
Fecal Feline Coronavirus RNA Shedding and Spike Gene Mutations in Cats with Feline Infectious Peritonitis Treated with GS-441524

 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vivo Study Design and Patients
2.2. Sample Collection for Determination of Viral Loads and Anti-FCoV Antibody Titers
2.3. Anti-FCoV Antibody Titers
2.4. FCoV RNA Loads in Feces, Blood, and Effusions
2.5. Sanger Sequencing to Detect Spike Gene Mutations in Fecal Samples, Blood, and Effusions
2.6. Statistics
3. Results
3.1. Treatment Study
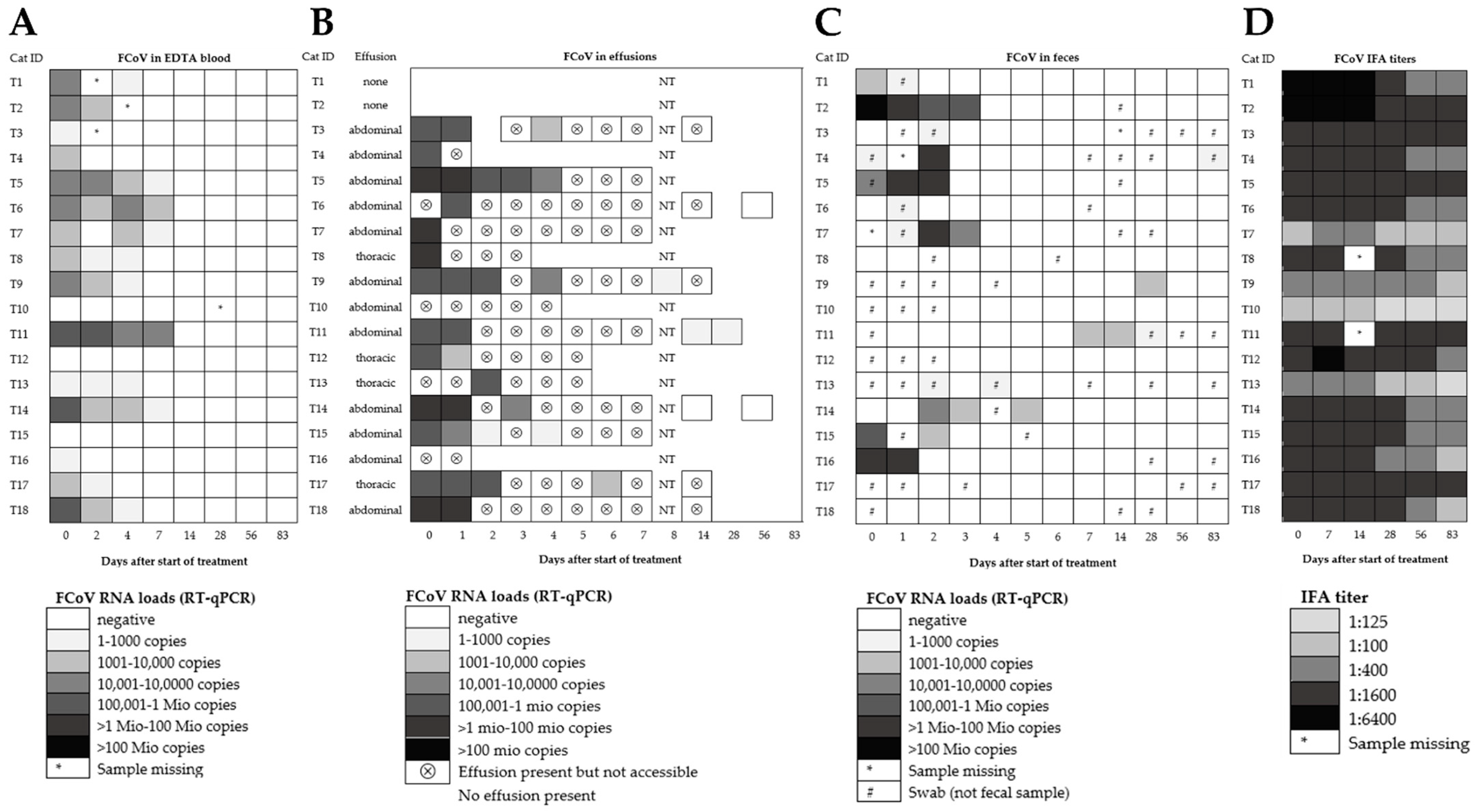
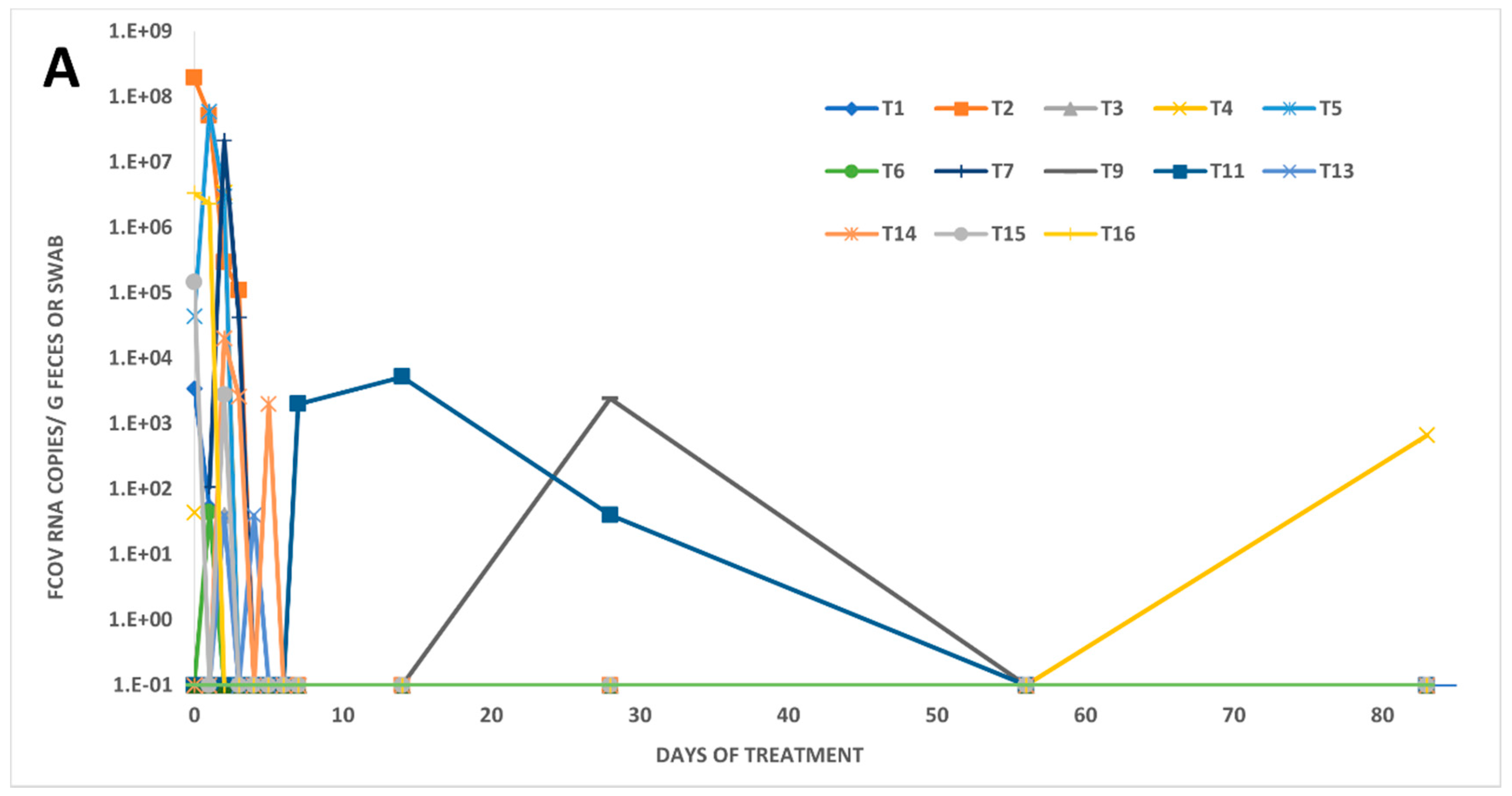
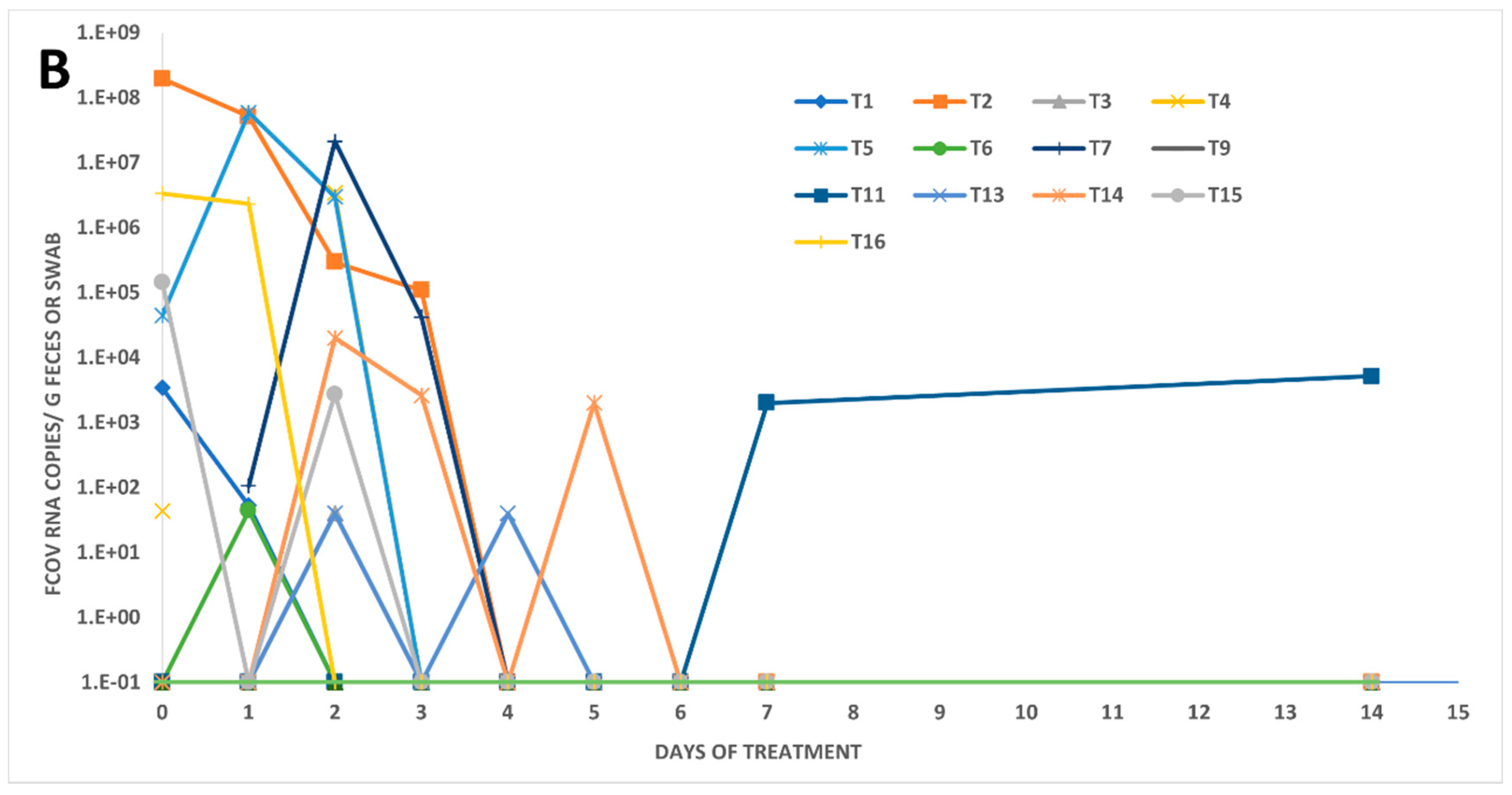
3.2. Fecal FCoV RNA Shedding and Viral Loads in Treated FIP Cats
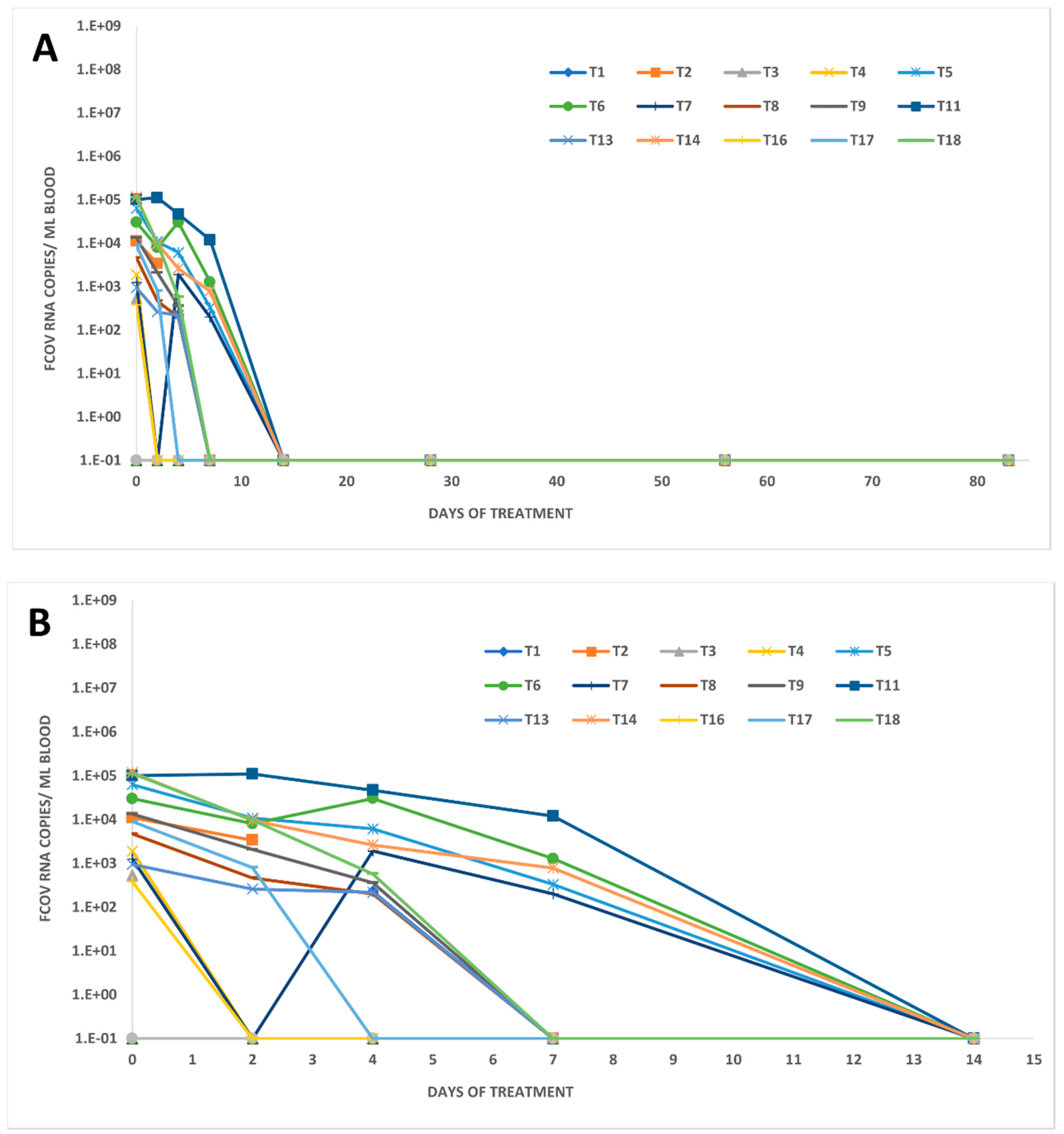
3.3. FCoV RNA Loads in Other Body Compartments of Treated FIP Cats
3.4. Anti-FCoV Antibody Titers in Treated FIP Cats
3.5. Spike Gene Mutations in Fecal Samples, Blood, and Effusions in Treated FIP Cats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
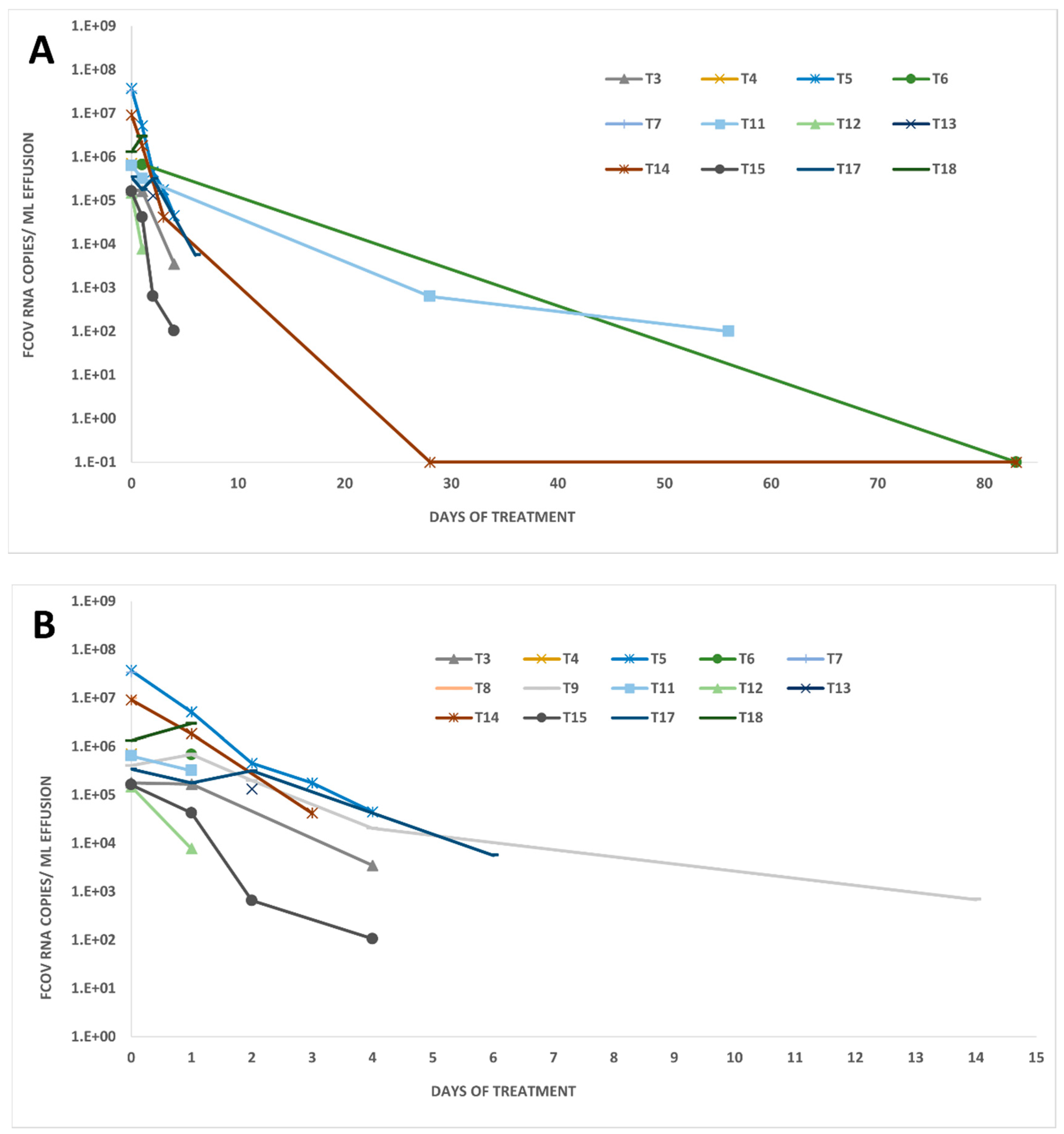{kind=link}
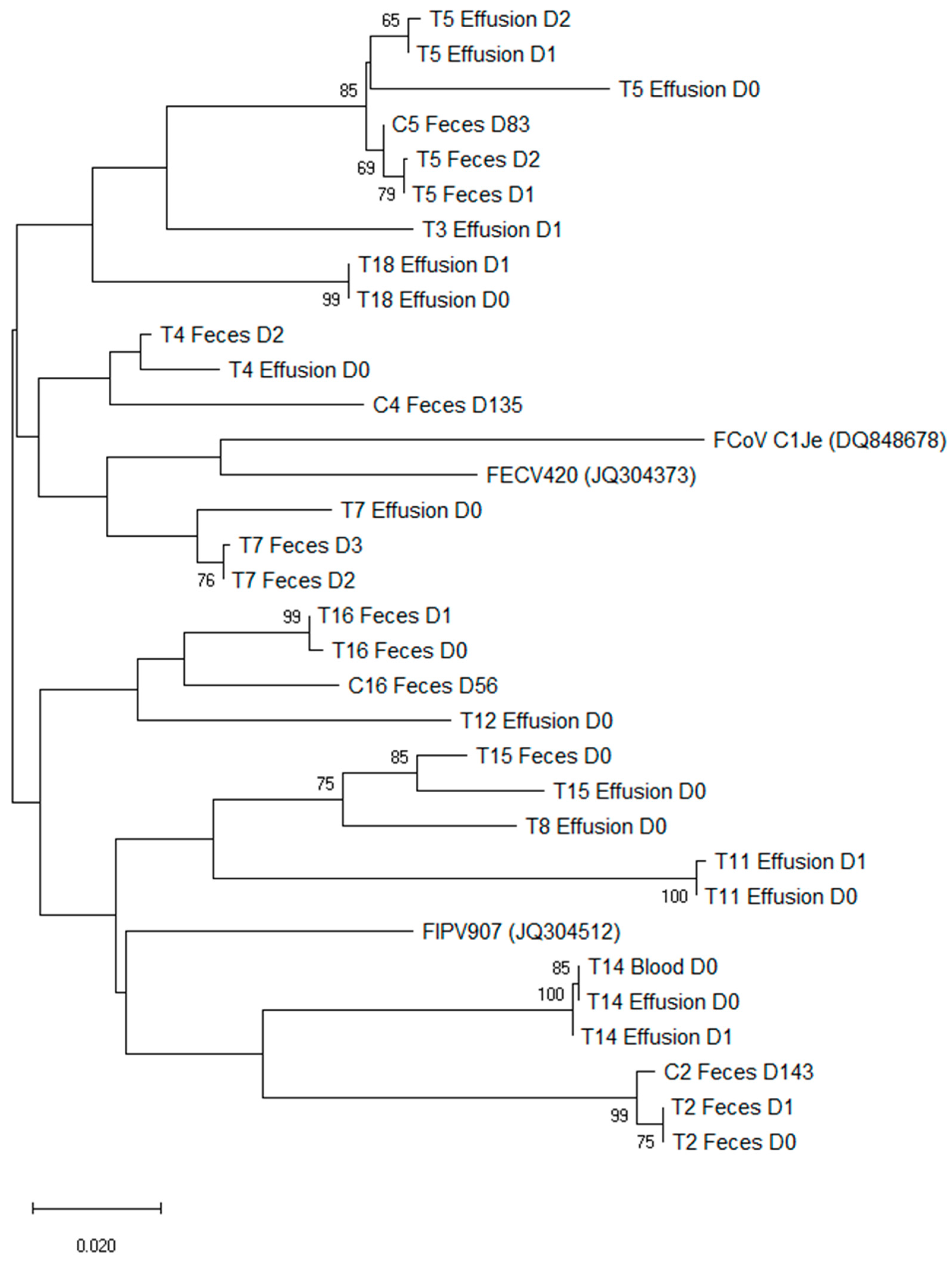{kind=link}
| Effusions | Blood | Fecal Samples | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cat Number | d0 | d1 | d2 | d3 | d4 | d0 | d2 | d4 | d7 | d0 | d1 | d2 | d3 | d4 | d5 | d7 | d14 | d28 | d56 | d83 |
| T1 | nb | x | n | x | x | n | n | n | n | n | n | n | n | n | ||||||
| T2 | nb | nb | n | MS | MS | nb | nb | n | n | n | n | n | n | n | ||||||
| T3 | nb | LS | nb | x | n | n | n | n | x | n | n | n | n | n | n | n | ||||
| T4 | LS | x | n | n | n | x | MS | n | n | n | n | n | n | n | x | |||||
| T5 | LS | LS | LS | nb | nb | nb | nb | nb | x | x | MS | MS | n | n | n | n | n | n | n | n |
| T6 | nb | nb | nb | nb | x | n | x | n | n | n | n | n | n | n | n | n | ||||
| T7 | LS | x | n | nb | x | x | MS | MS | n | n | n | n | n | n | n | |||||
| T8 | LS | nb | x | x | n | n | n | n | n | n | n | n | n | n | n | n | ||||
| T9 | nb | nb | nb | nb | nb | nb | x | n | n | n | n | n | n | n | n | n | x | n | n | |
| T10 | n | n | n | n | n | n | n | n | n | n | n | n | n | n | n | |||||
| T11 | FS * | FS * | nb | nb | nb | nb | n | n | n | n | n | n | x | x | x | n | n | |||
| T12 | MA * | nb | n | n | n | n | n | n | n | n | n | n | n | n | n | n | n | |||
| T13 | nb | x | x | x | n | n | n | x | n | x | n | n | n | n | n | n | ||||
| T14 | MA * | MA * | nb | MA * | nb | nb | x | n | n | nb | x | n | x | n | n | n | n | n | ||
| T15 | LS | nb | x | x | n | n | n | n | MS | n | x | n | n | n | n | n | n | n | n | |
| T16 | x | n | n | n | MS | MS | n | n | n | n | n | n | n | n | n | |||||
| T17 | nb | nb | nb | nb | x | n | n | n | n | n | n | n | n | n | n | n | n | n | ||
| T18 | LS | LS | nb | nb | x | n | n | n | n | n | n | n | n | n | n | n | n | |||
3.6. Fecal FCoV Shedding, Spike Gene Mutations, and Anti-FCoV Antibody Titers in Companion Cats
3.7. Phylogenetic Analysis of Sequences of Treated FIP Cats and Companion Cats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzalez, J.M.; Gomez-Puertas, P.; Cavanagh, D.; Gorbalenya, A.E.; Enjuanes, L. A comparative sequence analysis to revise the current taxonomy of the family Coronaviridae. Arch. Virol. 2003, 148, 2207–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauring, A.S.; Andino, R. Quasispecies theory and the behavior of RNA viruses. PLoS Pathog. 2010, 6, e1001005. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, Y.; Alcaraz, A.; Bossong, F.J.; Collisson, E.W.; Diniz, P.P. Feline coronavirus in multicat environments. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1133–1169. [Google Scholar] [CrossRef] [PubMed]
- Felten, S.; Klein-Richers, U.; Hofmann-Lehmann, R.; Bergmann, M.; Unterer, S.; Leutenegger, C.M.; Hartmann, K. Correlation of Feline Coronavirus Shedding in Feces with Coronavirus Antibody Titer. Pathogens 2020, 9, 598. [Google Scholar] [CrossRef]
- Klein-Richers, U.; Hartmann, K.; Hofmann-Lehmann, R.; Unterer, S.; Bergmann, M.; Rieger, A.; Leutenegger, C.; Pantchev, N.; Balzer, J.; Felten, S. Prevalence of Feline Coronavirus Shedding in German Catteries and Associated Risk Factors. Viruses 2020, 12, 1000. [Google Scholar] [CrossRef]
- Pedersen, N.C. A review of feline infectious peritonitis virus infection: 1963–2008. J. Feline Med. Surg. 2009, 11, 225–258. [Google Scholar] [CrossRef]
- Foley, J.E.; Poland, A.; Carlson, J.; Pedersen, N.C. Risk factors for feline infectious peritonitis among cats in multiple-cat environments with endemic feline enteric coronavirus. J. Am. Vet. Med. Assoc. 1997, 210, 1313–1318. [Google Scholar]
- Pedersen, N.C.; Allen, C.E.; Lyons, L.A. Pathogenesis of feline enteric coronavirus infection. J. Feline Med. Surg. 2008, 10, 529–541. [Google Scholar] [CrossRef] [Green Version]
- Addie, D.; Belak, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; Lutz, H.; et al. Feline infectious peritonitis. ABCD guidelines on prevention and management. J. Feline Med. Surg. 2009, 11, 594–604. [Google Scholar] [CrossRef]
- Addie, D.D.; Toth, S.; Murray, G.D.; Jarrett, O. Risk of feline infectious peritonitis in cats naturally infected with feline coronavirus. Am. J. Vet. Res. 1995, 56, 429–434. [Google Scholar]
- Kipar, A.; May, H.; Menger, S.; Weber, M.; Leukert, W.; Reinacher, M. Morphologic features and development of granulomatous vasculitis in feline infectious peritonitis. Vet. Pathol. 2005, 42, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Vennema, H.; Poland, A.; Foley, J.; Pedersen, N.C. Feline infectious peritonitis viruses arise by mutation from endemic feline enteric coronaviruses. Virology 1998, 243, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.C.; Boyle, J.F.; Floyd, K.; Fudge, A.; Barker, J. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. Am. J. Vet. Res. 1981, 42, 368–377. [Google Scholar] [PubMed]
- Dewerchin, H.L.; Cornelissen, E.; Nauwynck, H.J. Replication of feline coronaviruses in peripheral blood monocytes. Arch. Virol. 2005, 150, 2483–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipar, A.; Meli, M.L. Feline infectious peritonitis: Still an enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malbon, A.J.; Michalopoulou, E.; Meli, M.L.; Barker, E.N.; Tasker, S.; Baptiste, K.; Kipar, A. Colony Stimulating Factors in Early Feline Infectious Peritonitis Virus Infection of Monocytes and in End Stage Feline Infectious Peritonitis; A Combined In Vivo And In Vitro Approach. Pathogens 2020, 9, 893. [Google Scholar] [CrossRef]
- Stoddart, C.A.; Scott, F.W. Intrinsic resistance of feline peritoneal macrophages to coronavirus infection correlates with in vivo virulence. J. Virol. 1989, 63, 436–440. [Google Scholar] [CrossRef] [Green Version]
- Gunn-Moore, D.A.; Gruffydd-Jones, T.J.; Harbour, D.A. Detection of feline coronaviruses by culture and reverse transcriptase-polymerase chain reaction of blood samples from healthy cats and cats with clinical feline infectious peritonitis. Vet. Microbiol. 1998, 62, 193–205. [Google Scholar] [CrossRef]
- Meli, M.; Kipar, A.; Muller, C.; Jenal, K.; Gonczi, E.; Borel, N.; Gunn-Moore, D.; Chalmers, S.; Lin, F.; Reinacher, M.; et al. High viral loads despite absence of clinical and pathological findings in cats experimentally infected with feline coronavirus (FCoV) type I and in naturally FCoV-infected cats. J. Feline Med. Surg. 2004, 6, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Hornyak, A.; Balint, A.; Farsang, A.; Balka, G.; Hakhverdyan, M.; Rasmussen, T.B.; Blomberg, J.; Belak, S. Detection of subgenomic mRNA of feline coronavirus by real-time polymerase chain reaction based on primer-probe energy transfer (P-sg-QPCR). J. Virol. Methods 2012, 181, 155–163. [Google Scholar] [CrossRef]
- Kipar, A.; Baptiste, K.; Barth, A.; Reinacher, M. Natural FCoV infection: Cats with FIP exhibit significantly higher viral loads than healthy infected cats. J. Feline Med. Surg. 2006, 8, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Simons, F.A.; Vennema, H.; Rofina, J.E.; Pol, J.M.; Horzinek, M.C.; Rottier, P.J.; Egberink, H.F. A mRNA PCR for the diagnosis of feline infectious peritonitis. J. Virol. Methods 2005, 124, 111–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipar, A.; Meli, M.L.; Baptiste, K.E.; Bowker, L.J.; Lutz, H. Sites of feline coronavirus persistence in healthy cats. J. Gen. Virol. 2010, 91, 1698–1707. [Google Scholar] [CrossRef] [Green Version]
- Addie, D.D.; Schaap, I.A.; Nicolson, L.; Jarrett, O. Persistence and transmission of natural type I feline coronavirus infection. J. Gen. Virol. 2003, 84, 2735–2744. [Google Scholar] [CrossRef]
- Vogel, L.; Van der Lubben, M.; te Lintelo, E.G.; Bekker, C.P.; Geerts, T.; Schuijff, L.S.; Grinwis, G.C.; Egberink, H.F.; Rottier, P.J. Pathogenic characteristics of persistent feline enteric coronavirus infection in cats. Vet. Res. 2010, 41, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addie, D.D.; Jarrett, O. Use of a reverse-transcriptase polymerase chain reaction for monitoring the shedding of feline coronavirus by healthy cats. Vet. Rec. 2001, 148, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.E.; Poland, A.; Carlson, J.; Pedersen, N.C. Patterns of feline coronavirus infection and fecal shedding from cats in multiple-cat environments. J. Am. Vet. Med. Assoc. 1997, 210, 1307–1312. [Google Scholar] [PubMed]
- Harpold, L.M.; Legendre, A.M.; Kennedy, M.A.; Plummer, P.J.; Millsaps, K.; Rohrbach, B. Fecal shedding of feline coronavirus in adult cats and kittens in an Abyssinian cattery. J. Am. Vet. Med. Assoc. 1999, 215, 948–951. [Google Scholar]
- Herrewegh, A.A.; Mahler, M.; Hedrich, H.J.; Haagmans, B.L.; Egberink, H.F.; Horzinek, M.C.; Rottier, P.J.; de Groot, R.J. Persistence and evolution of feline coronavirus in a closed cat-breeding colony. Virology 1997, 234, 349–363. [Google Scholar] [CrossRef] [Green Version]
- Desmarets, L.M.; Vermeulen, B.L.; Theuns, S.; Conceicao-Neto, N.; Zeller, M.; Roukaerts, I.D.; Acar, D.D.; Olyslaegers, D.A.; Van Ranst, M.; Matthijnssens, J.; et al. Experimental feline enteric coronavirus infection reveals an aberrant infection pattern and shedding of mutants with impaired infectivity in enterocyte cultures. Sci. Rep. 2016, 6, 20022. [Google Scholar] [CrossRef]
- Addie, D.D.; le Poder, S.; Burr, P.; Decaro, N.; Graham, E.; Hofmann-Lehmann, R.; Jarrett, O.; McDonald, M.; Meli, M.L. Utility of feline coronavirus antibody tests. J. Feline Med. Surg. 2015, 17, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Lutz, H.; Gut, M.; Leutenegger, C.M.; Schiller, I.; Meli, M.L. Inetics of FCoV Infection in Kittens Born in Catteries of High risk for FIP under Different Rearing Conditions. In Proceedings of the Second International Feline Coronavirus/Feline Infectious Peritonitis Symposium, Glasgow, Scotland, 4–7 August 2002. [Google Scholar]
- Hartmann, K. Feline infectious peritonitis. Vet. Clin. N. Am. Small Anim. Pract. 2005, 35, 39–79. [Google Scholar] [CrossRef] [PubMed]
- Rohner-Mächler, M. Bestimmung der Ausscheidungskinetik von Felinen Coronaviren unter Feldbedingungen. Inaugural Thesis, University of Zurich, Switzerland, Zurich, 1999. [Google Scholar]
- Bell, E.T.; Toribio, J.A.; White, J.D.; Malik, R.; Norris, J.M. Seroprevalence study of feline coronavirus in owned and feral cats in Sydney, Australia. Aust. Vet. J. 2006, 84, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Malbon, A.J.; Russo, G.; Burgener, C.; Barker, E.N.; Meli, M.L.; Tasker, S.; Kipar, A. The Effect of Natural Feline Coronavirus Infection on the Host Immune Response: A Whole-Transcriptome Analysis of the Mesenteric Lymph Nodes in Cats with and without Feline Infectious Peritonitis. Pathogens 2020, 9, 524. [Google Scholar] [CrossRef]
- Mustaffa-Kamal, F.; Liu, H.; Pedersen, N.C.; Sparger, E.E. Characterization of antiviral T cell responses during primary and secondary challenge of laboratory cats with feline infectious peritonitis virus (FIPV). BMC Vet. Res. 2019, 15, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaplace, M.; Huet, H.; Gambino, A.; Le Poder, S. Feline Coronavirus Antivirals: A Review. Pathogens 2021, 10, 1150. [Google Scholar] [CrossRef]
- Dickinson, P.J.; Bannasch, M.; Thomasy, S.M.; Murthy, V.D.; Vernau, K.M.; Liepnieks, M.; Montgomery, E.; Knickelbein, K.E.; Murphy, B.; Pedersen, N.C. Antiviral treatment using the adenosine nucleoside analogue GS-441524 in cats with clinically diagnosed neurological feline infectious peritonitis. J. Vet. Intern. Med. 2020, 34, 1587–1593. [Google Scholar] [CrossRef]
- Murphy, B.G.; Perron, M.; Murakami, E.; Bauer, K.; Park, Y.; Eckstrand, C.; Liepnieks, M.; Pedersen, N.C. The nucleoside analog GS-441524 strongly inhibits feline infectious peritonitis (FIP) virus in tissue culture and experimental cat infection studies. Vet. Microbiol. 2018, 219, 226–233. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Kim, Y.; Liu, H.; Galasiti Kankanamalage, A.C.; Eckstrand, C.; Groutas, W.C.; Bannasch, M.; Meadows, J.M.; Chang, K.O. Efficacy of a 3C-like protease inhibitor in treating various forms of acquired feline infectious peritonitis. J. Feline Med. Surg. 2018, 20, 378–392. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Perron, M.; Bannasch, M.; Montgomery, E.; Murakami, E.; Liepnieks, M.; Liu, H. Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis. J. Feline Med. Surg. 2019, 21, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Krentz, D.; Zenger, K.; Alberer, M.; Felten, S.; Bergmann, M.; Dorsch, R.; Matiasek, K.; Kolberg, L.; Hofmann-Lehmann, R.; Meli, M.L.; et al. Curing Cats with Feline Infectious Peritonitis with an Oral Multi-Component Drug Containing GS-441524. Viruses 2021, 13, 2228. [Google Scholar] [CrossRef]
- Addie, D.D.; Covell-Ritchie, J.; Jarrett, O.; Fosbery, M. Rapid Resolution of Non-Effusive Feline Infectious Peritonitis Uveitis with an Oral Adenosine Nucleoside Analogue and Feline Interferon Omega. Viruses 2020, 12, 1216. [Google Scholar] [CrossRef] [PubMed]
- Addie, D.D.; Curran, S.; Bellini, F.; Crowe, B.; Sheehan, E.; Ukrainchuk, L.; Decaro, N. Oral Mutian(R)X stopped faecal feline coronavirus shedding by naturally infected cats. Res. Vet. Sci. 2020, 130, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottier, P.J. Spike protein fusion peptide and feline coronavirus virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Brunner, C.; Kanellos, T.; Meli, M.L.; Sutton, D.J.; Gisler, R.; Gomes-Keller, M.A.; Hofmann-Lehmann, R.; Lutz, H. Antibody induction after combined application of an adjuvanted recombinant FeLV vaccine and a multivalent modified live virus vaccine with a chlamydial component. Vaccine 2006, 24, 1838–1846. [Google Scholar] [CrossRef]
- Lutz, H.; Hauser, B.; Horzinek, M. On the serological diagnosis of feline infectious peritonitis. Prakt Tierarzt 1984, 65, 406–408. [Google Scholar]
- Osterhaus, A.D.; Horzinek, M.C.; Reynolds, D.J. Seroepidemiology of feline infectious peritonitis virus infections using transmissible gastroenteritis virus as antigen. Zent. Vet. B 1977, 24, 835–841. [Google Scholar] [CrossRef]
- Gut, M.; Leutenegger, C.M.; Huder, J.B.; Pedersen, N.C.; Lutz, H. One-tube fluorogenic reverse transcription-polymerase chain reaction for the quantitation of feline coronaviruses. J. Virol. Methods 1999, 77, 37–46. [Google Scholar] [CrossRef]
- Tandon, R.; Cattori, V.; Gomes-Keller, M.A.; Meli, M.L.; Golder, M.C.; Lutz, H.; Hofmann-Lehmann, R. Quantitation of feline leukaemia virus viral and proviral loads by TaqMan real-time polymerase chain reaction. J. Virol. Methods 2005, 130, 124–132. [Google Scholar] [CrossRef]
- Lutz, M.; Steiner, A.R.; Cattori, V.; Hofmann-Lehmann, R.; Lutz, H.; Kipar, A.; Meli, M.L. FCoV Viral Sequences of Systemically Infected Healthy Cats Lack Gene Mutations Previously Linked to the Development of FIP. Pathogens 2020, 9, 603. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzhetsky, A.; Nei, M. Statistical properties of the ordinary least-squares, generalized least-squares, and minimum-evolution methods of phylogenetic inference. J. Mol. Evol. 1992, 35, 367–375. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Chang, H.W.; de Groot, R.J.; Egberink, H.F.; Rottier, P.J. Feline infectious peritonitis: Insights into feline coronavirus pathobiogenesis and epidemiology based on genetic analysis of the viral 3c gene. J. Gen. Virol. 2010, 91, 415–420. [Google Scholar] [CrossRef]
- Barker, E.N.; Stranieri, A.; Helps, C.R.; Porter, E.L.; Davidson, A.D.; Day, M.J.; Knowles, T.; Kipar, A.; Tasker, S. Limitations of using feline coronavirus spike protein gene mutations to diagnose feline infectious peritonitis. Vet. Res. 2017, 48, 60. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, N.C.; Liu, H.; Scarlett, J.; Leutenegger, C.M.; Golovko, L.; Kennedy, H.; Kamal, F.M. Feline infectious peritonitis: Role of the feline coronavirus 3c gene in intestinal tropism and pathogenicity based upon isolates from resident and adopted shelter cats. Virus Res. 2012, 165, 17–28. [Google Scholar] [CrossRef]
- Potten, C.S. Stem cells in gastrointestinal epithelium: Numbers, characteristics and death. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish, E.J.; Diniz, P.P.V.; Juan, Y.C.; Bossong, F.; Collisson, E.W.; Drechsler, Y.; Kaltenboeck, B. Cross-sectional quantitative RT-PCR study of feline coronavirus viremia and replication in peripheral blood of healthy shelter cats in Southern California. J. Feline Med. Surg. 2018, 20, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Doenges, S.J.; Weber, K.; Dorsch, R.; Fux, R.; Hartmann, K. Comparison of real-time reverse transcriptase polymerase chain reaction of peripheral blood mononuclear cells, serum and cell-free body cavity effusion for the diagnosis of feline infectious peritonitis. J. Feline Med. Surg. 2017, 19, 344–350. [Google Scholar] [CrossRef]
- Felten, S.; Leutenegger, C.M.; Balzer, H.J.; Pantchev, N.; Matiasek, K.; Wess, G.; Egberink, H.; Hartmann, K. Sensitivity and specificity of a real-time reverse transcriptase polymerase chain reaction detecting feline coronavirus mutations in effusion and serum/plasma of cats to diagnose feline infectious peritonitis. BMC Vet. Res. 2017, 13, 228. [Google Scholar] [CrossRef] [PubMed]
- Felten, S.; Weider, K.; Doenges, S.; Gruendl, S.; Matiasek, K.; Hermanns, W.; Mueller, E.; Matiasek, L.; Fischer, A.; Weber, K.; et al. Detection of feline coronavirus spike gene mutations as a tool to diagnose feline infectious peritonitis. J. Feline Med. Surg. 2017, 19, 321–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.C.; Eckstrand, C.; Liu, H.; Leutenegger, C.; Murphy, B. Levels of feline infectious peritonitis virus in blood, effusions, and various tissues and the role of lymphopenia in disease outcome following experimental infection. Vet. Microbiol. 2015, 175, 157–166. [Google Scholar] [CrossRef]
- Longstaff, L.; Porter, E.; Crossley, V.J.; Hayhow, S.E.; Helps, C.R.; Tasker, S. Feline coronavirus quantitative reverse transcriptase polymerase chain reaction on effusion samples in cats with and without feline infectious peritonitis. J. Feline Med. Surg. 2017, 19, 240–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stranieri, A.; Giordano, A.; Paltrinieri, S.; Giudice, C.; Cannito, V.; Lauzi, S. Comparison of the performance of laboratory tests in the diagnosis of feline infectious peritonitis. J. Vet. Diagn. Investig. 2018, 30, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Jacobse-Geels, H.E.; Daha, M.R.; Horzinek, M.C. Antibody, immune complexes, and complement activity fluctuations in kittens with experimentally induced feline infectious peritonitis. Am. J. Vet. Res. 1982, 43, 666–670. [Google Scholar]
- Olsen, C.W.; Corapi, W.V.; Ngichabe, C.K.; Baines, J.D.; Scott, F.W. Monoclonal antibodies to the spike protein of feline infectious peritonitis virus mediate antibody-dependent enhancement of infection of feline macrophages. J. Virol. 1992, 66, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, N.C. Virologic and immunologic aspects of feline infectious peritonitis virus infection. Adv. Exp. Med. Biol. 1987, 218, 529–550. [Google Scholar]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Whittaker, G.R. Host cell proteases: Critical determinants of coronavirus tropism and pathogenesis. Virus Res. 2015, 202, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Mari, V.; Lanave, G.; Lorusso, E.; Lucente, M.S.; Desario, C.; Colaianni, M.L.; Elia, G.; Ferringo, F.; Alfano, F.; et al. Mutation analysis of the spike protein in Italian feline infectious peritonitis virus and feline enteric coronavirus sequences. Res. Vet. Sci. 2021, 135, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Porter, E.; Tasker, S.; Day, M.J.; Harley, R.; Kipar, A.; Siddell, S.G.; Helps, C.R. Amino acid changes in the spike protein of feline coronavirus correlate with systemic spread of virus from the intestine and not with feline infectious peritonitis. Vet. Res. 2014, 45, 49. [Google Scholar] [CrossRef] [Green Version]
- Jähne, S.; Felten, S.; Bergmann, M.; Erber, K.; Matiasek, K.; Meli, M.L.; Hofmann-Lehmann, R.; Hartmann, K. Detection of feline coronavirus variants in cats without feline infectious peritonitis. Viruses, 2022; Submitted. [Google Scholar]
- Brown, M.A.; Troyer, J.L.; Pecon-Slattery, J.; Roelke, M.E.; O’Brien, S.J. Genetics and pathogenesis of feline infectious peritonitis virus. Emerg. Infect. Dis. 2009, 15, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Liu, H.; Dodd, K.A.; Pesavento, P.A. Significance of coronavirus mutants in feces and diseased tissues of cats suffering from feline infectious peritonitis. Viruses 2009, 1, 166–184. [Google Scholar] [CrossRef] [Green Version]
- Fischer, Y.; Ritz, S.; Weber, K.; Sauter-Louis, C.; Hartmann, K. Randomized, placebo controlled study of the effect of propentofylline on survival time and quality of life of cats with feline infectious peritonitis. J. Vet. Intern. Med. 2011, 25, 1270–1276. [Google Scholar] [CrossRef]
- Ritz, S.; Egberink, H.; Hartmann, K. Effect of feline interferon-omega on the survival time and quality of life of cats with feline infectious peritonitis. J. Vet. Intern. Med. 2007, 21, 1193–1197. [Google Scholar] [CrossRef]






| Identification of Treated FIP Cats | Age (Months) | Sex 1 | FIP-Associated Cardinal Signs 2 | Number of Companion Cats in the Same Household | Identification of Companion Cats with Samples Available | Collection Time Point (Day of Collection) 3 of Fecal and Serum Samples from Companion Cats |
|---|---|---|---|---|---|---|
| T1 | 6 | mn | OS | 1 | C1 4 | 145 |
| T2 | 6 | mi | NS + OS | 1 | C2 | 143 |
| T3 | 10 | mn | AE | 1 | ||
| T4 | 7 | mi | AE | 1 | C4 | 135 |
| T5 | 6 | fi | AE | 1 | C5 | 83 |
| T6 | 11 | mn | AE | 1 | ||
| T7 | 5 | mi | AE + TE | 1 | ||
| T8 | 6 | mi | TE | 0 | ||
| T9 | 9 | mn | AE | 3 | ||
| T10 | 39 | fn | AE | 3 | ||
| T11 | 57 | fn | AE | 3 | C11A, C11B, C11C | 83 |
| T12 | 12 | mn | TE | 1 | C12 | 83 |
| T13 | 29 | fi | TE | 9 | ||
| T14 | 8 | mi | AE | 1 | C14 | 56 |
| T15 | 8 | mi | AE | 1 | C15 | 56 |
| T16 | 9 | fn | AE | 1 | C16 | 56 |
| T17 | 8 | mi | TE | 1 | ||
| T18 | 8 | fi | AE | 2 | C18 | 56 |
| Companion Cat Number | Corresponding Treated FIP Cat | Day of Sample Collection in Companion Cats | Fecal Samples Collected | Fecal RNA Loads (Copies per g Feces or per Swab) | Sequencing Result | Anti-FCoV Antibody Titer |
|---|---|---|---|---|---|---|
| C1 | T1 | 145 | Swab | n | n | 1:1600 |
| C2 | T2 | 143 | Feces | 66,337,219 | MS | 1:100 |
| C4 | T4 1 | 135 | Swab | 27,545 | MS | 1:400 |
| C5 | T5 | 83 | Feces | 12,990,572 | MS | 1:1600 |
| C11A | T11 2 | 83 | Swab | 1906 | nb | 1:100 |
| C11B | T11 2 | 83 | Swab | 346,600 | nb | 1:400 |
| C11C | T11 2 | 83 | Swab | 178,345 | nb | 1:400 |
| C12 | T12 | 83 | Feces | n | n | 1:100 |
| C14 | T14 | 56 | Swab | n | n | 1:25 * |
| C15 | T15 | 56 | Feces | 159,536 | nb | 1:400 |
| C16 | T16 | 56 | Feces | 74,714,117 | MS | 1:100 |
| C18 | T18 | 56 | Swab | n | n | 1:100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meli, M.L.; Spiri, A.M.; Zwicklbauer, K.; Krentz, D.; Felten, S.; Bergmann, M.; Dorsch, R.; Matiasek, K.; Alberer, M.; Kolberg, L.; et al. Fecal Feline Coronavirus RNA Shedding and Spike Gene Mutations in Cats with Feline Infectious Peritonitis Treated with GS-441524. Viruses 2022, 14, 1069. https://0-doi-org.brum.beds.ac.uk/10.3390/v14051069
Meli ML, Spiri AM, Zwicklbauer K, Krentz D, Felten S, Bergmann M, Dorsch R, Matiasek K, Alberer M, Kolberg L, et al. Fecal Feline Coronavirus RNA Shedding and Spike Gene Mutations in Cats with Feline Infectious Peritonitis Treated with GS-441524. Viruses. 2022; 14(5):1069. https://0-doi-org.brum.beds.ac.uk/10.3390/v14051069
Chicago/Turabian StyleMeli, Marina L., Andrea M. Spiri, Katharina Zwicklbauer, Daniela Krentz, Sandra Felten, Michèle Bergmann, Roswitha Dorsch, Kaspar Matiasek, Martin Alberer, Laura Kolberg, and et al. 2022. "Fecal Feline Coronavirus RNA Shedding and Spike Gene Mutations in Cats with Feline Infectious Peritonitis Treated with GS-441524" Viruses 14, no. 5: 1069. https://0-doi-org.brum.beds.ac.uk/10.3390/v14051069