
Molecular and Serological Characterization of the SARS-CoV-2 Delta Variant in Bangladesh in 2021

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Sample Collections
2.2. Quantification of Viral RNA
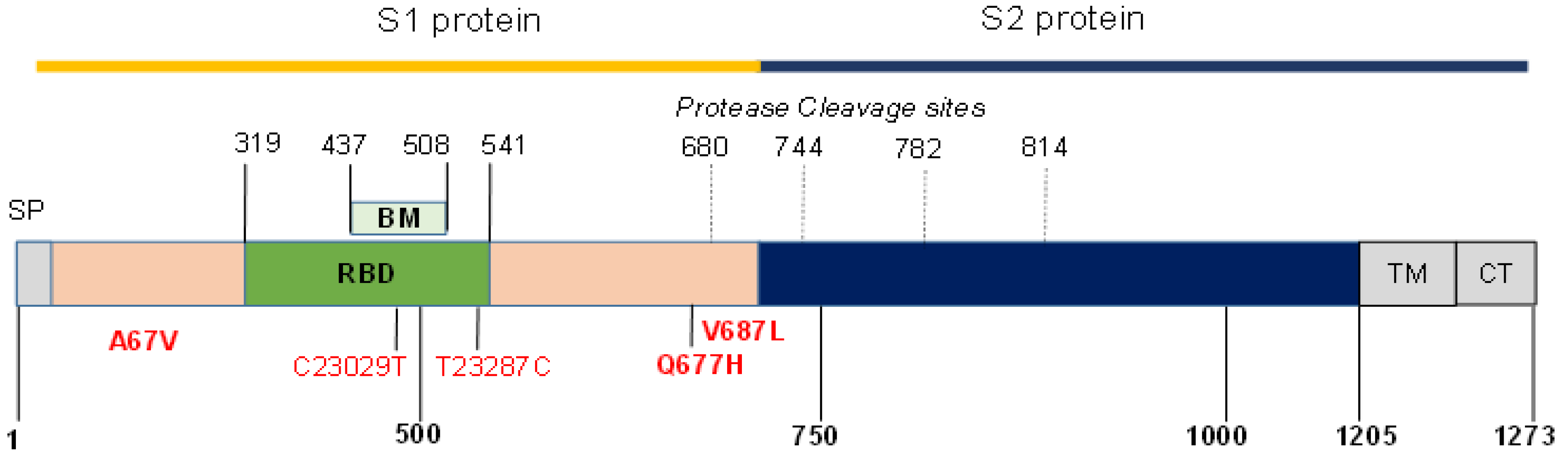
2.3. Screening of Spike Protein Mutations and Sequencing
2.4. Bioinformatics
2.5. Assessment of Anti-SARS CoV-2 Antibodies
2.6. Statistical Analysis
3. Results
3.1. SARS CoV-2 Variants
3.2. Bangladeshi COVID-19 Patient Characteristics
3.3. SARS CoV-2 Variant Viral Loads
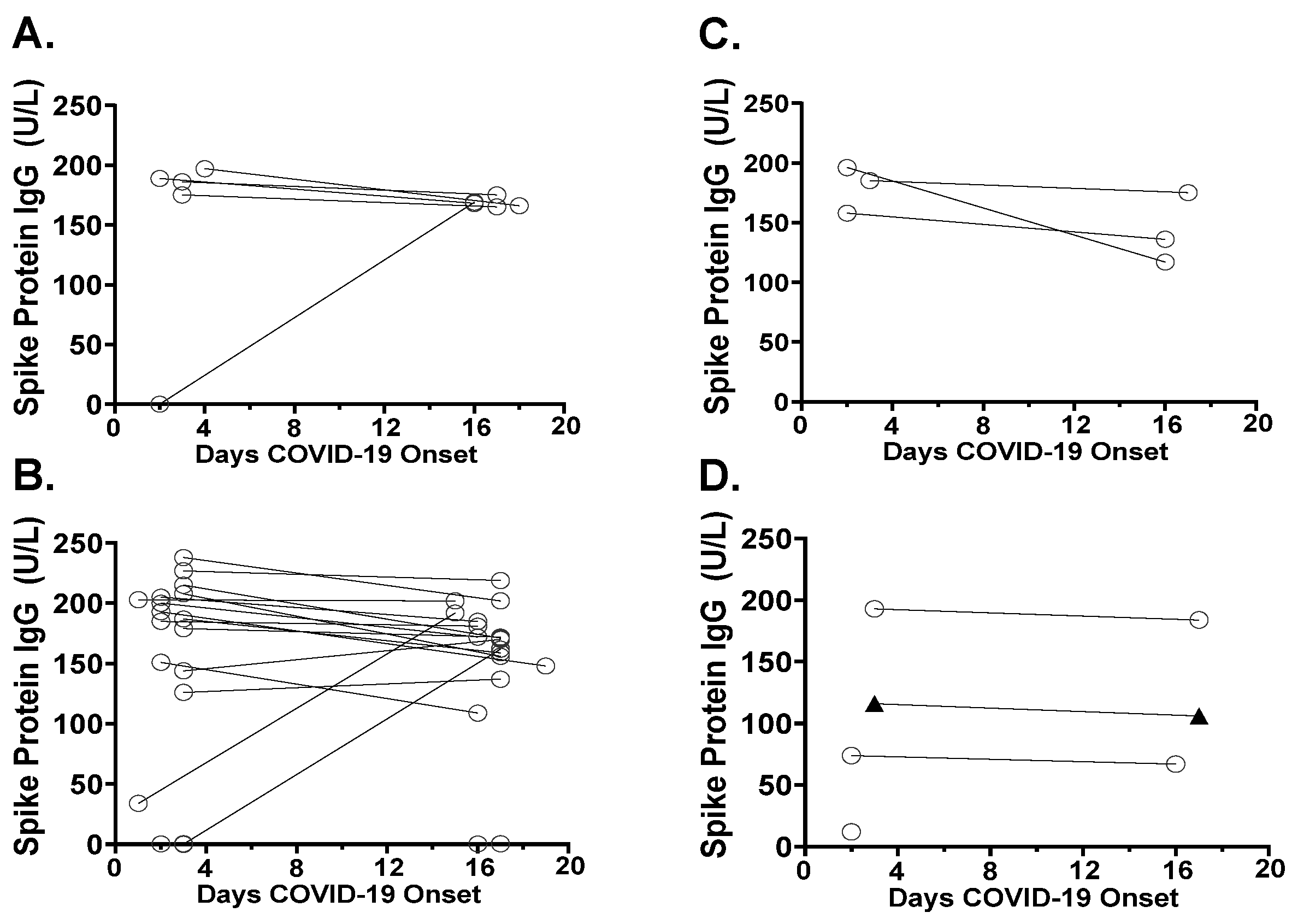
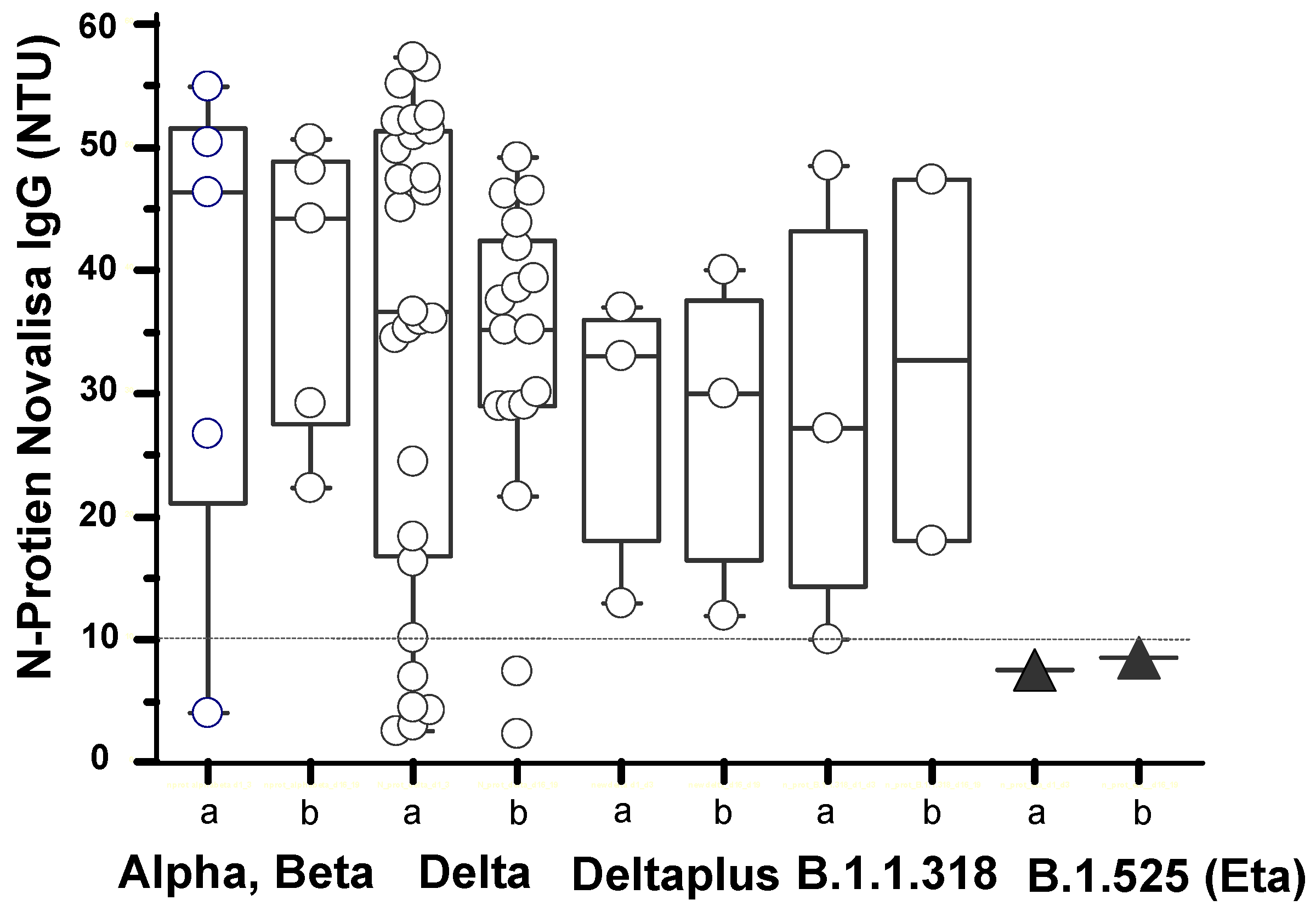
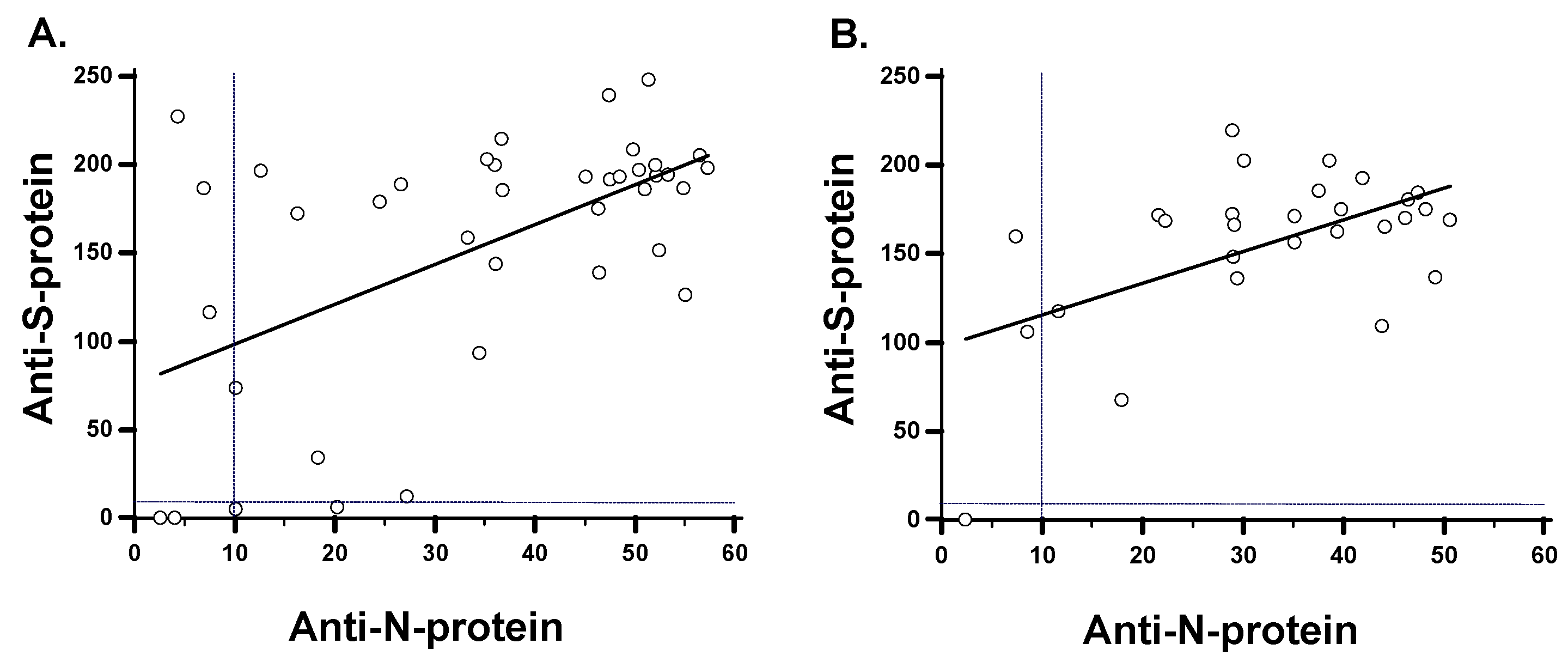
3.4. Antibody Responses in SARS-CoV-2 Variants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- John Hopkins University of Medicine Coronavirus. Available online: https://0-coronavirus-jhu-edu.brum.beds.ac.uk/ (accessed on 20 September 2021).
- European Medical Agency EMA. Available online: https://www.ema.europa.eu/en (accessed on 9 November 2021).
- Ball, P. The speedy approach used to tackle SARS-CoV-2 could change the future of vaccine science. Nature 2021, 589, 16–18. [Google Scholar] [CrossRef]
- Thanh, T.; Andreadakis, Z.; Kumar, A.; Gomez-Roman, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. 2020, 19, 305–306. [Google Scholar] [CrossRef]
- De Oliveira, B.R.; Da Penha Sobral, A.I.G.; Marinho, M.L.; Sobral, M.F.F.; De Souza Melo, A.; Duarte, B.G. Determinants of access to the SARS-CoV-2 vaccine: A preliminary approach. Int. J. Equity Health 2021, 20, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Kaim, A.; Siman-Tov, M.; Jaffe, E.; Adini, B. From isolation to containment: Perceived fear of infectivity and protective behavioral changes during the COVID-19 vaccination campaign. Int. J. Environ. Res. Public Health 2021, 18, 6503. [Google Scholar] [CrossRef]
- Siam, H.B.; Hasan, M.; Tashrif, S.M.; Khan, H.R.; Raheem, E.; Hossain, M.S. Insights into the first seven-months of COVID-19 pandemic in Bangladesh: Lessons learned from a high risk country. Heliyon 2021, 7, e07385. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.L.; Wiysonge, C. Social media and vaccine hesitancy. BMJ Glob. Health 2020, 5, e004206. [Google Scholar] [CrossRef]
- Rella, S.A.; Kulikova, Y.A.; Dermitzakis, E.T.; Kondrashov, F.A. Rates of SARS-CoV-2 transmission and vaccination impact the fate of vaccine-resistant strains. Nat. Sci. Rep. 2021, 11, 15279. [Google Scholar] [CrossRef] [PubMed]
- Kames, J.; Holcomb, D.D.; Kimchi, O.; DiCuccio, M.; Hamasaki-Katagiri, N.; Wang, T.; Komar, A.A.; Alexaki, A.; Kimchi-Sarfaty, C. Sequence analysis of SARS-CoV-2 genome reveals features important for vaccine design. Nat. Sci. Rep. 2020, 10, 15643. [Google Scholar] [CrossRef]
- Pritchard, E.; Matthews, P.C.; Stoesser, N.; Eyre, D.W.; Gethings, O.; Vihta, K.D.; Jones, J.; House, T.; VanSteenHouse, H.; Bell, I.; et al. Impact of vaccination on new SARS-CoV-2 infections in the United Kingdom. Nat. Med. 2021, 27, 1370–1378. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Tho mson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A. COVID-19 Genomics UK (COG-UK) Consortium, Peacock SJ and Robertson DL, SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Farinholt, T.; Doddapaneni, H.; Qin, X.; Menon, V.; Meng, Q.; Metcalf, G.; Chao, H.; Gingras, M.C.; Farinholt, P.; Agrawal, C.; et al. Transmission event of SARS-CoV-2 Delta variant reveals multiple vaccine breakthrough infections. medRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Elbe, S.; Buckland-Merrett, G. Data, disease and diplomacy: GISAID’s innovative contribution to global health. Glob. Chall. 2017, 1, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.M.; Kim, I.H.; Kim, S. Nucleic acid testing of SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 6150. [Google Scholar] [CrossRef]
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; Du Plessis, L.; Pybus, O.G. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef]
- Rules for the Designation and Naming of Pango Lineages. Available online: https://www.pango.network/the-pango-nomenclature-system/statement-of-nomenclature-rules/ (accessed on 9 November 2021).
- SARS-CoV-2 Variant Classifications and Definitions. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/variant-info.html (accessed on 4 October 2021).
- Investigation of SARS-CoV-2 Variants of Concern: Technical Briefings. Available online: https://www.gov.uk/government/publications/investigation-of-novel-sars-cov-2-variant-variant-of-concern-20201201 (accessed on 9 November 2021).
- Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 10 November 2021).
- Barros-Martins, J.; Hammerschmidt, S.I.; Cossmann, A.; Odak, I.; Stankov, M.V.; Morillas Ramos, G.; Dopfer-Jablonka, A.; Heidemann, A.; Ritter, C.; Friedrichsen, M.; et al. Immune responses against SARS-CoV-2 variants after heterologous and homologous ChAdOx1 nCoV-19/BNT162b2 vaccination. Nat. Med. 2021, 27, 1525–1529. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Bernal, J.L.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of Covid-19 Vaccines against the B.1.617.2 (Delta) Variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef]
- Bangladesh Directorate General of Health Services, Ministry of Health and Family Welfare. Coronavirus COVID-19 Dashboard. 2021. Available online: http://103.247.238.92/webportal/pages/covid19.php (accessed on 23 September 2021).
- Saha, S.; Tanmoy, A.M.; Hooda, Y.; Akter, A.T.; Goswami, S.; Sium, S.M.; Islam, S.M.; Malaker, R.; Islam, S.; Rahman, H.; et al. COVID-19 rise in Bangladesh correlates with increasing detection of B.1.351 variant. Brit. Med. J. Glob. Health 2021, 6, e006012. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Shirin, T.; Rahman, S.; Rahman, M.M.; Hossain, M.E.; Hossain, M.K.; Rahman, M.Z.; El Arifeen, S.; Ahmed, T. The emergence of SARS-CoV-2 variants in Dhaka city, Bangladesh. Transbound. Emerg. Dis. 2021, 1, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Sayeed, M.A.; Rahman, M.K.; Zamil, S.; Abedin, J.; Saha, O.; Hassan, M.M. Assessment of basic reproduction number (R0), spatial and temporal epidemiological determinants, and genetic characterization of SARS-CoV-2 in Bangladesh. Infect. Genet. Evol. 2021, 92, 104884. [Google Scholar] [CrossRef]
- Hasan, M.M.; Rocha, I.C.N.; Ramos, K.G.; Cedeño, T.D.D.; Dos Santos Costa, A.C.; Tsagkaris, C.; Billah, M.; Ahmad, S.; Essar, M.Y. Emergence of highly infectious SARS-CoV-2 variants in Bangladesh: The need for systematic genetic surveillance as a public health strategy. Trop. Med. Health BMC 2021, 46, 69. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.; Bhattacharjee, A.; Rahmat, R.; Islam, S.R.; Akhter, T.; Ahammad, I.; Hossain, M.U.; Munshi, S.U.; Salimullahd, M. Genomic surveillance of SARS-CoV-2 viruses collected during the ending phase of the first wave of the COVID-19 pandemic in Bangladesh. Microbiol. Resour. Announc. 2021, 29, e00496-21. [Google Scholar] [CrossRef]
- Hasan, M.M.; Das, R.; Rasheduzzaman, M.; Hussain, M.H.; Muzahid, N.H.; Salauddin, A.; Rumi, M.H.; Rashid, S.M.M.; Siddiki, Z.; Mannan, A. Global and local mutations in Bangladeshi SARS-CoV-2 genomes. Virus Res. 2021, 297, 198390. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.E.; Rahman, M.M.; Alam, M.S.; Karim, Y.; Hoque, A.F.; Rahman, S.; Rahman, M.Z.; Rahmana, M. Genome sequence of a SARS-CoV-2 strain from Bangladesh that is nearly identical to United Kingdom SARS-CoV-2 variant B.1.1.7. Microbiol. Resour. Announc. 2021, 10, e00100-21. [Google Scholar] [CrossRef]
- Sarkar, M.M.H.; Rabbi, M.F.A.; Akter, S.; Banu, T.A.; Goswami, B.; Jahan, I.; Hossain, M.S.; Osman, E.; Uzzaman, M.S.; Habib, A.; et al. Genome sequence of a SARS-CoV-2 P.1 variant of concern (20J/501Y.V3) from Bangladesh. Microbiol. Resour. Announc. 2021, 10, e00524-21. [Google Scholar] [CrossRef] [PubMed]
- Daria, A.A.; Assaduzzaman, M.S.; Islam, M.R. Bangladesh reported delta variant of coronavirus among its citizen: Actionable items to tackle the potential massive third wave. Infect. Prev. Pract. 2021, 3, 100159. [Google Scholar]
- Das, A.; Khurshid, S.; Ferdausi, A.; Nipu, E.S.; Das, A.; Ahmed, F.F. Molecular insight into the genomic variation of SARS-CoV-2 strains from current outbreak. Comput. Biol. Chem. 2021, 93, 10753. [Google Scholar] [CrossRef]
- Nagy, A.; Basiouni, S.; Prvin, R.; Hafez, H.; Shehata, A.A. Evolutionary insights into the furin cleavage sites of SARS-CoV-2 variants from humans and animals. Arch. Virol. 2021, 166, 2541–2549. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, F.; Preibisch, G.; Gizinaki, S.; Kochanczyk, M.; Lipniack, T. SARS-CoV-2 variant of concern 202012/01 has about twofold replicative advantage and acquires concerning mutations. Viruses 2021, 13, 392. [Google Scholar] [CrossRef]
- Grabowski, F.; Preibisch, G.; Gizinski, S.; Kochanczyk, M.; Lipniack, T. Immune evasion of SARS-CoV-2 emerging variants: What have we learnt so far? Viruses 2021, 13, 1192. [Google Scholar] [CrossRef]
- Tegally, H.; Ramuth, M.; Amoaka, D.; Scheepers, C.; Wilkinson, E.; Giovanetti, M.; Lessells, R.J.; Giandhari, J.; Ismail, A.; Martin, D.; et al. Genomic epidemiology of SARS-CoV-2 in Mauritius reveals a new wave of infections dominated by the B.1.1.318, a variant under investigation. medRxiv 2021. [Google Scholar] [CrossRef]
- Wei, J.; Matthews, P.C.; Stoesser, N.; Maddox, T.; Lorenzi, L.; Studley, R.; Bell, J.I.; Newton, J.N.; Farrar, J.; Diamond, I.; et al. Anti-spike antibody response to natural SARS-CoV-2 infection in the general population. Nat. Commun. 2021, 12, 6250. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Dolscheid-Pommerich, R.; Bartok, E.; Renn, M.; Kümmerer, B.M.; Schulte, B.; Schmithausen, R.M.; Stoffel-Wagner, B.; Streeck, H.; Saschenbrecker, S.; Steinhagen, K.; et al. Correlation between a quantitative SARS CoV-2 IgG ELISA and neutralization activity. J. Med. Virol. 2021, 94, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Brochot, E.; Demey, B.; Touzé, A.; Belouzard, S.; Dubuisson, J.; Schmit, J.L.; Duverlie, G.; Francois, C.; Castelain, S.; Helle, F. Anti-spike, anti-nucleocapsid and neutralizing antibodies in SARS-CoV-2 in patients and asymptomatic individuals. Front. Microbiol. 2020, 11, 584251. [Google Scholar] [CrossRef] [PubMed]
- Lytton, S.; Yeasmin, M.; Ghosh, A.; Bulbul, R.H.; Molla, M.A.; Herr, M.; Duchmann, H.; Sharif, M.; Nafisa, T.; Amin, R.; et al. Detection of anti-nucleocapsid antibody in COVID-19 patients in Bangladesh is not correlated with previous dengue infection. Pathogens 2021, 10, 637. [Google Scholar] [CrossRef]
- Egbert, E.R.; Xiao, S.; Colantuoni, E.; Caturegli, P.; Gadala, A.; Milstone, A.M.; Debes, A.K. Durability of spike immunoglobin G antibodies to SARS-CoV-2 among health care workers with prior infection. JAMA 2021, 4, e2123256. [Google Scholar] [CrossRef]
- Swadźba, J.; Bednarczyk, M.; Anyszek, T.; Kozlowska, D.; Panek, A.; Martin, E. The real life performance of 7 automated anti-SARS-CoV-2 IgG and IgM/IgA immunoassays. Pract. Lab. Med. 2021, 25, e00212. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | SARS-CoV-2 Alpha, Beta Group 1 | SARS-CoV-2 Delta Group 2 | SARS-CoV-2 Delta Plus Group 3 | SARS-CoV-2 B.1.1.318, Eta Group 4 | p Value 1 vs. 2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Number of cases, N | 5 | 24 | 3 | 4 | NR | ||||
| SARS CoV-2 Variant, % | 14 | 67 | 8 | 11 | 0.03 | ||||
| Age as a median (range), years | 45 (30–55) | 48 (29–87) | 44 (32–55) | 40 (28–52) | 0.39 (b) | ||||
| Gender, M/F | 2/3 | 17/7 | 0/3 | 2/2 | 0.2 | ||||
| Hospitalized, n (%) | 5 (100) | 23 (96) | 2 (67) | 4 (100) | 0.65 | ||||
| Oxygen support, n (%) | 1 (20) | 11 (46) | 0 (0) | 3 (75) | 0.29 | ||||
| Difficulty breathing, n (%) | 1 (20) | 22 (46) | 0 (0) | 3 (75) | 0.29 | ||||
| Days post onset of COVID-19 symptoms as a median (range) | 3 (2–4) | 2 (1–3) | 2 (2–3) | 2 (2–3) | 0.65 (b) | ||||
| COVID-19 symptoms | |||||||||
| Fever, n (%) | 5 (100) | 23 (96) | 3 (100) | 3 (75) | 0.9 | ||||
| Cough, n (%) | 2 (40) | 17 (71) | 0 (0) | 4 (100) | 0.19 | ||||
| Shortness of breath, n (%) | 1 (20) | 11 (46) | 1 (33) | 2 (50) | 0.29 | ||||
| Loss of taste or smell, n (%) | 4 (80) | 6 (25) | 2 (67) | 0 (0) | 0.02 | ||||
| Muscle or body pain, or headache | 0 (0) | 3 (13) | 0 (0) | 2 (67) | 0.4 | ||||
| Viral RNA Cp median (95% CI) | 31 (26–32) | 24 (19–27) | 27 (23–31) | 24 (19–30) | 0.049 (b) | ||||
| Number of cases, N | 5 (A) | 5 (B) | 24 (A) | 17 (B) | 3 (A) | 3 (B) | 4 (A) | 4 (B) | NR |
| Antibody levels mean OD450–620nm (SD) | |||||||||
| Anti-S-protein IgA | 3.7 (2) | 4 (1) | 2.8 (1.8) | 2.5 (1.75) | 3.4 (2.3) | 3 (1.8) | 1.4 (0.25) | 2.3 (1.7) | 0.37 (b) |
| Anti-S-protein IgG | 1.9 (1) | 2 (0.05) | 1.96 (1) | 2 (0.6) | 2.3 (0.2) | 1.9 (0.3) | 0.65 (0.4) | 1.7 (0.7) | 0.9 (b) |
| Anti-N-protein IgG | 1.8 (1) | 1.9 (0.6) | 1.6 (0.9) | 1.6 (0.7) | 1.3 (0.3) | 1.3 (0.4) | 0.9 (0.6) | 1.6 (1) | 0.73 (b) |
| Sample Nr | PANGO Lineage | Variant | a.a. Substitution, Sequenced Spike-Protein Fragment | Nucleotide Substitution without a.a. Substitution | GISAID Accession-ID |
|---|---|---|---|---|---|
| 09 | B.1.1.318 | T95I, del144, E484K, D614G, P681H | C21846T, del21994-21996, G23012A, T23287C, A23403G, C23604A | EPI_ISL_2956623 | |
| 16 | B.1.1.318 | T95I, del144, E484K, D614G, P681H | C21846T, del21994-21996, G23012A, T23287C, A23403G, C23604A | EPI_ISL_2956624 | |
| 28 | B.1.617.2 | Delta | G142D, E156G, del157/158, L452R, T478K, D614G, Q677H, P681R | G21987A, del22029-22034, T22917G, C22995A, A23403G, G23593T, C23604G | EPI_ISL_2956625 |
| 31 | B.1.525 | Eta | Q52R, A67V, del69/70, del144, E484K, D614G, Q677H, V687L | A21717G, C21762T, del21766-21771, del21994-21996, G23012A, A23403G, G23593C, G23621T | EPI_ISL_2956628 |
| 39 | B.1.617.2 | Delta | G142D, E156G, del157/158, L452R, T478K, D614G, P681R | G21987A, del22029-22034, T22917G, C22995A, C23029T, A23403G, C23604G | EPI_ISL_2956629 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, A.K.; Kaiser, M.; Molla, M.M.A.; Nafisa, T.; Yeasmin, M.; Ratul, R.H.; Sharif, M.M.; Akram, A.; Hosen, N.; Mamunur, R.; et al. Molecular and Serological Characterization of the SARS-CoV-2 Delta Variant in Bangladesh in 2021. Viruses 2021, 13, 2310. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112310
Ghosh AK, Kaiser M, Molla MMA, Nafisa T, Yeasmin M, Ratul RH, Sharif MM, Akram A, Hosen N, Mamunur R, et al. Molecular and Serological Characterization of the SARS-CoV-2 Delta Variant in Bangladesh in 2021. Viruses. 2021; 13(11):2310. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112310
Chicago/Turabian StyleGhosh, Asish Kumar, Marco Kaiser, Md. Maruf Ahmed Molla, Tasnim Nafisa, Mahmuda Yeasmin, Rifat Hossain Ratul, Md. Mohiuddin Sharif, Arifa Akram, Nur Hosen, Rashid Mamunur, and et al. 2021. "Molecular and Serological Characterization of the SARS-CoV-2 Delta Variant in Bangladesh in 2021" Viruses 13, no. 11: 2310. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112310