
VP8, the Major Tegument Protein of Bovine Herpesvirus-1, Is Partially Packaged during Early Tegument Formation in a VP22-Dependent Manner

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Viruses
2.2. Plasmids and Antibodies
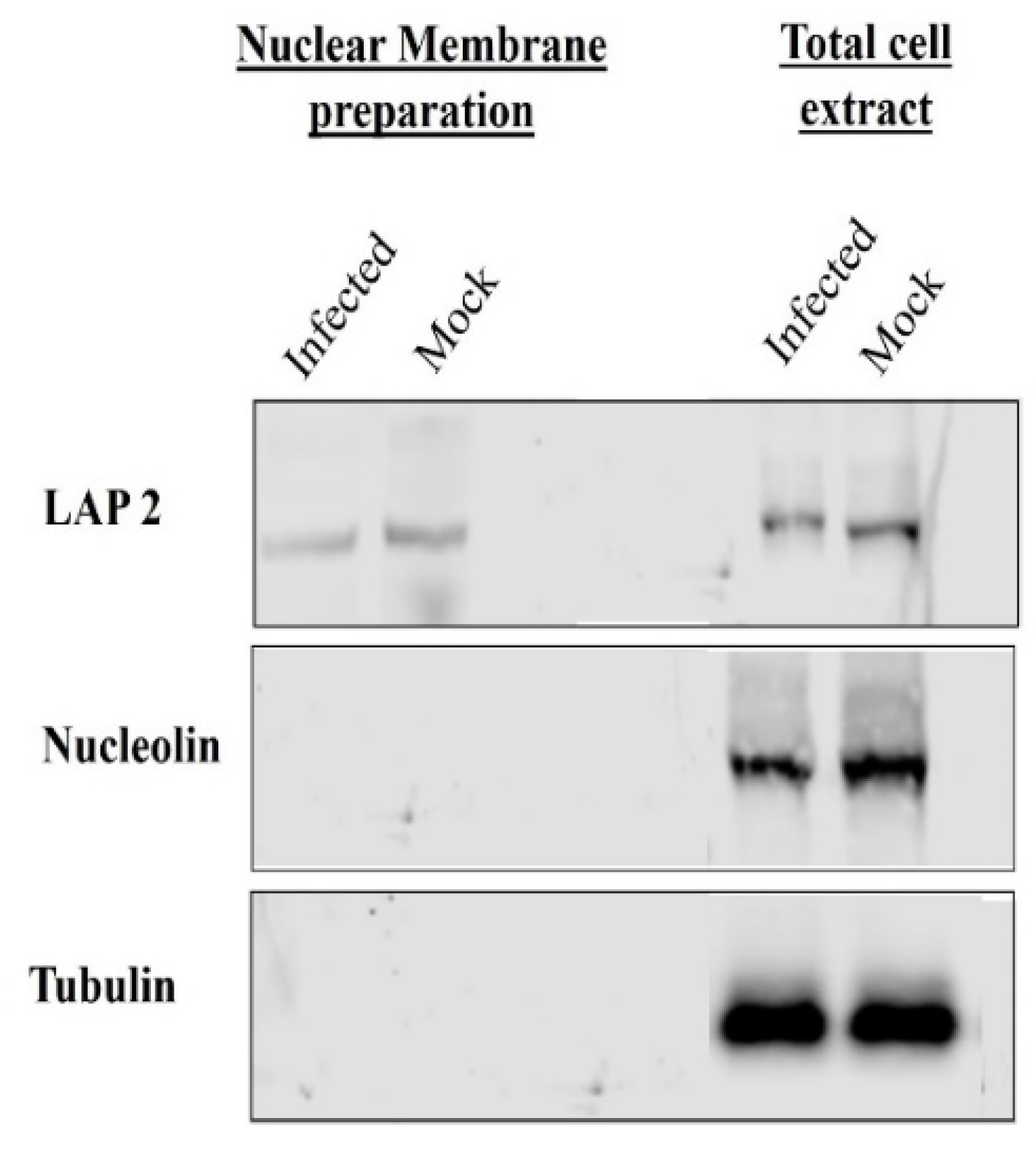
2.3. Isolation of Nuclear Membrane
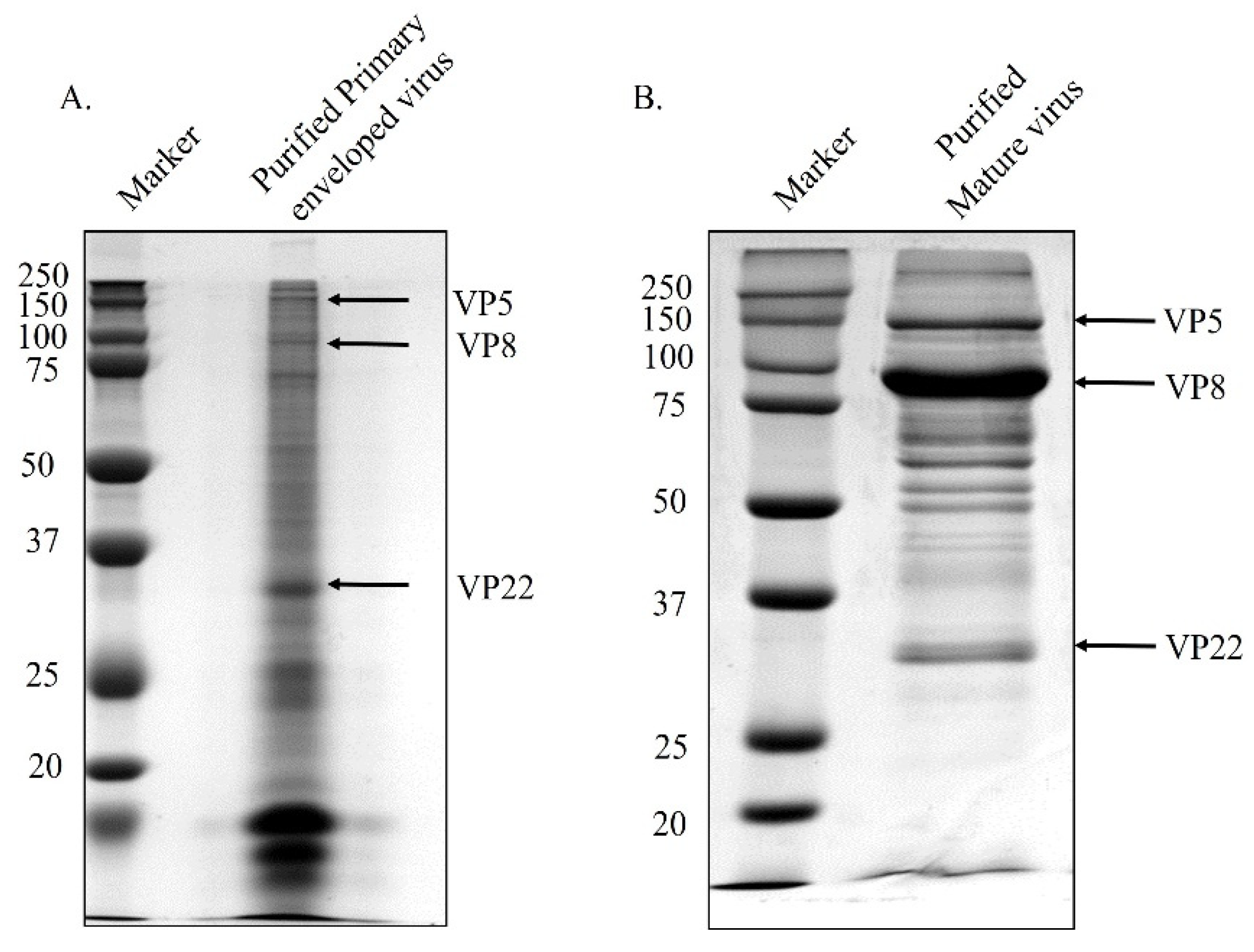
2.4. Isolation of Primary Enveloped Virions
2.5. Purification of Mature Virus
2.6. Gel Electrophoresis
2.7. Mass Spectrometry Sample Preparation and Analysis
2.8. Transfections
2.9. Preparation of Cell Lysates
2.10. Immunoprecipitation and Western Blotting
2.11. Confocal Microscopy
3. Results
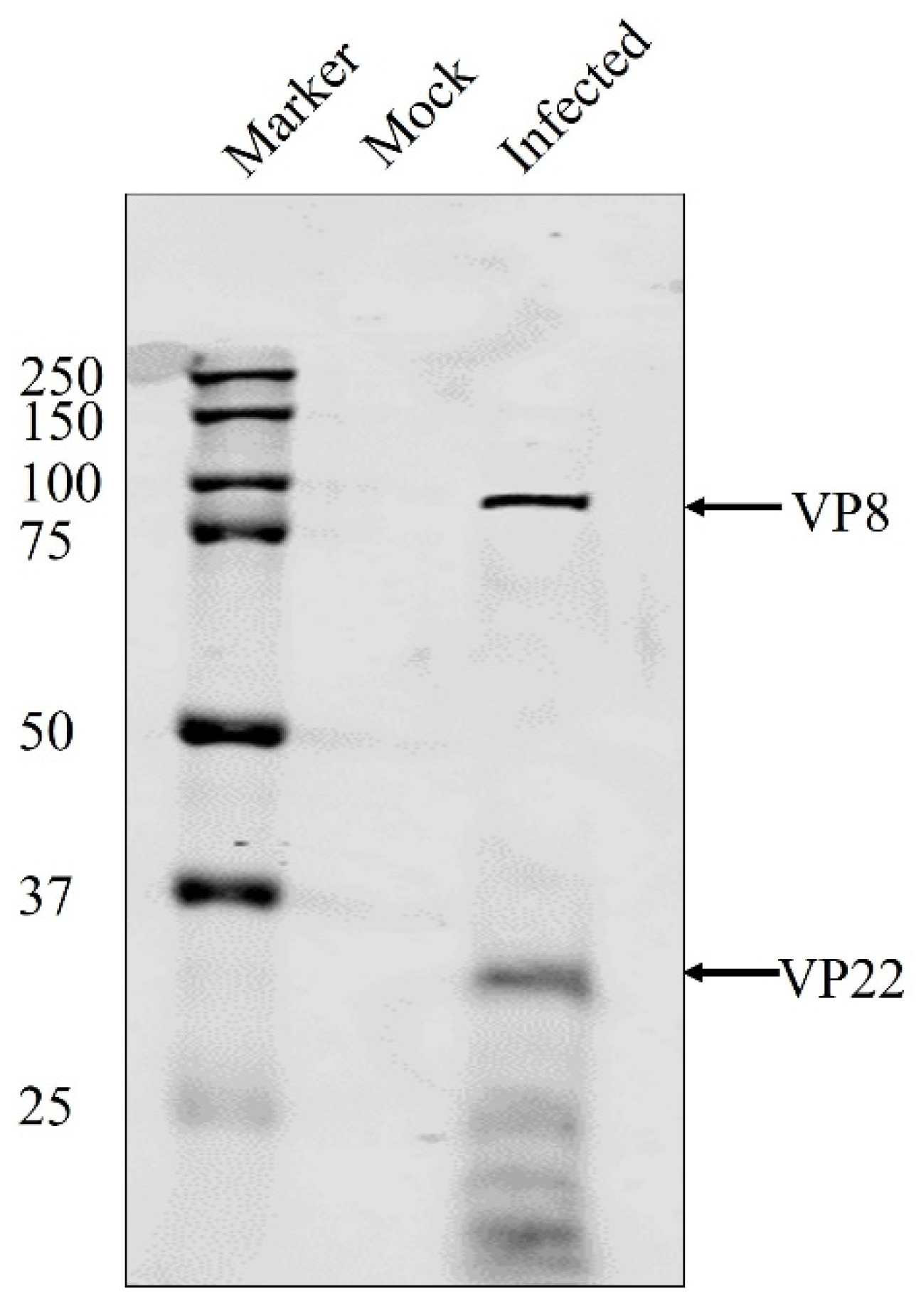
3.1. VP8 and VP22 Are Packaged at the Early Tegumentation Stage
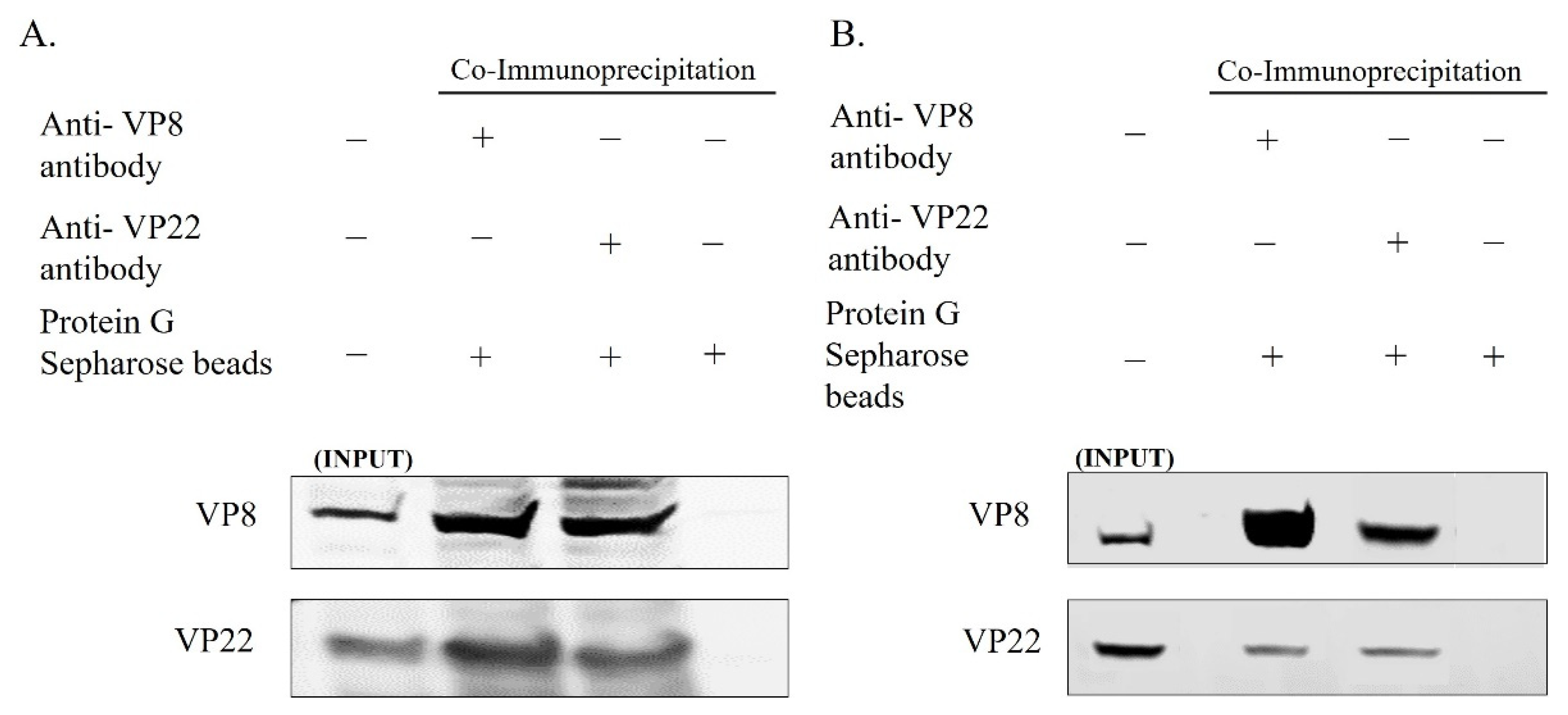
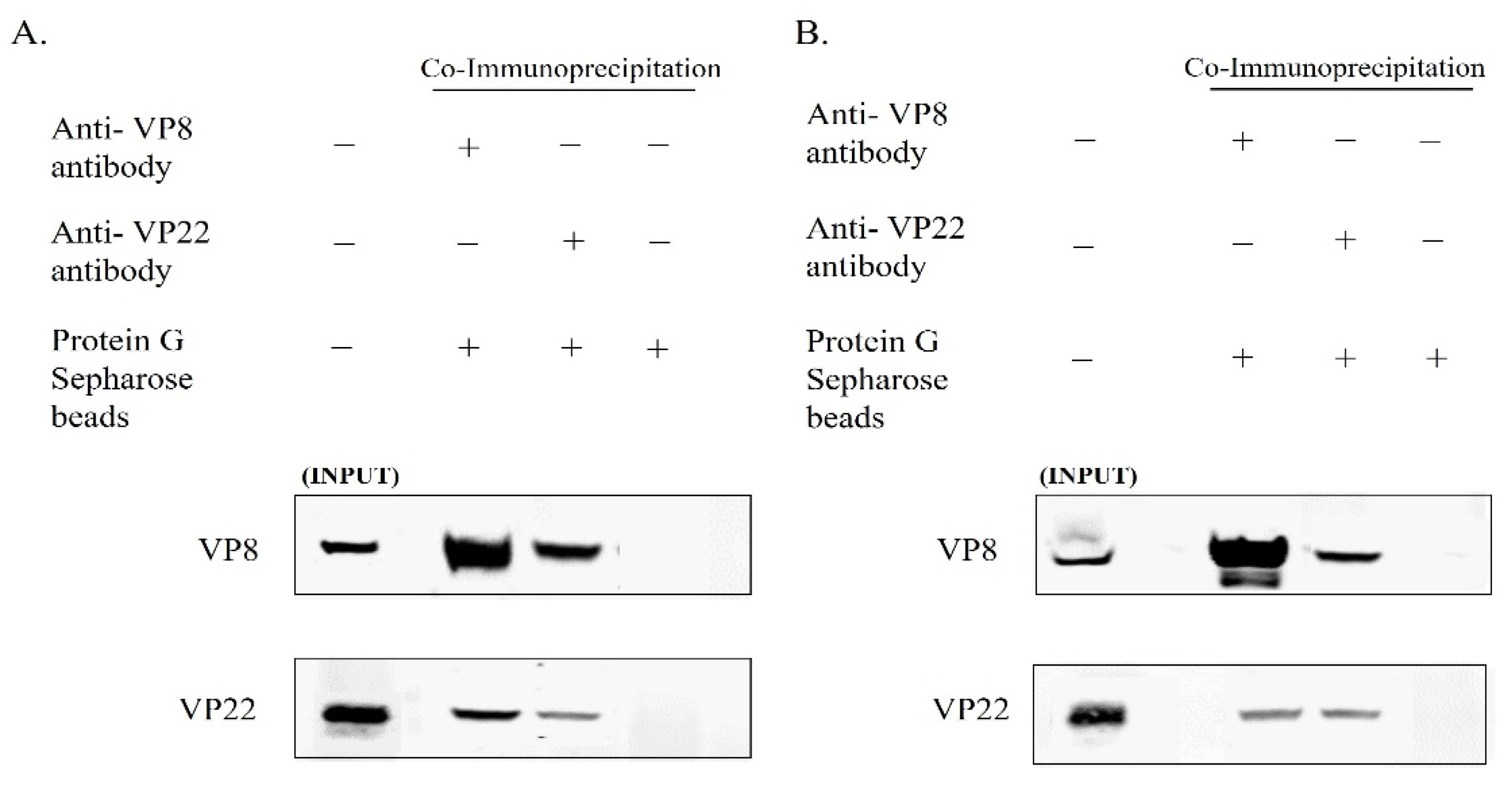
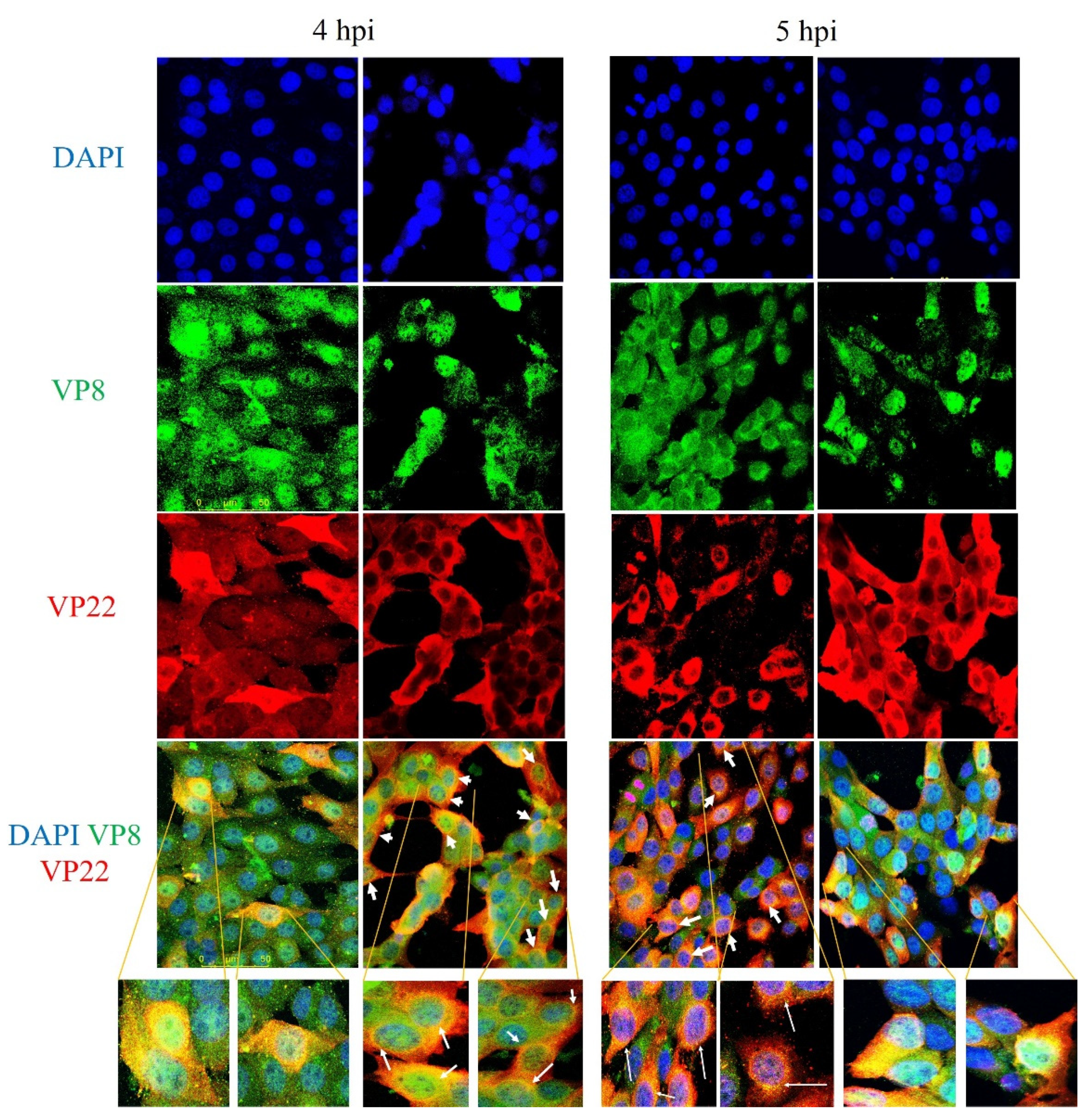
3.2. VP8 Interacts with VP22 in the Perinuclear Region and in Mature Virus in BoHV-1-Infected Cells and in BoHV1-YmVP8-Infected Cells
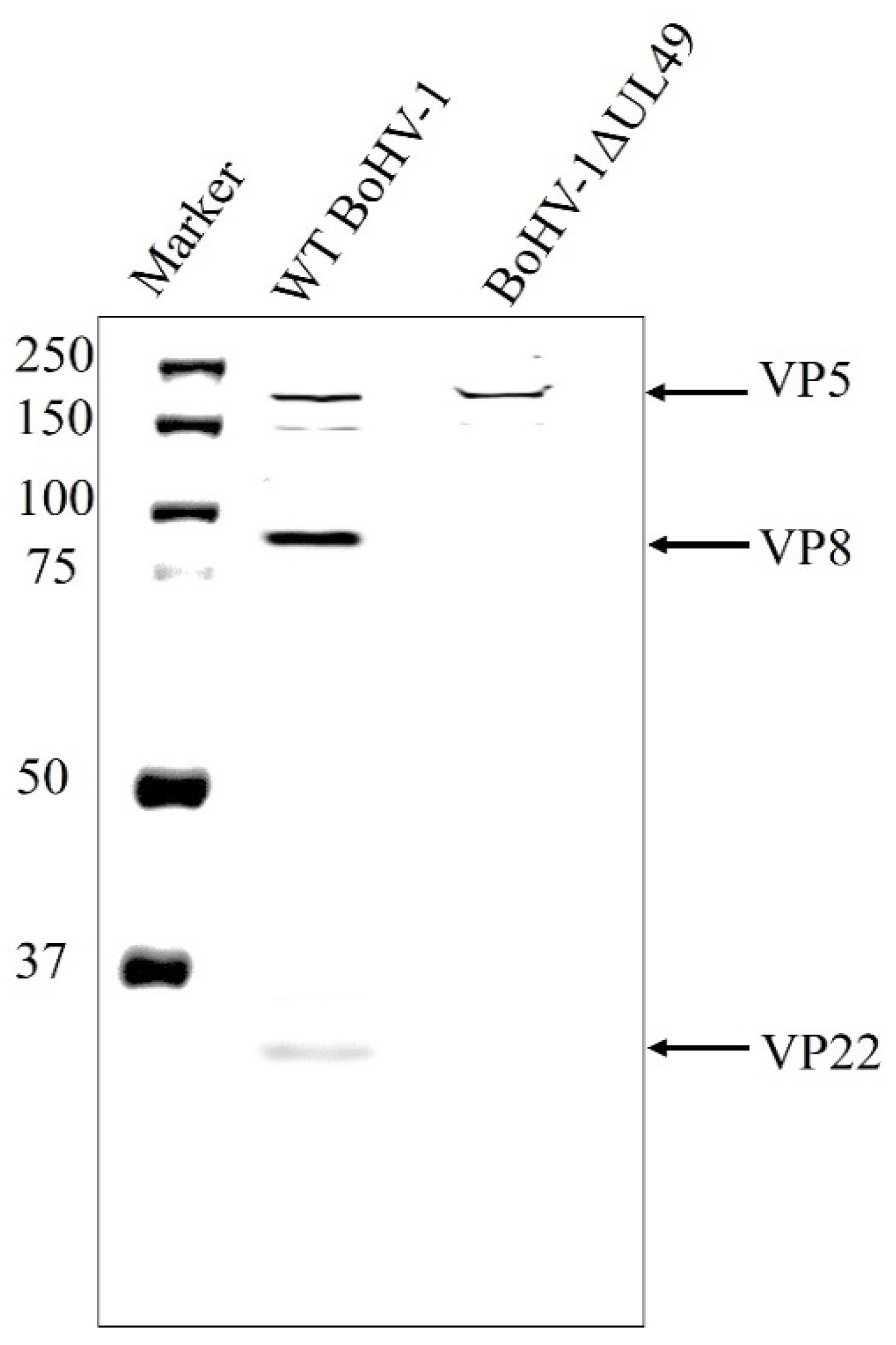
3.3. VP8 Is Absent in the Primary Enveloped Virus in the Absence of VP22
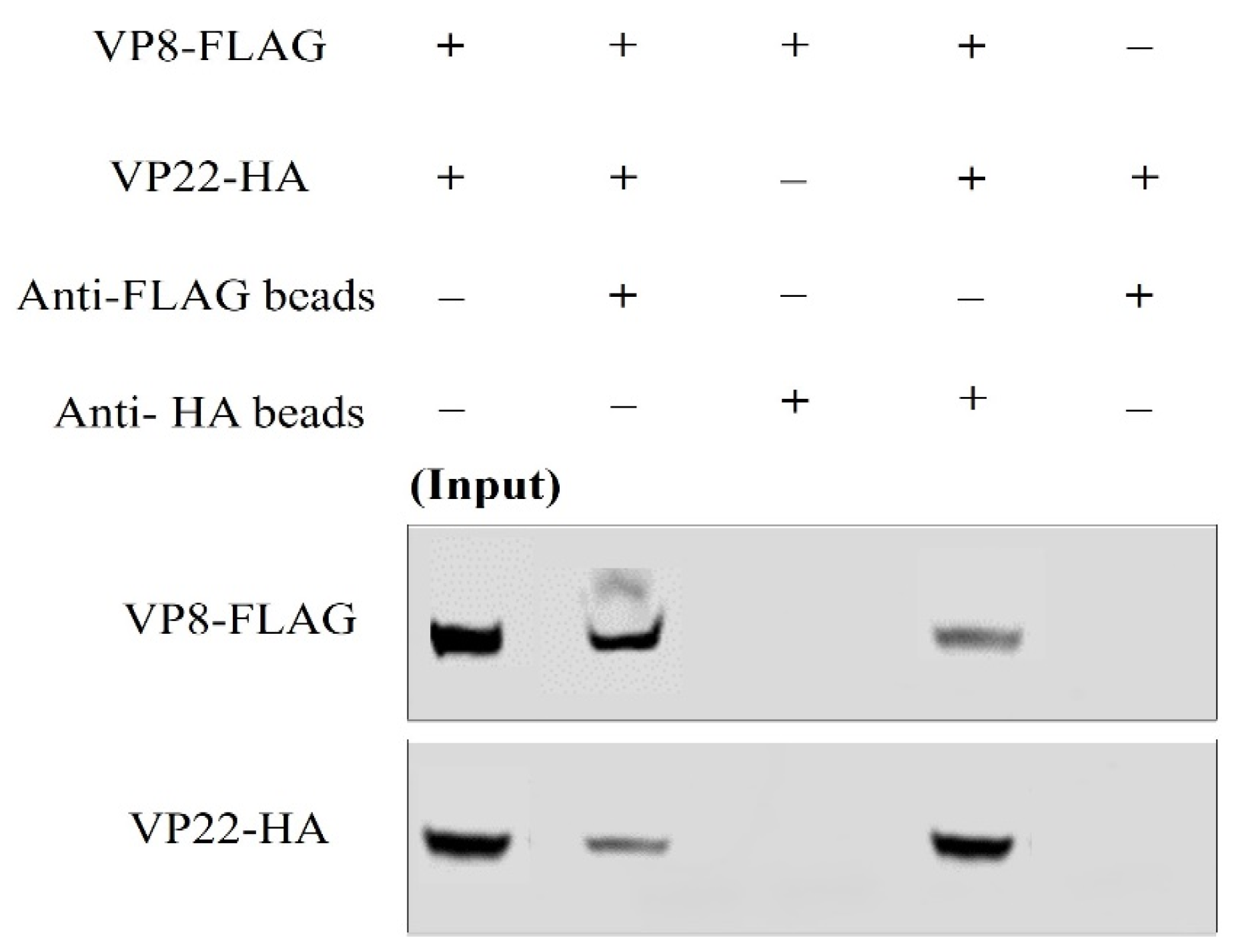
3.4. VP8 and VP22 Interact with one Another Outside the Context of Infection
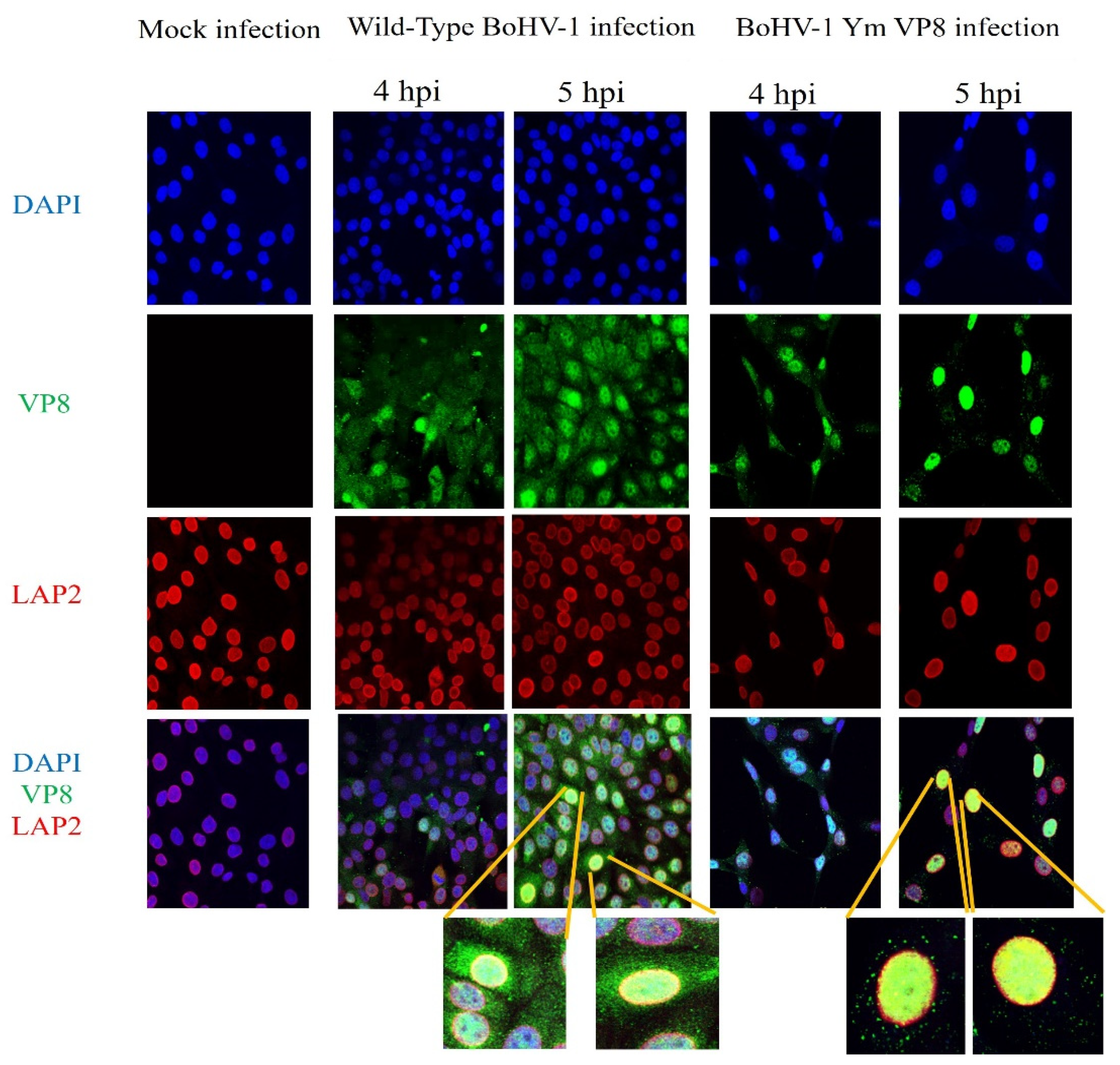
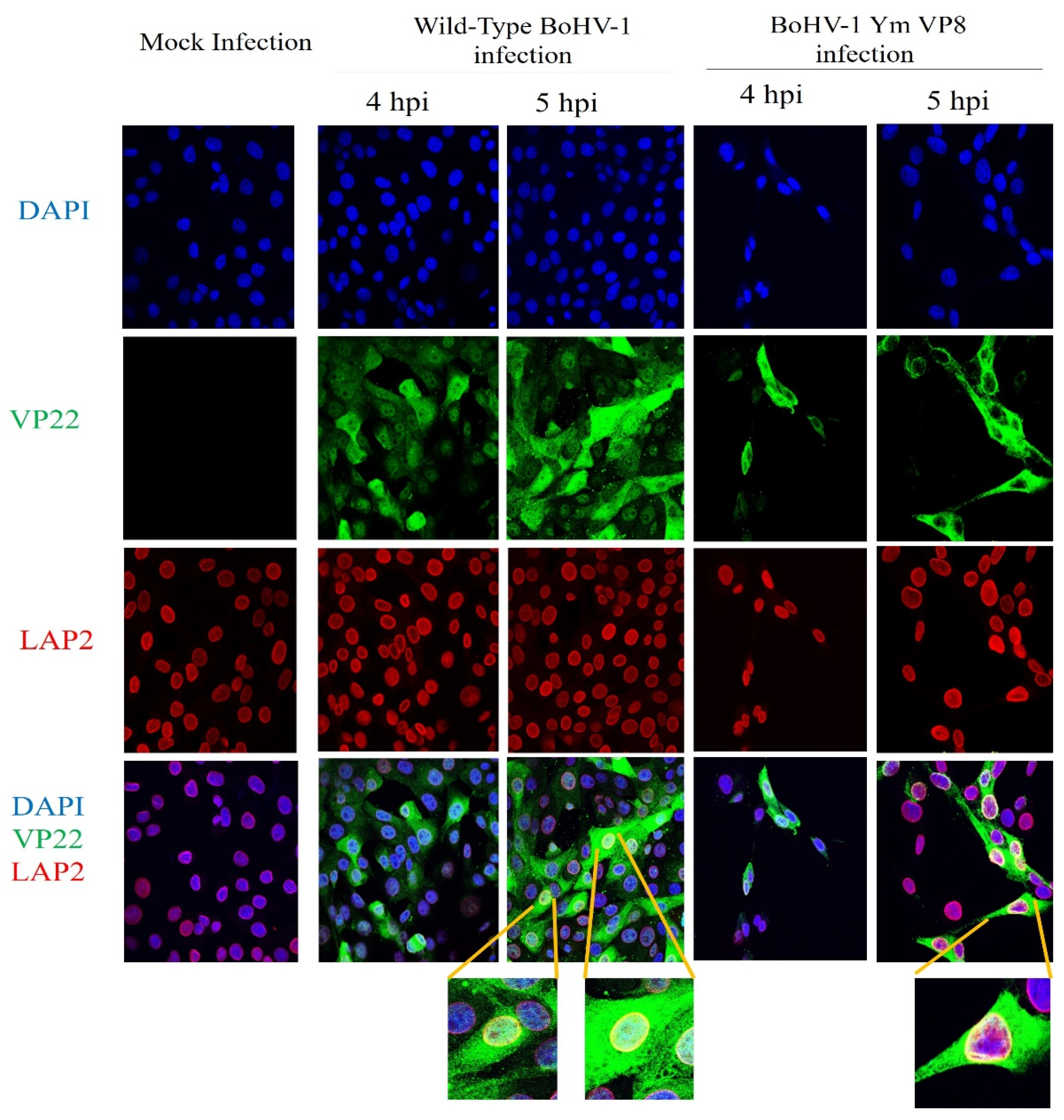
3.5. VP8 and VP22 Localise in the Perinuclear Region in Cells Infected with Wild-Type BoHV-1 or BoHV-1YmVP8
3.6. VP22 Co-Localises with VP8
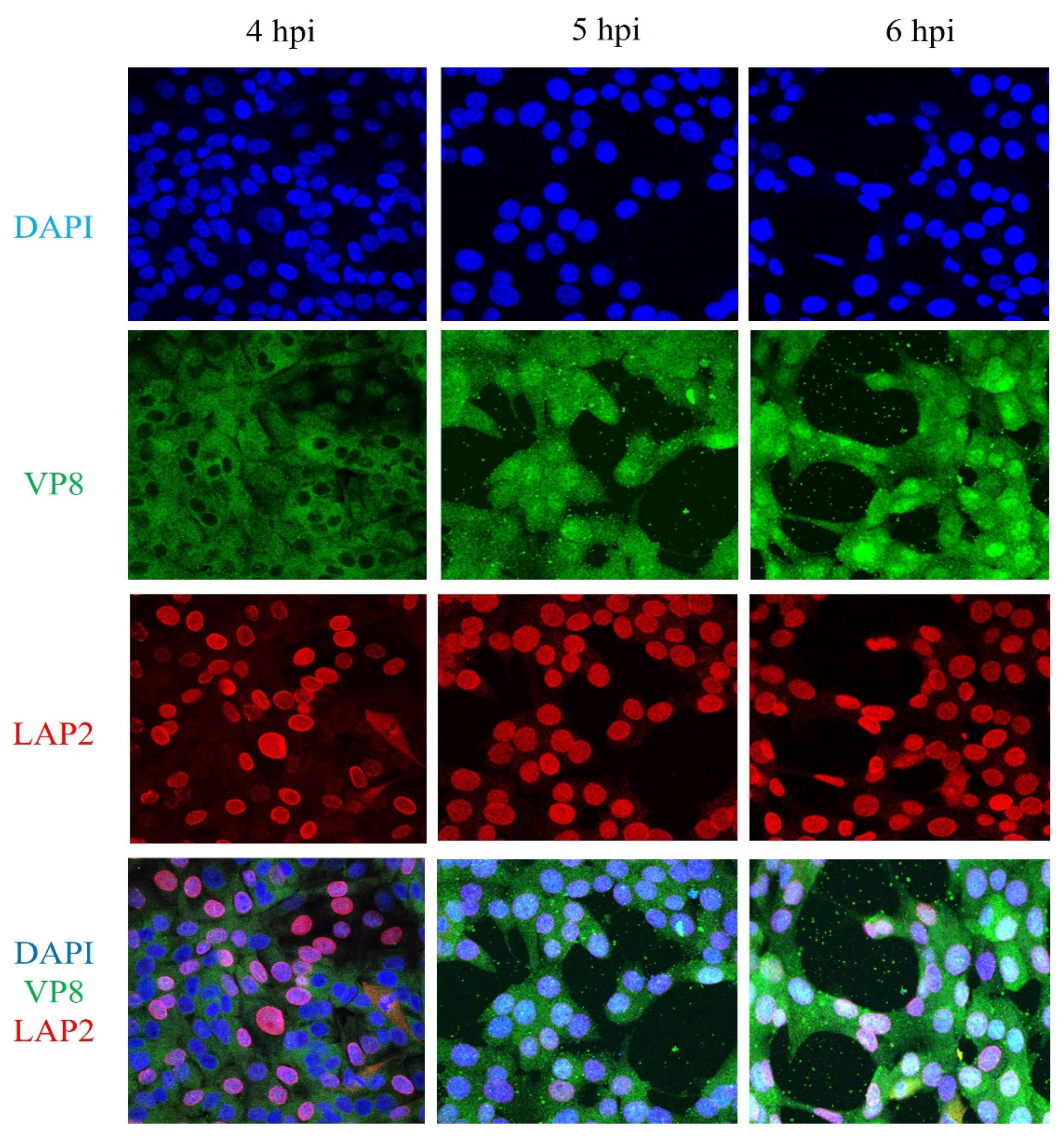
3.7. VP8 Is not Localised to the Perinuclear Region in BoHV-1ΔUL49-Infected Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, C. Bovine Herpesvirus 1 Counteracts Immune Responses and Immune-Surveillance to Enhance Pathogenesis and Virus Transmission. Front. Immunol. 2019, 10, 1008. [Google Scholar] [CrossRef] [Green Version]
- Lucchese, L.; Benkirane, A.; Hakimi, I.; El Idrissi, A.; Natale, A. Seroprevalence study of the main causes of abortion in dairy cattle in Morocco. Vet. Ital. 2016, 52, 13–19. [Google Scholar]
- Haanes, E.J.; Thomsen, D.R.; Martin, S.; Homa, F.L.; Lowery, D.E. The bovine herpesvirus 1 maturational proteinase and scaffold proteins can substitute for the homologous herpes simplex virus type 1 proteins in the formation of hybrid type B capsids. J. Virol. 1995, 69, 7375–7379. [Google Scholar] [CrossRef] [Green Version]
- Lobanov, V.A.; Maher-Sturgess, S.L.; Snider, M.G.; Lawman, Z.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S.A. UL47 gene deletion mutant of bovine herpesvirus type 1 exhibits impaired growth in cell culture and lack of virulence in cattle. J. Virol. 2010, 84, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Owen, D.J.; Crump, C.M.; Graham, S.C. Tegument Assembly and Secondary Envelopment of Alphaherpesviruses. Viruses 2015, 7, 5084–5114. [Google Scholar] [CrossRef] [Green Version]
- Metrick, C.M.; Koenigsberg, A.L.; Heldwein, E.E. Conserved Outer Tegument Component UL11 from Herpes Simplex Virus 1 Is an Intrinsically Disordered, RNA-Binding Protein. mBio 2020, 11, e00810-20. [Google Scholar] [CrossRef]
- Mettenleiter, T.C.; Klupp, B.G.; Granzow, H. Herpesvirus assembly: A tale of two membranes. Curr. Opin. Microbiol. 2006, 9, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Muylkens, B.; Thiry, J.; Kirten, P.; Schynts, F.; Thiry, E. Bovine herpesvirus 1 infection and infectious bovine rhinotracheitis. Vet. Res. 2007, 38, 181–209. [Google Scholar] [CrossRef] [Green Version]
- Homa, F.L.; Brown, J.C. Capsid assembly and DNA packaging in herpes simplex virus. Rev. Med. Virol. 1997, 7, 107–122. [Google Scholar] [CrossRef]
- Crump, C. Virus Assembly and Egress of HSV. Adv. Exp. Med. Biol. 2018, 1045, 23–44. [Google Scholar] [PubMed]
- Mettenleiter, T.C.; Klupp, B.G.; Granzow, H. Herpesvirus assembly: An update. Virus Res. 2009, 143, 222–234. [Google Scholar] [CrossRef]
- Granzow, H.; Klupp, B.G.; Fuchs, W.; Veits, J.; Osterrieder, N.; Mettenleiter, T.C. Egress of alphaherpesviruses: Comparative ultrastructural study. J. Virol. 2001, 75, 3675–3684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mettenleiter, T.C. Budding events in herpesvirus morphogenesis. Virus Res. 2004, 106, 167–180. [Google Scholar] [CrossRef]
- Carpenter, D.E.; Misra, V. The Most Abundant Protein in Bovine Herpes 1 Virions is a Homologue of Herpes Simplex Virus Type 1 UL47. J. Gen. Virol. 1991, 72 Pt 12, 3077–3084. [Google Scholar] [CrossRef]
- van Drunen Littel-van den Hurk, S.; Garzon, S.; van den Hurk, J.V.; Babiuk, L.A.; Tijssen, P. The role of the major tegument protein VP8 of bovine herpesvirus-1 in infection and immunity. Virology 1995, 206, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Afroz, S.; Brownlie, R.; Snider, M.; van Drunen Littel-van den Hurk, S. Regulation and function of phosphorylation on VP8, the major tegument protein of bovine herpesvirus 1. J. Virol. 2015, 89, 4598–4611. [Google Scholar] [CrossRef] [Green Version]
- Afroz, S.; Garg, R.; Fodje, M.; van Drunen Littel-van den Hurk, S. The Major Tegument Protein of Bovine Herpesvirus 1, VP8, Interacts with DNA Damage Response Proteins and Induces Apoptosis. J. Virol. 2018, 92, e00773-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhagen, J.; Hutchinson, I.; Elliott, G. Nucleocytoplasmic Shuttling of Bovine Herpesvirus 1 UL47 Protein in Infected Cells. J. Virol. 2006, 80, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, M.; Elliott, G. Nuclear localization and shuttling of herpes simplex virus tegument protein VP13/14. J. Virol. 2001, 75, 2566–2574. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Brownlie, R.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Characterization of nuclear localization and export signals of the major tegument protein VP8 of bovine herpesvirus-1. Virology 2004, 324, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Donovan, T.; Sucharita, S.; Brownlie, R.; Snider, M.; Tikoo, S.K.; van Drunen Littel-van den Hurk, S. US3 Kinase-Mediated Phosphorylation of Tegument Protein VP8 Plays a Critical Role in the Cellular Localization of VP8 and Its Effect on the Lipid Metabolism of Bovine Herpesvirus 1-Infected Cells. J. Virol. 2019, 93, e02151-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Brownlie, R.; Snider, M.; Hurk, S.V.D.L.-V.D. Phosphorylation of Bovine Herpesvirus 1 VP8 Plays a Role in Viral DNA Encapsidation and Is Essential for Its Cytoplasmic Localization and Optimal Virion Incorporation. J. Virol. 2016, 90, 4427–4440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Chow, B.; Li, Y.; Raggo, C.; Yoo, D.; Attah-Poku, S.; Babiuk, L.A. Characterization of bovine herpesvirus 1 UL49 homolog gene and product: Bovine herpesvirus 1 UL49 homolog is dispensable for virus growth. J. Virol. 1995, 69, 3863–3867. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Harms, J.S.; Splitter, G.A. Bovine Herpesvirus 1 Tegument Protein VP22 Interacts with Histones, and the Carboxyl Terminus of VP22 Is Required for Nuclear Localization. J. Virol. 2001, 75, 8251–8258. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, E.F.M.; Blaho, J.A. A conserved carboxy-terminal domain in the major tegument structural protein VP22 facilitates virion packaging of a chimeric protein during productive herpes simplex virus 1 infection. Virology 2009, 387, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Harms, J.S.; Ren, X.; Oliveira, S.C.; Splitter, G.A. Distinctions between Bovine Herpesvirus 1 and Herpes Simplex Virus Type 1 VP22 Tegument Protein Subcellular Associations. J. Virol. 2000, 74, 3301–3312. [Google Scholar] [CrossRef] [Green Version]
- Padula, M.E.; Sydnor, M.L.; Wilson, D.W. Isolation and Preliminary Characterization of Herpes Simplex Virus 1 Primary Enveloped Virions from the Perinuclear Space. J. Virol. 2009, 83, 4757–4765. [Google Scholar] [CrossRef] [Green Version]
- Lobanov, V.A.; Zheng, C.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Intracellular trafficking of VP22 in bovine herpesvirus-1 infected cells. Virology 2010, 396, 189–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Brattain, M.G. The maximal size of protein to diffuse through the nuclear pore is larger than 60 kDa. FEBS Lett. 2007, 581, 3164–3170. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Cheng, A.; Wang, M.; Jia, R.; Yang, Q.; Wu, Y.; Zhu, D.; Zhao, X.; Chen, S.; Liu, M.; et al. Alphaherpesvirus Major Tegument Protein VP22: Its Precise Function in the Viral Life Cycle. Front. Microbiol. 2020, 11, 1908. [Google Scholar] [CrossRef]
- Labiuk, S.L.; Babiuk, L.A.; Hurk, S.V.D.L.-V.D. Major tegument protein VP8 of bovine herpesvirus 1 is phosphorylated by viral US3 and cellular CK2 protein kinases. J. Gen. Virol. 2009, 90, 2829–2839. [Google Scholar] [CrossRef] [PubMed]
- Loret, S.; Guay, G.; Lippé, R. Comprehensive Characterization of Extracellular Herpes Simplex Virus Type 1 Virions. J. Virol. 2008, 82, 8605–8618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mocarski Jr, E.S. Comparative analysis of herpesvirus-common proteins. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press Copyright © Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Barber, K.A.; Daugherty, H.C.; Ander, S.E.; Jefferson, V.A.; Shack, L.A.; Pechan, T.; Nanduri, B.; Meyer, F. Protein Composition of the Bovine Herpesvirus 1.1 Virion. Vet. Sci. 2017, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannhorst, K.; Wei, H.; Yezid, H.; He, J.; Chowdhury, S.I. Bovine Herpesvirus 1 U(L)49.5 Interacts with gM and VP22 To Ensure Virus Cell-to-Cell Spread and Virion Incorporation: Novel Role for VP22 in gM-Independent U(L)49.5 Virion Incorporation. J. Virol. 2018, 92, e00240-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuRaine, G.; Wisner, T.W.; Johnson, D.C. Characterization of the Herpes Simplex Virus (HSV) Tegument Proteins That Bind to gE/gI and US9, Which Promote Assembly of HSV and Transport into Neuronal Axons. J. Virol. 2020, 94, e01113-20. [Google Scholar] [CrossRef]
- Xue, X.; Huang, J.; Wang, H. The Study of the Intercellular Trafficking of the Fusion Proteins of Herpes Simplex Virus Protein VP22. PLoS ONE 2014, 9, e100840. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Babiuk, L.A.; Hurk, S.V.D.L.-V.D. Bovine Herpesvirus 1 VP22 Enhances the Efficacy of a DNA Vaccine in Cattle. J. Virol. 2005, 79, 1948–1953. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Xiao, S.; Jiang, Y.; Fang, L.; Chen, H.; Jin, M. A DNA vaccine encoding the FMDV capsid precursor polypeptide P1 and the enhancing effect of bovine herpesvirus 1 VP22 protein as molecular adjuvant. Acta Virol. 2012, 56, 111–117. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein | Present in Primary Enveloped Virus | Present in Mature Virus | Molecular Weight (kDa) | Number of Unique Peptides | Peptide Coverage (%) |
|---|---|---|---|---|---|---|
| Glycoproteins | ||||||
| UL27 | Glycoprotein B (gB) | x | √ | 101.1 | 200 | 63 |
| UL44 | Glycoprotein C (gC) | x | √ | 55.3 | 134 | 60 |
| US6 | Glycoprotein D (gD) | x | √ | 44.5 | 19 | 73 |
| US8 | Glycoprotein E (gE) | x | √ | 61.1 | 65 | 46 |
| UL22 | Glycoprotein H (gH) | x | √ | 88.3 | 82 | 57 |
| UL1 | Glycoprotein L (gL) | x | √ | 17.1 | 8 | 23 |
| US4 | Glycoprotein G (gG) | x | √ | 46.5 | 67 | 48 |
| UL10 | Glycoprotein M (gM) | x | √ | 45.5 | 6 | 13 |
| UL49.5 | Glycoprotein N (gN) | x | √ | 10.2 | 5 | 25 |
| UL53 | Glycoprotein K (gK) | x | √ | 35.8 | 5 | 25 |
| US7 | Glycoprotein I (gI) | x | √ | 39.9 | 100 | 45 |
| Envelope Proteins | ||||||
| UL20 | pUL20 | x | √ | 25.6 | 5 | 37 |
| UL34 | pUL34 | x | √ | 27.1 | 55 | 54 |
| UL43 | pUL43 | x | √ | 36.9 | 9 | 25 |
| Capsid Proteins | ||||||
| UL6 | Capsid portal protein | √ | √ | 75.1 | 30 | 86 |
| UL18 | Capsid triplex subunit | √ | √ | 33.3 | 37 | 46 |
| UL19 | VP5 major capsid protein | √ | √ | 105.1 | 106 | 52 |
| Tegument Proteins | ||||||
| UL2 | pUL2 | √ | x | 33.8 | 8 | 18 |
| UL7 | pUL7 | x | √ | 32.5 | 21 | 63 |
| UL11 | pUL11 | x | √ | 9.7 | 15 | 96 |
| UL14 | pUL14 | x | √ | 23.2 | 31 | 76 |
| UL16 | pUL16 | x | √ | 36.4 | 12 | 25 |
| UL17 | Cleavage protein | x | √ | 72.6 | 59 | 83 |
| UL21 | pUL21 | × | √ | 60.2 | 47 | 48 |
| UL25 | Cleavage protein | × | √ | 63.1 | 62 | 63 |
| UL26 | Maturational protease | √ | √ | 63.7 | 14 | 46 |
| UL29 | Major DNA binding protein | √ | √ | 78.9 | 13 | 29 |
| UL31 | pUL31 | x | √ | 39.5 | 35 | 72 |
| UL32 | pUL32 | x | √ | 62.9 | 4 | 23 |
| UL36 | Egress regulating protein | √ | x | 327.2 | 79 | 57 |
| UL37 | Egress regulating protein | √ | x | 105.9 | 86 | 67 |
| UL39 | pUL39 | √ | √ | 86.1 | 27 | 58 |
| UL40 | pUL40 | √ | √ | 31.1 | 15 | 57 |
| UL41 | Host shut off protein | x | √ | 50.1 | 41 | 65 |
| UL42 | DNA processivity protein | x | √ | 42.6 | 67 | 63 |
| UL46 | VP11/12 | x | √ | 77.6 | 26 | 38 |
| UL47 | VP8 | √ | √ | 80.7 | 47 | 65 |
| UL48 | VP16 | x | √ | 54.1 | 55 | 58 |
| UL49 | VP22 | √ | √ | 26.8 | 148 | 84 |
| UL50 | pUL50 | x | √ | 34.1 | 18 | 63 |
| UL51 | pUL51 | x | √ | 24.9 | 16 | 86 |
| UL54 | bICP27 | x | √ | 43.3 | 26 | 63 |
| US1.67 | pUS1.67 | x | √ | 27.2 | 28 | 44 |
| US3 | Serine/threonine kinase | x | √ | 49.9 | 27 | 48 |
| Host Proteins | ||||||
| HSP70 | x | √ | 70.8 | 4 | 18 | |
| Eukaryotic initiation factor 4H | x | √ | 30.8 | 26 | 18 | |
| Annexin | x | √ | 38.9 | 15 | 75 | |
| Alpha tubulin | x | √ | 50 | 5 | 23 | |
| Beta actin | x | √ | 40.3 | 9 | 13 | |
| Histone | √ | √ | 13.9 | 5 | 29 | |
| PCNA | √ | √ | 28.8 | 11 | 26 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sucharita, S.; Zhang, K.; van Drunen Littel-van den Hurk, S. VP8, the Major Tegument Protein of Bovine Herpesvirus-1, Is Partially Packaged during Early Tegument Formation in a VP22-Dependent Manner. Viruses 2021, 13, 1854. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091854
Sucharita S, Zhang K, van Drunen Littel-van den Hurk S. VP8, the Major Tegument Protein of Bovine Herpesvirus-1, Is Partially Packaged during Early Tegument Formation in a VP22-Dependent Manner. Viruses. 2021; 13(9):1854. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091854
Chicago/Turabian StyleSucharita, Soumya, Kuan Zhang, and Sylvia van Drunen Littel-van den Hurk. 2021. "VP8, the Major Tegument Protein of Bovine Herpesvirus-1, Is Partially Packaged during Early Tegument Formation in a VP22-Dependent Manner" Viruses 13, no. 9: 1854. https://0-doi-org.brum.beds.ac.uk/10.3390/v13091854