
Development of an Indirect ELISA Based on Spike Protein to Detect Antibodies against Feline Coronavirus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Serum Samples and Antibodies
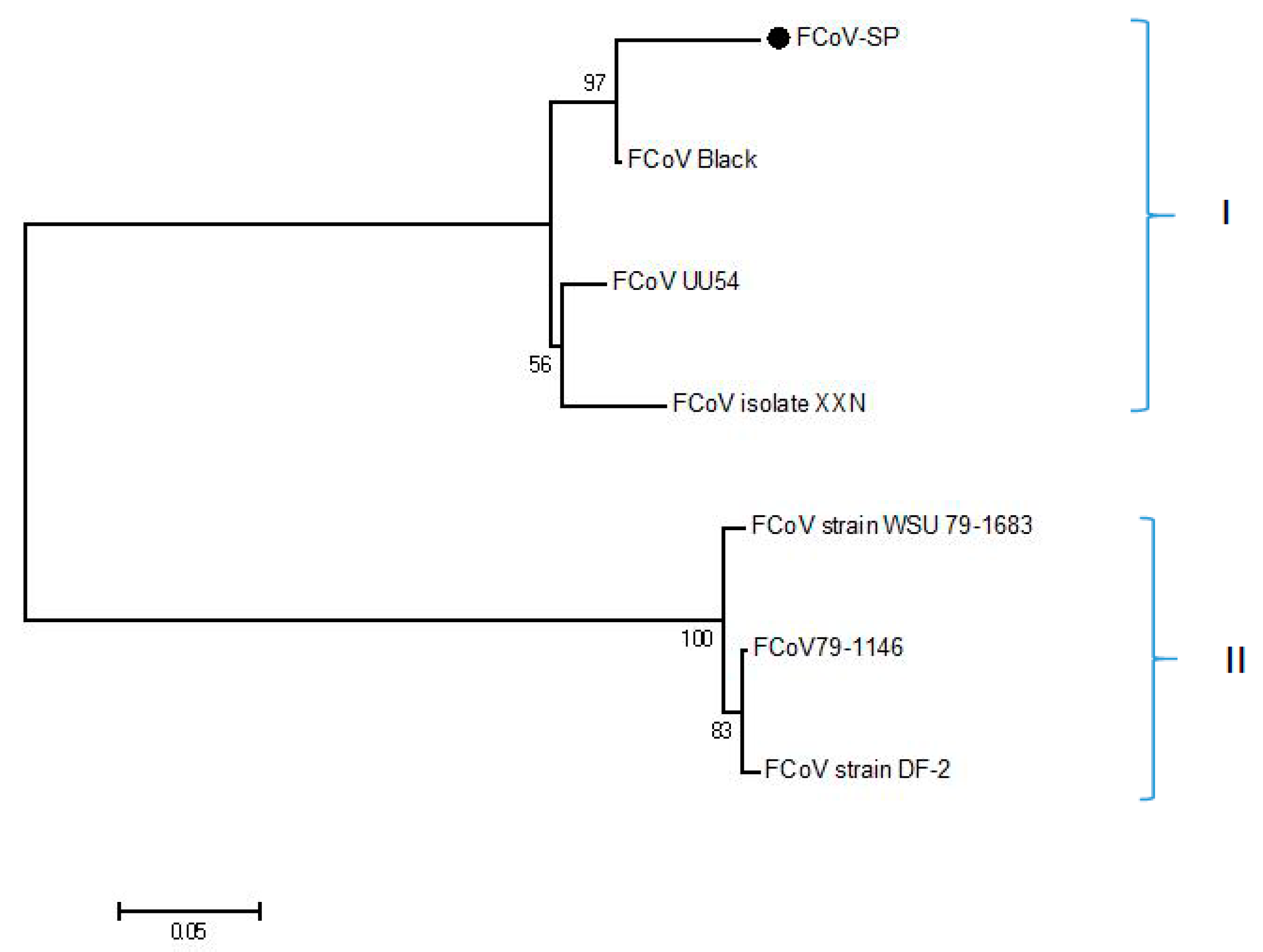
2.2. Antigen Selection and Vector Construction
2.3. Expression of Recombinant FCoV-SP Protein
2.4. Western Blotting of the FCoV-SP Protein
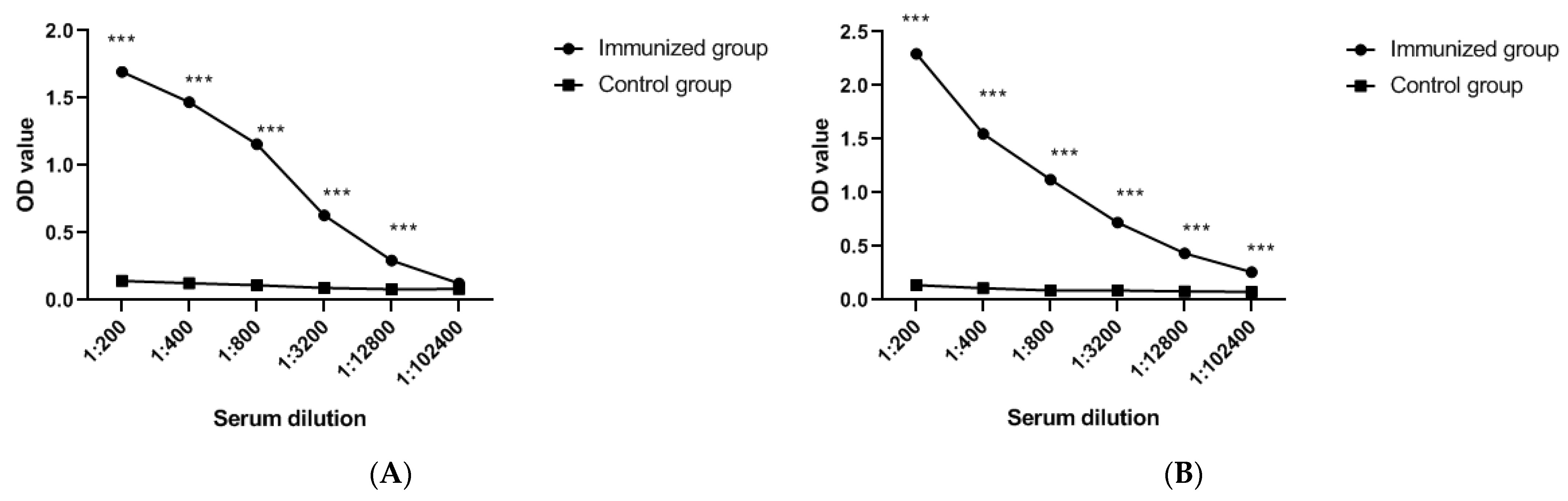
2.5. Immunogenicity Assessment
2.6. iELISA Procedures for Detecting Antibodies against FCoV
2.7. Determination of the Cut-Off Value
2.8. Check for the Coefficient of Variation (CV)
2.9. Specificity Test
2.10. Assessment of the Diagnostic Sensitivity and Specificity
2.11. Detection of FCoV in Field Samples
2.12. Statistical Analysis
3. Results
3.1. Production of the FCoV-SP Protein
3.2. Immunogenicity Evaluation of FCoV-SP Protein
3.3. Cut-Off Value of the iELISA
3.4. Repeatability of the iELISA
3.5. Specificity Assessment
3.6. Diagnostic Sensitivity and Specificity of iELISA
3.7. Antibody Identification in Field Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pedersen, N.C.; Allen, C.E.; Lyons, L. Pathogenesis of feline enteric coronavirus infection. J. Feline Med. Surg. 2008, 10, 529–541. [Google Scholar] [CrossRef] [Green Version]
- Pratelli, A. Comparison of Serologic Techniques for the Detection of Antibodies against Feline Coronaviruses. J. Vet. Diagn. Investig. 2008, 20, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Vennema, H.; Poland, A.; Foley, J.; Pedersen, N.C. Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 1998, 243, 150–157. [Google Scholar] [CrossRef] [Green Version]
- Olyslaegers, D.A.; Dedeurwaerder, A.; Desmarets, L.M.; Vermeulen, B.L.; Dewerchin, H.L.; Nauwynck, H.J. Altered ex-pression of adhesion molecules on peripheral blood leukocytes in feline infectious peritonitis. Vet. Microbiol. 2013, 166, 438–449. [Google Scholar] [CrossRef]
- Dye, C.; Siddell, S.G. Genomic RNA sequence of Feline coronavirus strain FIPV WSU-79/1146. J. Gen. Virol. 2005, 86, 2249–2253. [Google Scholar] [CrossRef]
- Porter, E.; Tasker, S.; Day, M.J.; Harley, R.; Kipar, A.; Siddell, S.G.; Helps, C.R. Amino acid changes in the spike protein of feline coronavirus correlate with systemic spread of virus from the intestine and not with feline infectious peritonitis. Vet. Res. 2014, 45, 1297–9716. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottier, P.J. Spike protein fusion peptide and feline coronavirus virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095. [Google Scholar] [CrossRef]
- Licitra, B.N.; Millet, J.K.; Regan, A.D.; Hamilton, B.S.; Rinaldi, V.; Duhamel, G.E.; Whittaker, G.R. Mutation in Spike Protein Cleavage Site and Pathogenesis of Feline Coronavirus. Emerg. Infect. Dis. 2013, 19, 1066–1073. [Google Scholar] [CrossRef]
- Sun, D.; Feng, L.; Shi, H.; Chen, J.; Cui, X.; Chen, H.; Liu, S.; Tong, Y.; Wang, Y.; Tong, G. Identification of two novel B cell epitopes on porcine epidemic diarrhea virus spike protein. Vet. Microbiol. 2008, 131, 73–81. [Google Scholar] [CrossRef]
- Jaimes, J.A.; Whittaker, G.R. Feline coronavirus: Insights into viral pathogenesis based on the spike protein structure and function. Virology 2018, 517, 108–121. [Google Scholar] [CrossRef]
- Lewis, C.S.; Porter, E.; Matthews, D.; Kipar, A.; Tasker, S.; Helps, C.R.; Siddell, S.G. Genotyping coronaviruses associated with feline infectious peritonitis. J. Gen. Virol. 2015, 96, 1358–1368. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, G.R.; André, N.M.; Millet, J.K. Improving Virus Taxonomy by Recontextualizing Sequence-Based Classification with Biologically Relevant Data: The Case of the Alphacoronavirus 1 Species. mSphere 2018, 3, e00463-17. [Google Scholar] [CrossRef] [Green Version]
- Benetka, V.; Kübber-Heiss, A.; Kolodziejek, J.; Nowotny, N.; Hofmann-Parisot, M.; Möstl, K. Prevalence of feline corona-virus types I and II in cats with histopathologically verified feline infectious peritonitis. Vet. Microbiol. 2004, 99, 31–42. [Google Scholar] [CrossRef]
- Kummrow, M.; Meli, M.L.; Haessig, M.; Goenczi, E.; Poland, A.; Pedersen, N.C.; Hofmann-Lehmann, R.; Lutz, H. Feline coronavirus serotypes 1 and 2: Seroprevalence and association with disease in Switzerland. Clin. Diagn. Lab. Immunol. 2005, 12, 1209–1215. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Li, H.; Han, M.; Jia, S.; Feng, B.; Gao, X.; Wang, Z.; Jiang, Y.; Cui, W.; Wang, L.; et al. Epidemiological investigation of feline infectious peritonitis in cats living in Harbin, Northeast China from 2017 to 2019 using a combination of an EvaGreen-based real-time RT-PCR and serum chemistry assays. Mol. Cell Probes 2020, 49, 14. [Google Scholar] [CrossRef]
- Li, C.; Liu, Q.; Kong, F.; Guo, D.; Zhai, J.; Su, M.; Sun, D. Circulation and genetic diversity of Feline coronavirus type I and II from clinically healthy and FIP-suspected cats in China. Transbound. Emerg. Dis. 2019, 66, 763–775. [Google Scholar] [CrossRef] [Green Version]
- Addie, D.D.; Jarrett, O. Control of feline coronavirus infection in kittens. Vet. Rec. 1990, 126, 164. [Google Scholar]
- Addie, D.D.; Jarrett, O. A study of naturally occurring feline coronavirus infections in kittens. Vet. Rec. 1992, 130, 133–137. [Google Scholar] [CrossRef]
- Addie, D.D.; Jarrett, O. Use of a reverse-transcriptase polymerase chain reaction for monitoring the shedding of feline coronavirus by healthy cats. Vet. Rec. 2001, 148, 649–653. [Google Scholar] [CrossRef]
- Sparkes, A.H.; Gruffydd-Jones, T.J.; Harbour, D.A. An appraisal of the value of laboratory tests in the diagnosis of feline infectious peritonitis. Am. Anim. Hosp. Assoc. 1995, 30, 345–350. [Google Scholar]
- Okda, F.; Liu, X.; Singrey, A.; Clement, T.; Nelson, J.; Christopher-Hennings, J.; Nelson, E.A.; Lawson, S. Development of an indirect ELISA, blocking ELISA, fluorescent microsphere immunoassay and fluorescent focus neutralization assay for serologic evaluation of exposure to North American strains of Porcine Epidemic Diarrhea Virus. BMC Vet. Res. 2015, 11, 180. [Google Scholar] [CrossRef] [Green Version]
- Kit, S.; Otsuka, H.; Kit, M. Expression of porcine pseudorabies virus genes by a bovine herpesvirus-1 (infectious bovine rhinotracheitis virus) vector. Arch. Virol. 1992, 124, 1–20. [Google Scholar] [CrossRef]
- Hofmann, M.; Wyler, R. Enzyme-linked immunosorbent assay for the detection of porcine epidemic diarrhea coronavirus antibodies in swine sera. Vet. Microbiol. 1990, 21, 263–273. [Google Scholar] [CrossRef]
- White, J.M.; Delos, S.E.; Brecher, M.; Schornberg, K. Structures and Mechanisms of Viral Membrane Fusion Proteins: Multiple Variations on a Common Theme. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 189–219. [Google Scholar] [CrossRef] [Green Version]
- Knuchel, M.; Ackermann, M.; Müller, H.K.; Kihm, U. An ELISA for detection of antibodies against porcine epidemic diarrhoea virus (PEDV) based on the specific solubility of the viral surface glycoprotein. Vet. Microbiol. 1992, 32, 117–134. [Google Scholar] [CrossRef]
- Felten, S.; Hartmann, K. Diagnosis of Feline Infectious Peritonitis: A Review of the Current Literature. Viruses 2019, 11, 1068. [Google Scholar] [CrossRef] [Green Version]
- DeSilva, B.; Smith, W.; Weiner, R.; Kelley, M.; Smolec, J.; Lee, B.; Khan, M.; Tacey, R.; Hill, H.; Celniker, A. Recommendations for the bioanalytical method validation of ligand-binding assays to support pharmacokinetic assessments of macro-molecules. Pharm. Res. 2003, 20, 1885–1900. [Google Scholar] [CrossRef]
- Tvarijonaviciute, A.; Martínez-Subiela, S.; Ceron, J.J. Validation of two ELISA assays for total ghrelin measurement in dogs. J. Anim. Physiol. Anim. Nutr. 2012, 96, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, X.-X.; Li, C.; Wang, H.; Wang, H.; Meng, X.-Z.; Ma, J.; Ni, H.-B.; Zhang, X.; Qi, Y.; et al. Epidemiology of porcine epidemic diarrhea virus among Chinese pig populations: A meta-analysis. Microb. Pathog. 2019, 129, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zhang, D.; Zhou, J.; Xia, Z.; Lin, D.; Lou, Y.; Tan, W.; Qin, K. Prevalence and phylogenetic characterization of canine coronavirus from diseased pet dogs in Beijing, China. Sci. China Life Sci. 2016, 59, 860–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | X | SD | CV% | |
|---|---|---|---|---|
| Intra-assay | 1 | 2.03 | 0.03 | 1.63% |
| 2 | 2.26 | 0.03 | 1.15% | |
| 3 | 1.99 | 0.05 | 2.40% | |
| 4 | 0.14 | 0.01 | 4.13% | |
| 5 | 0.19 | 0.01 | 5.04% | |
| Inter-assay | 1 | 2.31 | 0.22 | 9.28% |
| 2 | 2.21 | 0.14 | 6.09% | |
| 3 | 2.07 | 0.09 | 4.28% | |
| 4 | 0.16 | 0.02 | 15.13% | |
| 5 | 0.21 | 0.01 | 6.80% |
| iELISA | Result of Western Blot | Total | |
|---|---|---|---|
| Positive | Negative | ||
| Positive | 30 | 2 | 32 |
| Negative | 0 | 28 | 28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, B.; Zhang, G.; Zhang, X.; Chen, X.; Zhang, M.; Li, L.; Lin, W. Development of an Indirect ELISA Based on Spike Protein to Detect Antibodies against Feline Coronavirus. Viruses 2021, 13, 2496. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122496
Dong B, Zhang G, Zhang X, Chen X, Zhang M, Li L, Lin W. Development of an Indirect ELISA Based on Spike Protein to Detect Antibodies against Feline Coronavirus. Viruses. 2021; 13(12):2496. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122496
Chicago/Turabian StyleDong, Bo, Gaoqiang Zhang, Xiaodong Zhang, Xufei Chen, Meiling Zhang, Linglin Li, and Weiming Lin. 2021. "Development of an Indirect ELISA Based on Spike Protein to Detect Antibodies against Feline Coronavirus" Viruses 13, no. 12: 2496. https://0-doi-org.brum.beds.ac.uk/10.3390/v13122496