
Construction of Baculovirus-Inducible CRISPR/Cas9 Antiviral System Targeting BmNPV in Bombyx mori


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silkworm Strains and Virus Stock
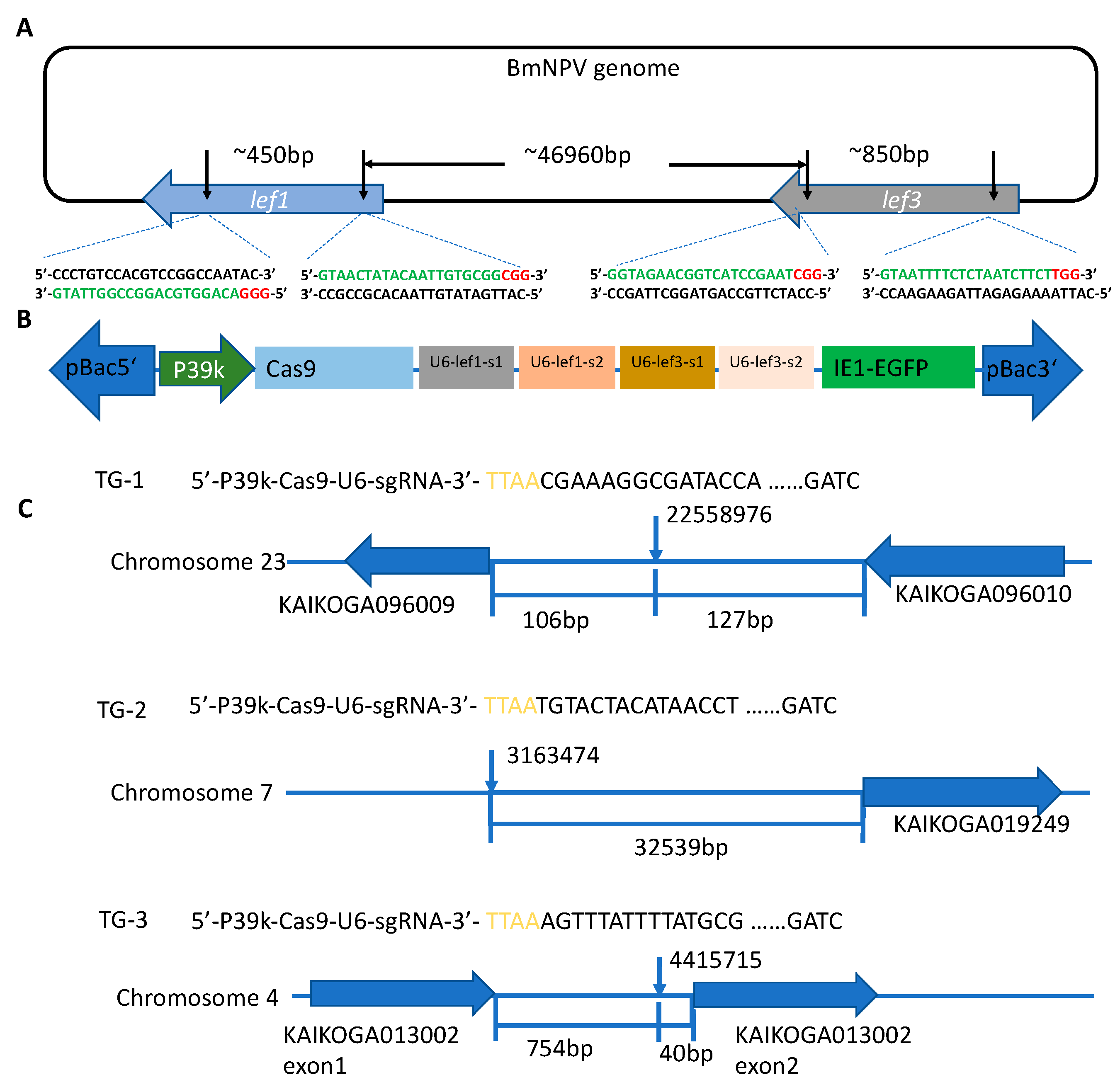
2.2. Target Gene Selection and Vector Construction
2.3. Silkworm Genetic Transformation
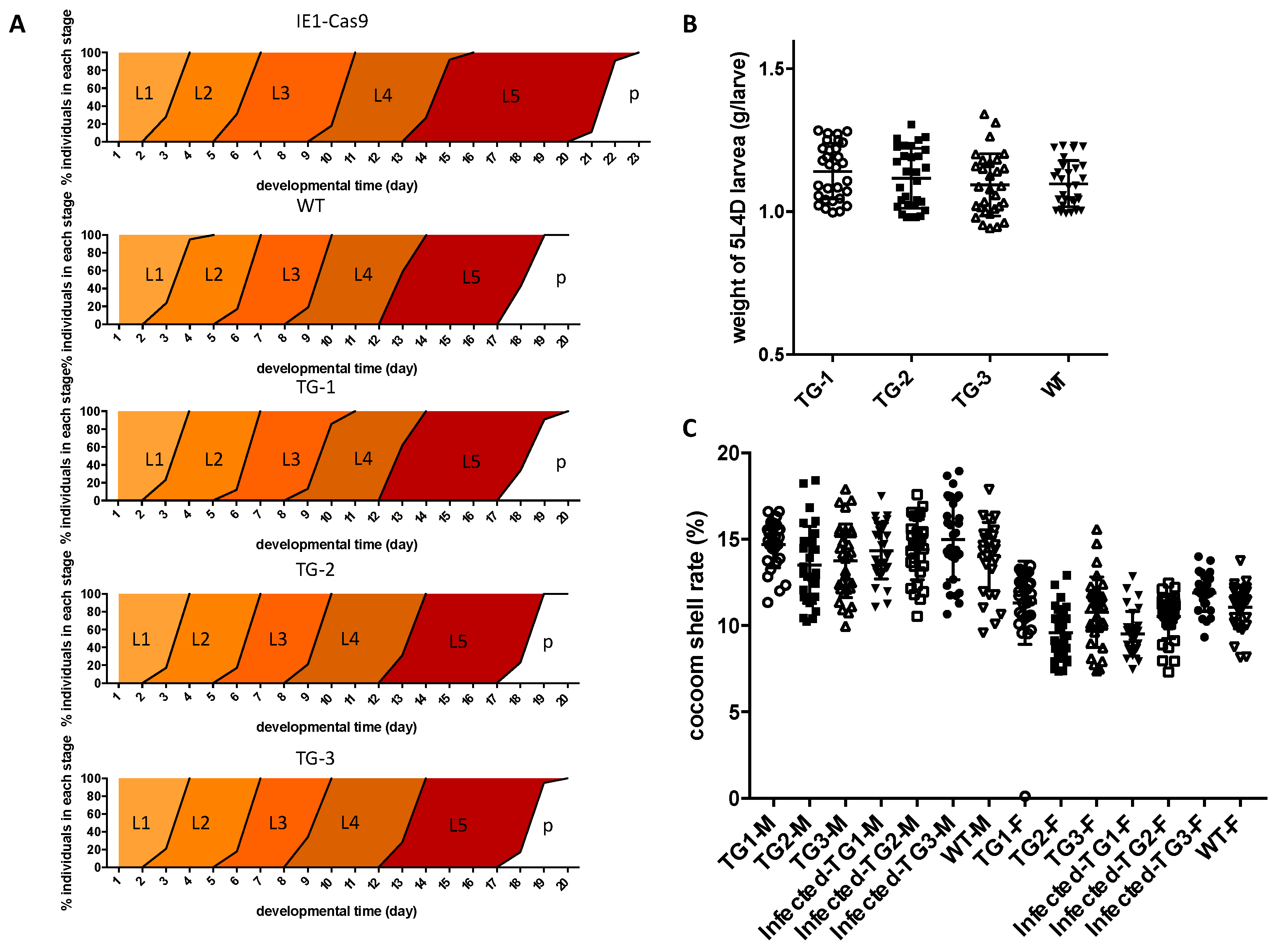
2.4. Analysis of Growth and Economic Characteristics
2.5. Viral Inoculation and Mortality Analyses
2.6. RNA Isolation and cDNA Synthesis
2.7. Time-Course qPCR Analysis of BmNPV DNA Copies after Oral Infection
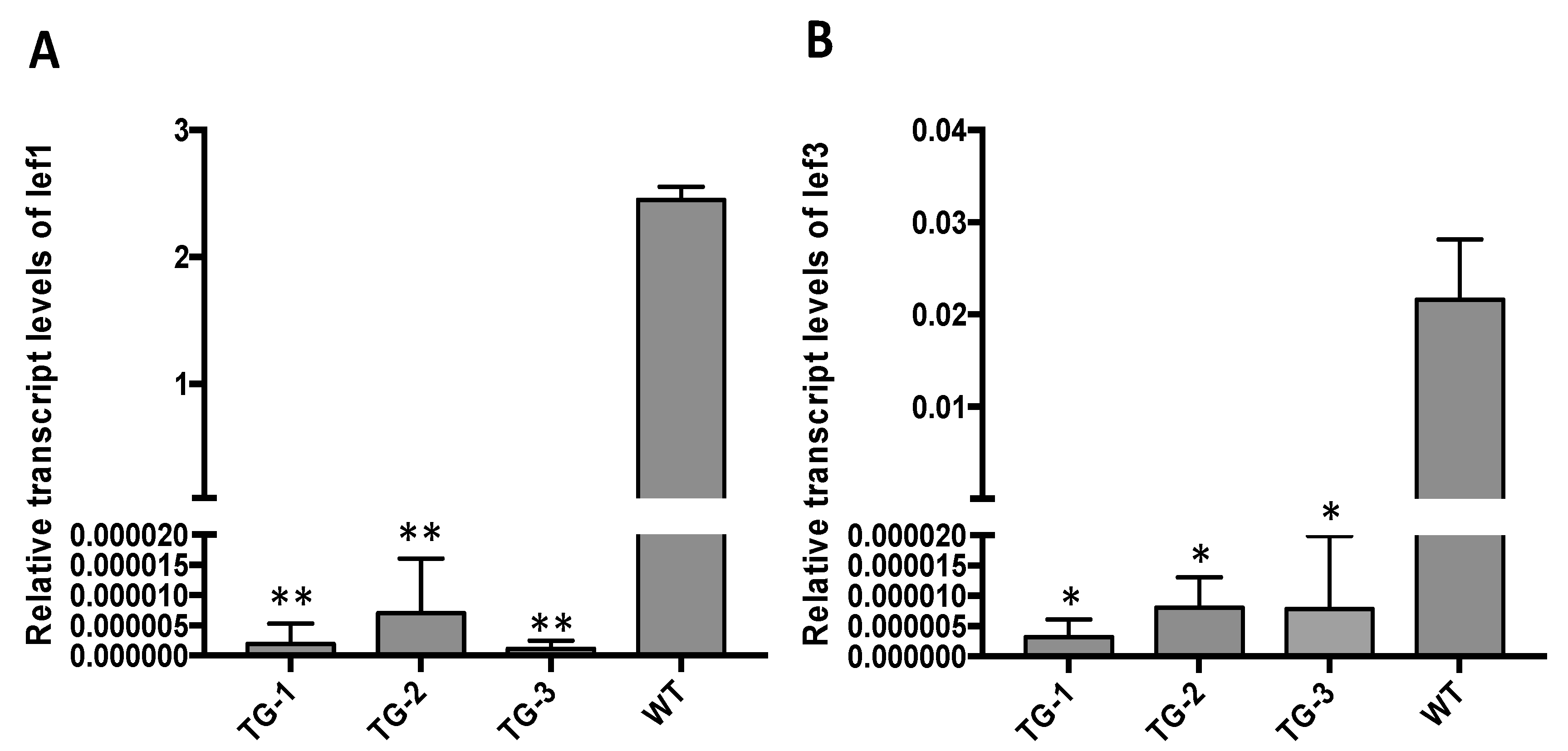
2.8. Quantification of BmNPV Gene Expression
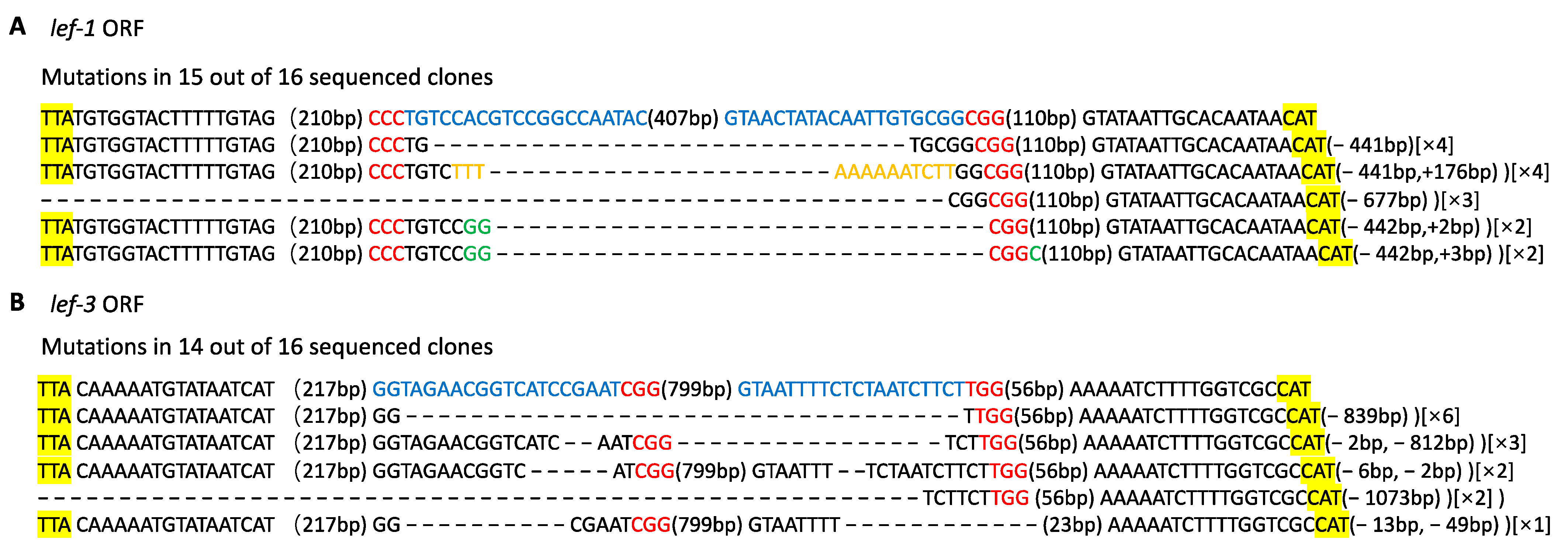
2.9. Mutagenesis Analysis of the Viral Target Genes
2.10. Statistical Analysis
3. Results
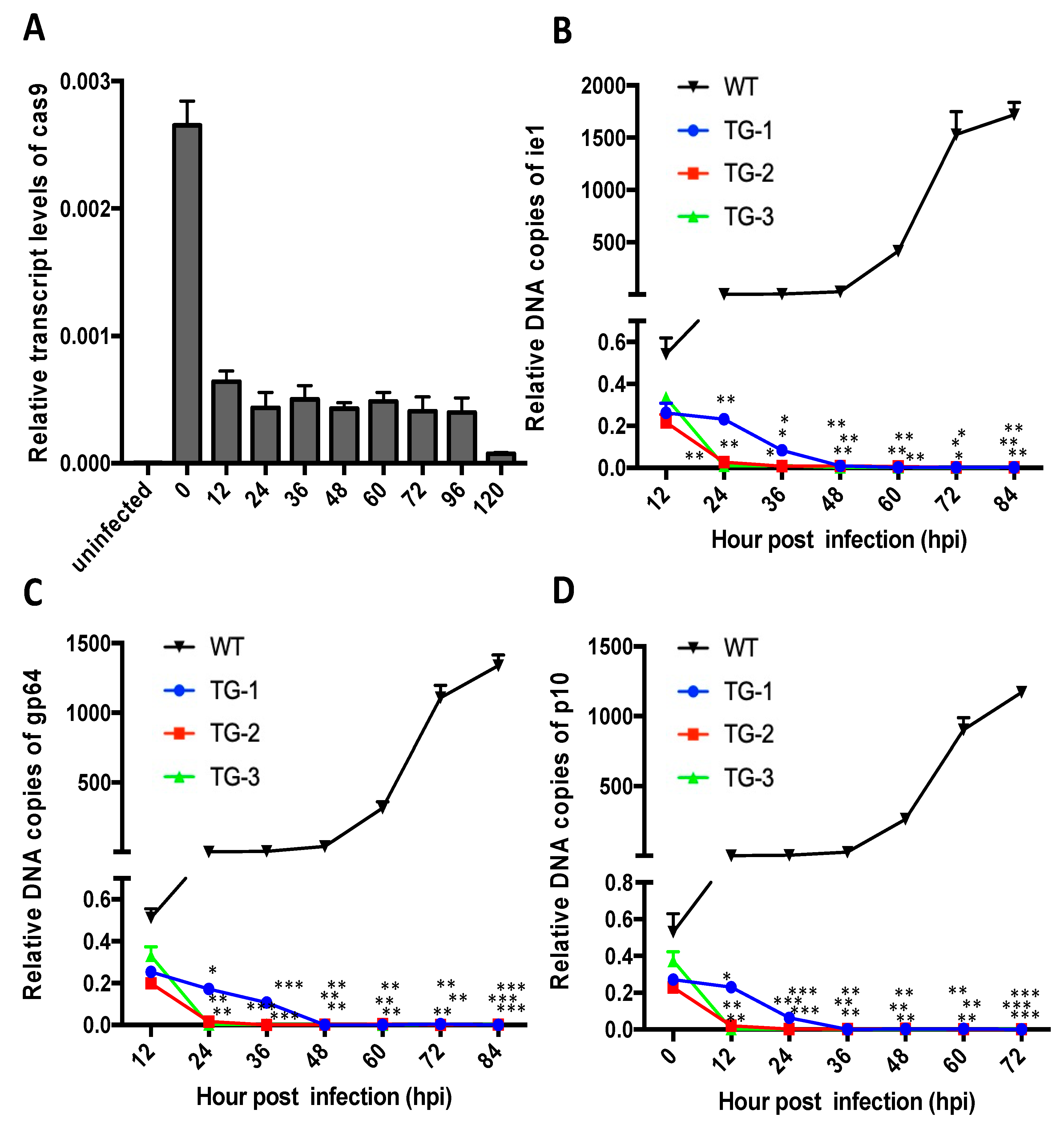
3.1. Construction of Baculovirus-Inducible CRISPR/Cas9 Gene Editing System
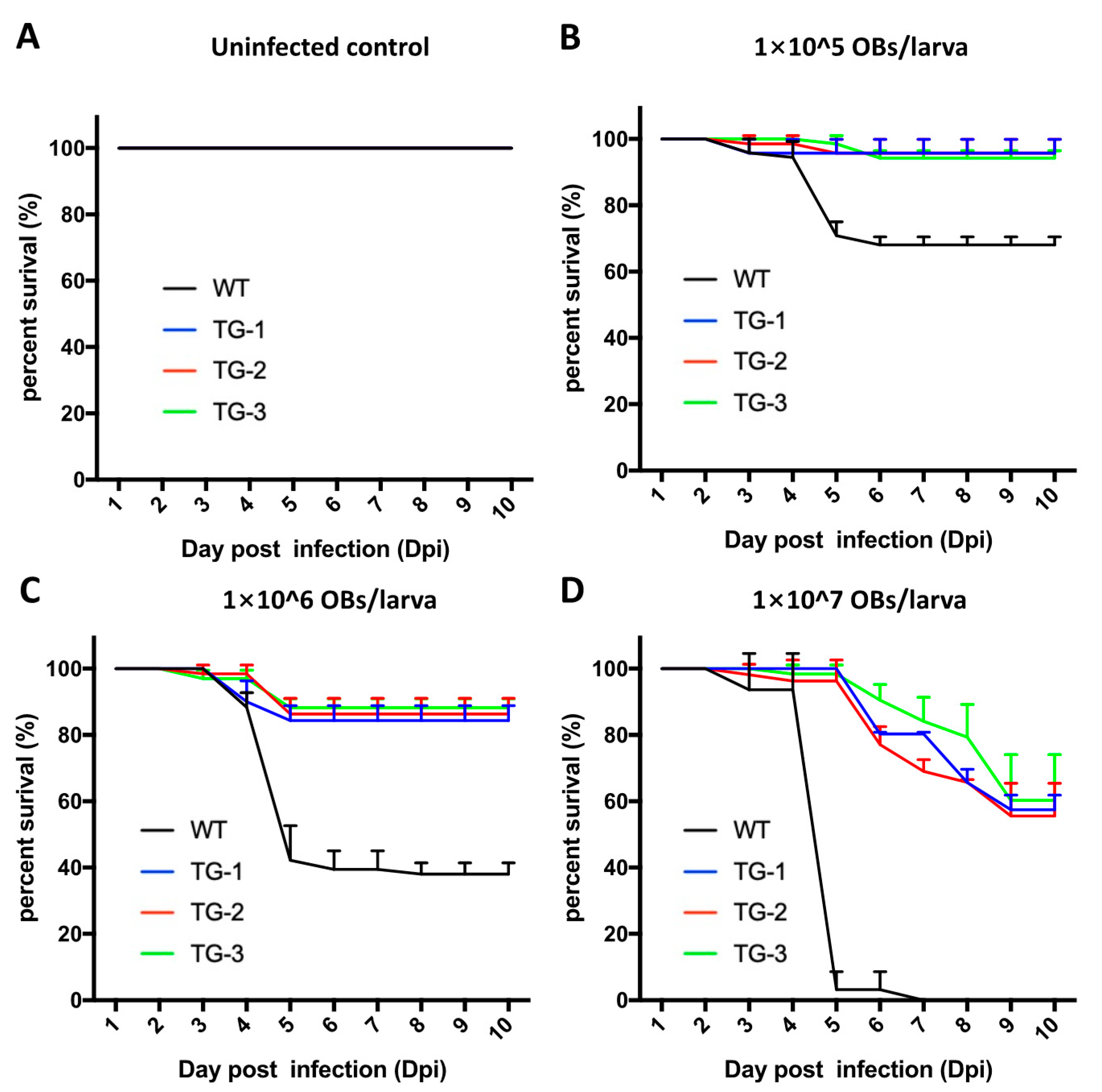
3.2. TG Silkworms Exhibit Higher Resistance to BmNPV Infection Than the Wild-Type Silkworms
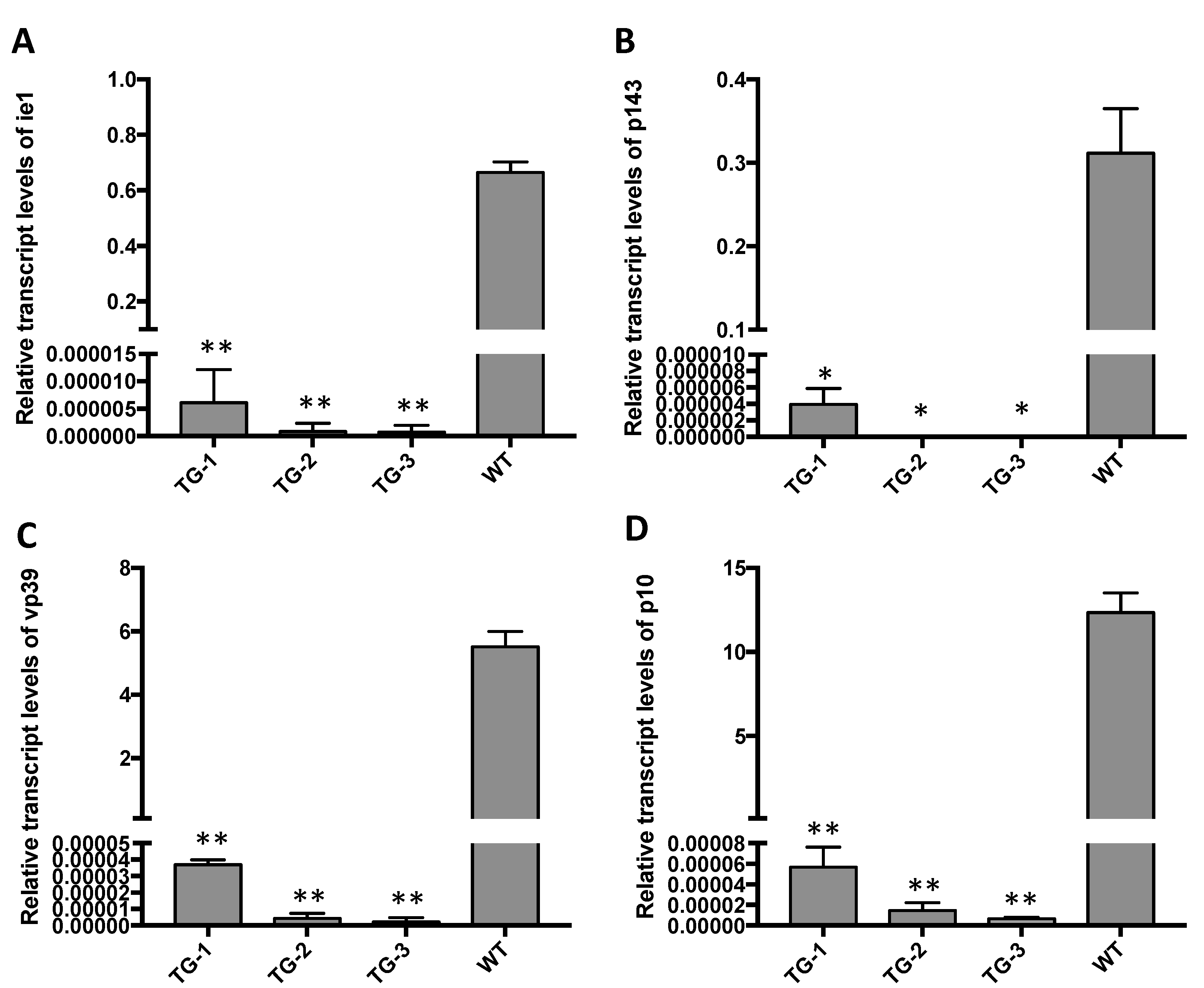
3.3. CRISPR/Cas9-Mediated BmNPV Genome Editing in Transgenic Silkworm
3.4. The Antivirus Efficiency of the Baculovirus-Inducing and Targeting CRISPR/Cas9 System Is High
3.5. Economically Important Characteristics Are Normal in the Transgenic Silkworm
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Annu. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef]
- Jiang, L.; Xia, Q. The progress and future of enhancing antiviral capacity by transgenic technology in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2014, 48, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, A.; Maeda, S. Host range expansion by recombination of the baculoviruses Bombyx mori nuclear polyhedrosis virus and Autographa californica nuclear polyhedrosis virus. J. Virol. 1991, 65, 3625–3632. [Google Scholar] [CrossRef] [Green Version]
- Blissard, G.W.; Theilmann, D.A. Baculovirus Entry and Egress from Insect Cells. Annu. Rev. Virol. 2018, 5, 113–139. [Google Scholar] [CrossRef]
- Kool, M.; Ahrens, C.H.; Goldbach, R.W.; Rohrmann, G.F.; Vlak, J.M. Identification of genes involved in DNA replication of the Autographa californica baculovirus. Proc. Natl. Acad. Sci. USA 1994, 91, 11212–11216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kool, M.; Ahrens, C.H.; Vlak, J.M.; Rohrmann, G.F. Replication of baculovirus DNA. J. Gen. Virol. 1995, 76, 2103–2118. [Google Scholar] [CrossRef] [PubMed]
- Vanarsdall, A.; Mikhailov, V.; Rohrmann, G. Baculovirus DNA Replication and Processing. Curr. Drug Targets 2007, 8, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, G. Baculovirus Molecular Biology; Bethesda: Rockville, MD, USA, 2019; pp. 1–2. [Google Scholar]
- Jiang, L.; Zhao, P.; Cheng, T.; Sun, Q.; Peng, Z.; Dang, Y.; Wu, X.; Wang, G.; Jin, S.; Lin, P.; et al. A transgenic animal with antiviral properties that might inhibit multiple stages of infection. Antivir. Res. 2013, 98, 171–173. [Google Scholar] [CrossRef]
- Ponnuvel, K.M.; Nakazawa, H.; Furukawa, S.; Asaoka, A.; Ishibashi, J.; Tanaka, H.; Yamakawa, M. A Lipase Isolated from the Silkworm Bombyx mori Shows Antiviral Activity against Nucleopolyhedrovirus. J. Virol. 2003, 77, 10725–10729. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, G.; Cheng, T.; Yang, Q.; Jin, S.; Lu, G.; Wu, F.; Xiao, Y.; Xu, H.; Xia, Q. Resistance to Bombyx mori nucleopolyhedrovirus via overexpression of an endogenous antiviral gene in transgenic silkworms. Arch. Virol. 2012, 157, 1323–1328. [Google Scholar] [CrossRef]
- Yao, H.; He, F.; Guo, A.; Cao, C.; Lu, X.; Wu, X. Gene analysis of an antiviral protein SP-2 from Chinese wild silkworm, Bombyx mandarina Moore and its bioactivity assay. Sci. China Ser. C Life Sci. 2008, 51, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-H.; Dong, X.-L.; Pan, C.-X.; Du, G.-Y.; Wu, Y.-F.; Yang, J.-G.; Chen, P.; Lu, C.; Pan, M.-H. A newly discovered member of the Atlastin family, BmAtlastin-n, has an antiviral effect against BmNPV in Bombyx mori. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isobe, R.; Kojima, K.; Matsuyama, T.; Quan, G.-X.; Kanda, T.; Tamura, T.; Sahara, K.; Asano, S.-I.; Bando, H. Use of RNAi technology to confer enhanced resistance to BmNPV on transgenic silkworms. Arch. Virol. 2004, 149, 1931–1940. [Google Scholar] [CrossRef]
- Zhang, J.; He, Q.; Zhang, C.-D.; Chen, X.-Y.; Chen, X.-M.; Dong, Z.-Q.; Li, N.; Kuang, X.-X.; Cao, M.-Y.; Lu, C.; et al. Inhibition of BmNPV replication in silkworm cells using inducible and regulated artificial microRNA precursors targeting the essential viral gene lef-11. Antivir. Res. 2014, 104, 143–152. [Google Scholar] [CrossRef]
- Kanginakudru, S.; Royer, C.; Edupalli, S.V.; Jalabert, A.; Mauchamp, B.; Prasad, S.V.; Chavancy, G.; Couble, P.; Nagaraju, J. Targeting ie-1 gene by RNAi induces baculoviral resistance in lepidopteran cell lines and in transgenic silkworms. Insect Mol. Biol. 2007, 16, 635–644. [Google Scholar] [CrossRef]
- Subbaiah, E.V.; Royer, C.; Kanginakudru, S.; Satyavathi, V.V.; Babu, A.S.; Sivaprasad, V.; Chavancy, G.; Darocha, M.; Jalabert, A.; Mauchamp, B.; et al. Engineering silkworms for resistance to baculovirus through multigene RNA interference. Genetics 2013, 193, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, J.; Lu, Y.; Hu, Y.; Xue, R.; Cao, G.; Gong, C. Resistance of transgenic silkworm to BmNPV could be improved by silencing ie-1 and lef-1 genes. Gene Ther. 2014, 21, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.-K.; Gu, Y.; Diaz, A.; Marlett, J.; Takahashi, Y.; Li, M.; Suzuki, K.; Xu, R.; Hishida, T.; Chang, C.-J.; et al. Use of the CRISPR/Cas9 system as an intracellular defense against HIV-1 infection in human cells. Nat. Commun. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebina, H.; Misawa, N.; Kanemura, Y.; Koyanagi, Y. Harnessing the CRISPR/Cas9 system to disrupt latent HIV-1 provirus. Sci. Rep. 2013, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zhen, S.; Lu, J.; Liu, Y.-H.; Chen, W.; Li, X. Synergistic antitumor effect on cervical cancer by rational combination of PD1 blockade and CRISPR-Cas9-mediated HPV knockout. Cancer Gene Ther. 2020, 27, 168–178. [Google Scholar] [CrossRef]
- Lin, S.-R.; Yang, H.-C.; Kuo, Y.-T.; Liu, C.-J.; Yang, T.-Y.; Sung, K.-C.; Lin, Y.-Y.; Wang, H.-Y.; Wang, C.-C.; Shen, Y.-C.; et al. The CRISPR/Cas9 system facilitates clearance of the intrahepatic HBV templates in vivo. Mol. Ther. Nucleic Acids 2014, 3, e186. [Google Scholar] [CrossRef]
- Yuen, K.-S.; Chan, C.-P.; Wong, N.-H.M.; Ho, C.-H.; Ho, T.-H.; Lei, T.; Deng, W.; Tsao, S.W.; Chen, H.; Kok, K.-H.; et al. CRISPR/Cas9-mediated genome editing of Epstein–Barr virus in human cells. J. Gen. Virol. 2015, 96, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Zaynab, M.; Sharif, Y.; Fatima, M.; Afzal, M.Z.; Aslam, M.M.; Raza, M.F.; Anwar, M.; Raza, M.A.; Sajjad, N.; Yang, X.; et al. CRISPR/Cas9 to generate plant immunity against pathogen. Microb. Pathog. 2020, 141, 103996. [Google Scholar] [CrossRef]
- Chen, S.; Hou, C.; Bi, H.; Wang, Y.; Xu, J.; Li, M.; James, A.A.; Huang, Y.; Tan, A. Transgenic Clustered Regularly Interspaced Short Palindromic Repeat/Cas9-Mediated Viral Gene Targeting for Antiviral Therapy of Bombyx mori Nucleopolyhedrovirus. J. Virol. 2017, 91, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Zhang, X.; Liu, Y.; Yang, D.; Liu, Z.; Chen, K.; Tang, L.; Wang, M.; Hu, Z.; Zhang, S.; et al. Transgenic genome editing-derived antiviral therapy to nucleopolyhedrovirus infection in the industrial strain of the silkworm. Insect Biochem. Mol. Biol. 2021, 139, 103672. [Google Scholar] [CrossRef]
- Dong, Z.; Dong, F.; Yu, X.; Huang, L.; Jiang, Y.; Hu, Z.; Chen, P.; Lu, C.; Pan, M. Excision of nucleopolyhedrovirus form transgenic silkworm using the CRISPR/Cas9 system. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Dong, Z.; Huang, L.; Dong, F.; Hu, Z.; Qin, Q.; Long, J.; Cao, M. Establishment of a baculovirus-inducible CRISPR / Cas9 system for antiviral research in transgenic silkworms. Appl. Microbiol. Biotechnol. 2018, 102, 9255–9265. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Hu, Z.; Qin, Q.; Dong, F.; Huang, L.; Long, J.; Chen, P.; Lu, C.; Pan, M. CRISPR/Cas9-mediated disruption of the immediate early-0 and 2 as a therapeutic approach to Bombyx mori nucleopolyhedrovirus in transgenic silkworm. Insect Mol. Biol. 2019, 28, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Qin, Q.; Hu, Z.; Chen, P.; Huang, L.; Zhang, X.; Tian, T.; Lu, C.; Pan, M. Construction of a One-Vector Multiplex CRISPR/Cas9 Editing System to Inhibit Nucleopolyhedrovirus Replication in Silkworms. Virol. Sin. 2019, 34, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef]
- Kim, D.; Bae, S.; Park, J.; Kim, E.; Kim, S.; Yu, H.R.; Hwang, J.; Kim, J.-I.; Kim, J.-S. Digenome-seq: Genome-wide profiling of CRISPR-Cas9 off-target effects in human cells. Nat. Methods 2015, 12, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Kramer, N.J.; Haney, M.S.; Morgens, D.W.; Jovičić, A.; Couthouis, J.; Li, A.; Ousey, J.; Ma, R.; Bieri, G.; Tsui, C.K.; et al. CRISPR-Cas9 screens in human cells and primary neurons identify modifiers of C9ORF72 dipeptide-repeat-protein toxicity. Nat. Genet. 2018, 50, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Kool, M.; Vlak, J.M. The structural and functional organization of the autographa californica nuclear polyhedrosis virus genome. Arch. Virol. 1993, 130, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.-Y.; Kuang, X.-X.; Li, H.-Q.; Lei, X.-I.; Xiao, W.-F.; Dong, Z.-Q.; Zhang, J.; Hu, N.; Chen, T.-T.; Lu, C.; et al. Screening and optimization of an efficient Bombyx mori nucleopolyhedrovirus inducible promoter. J. Biotechnol. 2016, 231, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.D.; Guarino, L.A.; Summers, M.D. Functional mapping of an AcNPV immediately early gene which augments expression of the IE-1 trans-activated 39K gene. Virology 1988, 162, 444–451. [Google Scholar] [CrossRef]
- Tan, A.; Fu, G.; Jin, L.; Guo, Q.; Li, Z.; Niu, B.; Meng, Z.; Morrison, N.I.; Alphey, L.; Huang, Y. Transgene-based, female-specific lethality system for genetic sexing of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2013, 110, 6766–6770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, Z.; Xu, J.; Zeng, B.; Ling, L.; You, L.; Chen, Y.; Huang, Y.; Tan, A. The CRISPR/Cas System mediates efficient genome engineering in Bombyx mori. Cell Res. 2013, 23, 1414–1416. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.-Q.; Hu, Z.-G.; Li, H.-Q.; Jiang, Y.-M.; Cao, M.-Y.; Chen, P.; Lu, C.; Pan, M.-H. Construction and characterization of a synthetic Baculovirus-inducible 39K promoter. J. Biol. Eng. 2018, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Goldsmith, M.R.; Xia, Q. Advances in the Arms Race Between Silkworm and Baculovirus. Front. Immunol. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Huang, L.; Dong, Z.Q.; Dong, F.F.; Yu, X.B.; Hu, Z.G.; Liao, N.C.; Chen, P.; Lu, C.; Pan, M.-H. Gene editing the BmNPV inhibitor of apoptosis protein 2 (iap2) as an antiviral strategy in transgenic silkworm. Int. J. Biol. Macromol. 2021, 166, 529–537. [Google Scholar] [CrossRef]
- Dong, Z.; Qin, Q.; Hu, Z.; Zhang, X.; Miao, J.; Huang, L.; Chen, P.; Lu, C.; Pan, M. CRISPR/Cas12a Mediated Genome Editing Enhances Bombyx mori Resistance to BmNPV. Front. Bioeng. Biotechnol. 2020, 8, 1–11. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR-Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freije, C.A.; Myhrvold, C.; Boehm, C.K.; Lin, A.E.; Welch, N.L.; Carter, A.; Metsky, H.C.; Luo, C.Y.; Abudayyeh, O.O.; Gootenberg, J.S.; et al. Programmable Inhibition and Detection of RNA Viruses Using Cas13. Mol. Cell 2019, 76, 826–837.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wang, S.; Dong, X.; Li, Q.; Li, M.; Li, J.; Guo, Y.; Jin, X.; Zhou, Y.; Song, H.; et al. CRISPR-Cas13a Cleavage of Dengue Virus NS3 Gene Efficiently Inhibits Viral Replication. Mol. Ther. Nucleic Acids 2020, 19, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Meng, X.; Xue, R.; Zhu, Y.; Zhang, X.; Pan, Z.; Zheng, X.; Gong, C. Characterization of the complete genome segments from BmCPV-SZ, a novel Bombyx mori cypovirus 1 isolate. Can. J. Microbiol. 2012, 58, 872–883. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Primer Purpose |
|---|---|---|
| P39k-F | AAGGCTGTCCTGCTGTGTGC | Plasmid construction |
| P39k-R | CTGGCAATTCGTTTGTGATG | Plasmid construction |
| P39k-cas9-F | ACTCACTATAGGGCGAATTGGGTACCGTCGACAAGGCTG TCCTGCTGTGTGC | Plasmid construction |
| P39k-cas9-R | ATGGAGTACTTCTTGTCCATGACGTCCTGGCAATTCGTTTGTGATG | Plasmid construction |
| lef-1-sg1-F | GTAACTATACAATTGTGCGGGTTTTAGAGCTAGAAATAGC | Plasmid construction |
| lef-1-sg1-R | CCGCACAATTGTATAGTTACACTTGTAGAGCACGATATTT | Plasmid construction |
| lef-1-sg2-F | GTATTGGCCGGACGTGGACAGTTTTAGAGCTAGAAATAGC | Plasmid construction |
| lef-1-sg2-R | TGTCCACGTCCGGCCAATACACTTGTAGAGCACGATATTT | Plasmid construction |
| lef-3-sg1-F | GTAATTTTCTCTAATCTTCTGTTTTAGAGCTAGAAATAGC | Plasmid construction |
| lef-3-g1-R | AGAAGATTAGAGAAAATTACACTTGTAGAGCACGATATTT | Plasmid construction |
| lef-3-g2-F | GGTAGAACGGTCATCCGAATGTTTTAGAGCTAGAAATAGC | Plasmid construction |
| lef-3-g2-R | ATTCGGATGACCGTTCTACCACTTGTAGAGCACGATATTT | Plasmid construction |
| U6-F1 | ACTCACTATAGGGCGAATTGGGTACCAGGTTATGTAGTACACATTG | Plasmid construction |
| Overlap-gBone-R | CCGCGGAGTCAATGGCTAGCAAAAAAAGCACCGACTCGGTG | Plasmid construction |
| Overlap-U6-F | GCTAGCCATTGACTCCGCGGAGGTTATGTAGTACACATTGTTGTA | Plasmid construction |
| gBone-HindIII-R | TTTTCTTGTTATAGATATCAAAAAAAAGCACCGACTCGGTG | Plasmid construction |
| ie1-qPCR-F | GCTCAAGACCACTGATAATCTC | qPCR |
| ie1-qPCR-R | AATCGTCCAAGTATTCGTCCA | qPCR |
| gp64-qPCR-F | CCTTCAGCCATGGAAGTGAT | qPCR |
| gp64-qPCR-R | GACGACGTCGAATTTTGGAT | qPCR |
| p10-qPCR-F | CCATTGCGGAAACTAACACA | qPCR |
| p10-qPCR-R | AGCAGTGTCACCGGTCAATA | qPCR |
| Bmrp49-qPCR-F | AAACATACAAGATGGCTATAAGACCTG | qPCR |
| Bmrp49-qPCR-R | TTTATAAATGACATGTGAACATACCTC | qPCR |
| Inverse PCR-F1 | CAGTGACACTTACCGCATTGA | Inverse PCR |
| Inverse PCR-R1 | CATTTTGACTCACGCGGTC | Inverse PCR |
| Inverse PCR-F2 | CGCTATTTAGAAAGAGAGAGCAA | Inverse PCR |
| Inverse PCR-R2 | ATCACGTAAGTAGAACATGAAATAACA | Inverse PCR |
| ie1-qRT-F | GCTCAAGACCACTGATAATCTC | qPCR |
| ie1-qRT-R | AATCGTCCAAGTATTCGTCCA | qPCR |
| p143-qRT-F | TGGCTTCATACTTTAGCAACC | qPCR |
| p143-qRT-R | GTTTGACGATGACAACCACAG | qPCR |
| vp39-qRT-F | TCTAAATCTCAATTCCTCCGTG | qPCR |
| vp39-qRT-R | GCATTCTAGACACCACAAACC | qPCR |
| p10-qRT-F | CCATTGCGGAAACTAACACA | qPCR |
| p10-qRT-R | AGCAGTGTCACCGGTCAATA | qPCR |
| Bmrp49-qRT-F | TCAATCGGATCGCTATGACA | qPCR |
| Bmrp49-qRT-R | ATGACGGGTCTTCTTGTTGG | qPCR |
| lef-1-mut-F1 | GCGCCCAACGAGTTGAGATC | Mutagenesis detection |
| lef-1-mut-F2 | GCAATCAAGTATGTCGTCGT | Mutagenesis detection |
| lef-1-mut-R1 | AACTTCCGTAATATTACCGC | Mutagenesis detection |
| lef-1-mut-R2 | TTGGGCTTGACGACCGTAAC | Mutagenesis detection |
| lef-3-mut-F1 | GACGAGCGATTCCAAAACTT | Mutagenesis detection |
| lef-3-mut-F2 | GCAGATCAGGCTGTCAAATC | Mutagenesis detection |
| lef-3-mut-R1 | GCTTCGTGTCGGTCGTACGG | Mutagenesis detection |
| lef-3-mut-R2 | ATCGTTAAATCGAGCGGGTC | Mutagenesis detection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Chen, D.; Zhang, X.; Chen, S.; Yang, D.; Tang, L.; Yang, X.; Wang, Y.; Luo, X.; Wang, M.; et al. Construction of Baculovirus-Inducible CRISPR/Cas9 Antiviral System Targeting BmNPV in Bombyx mori. Viruses 2022, 14, 59. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010059
Liu Y, Chen D, Zhang X, Chen S, Yang D, Tang L, Yang X, Wang Y, Luo X, Wang M, et al. Construction of Baculovirus-Inducible CRISPR/Cas9 Antiviral System Targeting BmNPV in Bombyx mori. Viruses. 2022; 14(1):59. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010059
Chicago/Turabian StyleLiu, Yujia, Dongbin Chen, Xiaoqian Zhang, Shuqing Chen, Dehong Yang, Linmeng Tang, Xu Yang, Yaohui Wang, Xingyu Luo, Manli Wang, and et al. 2022. "Construction of Baculovirus-Inducible CRISPR/Cas9 Antiviral System Targeting BmNPV in Bombyx mori" Viruses 14, no. 1: 59. https://0-doi-org.brum.beds.ac.uk/10.3390/v14010059