
Geographic Partitioning of Dengue Virus Transmission Risk in Florida

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mammalian Cell Culture and Virus Propagation
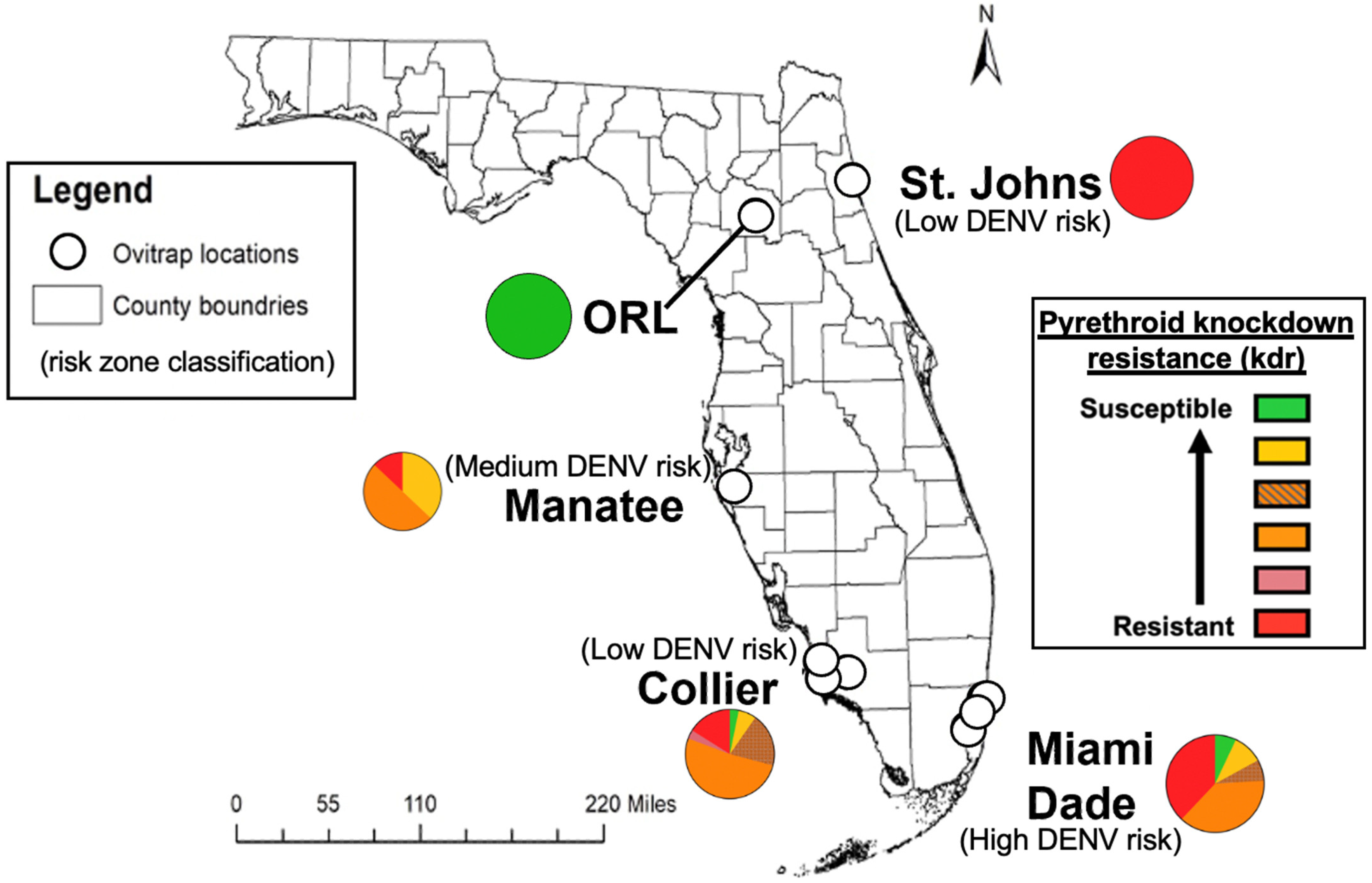
2.2. Risk Zone Classification
2.3. Field-Caught Ae. aegypti
2.4. Mosquito Rearing, Infection, and Tissue/Specimen Preparations
2.5. Mosquito Midgut and Saliva Analysis
2.6. Statistical Analyses
2.7. kdr Genotyping
3. Results
3.1. Serotype-Specific Vector Competence of the Ae. aegypti (ORL) Colony
3.2. Low Passage DENV Infections Have Elevated Midgut and Saliva Infection Intensities in Ae. aegypti ORL Mosquitoes
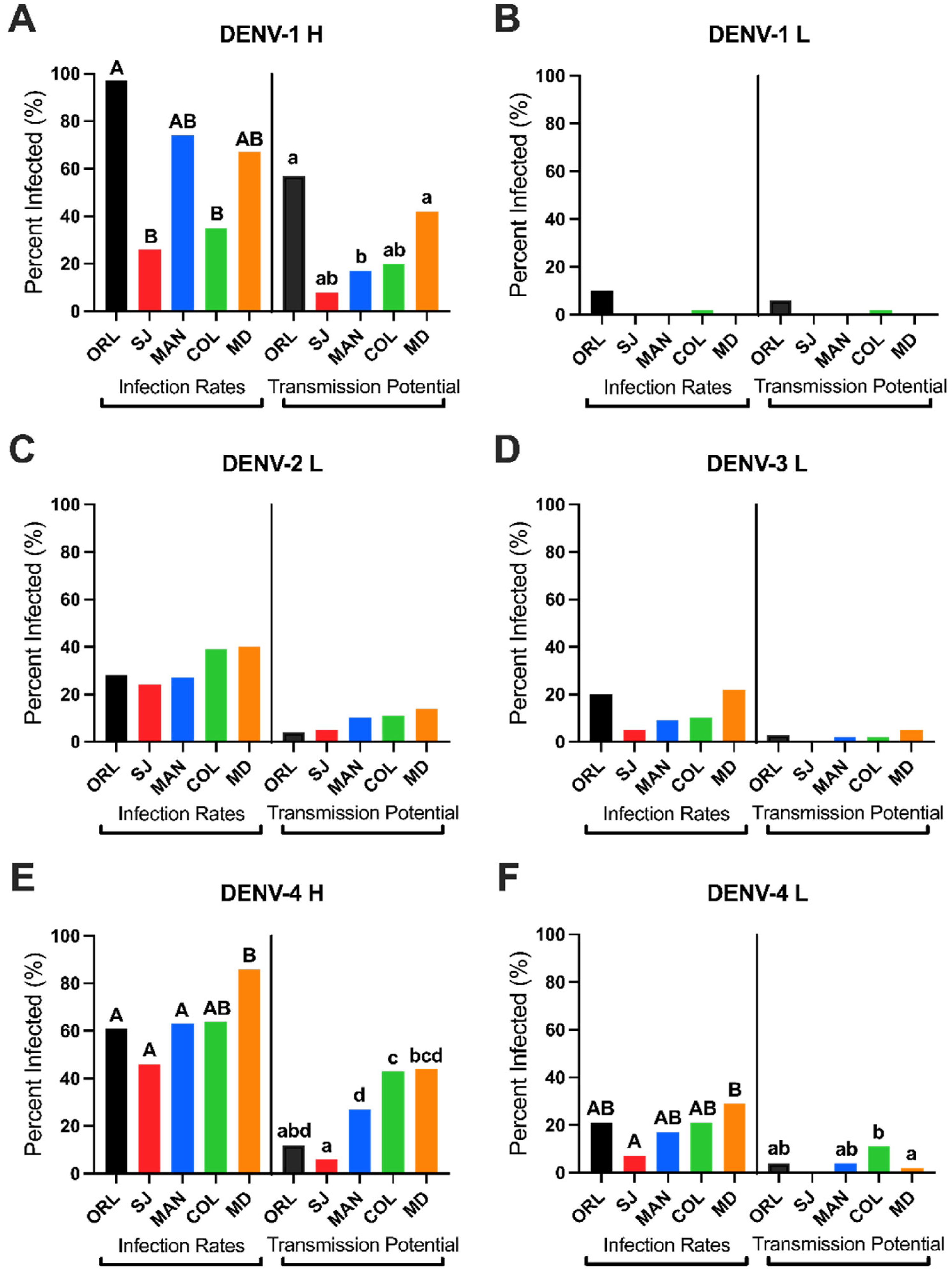
3.3. Field-Derived Ae. aegypti from South FL Have Comparatively Greater Vector Competence
3.4. Higher Infection and Transmission Potentials for Low Passage DENVs in Floridian Field Ae. aegypti
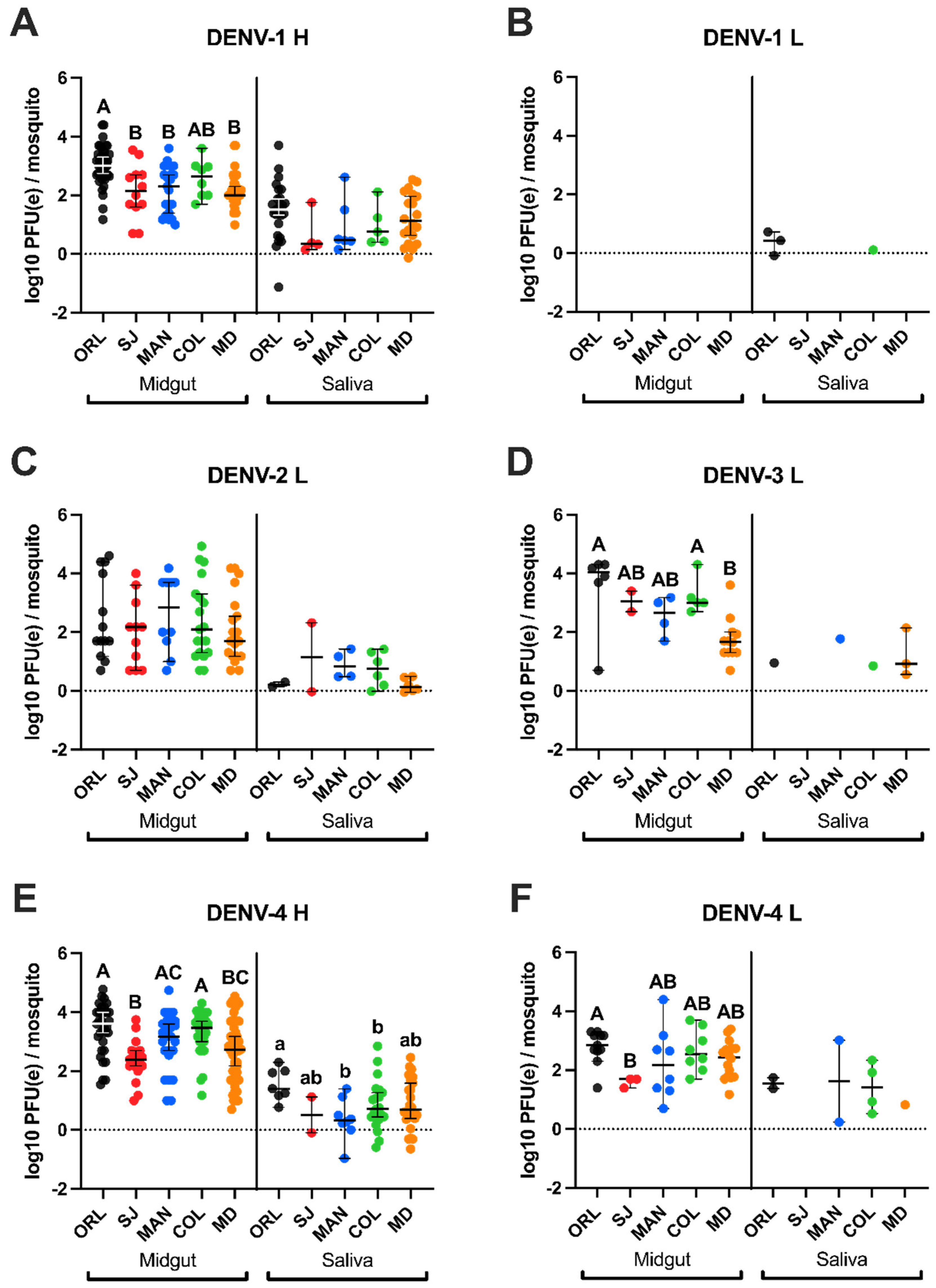
3.5. Low Passage DENV-4 Attained Highest Titers in Field Ae. aegypti Midgut Samples but Not the Highest Genome Copies in Saliva
3.6. Higher Infection Intensity for Laboratory Ae. aegypti (Orlando) Compared to Floridian Field Ae. aegypti
3.7. Knockdown Resistance Allele Profiles in Field-Derived Ae. aegypti Inversely Trends with Vector Competence for DENV
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Virus | Strain | Source | Year | GenBank ID | Average Blood Meal Titer (PFU/ mL) | Stock Titer (PFU/ mL) |
|---|---|---|---|---|---|---|
| DENV-1 H | Haiti/1207/2014 | Field isolate | 2014 | KT279761.2 | 6 × 106 | 8.71 × 106 |
| DENV-1 L | NR-82 Hawaii | ATCC | 1944 | KM204119 a | 6 × 105 | 9.5 × 105 |
| DENV-2 L | NR-84 New Guinea C [NGC] | ATCC | 1944 | KM204118 a | 5 × 106 | 6.17 × 106 |
| DENV-3 L | NR-80 Philippines/H87 | ATCC | 1956 | KU050695 a | 6.3 × 106 | 8.43 × 106 |
| DENV-4 H | Haiti/0075/2015 | Field isolate | 2015 | MK514144.1 | 2.5 × 106 | 1.68 × 107 |
| DENV-4 L | NR-86 H241 [TC] | ATCC | 1956 | KR011349 a | 1.6 × 106 | 8.7 × 106 |
| Primer Name | Sequence | Citation |
|---|---|---|
| DENV Forward 1 | 5′-AGGACYAGAGGTTAGAGGAGA-3′ | |
| DENV Reverse 1 | 5′-CGYTCTGTGCCTGGAWTGAT-3′ | [61] |
| DENV Probe 1 | 5′-FAM-ACAGCATATTGACGCTGGGARAGACC-BHQ1-3′ | |
| DENV Forward 2 | 5′ GGACTAGAGGTTAGAGGAGACCCC-3′ | [62] |
| DENV Reverse 2 | 5′-GAGACAGCAGGATCTCTGGTC-3′ | |
| DENV Probe 2 | 5′-FAM-AGCATATTGACGCTGGGA-BHQ1-3′ | |
| CFAV E Forward | 5′-GCTTCAAGTGGGGGATTGGA -3′ | [22] |
| CFAV E Reverse | 5′- CAACTTTCTCCATGCCGTGC -3′ | |
| V1016F | 5′- GCGGGCAGGGCGGCGGGGGCGGGGCCACAAATTGTTTCCCACCCGCACCGG -3′ | |
| I1016F | 5′-GCGGGCACAAATTGTTTCCCACCCGCACTGA-3′ | [43] |
| 1016R | 5′-GGATGAACCSAAATTGGACAAAAGC -3′ | |
| F1534F | 5′ GCGGGCTCTACTTTGTGTTCTTCATCATATT -3′ | |
| C1534F 1534R | 5′-GCGGGCAGGGCGGCGGGGGCGGGGCCTCTACTTTGTGTTCTTCATCATGTG-3′ 5′-TCTGCTCGTTGAAGTTGTCGAT-3′ | [44] |
| Mosquito Strain/DENV Strain | ORL | SJ | MD | COL | MAN |
|---|---|---|---|---|---|
| DENV-1 H | 46/44/26 | 43/12/4 | 46/31/19 | 29/8/5 | 32/21/6 |
| DENV-1 L | 66/6/3 | 46/0/0 | 36/0/0 | 29/1/1 | 31/0/0 |
| DENV-2 L | 50/15/2 | 46/11/2 | 53/21/6 | 43/19/6 | 35/10/4 |
| DENV-3 L | 52/8/1 | 40/2/0 | 49/12/3 | 47/5/1 | 44/4/1 |
| DENV-4 H | 55/33/7 | 35/17/2 | 53/46/24 | 40/28/21 | 39/25/11 |
| DENV-4 L | 51/11/2 | 37/3/0 | 54/16/1 | 36/8/4 | 48/8/2 |
| Genotypes | St. Johns | Manatee | Miami-Dade | Collier | ORL |
|---|---|---|---|---|---|
| VVFF | 0 | 0 | 0 | 0 | 100 |
| VVFC | 0 | 0 | 7 | 3 | 0 |
| VVCC | 0 | 37 | 10 | 6 | 0 |
| VIFF | 0 | 0 | 0 | 0 | 0 |
| VIFC | 0 | 0 | 7 | 19 | 0 |
| VICC | 0 | 50 | 38 | 52 | 0 |
| IIFF | 0 | 0 | 0 | 0 | 0 |
| IIFC | 0 | 0 | 0 | 3 | 0 |
| IICC | 100 | 13 | 38 | 16 | 0 |
References
- World Health Organization. Dengue and Severe Dengue. World Health Organization. Regional Office for the Eastern Mediterranean, 2014. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 21 April 2021).
- Bhatt, S.; Gething, P.; Brady, O.; Messina, J.P.; Farlow, A.W.; Moyes, C.; Drake, J.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.G.; Wint, G.R.W.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, J.D.; Shepard, D.S.; A Undurraga, E.; A Halasa, Y.; Coffeng, L.E.; Brady, O.; Hay, S.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.; et al. The global burden of dengue: An analysis from the Global Burden of Disease Study 2013. Lancet Infect. Dis. 2016, 16, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Halstead, S.B. Dengvaxia sensitizes seronegatives to vaccine enhanced disease regardless of age. Vaccine 2017, 35, 6355–6358. [Google Scholar] [CrossRef] [PubMed]
- Camargo, S. History of Aedes aegypti eradication in the Americas. Bull. World Health Organ. 1967, 36, 602–603. [Google Scholar]
- Hladish, T.J.; Pearson, C.A.B.; Rojas, D.P.; Gomez-Dantes, H.; Halloran, M.E.; Vazquez-Prokopec, G.M.; Longini, I.M. Forecasting the effectiveness of indoor residual spraying for reducing dengue burden. PLoS Negl. Trop. Dis. 2018, 12, e0006570. [Google Scholar] [CrossRef]
- Marini, G.; Guzzetta, G.; Toledo, C.A.M.; Teixeira, M.; Rosà, R.; Merler, S. Effectiveness of Ultra-Low Volume insecticide spraying to prevent dengue in a non-endemic metropolitan area of Brazil. PLoS Comput. Biol. 2019, 15, e1006831. [Google Scholar] [CrossRef] [Green Version]
- Guzzetta, G.; Trentini, F.; Poletti, P.; Baldacchino, F.A.; Montarsi, F.; Capelli, G.; Rizzoli, A.; Rosà, R.; Merler, S.; Melegaro, A. Effectiveness and economic assessment of routine larviciding for prevention of chikungunya and dengue in temperate urban settings in Europe. PLoS Negl. Trop. Dis. 2017, 11, e0005918. [Google Scholar] [CrossRef]
- Mahmud, M.A.F.; Mutalip, M.H.A.; Lodz, N.A.; Muhammad, E.N.; Yoep, N.; Hashim, M.H.; Paiwai, F.; Rajarethinam, J.; Aik, J.; Muhammad, N.A. Environmental management for dengue control: A systematic review protocol. BMJ Open 2019, 9, e026101. [Google Scholar] [CrossRef] [PubMed]
- Flores, H.; Neill, S.L.O. Controlling vector-borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Ba, Y.; Faye, O.; Soumare, M.L.; Dia, I.; Sall, A.A. Vector competence of Aedes aegypti populations from Senegal for sylvatic and epidemic dengue 2 virus isolated in West Africa. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Knox, T.B.; Kay, B.H.; Hall, R.A.; Ryan, P.A. Enhanced Vector Competence of Aedes aegypti (Diptera: Culicidae) from the Torres Strait Compared with Mainland Australia for Dengue 2 and 4 Viruses. J. Med. Entomol. 2003, 40, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; Anderson, S.L.; Alto, B.W. Vector competence of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) for dengue virus in the Florida Keys. J. Med. Entomol. 2012, 49, 942–946. [Google Scholar] [CrossRef]
- Gubler, D.J.; Nalim, S.; Tan, R.; Saipan, H.; Saroso, J.S. Variation in Susceptibility to Oral Infection with Dengue Viruses among Geographic Strains of Aedes aegypti. Am. J. Trop. Med. Hyg. 1979, 28, 1045–1052. [Google Scholar] [CrossRef]
- Stephenson, C.J.; Coatsworth, H.; Kang, S.; Lednicky, J.A.; Dinglasan, R.R. Transmission Potential of Floridian Aedes aegypti Mosquitoes for Dengue Virus Serotype 4: Implications for Estimating Local Dengue Risk. mSphere 2021, 6, e0027121. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L. Quantitative genetics of Aedes aegypti vector competence for dengue viruses: Towards a new paradigm? Trends Parasitol. 2011, 27, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic specificity and potential for local adaptation between dengue viruses and mosquito vectors. BMC Evol. Biol. 2009, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Lambrechts, L.; Quillery, E.; Noël, V.; Richardson, J.H.; Jarman, R.G.; Scott, T.W.; Chevillon, C. Specificity of resistance to dengue virus isolates is associated with genotypes of the mosquito antiviral gene Dicer-2. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122437. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, A.; Lequime, S.; Moltini-Conclois, I.; Jiolle, D.; Leparc-Goffart, I.; Reiner, R.C., Jr.; Lambrechts, L. Epidemiological significance of dengue virus genetic variation in mosquito infection dynamics. PLoS Pathog. 2018, 14, e1007187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baidaliuk, A.; Miot, E.F.; Lequime, S.; Moltini-Conclois, I.; Delaigue, F.; Dabo, S.; Dickson, L.B.; Aubry, F.; Merkling, S.H.; Cao-Lormeau, V.M.; et al. Cell-Fusing Agent Virus Reduces Arbovirus Dissemination in Aedes aegypti Mos-quitoes In Vivo. J. Virol. 2019, 18, e00705-19. [Google Scholar]
- Morales-Vargas, R.E.; Missé, D.; Chavez, I.F.; Kittayapong, P. Vector Competence for Dengue-2 Viruses Isolated from Patients with Different Disease Severity. Pathogens 2020, 9, 859. [Google Scholar] [CrossRef]
- Gutiérrez-Bugallo, G.; Boullis, A.; Martinez, Y.; Hery, L.; Rodríguez, M.; Bisset, J.A.; Vega-Rúa, A. Vector competence of Aedes aegypti from Havana, Cuba, for dengue virus type 1, chikungunya, and Zika viruses. PLoS Negl. Trop. Dis. 2020, 14, e0008941. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Bargielowski, I.; Carrasquilla, M.C.; Nishimura, N. Coexistence of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in peninsular Florida two decades after competitive displacements. J. Med. Entomol. 2016, 53, 1385–1390. [Google Scholar] [CrossRef]
- CDC. Centers for Disease Control and Prevention. ArboNET Disease Maps. Available online: https://wwwn.cdc.gov/arbonet/Maps/ADB_Diseases_Map/index.html (accessed on 21 April 2021).
- Teets, F.D.; Ramgopal, M.N.; Sweeney, K.D.; Graham, A.S.; Michael, S.F.; Isern, S. Origin of the dengue virus outbreak in Martin County, Florida, USA 2013. Virol. Rep. 2014, 1–2, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Morrison, A.; Rowe, D.; McDermott, C.; Stanek, D.; Heberlein-Larson, L.; LaCrue, A.; Castaneda, M.; Mock, V.; Blackmore, C. Florida Arbovirus Surveillance Report. Week 51: 22–28 December, 2019. Available online: http://www.floridahealth.gov/diseases-and-conditions/mosquito-borne-diseases/_documents/2019-week-51-arbovirus-surveillance-report.pdf (accessed on 21 April 2021).
- Buckner, E.A.; Alto, B.W.; Lounibos, L.P. Vertical Transmission of Key West Dengue-1 Virus by Aedes aegypti and Aedes albopictus (Diptera: Culicidae) Mosquitoes From Florida. J. Med. Entomol. 2013, 50, 1291–1297. [Google Scholar] [CrossRef] [Green Version]
- Rey, J.R. Dengue in Florida (USA). Insects 2014, 5, 991–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, A.S.; Pruszynski, C.A.; Hribar, L.J.; DeMay, D.J.; Tambasco, A.N.; Hartley, A.E.; Fussell, E.M.; Michael, S.F.; Isern, S. Mosquito-associated Dengue Virus, Key West, Florida, USA, 2010. Emerg. Infect. Dis. 2011, 17, 2074–2075. [Google Scholar] [CrossRef] [PubMed]
- Radke, E.; Gregory, C.J.; Kintziger, K.; Sauber-Schatz, E.K.; Hunsperger, E.A.; Gallagher, G.R.; Barber, J.M.; Biggerstaff, B.J.; Stanek, D.R.; Tomashek, K.M.; et al. Dengue Outbreak in Key West, Florida, USA, 2009. Emerg. Infect. Dis. 2012, 18, 135–137. [Google Scholar] [CrossRef]
- Morrison, A.; Rowe, D.; McDermott, C.; Stanek, D.; Heberlein-Larson, L.; LaCrue, A.; Castaneda, M.; Mock, V.; Blackmore, C. Florida Arbovirus Surveillance Week 52: 20–26 December, 2020. Available online: http://www.floridahealth.gov/diseases-and-conditions/mosquito-borne-diseases/_documents/2020-week-52-arbovirus-surveillance-report.pdf (accessed on 21 April 2021).
- Alto, B.W.; Smartt, C.T.; Shin, D.; Bettinardi, D.; Malicoate, J.; Anderson, S.L.; Richards, S.L. Susceptibility of Florida Aedes aegypti and Aedes albopictus to dengue viruses from Puerto Rico. J. Vector Ecol. 2014, 39, 406–413. [Google Scholar] [CrossRef]
- Mundis, S.J.; Estep, A.S.; Waits, C.M.; Ryan, S.J. Spatial variation in the frequency of knockdown resistance genotypes in Florida Aedes aegypti populations. Parasites Vectors 2020, 13, 1–12. [Google Scholar] [CrossRef]
- Estep, A.S.; Sanscrainte, N.D.; Waits, C.M.; Bernard, S.J.; Lloyd, A.M.; Lucas, K.J.; Buckner, E.A.; Vaidyanathan, R.; Morreale, R.; Conti, L.A.; et al. Quantification of permethrin resistance and kdr alleles in Florida strains of Aedes aegypti (L.) and Aedes albopictus (Skuse). PLoS Negl. Trop. Dis. 2018, 12, e0006544. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-Y.; Smartt, C.; Shin, D. Permethrin Resistance in Aedes aegypti Affects Aspects of Vectorial Capacity. Insects 2021, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Parker-Crockett, C.; Connelly, C.R.; Siegfried, B.; Alto, B. Influence of Pyrethroid Resistance on Vector Competency for Zika Virus by Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2021, 58, 1908–1916. [Google Scholar] [CrossRef] [PubMed]
- Dick, O.B.; Martín, J.L.S.; Del Diego, J.; Montoya, R.H.; Dayan, G.H.; Zambrano, B. The History of Dengue Outbreaks in the Americas. Am. J. Trop. Med. Hyg. 2012, 87, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Boyles, S.M.; Mavian, C.N.; Finol, E.; Ukhanova, M.; Stephenson, C.J.; Hamerlinck, G.; Kang, S.; Baumgartner, C.; Geesey, M.; Stinton, I.; et al. Under-the-Radar Dengue Virus Infections in Natural Populations of Aedes aegypti Mosquitoes. mSphere 2020, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.W.; Comrie, A.C.; Ernst, K. Climate and Dengue Transmission: Evidence and Implications. Environ. Health Perspect. 2013, 121, 1264–1272. [Google Scholar] [CrossRef]
- Li, Y.; Dou, Q.; Lu, Y.; Xiang, H.; Yu, X.; Liu, S. Effects of ambient temperature and precipitation on the risk of dengue fever: A systematic review and updated meta-analysis. Environ. Res. 2020, 191, 110043. [Google Scholar] [CrossRef]
- Saavedra-Rodriguez, K.; Urdaneta-Marquez, L.; Rajatileka, S.; Moulton, M.; Flores, A.E.; Fernandez-Salas, I.; Bisset, J.; Rodriguez, M.; Mccall, P.J.; Donnelly, M.J.; et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 2007, 16, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Somwang, P.; Prapanthadara, L.-A. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop. Med. Int. Health 2011, 16, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Black IV, W.C.; Bennett, K.E.; Gorrochótegui-Escalante, N.; Barillas-Mury, C.V.; Fernández-Salas, I.; de Lourdes Muñoz, M.; Farfán-Alé, J.A.; Olson, K.E.; Beaty, B.J. Flavivirus susceptibility in Aedes aegypti. Arch. Med. Res. 2002, 33, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Mercado-Curiel, R.F.; Esquinca-Avilés, H.A.; Tovar, R.; Díaz-Badillo, Á.; Camacho-Nuez, M.; Muñoz, M.D.L. The four serotypes of dengue recognize the same putative receptors in Aedes aegypti midgut and Ae. albopictus cells. BMC Microbiol. 2006, 6, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, M.S.; Edgil, D.; Roberts, T.G.; Lu, B.; Harris, E. Infection of human cells by dengue virus is modulated by dif-ferent cell types and viral strains. J. Virol. 2000, 74, 7814–7823. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Brown, H.E.; Rico-Hesse, R. Variation in Vector Competence for Dengue Viruses Does Not Depend on Mosquito Midgut Binding Affinity. PLoS Negl. Trop. Dis. 2011, 5, e1172. [Google Scholar] [CrossRef] [Green Version]
- Khoo, C.C.; Doty, J.B.; Held, N.L.; Olson, K.E.; Franz, A.W. Isolation of midgut escape mutants of two American gen-otype dengue 2 viruses from Aedes aegypti. Virol. J. 2013, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.-S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya Isolates from the Outbreak of La Reunion (Indian Ocean) Exhibit Different Patterns of Infection in the Mosquito, Aedes albopictus. PLoS ONE 2007, 2, e1168. [Google Scholar] [CrossRef] [PubMed]
- Blok, J. Genetic Relationships of the Dengue Virus Serotypes. J. Gen. Virol. 1985, 66, 1323–1325. [Google Scholar] [CrossRef] [PubMed]
- Añez, G.; Heisey, D.A.; Volkova, E.; Rios, M. Complete Genome Sequences of Dengue Virus Type 1 to 4 Strains Used for the Development of CBER/FDA RNA Reference Reagents and WHO International Standard Candidates for Nucleic Acid Testing. Genome Announc. 2016, 4, e01583-15. [Google Scholar] [CrossRef] [Green Version]
- Kuno, G.; Chang, G.-J.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angleró-Rodríguez, Y.I.; MacLeod, H.J.; Kang, S.; Carlson, J.S.; Jupatanakul, N.; Dimopoulos, G. Aedes aegypti Molecular Responses to Zika Virus: Modulation of Infection by the Toll and Jak/Stat Immune Pathways and Virus Host Factors. Front. Microbiol. 2017, 8, 2050. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.; Jupatanakul, N.; Ramirez, J.L.; Kang, S.; Romero-Vivas, C.M.; Mohammed, H.; Dimopoulos, G. Transcriptomic profiling of diverse Aedes aegypti strains reveals increased basal-level immune activation in dengue virus-refractory pop-ulations and identifies novel virus-vector molecular interactions. PLoS Negl. Trop. Dis. 2013, 7, e2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taracena, M.L.; Bottino-Rojas, V.; Talyuli, O.A.; Walter-Nuno, A.B.; Oliveira, J.H.M.; Angleró-Rodriguez, Y.I.; Wells, M.B.; Dimopoulos, G.; Oliveira, P.L.; Paiva-Silva, G.O. Regulation of midgut cell proliferation impacts Aedes aegypti sus-ceptibility to dengue virus. PLoS Negl. Trop. Dis. 2018, 12, e0006498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerdan, B.R. Dengue fever/dengue hemorrhagic fever. Am. J. Clin. Med. 2010, 7, 51–53. [Google Scholar]
- de Lima, V.H.F.; Andrade, P.D.S.; Thomazelli, L.M.; Marrelli, M.T.; Urbinatti, P.R.; Almeida, R.M.M.D.S.; Lima-Camara, T.N. Silent circulation of dengue virus in Aedes albopictus (Diptera: Culicidae) resulting from natural vertical transmission. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Sánchez-Vargas, I.; Harrington, L.C.; Doty, J.B.; Black 4th, W.C.; Olson, K.E. Demonstration of efficient vertical and venereal transmission of dengue virus type-2 in a genetically diverse laboratory strain of Aedes aegypti. PLoS. Negl. Trop. Dis. 2018, 12, e0006754. [Google Scholar] [CrossRef] [Green Version]
- Ayers, J.B.; Xie, X.; Coatsworth, H.; Stephenson, C.J.; Waits, C.M.; Shi, P.-Y.; Dinglasan, R.R. Ecology and transmission of a dengue virus serotype 4 identified in wild Aedes aegypti in Florida. Front. Microbiol. 2021, 12, 734903. [Google Scholar] [CrossRef]
- Leparc-Goffart, I.; Baragatti, M.; Temmam, S.; Tuiskunen, A.; Moureau, G.; Charrel, R.; de Lamballerie, X. Development and validation of real-time one-step reverse transcription-PCR for the detection and typing of dengue viruses. J. Clin. Virol. 2009, 45, 61–66. [Google Scholar] [CrossRef]
- Huhtamo, E.; Hasu, E.; Uzcátegui, N.Y.; Erra, E.; Nikkari, S.; Kantele, A.; Vapalahti, O.; Piiparinen, H. Early diagnosis of dengue in travelers: Comparison of a novel real-time RT-PCR, NS1 antigen detection and serology. J. Clin. Virol. 2010, 47, 49–53. [Google Scholar] [CrossRef]



| Mosquito Strain/ DENV Strain * | ORL | SJ | MAN | COL | MD | |
|---|---|---|---|---|---|---|
| Infection Rates (%) | DENV-1 H | a 97 A | acd 26 B | ad 74 AB | ad 35 B | a 67 AB |
| DENV-1 L | b 10 | 0 | 0 | b 2 | 0 | |
| DENV-2 L | c 28 | c 24 | b 27 | a 39 | b 40 | |
| DENV-3 L | bc 20 | b 5 | bc 9 | d 10 | c 22 | |
| DENV-4 H | d 61 A | d 46 A | d 63 A | c 64 AB | d 86 B | |
| DENV-4 L | bc 21 AB | bc 7 A | b 17 AB | abd 21 AB | bc 29 B | |
| Transmission Potentials (%) | DENV-1 H | a 57 A | 8 AB | 17 B | ab 20 AB | a 42 A |
| DENV-1 L | ab 6 | 0 | 0 | ab 2 | 0 | |
| DENV-2 L | b 4 | 5 | 10 | a 11 | bc 14 | |
| DENV-3 L | b 3 | 0 | 2 | a 2 | bc 5 | |
| DENV-4 H | b 12 ABD | 6 A | 27 D | b 43 C | ac 44 BCD | |
| DENV-4 L | b 4 AB | 0 | 4 AB | ab 11 B | b 2 A |
| Mosquito Strain/ DENV Strain * | ORL | SJ | MAN | COL | MD | |
|---|---|---|---|---|---|---|
| Midgut (average [SD]) | DENV-1 H | a 3.03 (0.65) A | 2.13 (0.92) B | a 2.18 (0.80) B | a 2.57 (0.65) AB | a 2.18 (0.60) B |
| DENV-1 L | -- | -- | -- | -- | -- | |
| DENV-2 L | b 2.46 (1.41) | 2.00 (1.16) | ab 2.64 (1.29) | a 2.41 (1.33) | a 2.09 (1.14) | |
| DENV-3 L | ac 3.51 (1.40) A | 3.05 (0.49) AB | ab 2.54 (0.68) AB | ab 3.24 (0.62) A | a 1.75 (0.74) B | |
| DENV-4 H | c 3.43 (0.85) A | 2.37 (0.71) B | b 3.02 (0.97) AC | b 3.28 (0.76) A | b 2.73 (1.09) BC | |
| DENV-4 L | ab 2.80 (0.56) A | 1.60 (0.17) B | a 2.25 (1.21) AB | ab 2.67 (0.71) AB | ab 2.36 (0.63) AB | |
| Saliva (average [SD]) | DENV-1 H | a 1.48 (0.96) | 0.65 (0.74) | ab 0.95 (0.94) | 0.99 (0.72) | a 1.22 (0.84) |
| DENV-1 L | b 0.36 (0.41) | -- | -- | 0.11 | -- | |
| DENV-2 L | b 0.22 (0.11) | 1.14 (1.66) | ab 0.89 (0.48) | 0.74 (0.60) | b 0.19 (0.23) | |
| DENV-3 L | 0.95 | -- | ab 1.77 | 0.85 | ab 1.2 (0.83) | |
| DENV-4 H | a 1.55 (0.55) A | 0.51 (0.86) AB | a 0.37 (0.72) B | 0.85 (0.84) B | ab 0.87 (0.87) AB | |
| DENV-4 L | ab 1.56 (0.26) | -- | b 1.63 (1.96) | 1.43 (0.84) | ab 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stephenson, C.J.; Coatsworth, H.; Waits, C.M.; Nazario-Maldonado, N.M.; Mathias, D.K.; Dinglasan, R.R.; Lednicky, J.A. Geographic Partitioning of Dengue Virus Transmission Risk in Florida. Viruses 2021, 13, 2232. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112232
Stephenson CJ, Coatsworth H, Waits CM, Nazario-Maldonado NM, Mathias DK, Dinglasan RR, Lednicky JA. Geographic Partitioning of Dengue Virus Transmission Risk in Florida. Viruses. 2021; 13(11):2232. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112232
Chicago/Turabian StyleStephenson, Caroline J., Heather Coatsworth, Christy M. Waits, Nicole M. Nazario-Maldonado, Derrick K. Mathias, Rhoel R. Dinglasan, and John A. Lednicky. 2021. "Geographic Partitioning of Dengue Virus Transmission Risk in Florida" Viruses 13, no. 11: 2232. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112232