
Outbreak of a Systemic Form of Camelpox in a Dromedary Herd (Camelus dromedarius) in the United Arab Emirates

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Evaluation of Diseased Animals
2.2. Histopathology and Polymerase Chain Reaction (PCR)
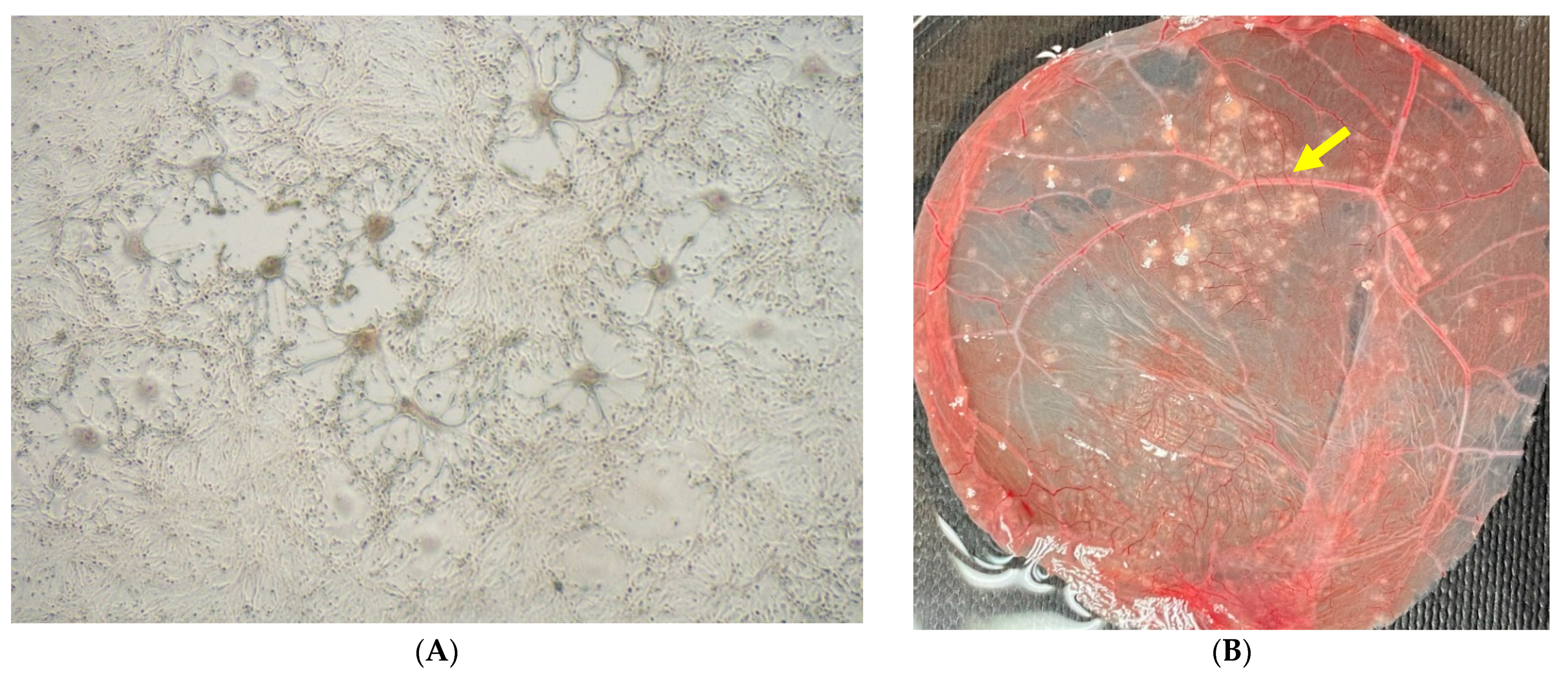
2.3. Virus Isolation
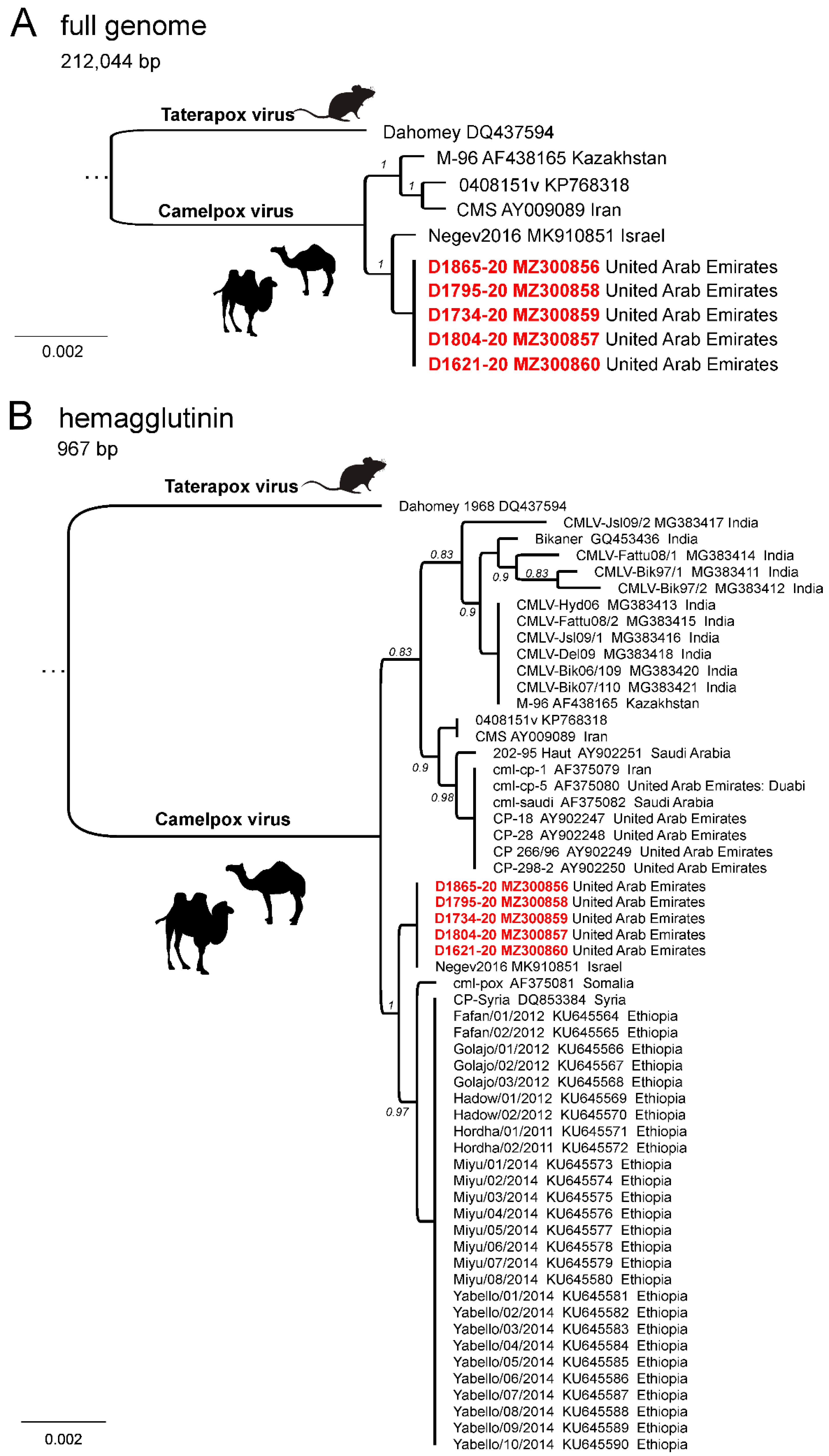
2.4. Sequencing and Phylogenetic Analysis
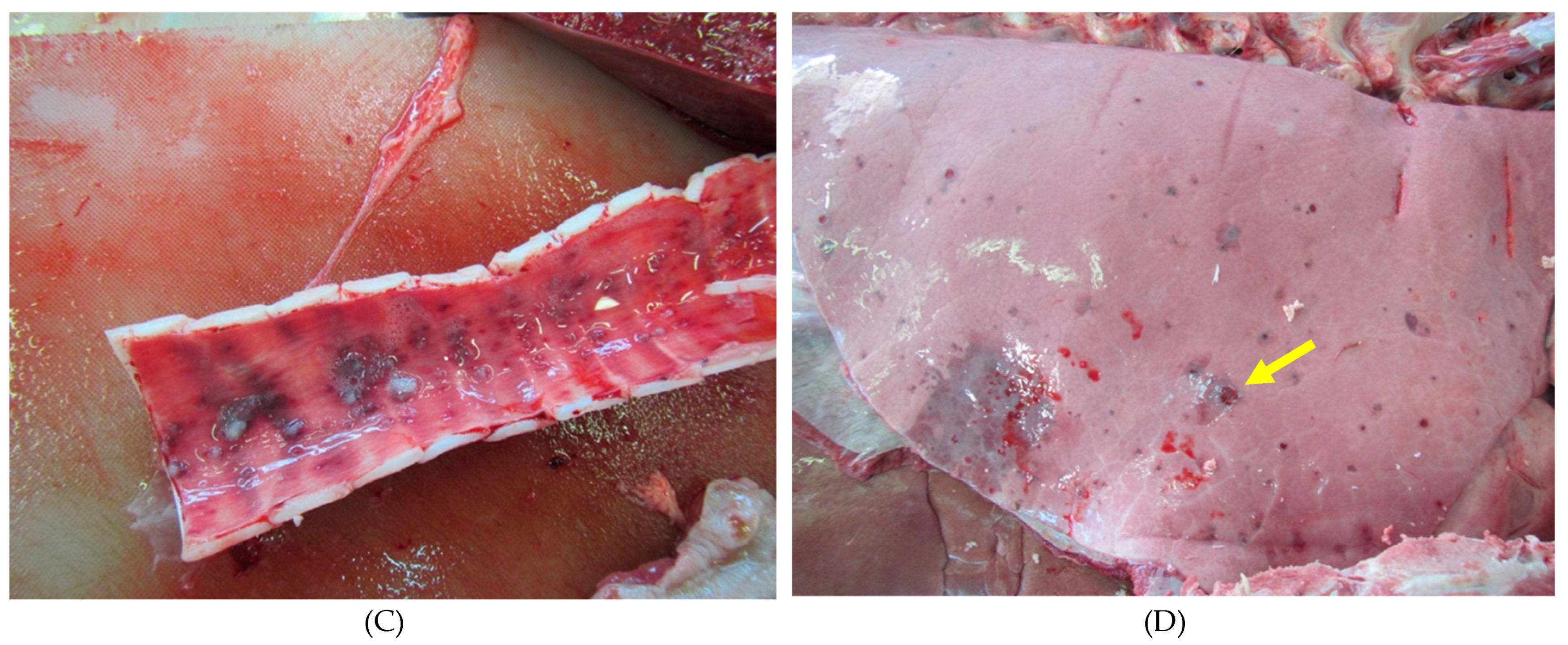
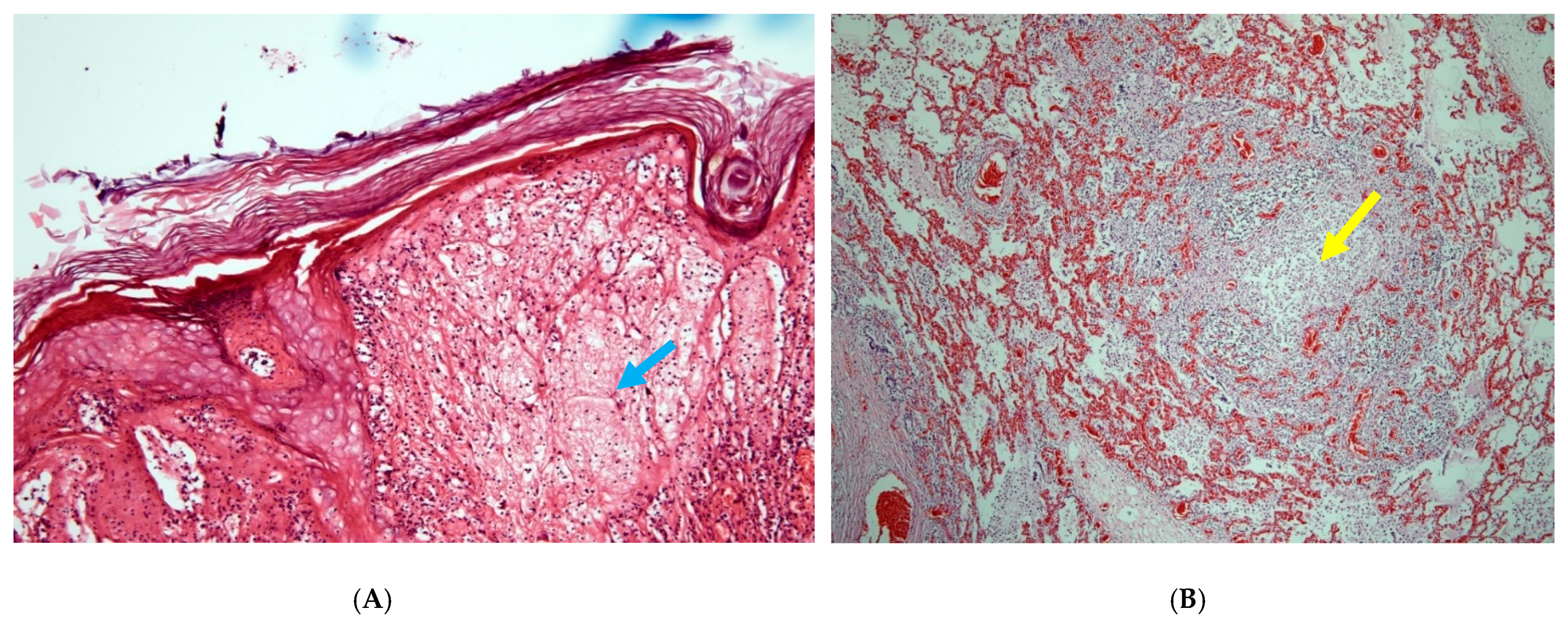
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wernery, U.; Kinne , J.; Zachariah, R. Experimental camelpox infection in vaccinated and unvaccinated guanacos. J. Camel Pract. Res. 2000, 7, 153–157. [Google Scholar]
- Gubser, C.; Smith, G.L. The sequence of camelpox virus shows it is most closely related to variola virus, the cause of smallpox. J. Gen. Virol. 2002, 83, 855–872. [Google Scholar] [CrossRef] [PubMed]
- Bera, B.; Shanmugasundaram, K.; Barua, S.; Venkatesan, G.; Virmani, N.; Riyesh, T.; Gulati, B.; Bhanuprakash, V.; Vaid, R.; Kakker, N.; et al. Zoonotic cases of camelpox infection in India. Veter- Microbiol. 2011, 152, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Khalafalla, A.I.; Abdelazim, F. Human and Dromedary Camel Infection with Camelpox Virus in Eastern Sudan. Vector-Borne Zoonotic Dis. 2017, 17, 281–284. [Google Scholar] [CrossRef] [PubMed]
- OIE. Chapter 3.5.1 camelpox. In Manual of Diagnostic Tests and Vaccines in Terrestrial Animals. 2021. Available online: https://www.oie.int/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 1 May 2021).
- Wernery, U.K.J.; Schuster, R.K. Camelid Infectious Disorders, 3rd ed.; OIE: Paris, France, 2014. [Google Scholar]
- Wernery, U.; Kaaden, O.R.; Ali, M. Orthopox virus infections in dromedary camels in United Arab Emirates (U.A.E.) during winter season. J. Camel Pract. Res. 1997, 4, 51–55. [Google Scholar]
- Khalafalla, A.I. Emerging infectious diseases in camelids. In Emerging and Re-Emerging Infectious Diseases of Livestock; Bayry, J., Ed.; Springer International Publishing: Basel, Switzerland, 2017; pp. 425–441. [Google Scholar]
- Cardeti, G.; Brozzi, A.; Eleni, C.; Polici, N.; D’Alterio, G.; Carletti, F.; Scicluna, M.T.; Castilletti, C.; Capobianchi, M.R.; Di Caro, A.; et al. Cowpox virus in llama, Italy. Emerg Infect Dis 2011, 17, 1513–1515. [Google Scholar] [CrossRef] [PubMed]
- Prkno, A.; Hoffmann, D.; Goerigk, D.; Kaiser, M.; Van Maanen, A.C.F.; Jeske, K.; Jenckel, M.; Pfaff, F.; Vahlenkamp, T.W.; Beer, M.; et al. Epidemiological Investigations of Four Cowpox Virus Outbreaks in Alpaca Herds, Germany. Viruses 2017, 9, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinne, J.; Cooper, J.E.; Wernery, U. Pathological studies on camelpox lesions of the respiratory system in the Anited Arab Emirates (UAE). J. Comp. Pathol. 1998, 118, 257–266. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and practice of histological techniques. In The Hematoxylins and Eosin; Churchill Livingstone: London, UK; New York, NY, USA, 2002; Chapter 8. [Google Scholar]
- Pfeffer, M.; Wernery , U.; Kaaden, O.R.; Meyer, H. Diagnostic procedures for poxvirus infections in camelids. J. Camel Pract. Res. 1998, 5, 189–195. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.J. 2011, 17, 3. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Reuter, K.; Drost, H.-G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. Mafft: A novel method for rapid multiple sequence alignment based on fast fourier transform. Nucleic Acids Res 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A. FastTree 2–Approximately Maximum-Likelihood Trees for Large Alignments. PLOS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Israeli, O.; Cohen-Gihon, I.; Zvi, A.; Shifman, O.; Melamed, S.; Paran, N.; Laskar-Levy, O.; Beth-Din, A. Complete Genome Sequence of the First Camelpox Virus Case Diagnosed in Israel. Microbiol. Resour. Announc. 2019, 8, e00671-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kříž, B. A study of camelpox in Somalia. J. Comp. Pathol. 1982, 92, 1–8. [Google Scholar] [CrossRef]
- Wernery, U.; Kaaden, O.R. Chapter: Camelpox. In Infectious Diseases of Camelids; Blackwell Science Ltd.: Oxford, UK, 2002. [Google Scholar]
- Duraffour, S.; Meyer, H.; Andrei, G.; Snoeck, R. Camelpox virus. Antivir. Res. 2011, 92, 167–186. [Google Scholar] [CrossRef]
- Narnaware, S.D.; Ranjan, R.; Dahiya, S.S.; Panchbuddhe, A.; Bajpai, D.; Tuteja, F.C.; Sawal, R.K. Pathological and molecular investigations of systemic form of camelpox in naturally infected adult male dromedary camels in India. Heliyon 2021, 7, e06186. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Dromedary | Number of Animals | Number of Diseased Animals | Morbidity Rate (%) | Number of Dead Animals | Mortality Rate (%) | Case Fatality Rate (%) * |
|---|---|---|---|---|---|---|
| Adult female | 2689 | 24 | 0.89 | 2 | 0.07 | 8.3 |
| Adult male | 48 | 1 | 2.08 | 1 | 2.08 | 100.0 |
| Calf < 1 year | 1128 | 9 | 0.80 | 1 | 0.09 | 11.1 |
| Yearling from 1 to 2 years | 480 | 2 | 0.42 | 0 | 0.00 | 0.00 |
| Yearling > 2 years | 740 | 20 | 2.70 | 1 | 0.27 | 5.0 |
| Total | 5085 | 56 | 1.10 | 5 | 0.10 | 8.9 |
| No | Diagnostic ID and Type of Dromedary | Weight (kg) | Clinical Signs |
|---|---|---|---|
| 1 | D1621/20 Adult male | 604 | Fever, nasal discharge, general oedema but particularly on the preputium, off feed, |
| 2 | D1734/20 Yearling > 2 years | 280 | Fever, oedema around larynx/distal neck, Swollen body lymph nodes, off feed |
| 3 | D1795/20 Yearling > 2 years | 211 | Fever, nasal discharge, abdominal oedema, off feed |
| 4 | D1804/20 Yearling > 2 years | 175 | Fever, nasal discharge, many ticks, off feed |
| 5 | D2053/ 20 Calf male < 1 year | 60 | Fever, nasal discharge, poor body condition, not suckling |
| 6 | D1865/20 Adult female | 456 | Fever, general oedema, swollen body lymph nodes, off feed |
| 7 | D2132/20Adult female | 310 | Fever, nasal discharge, off feed |
| No | Diagnostic ID and Type of Dromedary | Lesions | Histology Lesions | Successful Virus Isolation From | |
|---|---|---|---|---|---|
| External | Internal | ||||
| 1 | D1621/20 Adult male | Legs, preputium, scrotum, nostril, lips | Gum, trachea, oesophagus, lung | Proliferative dermatitis, ballooning, pox-like inclusion bodies and pneumonia | Skin, body lymph nodes, trachea, oesophagus, gum and lung |
| 2 | D1734/20 Yearling female >2 years | Legs | Gum, trachea, oesophagus, lung | Proliferative dermatitis with bacterial infection | Skin, udder lymph nodes, lung, oesophagus |
| 3 | D1795/20 Yearling female >2 years | Pock nodules all over the body | Lung | Proliferative dermatitis, ballooning, bronchitis pox-like inclusion bodies, pneumonia | Skin, lung |
| 4 | D 1804/20 Yearling female >2 years | Pock nodules, especially in inguinal and perianal regions, swollen body lymph nodes, subcutaneous haemorrhages at the head | Lung | Proliferative dermatitis with pox-like inclusion bodies | Skin, body lymph nodes, trachea, tonsil, liver, spleen, brain, kidney, gum and lung |
| 5 | D2053/20 Calf male <1 year | Pock nodules all over the body | Mouth mucosa, gum, tongue, oesophagus, lung | Proliferative bronchitis, pox-like inclusion bodies | Skin, lips, nasal swabs, lung |
| 6 | D 1865/20 Adult female | Ventral abdomen/inguinal and perianal regions | Gum, trachea | Massive congestion and marked proliferation of follicular and parafollicular lymphatic tissue | Skin, gum, trachea, lung, liver |
| 7 | D2132/20 Adult female | Pock nodules all over the body | Lung, trachea | Proliferative dermatitis | Skin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joseph, S.; Kinne, J.; Nagy, P.; Juhász, J.; Barua, R.; Patteril, N.A.G.; Hoffmann, D.; Pfaff, F.; Hoffmann, B.; Wernery, U. Outbreak of a Systemic Form of Camelpox in a Dromedary Herd (Camelus dromedarius) in the United Arab Emirates. Viruses 2021, 13, 1940. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101940
Joseph S, Kinne J, Nagy P, Juhász J, Barua R, Patteril NAG, Hoffmann D, Pfaff F, Hoffmann B, Wernery U. Outbreak of a Systemic Form of Camelpox in a Dromedary Herd (Camelus dromedarius) in the United Arab Emirates. Viruses. 2021; 13(10):1940. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101940
Chicago/Turabian StyleJoseph, Sunitha, Joerg Kinne, Péter Nagy, Jutka Juhász, Rajib Barua, Nissy Annie Georgy Patteril, Donata Hoffmann, Florian Pfaff, Bernd Hoffmann, and Ulrich Wernery. 2021. "Outbreak of a Systemic Form of Camelpox in a Dromedary Herd (Camelus dromedarius) in the United Arab Emirates" Viruses 13, no. 10: 1940. https://0-doi-org.brum.beds.ac.uk/10.3390/v13101940