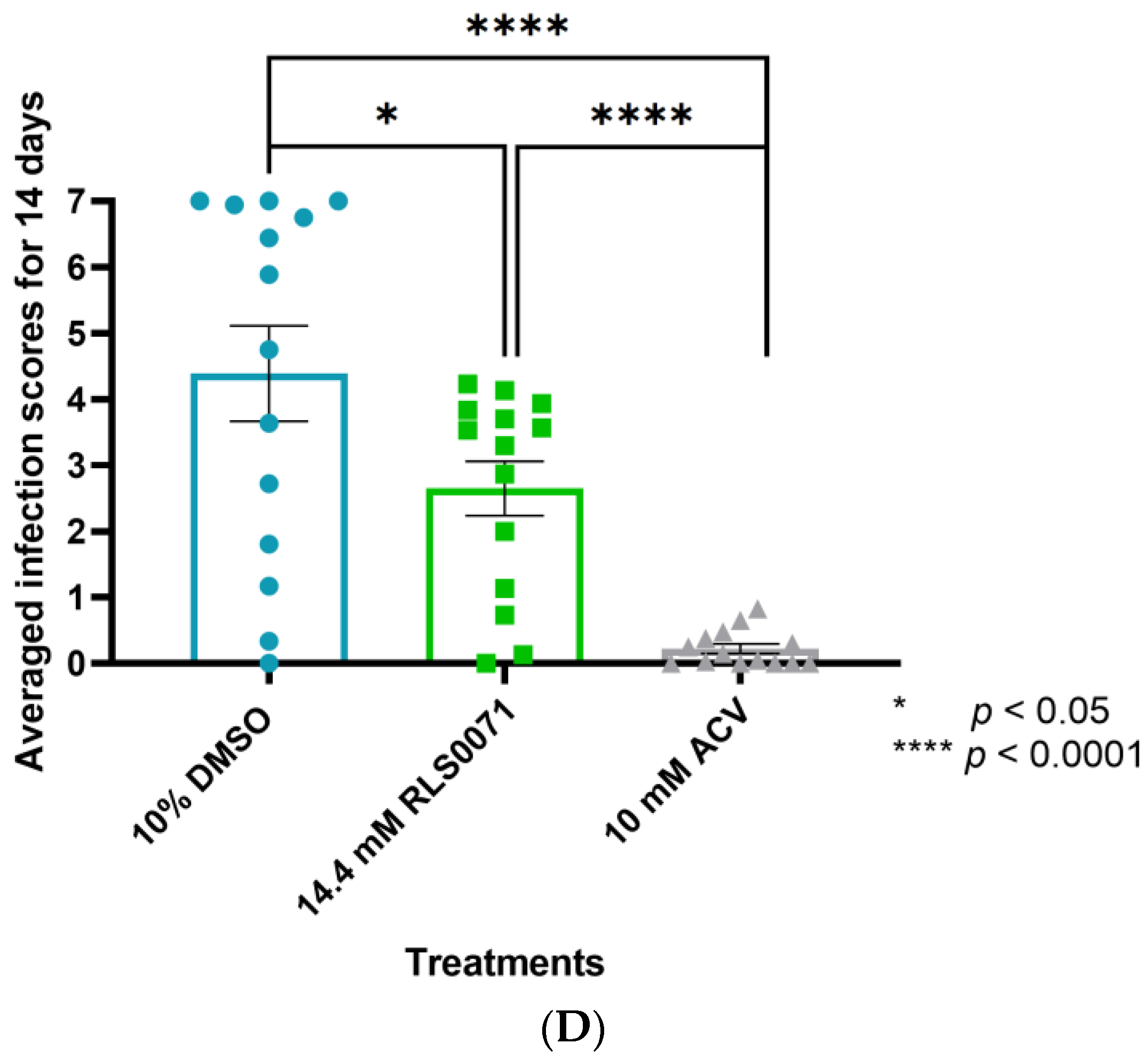
3.5. RLS-0071 Protects BALB/cJ Mice against Cutaneous ACVR-HSV-1 Strain 17+ Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Massive Proportion of World’s Population Are Living with Herpes Infection. Available online: https://www.who.int/news/item/01-05-2020-massive-proportion-world-population-living-with-herpes-infection#:~:text=An%20estimated%203.7%20billion%20people,the%20mouth%20(oral%20herpes) (accessed on 27 May 2021).
- Waisner, H.; Kalamvoki, M. The ICP0 Protein of Herpes Simplex Virus 1 (HSV-1) Downregulates Major Autophagy Adaptor Proteins Sequestosome 1 and Optineurin during the Early Stages of HSV-1 Infection. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yamamoto, T.; Aoyama, Y.; Fujimoto, W. Cell-to-cell transmission of HSV-1 in differentiated keratinocytes promotes multinucleated giant cell formation. J. Dermatol. Sci. 2019, 93, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Rahn, E.; Petermann, P.; Thier, K.; Bloch, W.; Morgner, J.; Wickström, S.A.; Knebel-Mörsdorf, D. Invasion of Herpes Simplex Virus Type 1 into Murine Epidermis: An Ex Vivo Infection Study. J. Investig. Dermatol. 2015, 135, 3009–3016. [Google Scholar] [CrossRef] [Green Version]
- Petermann, P.; Thier, K.; Rahn, E.; Rixon, F.J.; Bloch, W.; Özcelik, S.; Krummenacher, C.; Barron, M.; Dixon, M.J.; Scheu, S.; et al. Entry Mechanisms of Herpes Simplex Virus 1 into Murine Epidermis: Involvement of Nectin-1 and Herpesvirus Entry Mediator as Cellular Receptors. J. Virol. 2014, 89, 262–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, N.; Mao, H.; Rong, Q.; Docherty, J.J.; Zimmerman, D.; Rosenthal, K.S. The ability of an HSV strain to initiate zosteriform spread correlates with its neuroinvasive disease potential. Arch. Virol. 2002, 147, 763–773. [Google Scholar] [CrossRef]
- McGraw, H.M.; Awasthi, S.; Wojcechowskyj, J.A.; Friedman, H.M. Anterograde Spread of Herpes Simplex Virus Type 1 Requires Glycoprotein E and Glycoprotein I but Not Us9. J. Virol. 2009, 83, 8315–8326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frobert, E.; Ooka, T.; Cortay, J.C.; Lina, B.; Thouvenot, D.; Morfin, F. Herpes Simplex Virus Thymidine Kinase Mutations Associated with Resistance to Acyclovir: A Site-Directed Mutagenesis Study. Antimicrob. Agents Chemother. 2005, 49, 1055–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piret, J.; Boivin, G. Antiviral resistance in herpes simplex virus and varicella-zoster virus infections. Curr. Opin. Infect. Dis. 2016, 29, 654–662. [Google Scholar] [CrossRef]
- Rafail, S.; Kourtzelis, I.; Foukas, P.G.; Markiewski, M.M.; DeAngelis, R.A.; Guariento, M.; Ricklin, D.; Grice, E.; Lambris, J.D. Complement Deficiency Promotes Cutaneous Wound Healing in Mice. J. Immunol. 2015, 194, 1285–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazander, G.; Jukema, G.N.; Nibbering, P.H. Complement Activation and Inhibition in Wound Healing. Clin. Dev. Immunol. 2012, 2012, 534291. [Google Scholar] [CrossRef] [PubMed]
- Sharp, J.A.; Hair, P.S.; Pallera, H.K.; Kumar, P.S.; Mauriello, C.T.; Nyalwidhe, J.O.; Phelps, C.A.; Park, D.; Thielens, N.M.; Pascal, S.M.; et al. Peptide Inhibitor of Complement C1 (PIC1) Rapidly Inhibits Complement Activation after Intravascular Injection in Rats. PLoS ONE 2015, 10, e0132446. [Google Scholar] [CrossRef]
- Sharp, J.A.; Whitley, P.H.; Cunnion, K.M.; Krishna, N.K. Peptide Inhibitor of Complement C1, a Novel Suppressor of Classical Pathway Activation: Mechanistic Studies and Clinical Potential. Front. Immunol. 2014, 5, 406. [Google Scholar] [CrossRef] [Green Version]
- Hair, P.S.; Enos, A.I.; Krishna, N.K.; Cunnion, K.M. Inhibition of Immune Complex Complement Activation and Neutrophil Extracellular Trap Formation by Peptide Inhibitor of Complement C1. Front. Immunol. 2018, 9, 558. [Google Scholar] [CrossRef]
- Hair, P.S.; Sass, L.A.; Krishna, N.K.; Cunnion, K.M. Inhibition of Myeloperoxidase Activity in Cystic Fibrosis Sputum by Peptide Inhibitor of Complement C1 (PIC1). PLoS ONE 2017, 12, e0170203. [Google Scholar] [CrossRef] [Green Version]
- Cunnion, K.M.; Krishna, N.K.; Pallera, H.K.; Pineros-Fernandez, A.; Rivera, M.G.; Hair, P.S.; Lassiter, B.P.; Huyck, R.; Clements, M.A.; Hood, A.F.; et al. Complement Activation and STAT4 Expression Are Associated with Early Inflammation in Diabetic Wounds. PLoS ONE 2017, 12, e0170500. [Google Scholar] [CrossRef] [PubMed]
- Elliott, G.; O’Hare, P. Live-Cell Analysis of a Green Fluorescent Protein-Tagged Herpes Simplex Virus Infection. J. Virol. 1999, 73, 4110–4119. [Google Scholar] [CrossRef] [Green Version]
- Bhutta, M.S.; Shechter, O.; Gallo, E.S.; Martin, S.D.; Jones, E.; Doncel, G.F.; Borenstein, R. Ginkgolic Acid Inhibits Herpes Simplex Virus Type 1 Skin Infection and Prevents Zosteriform Spread in Mice. Viruses 2021, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Borenstein, R.; Hanson, B.A.; Markosyan, R.M.; Gallo, E.S.; Narasipura, S.D.; Bhutta, M.; Shechter, O.; Lurain, N.S.; Cohen, F.S.; Al-Harthi, L.; et al. Ginkgolic acid inhibits fusion of enveloped viruses. Sci. Rep. 2020, 10, 4746. [Google Scholar] [CrossRef] [PubMed]
- Hair, P.S.; Rivera, M.G.; Enos, A.I.; Pearsall, S.E.; Sharp, J.A.; Yousefieh, N.; Lattanzio, F.A.; Krishna, N.K.; Cunnion, K.M. Peptide Inhibitor of Complement C1 (PIC1) Inhibits Growth of Pathogenic Bacteria. Int. J. Pept. Res. Ther. 2019, 25, 83–92. [Google Scholar] [CrossRef]
- Karim, Q.A.; Karim, S.A.; Frohlich, J.A.; Grobler, A.; Baxter, C.; Mansoor, L.E.; Kharsany, A.B.M.; Sibeko, S.; Mlisana, K.; Omar, Z.; et al. Effectiveness and Safety of Tenofovir Gel, an Antiretroviral Microbicide, for the Prevention of HIV Infection in Women. Science 2010, 329, 1168–1174. [Google Scholar] [CrossRef] [Green Version]
- Tien, D.; Schnaare, R.L.; Kang, F.; Cohl, G.; McCormick, T.J.; Moench, T.R.; Doncel, G.; Watson, K.; Buckheit, R.W.; Lewis, M.G.; et al. In Vitroandin VivoCharacterization of a Potential Universal Placebo Designed for Use in Vaginal Microbicide Clinical Trials. AIDS Res. Hum. Retroviruses 2005, 21, 845–853. [Google Scholar] [CrossRef]
- Sellar, R.S.; Peggs, K.S. Management of multidrug-resistant viruses in the immunocompromised host. Br. J. Haematol. 2011, 156, 559–572. [Google Scholar] [CrossRef]
- Sauerbrei, A.; Bohn, K.; Heim, A.; Hofmann, J.; Weiβbrich, B.; Schnitzler, P.; Hoffmann, D.; Zell, R.; Jahn, G.; Wutzler, P.; et al. Novel resistance-associated mutations of thymidine kinase and DNA polymerase genes of herpes simplex virus type 1 and type 2. Antivir. Ther. 2011, 16, 1297–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauerbrei, A.; Bohn-Wippert, K.; Kaspar, M.; Krumbholz, A.; Karrasch, M.; Zell, R. Database on natural polymorphisms and resistance-related non-synonymous mutations in thymidine kinase and DNA polymerase genes of herpes simplex virus types 1 and 2. J. Antimicrob. Chemother. 2015, 71, 6–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betz, U.A.K.; Fischer, R.; Kleymann, G.; Hendrix, M.; Rubsamen-Waigmann, H. Potent in Vivo Antiviral Activity of the Herpes Simplex Virus Primase-Helicase Inhibitor BAY 57-1293. Antimicrob. Agents Chemother. 2002, 46, 1766–1772. [Google Scholar] [CrossRef] [Green Version]
- Stock, A.T.; Smith, J.; Carbone, F.R. Type I IFN suppresses Cxcr2 driven neutrophil recruitment into the sensory ganglia during viral infection. J. Exp. Med. 2011, 11, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Paludan, S.R.; Bowie, A.G.; Horan, K.A.; Fitzgerald, K.A. Recognition of herpesviruses by the innate immune system. Nat. Rev. Immunol. 2013, 11, 143–154. [Google Scholar] [CrossRef]
- Filtjens, J.; Roger, A.; Quatrini, L.; Wieduwild, E.; Gouilly, J.; Hoeffel, G.; Rossignol, R.; Daher, C.; Debroas, G.; Henri, S.; et al. Nociceptive sensory neurons promote CD8 T cell responses to HSV-1 infection. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hor, J.L.; Heath, W.; Mueller, S.N. Neutrophils are dispensable in the modulation of T cell immunity against cutaneous HSV-1 infection. Sci. Rep. 2017, 7, srep41091. [Google Scholar] [CrossRef]
- Lei, V.; Petty, A.J.; Atwater, A.R.; Wolfe, S.A.; MacLeod, A.S. Skin Viral Infections: Host Antiviral Innate Immunity and Viral Immune Evasion. Front. Immunol. 2020, 11, 2791. [Google Scholar] [CrossRef]
- Wang, L.; Wang, R.; Xu, C.; Zhou, H. Pathogenesis of Herpes Stromal Keratitis: Immune Inflammatory Response Mediated by Inflammatory Regulators. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Wojtasiak, M.; Pickett, D.L.; Tate, M.D.; Bedoui, S.; Job, E.R.; Whitney, P.G.; Brooks, A.G.; Reading, P.C. Gr-1+ cells, but not neutrophils, limit virus replication and lesion development following flank infection of mice with herpes simplex virus type-1. Virology 2010, 407, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, S.-L.; Chiang, H.-H.; Wu, C.-Y.; Hsu, M.-J.; Chen, Y.-T. Effects of herpes simplex virus type 1 infection on immune functions of human neutrophils. J. Periodontal Res. 2012, 47, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Gaudreau, A.; Hill, E.; Balfour, H.H.; Erice, A.; Boivin, G. Phenotypic and Genotypic Characterization of Acyclovir-Resistant Herpes Simplex Viruses from Immunocompromised Patients. J. Infect. Dis. 1998, 178, 297–303. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Melting Temperature | Sequences |
|---|---|---|
| Forward | 54.3 °C | 5′- CTT AAC AGC GTC AAC AGC G -3′ |
| Reverse | 54.5 °C | 5′- CAC CCG TGC GTT TTA TTC TG -3′ |
| Grade | Skin Outcomes |
|---|---|
| 0 | no lesions |
| 1, 2 | local site lesions |
| 3, 4, 5 | distant site zosteriform lesions along the dermatome |
| 6 | progression to severely compromised health |
| 7 | mortality (succumbed to infection) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhutta, M.S.; Sausen, D.G.; Reed, K.M.; Gallo, E.S.; Hair, P.S.; Lassiter, B.P.; Krishna, N.K.; Cunnion, K.M.; Borenstein, R. Peptide Inhibitor of Complement C1, RLS-0071, Reduces Zosteriform Spread of Herpes Simplex Virus Type 1 Skin Infection and Promotes Survival in Infected Mice. Viruses 2021, 13, 1422. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081422
Bhutta MS, Sausen DG, Reed KM, Gallo ES, Hair PS, Lassiter BP, Krishna NK, Cunnion KM, Borenstein R. Peptide Inhibitor of Complement C1, RLS-0071, Reduces Zosteriform Spread of Herpes Simplex Virus Type 1 Skin Infection and Promotes Survival in Infected Mice. Viruses. 2021; 13(8):1422. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081422
Chicago/Turabian StyleBhutta, Maimoona S., Daniel G. Sausen, Kirstin M. Reed, Elisa S. Gallo, Pamela S. Hair, Brittany P. Lassiter, Neel K. Krishna, Kenji M. Cunnion, and Ronen Borenstein. 2021. "Peptide Inhibitor of Complement C1, RLS-0071, Reduces Zosteriform Spread of Herpes Simplex Virus Type 1 Skin Infection and Promotes Survival in Infected Mice" Viruses 13, no. 8: 1422. https://0-doi-org.brum.beds.ac.uk/10.3390/v13081422