1. Introduction
Mesenchymal stem cells (MSCs) can be isolated from a variety of tissues, such as bone marrow, umbilical cord, and adipose tissue. Adipose tissue-derived MSCs (AD-MSCs) are easy to harvest, have excellent proliferation potential, and are progenitor cells that harbor valuable therapeutic and biological activities [
1,
2]. Studies have revealed the many abilities of AD-MSCs, such as reducing inflammation, modulating immune responses, and promoting tissue regeneration [
3,
4,
5]. The therapeutic capacities of MSCs mainly rely on the variety of their secretomes [
6,
7,
8]. Recently, several clinical studies involving the use of MSCs and/or their secretomes to treat viral infection in human beings, such as hepatitis B virus, human immunodeficiency virus, and severe acute respiratory syndrome coronavirus 2, have been registered [
9].
Feline calicivirus (FCV) is a highly contagious pathogen of cats that is found worldwide and causes upper respiratory tract and oral disease [
10]. Feline herpesvirus 1 (FHV1) is one of the most common viruses among cats, and FHV1 infection is associated with respiratory and ocular diseases [
11]. Vaccinations against these viruses are widespread throughout the world, but they are not fully effective [
12,
13]; thus, it is of urgency that we develop effective and safe antiviral drugs for veterinary medicine. Commercial formulation of recombinant type I interferons (IFNs) is sometimes use against FCV. IFNs are involved in antiviral responses and have broad spectrum antiviral activities [
14]; however, many viruses have required multiple strategies to escape or inhibit the IFN response [
15,
16,
17]. Moreover, some FCV strains do not promote the activation of the IFNβ promoter, allowing these viruses to evade the IFN response [
18]. Antiviral drugs against FHV1 have also been examined for their effects on plaque number and plaque size in vitro, and it was demonstrated that ganciclovir, 9-(2-phosphonylmethoxyethyl)-2,6-diaminopurine (PMEDAP), and cidofovir are the most potent inhibitors of FHV1 replication in Crandell–Reese feline kidney (CRFK) cells [
19].
Feline chronic gingivostomatitis (FCGS) is a multifactorial disease and FCV is one of the causes associated with it. FCGS-affected cats with FCV and puma feline foamy virus may poorly respond to treatment [
20]. Recently, feline chronic gingivostomatitis (FCGS) was reportedly treated with AD-MSCs [
21,
22,
23]. In the report, feline AD-MSCs administered systemically resulted in favorable clinical, histologic, and systemic responses in over 70% of cats [
23]. However, it remains unknown whether feline AD-MSCs have antiviral effects on FCV and FHV1. In this study, the antiviral effects of secretome from feline AD-MSCs on FCV and FHV1 replication were investigated by measuring changes in viral mRNA (vmRNA) levels and virus yields in CRFK cells. Then, gene expression was performed using RNA sequencing to detect any possible alterations related to the inhibition of these viruses between the mock CRFK cells and the CRFK cells sensitized to feline AD-MSC secretome.
2. Materials and Methods
2.1. Isolation and Expansion of Feline AD-MSCs
Adipose tissue was aseptically collected from falciform ligament fat of three healthy anaesthetized client-owned Japanese bobtail short-hair cats (female, mean body weight of 3.3 kg, mean age of 9 months) when they were scheduled to be spayed. The tissue was washed extensively in a phosphate buffer solution (PBS), minced, and digested with collagenase type I (Sigma-Aldrich) at 37 °C for 45 min with shaking. After washing with PBS and centrifuging, the pellets containing the stromal vascular fraction were resuspended, filtered through a 100-μm nylon mesh, and incubated overnight in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS; Capricorn Scientific) and a 1% antibiotic–antimycotic solution (Thermo Fisher Scientific) in a humidified atmosphere with 5% CO2 at 37 °C. Unattached cells were removed by changing the medium, and the attached cells were washed twice with PBS. Thereafter, the medium was replaced every 3–4 days. At 80–90% confluence, the cells were detached with trypsin-EDTA solution (Sigma-Aldrich) and passaged repeatedly.
2.2. Characterization of Feline AD-MSCs
The expression of several markers, such as CD14-FITC, CD29-PE, CD34-FITC, CD44-PE, CD45-PE, CD90-PE, CD105-PE, and MHC-II-FITC on passage 2 feline AD-MSCs was determined by flow cytometry using a CytoFLEX (BECKMAN COULTER). The capacity of the cells to differentiate into osteogenic, chondrogenic, and adipogenic lineages was evaluated.
2.3. Production of fADSC-CM
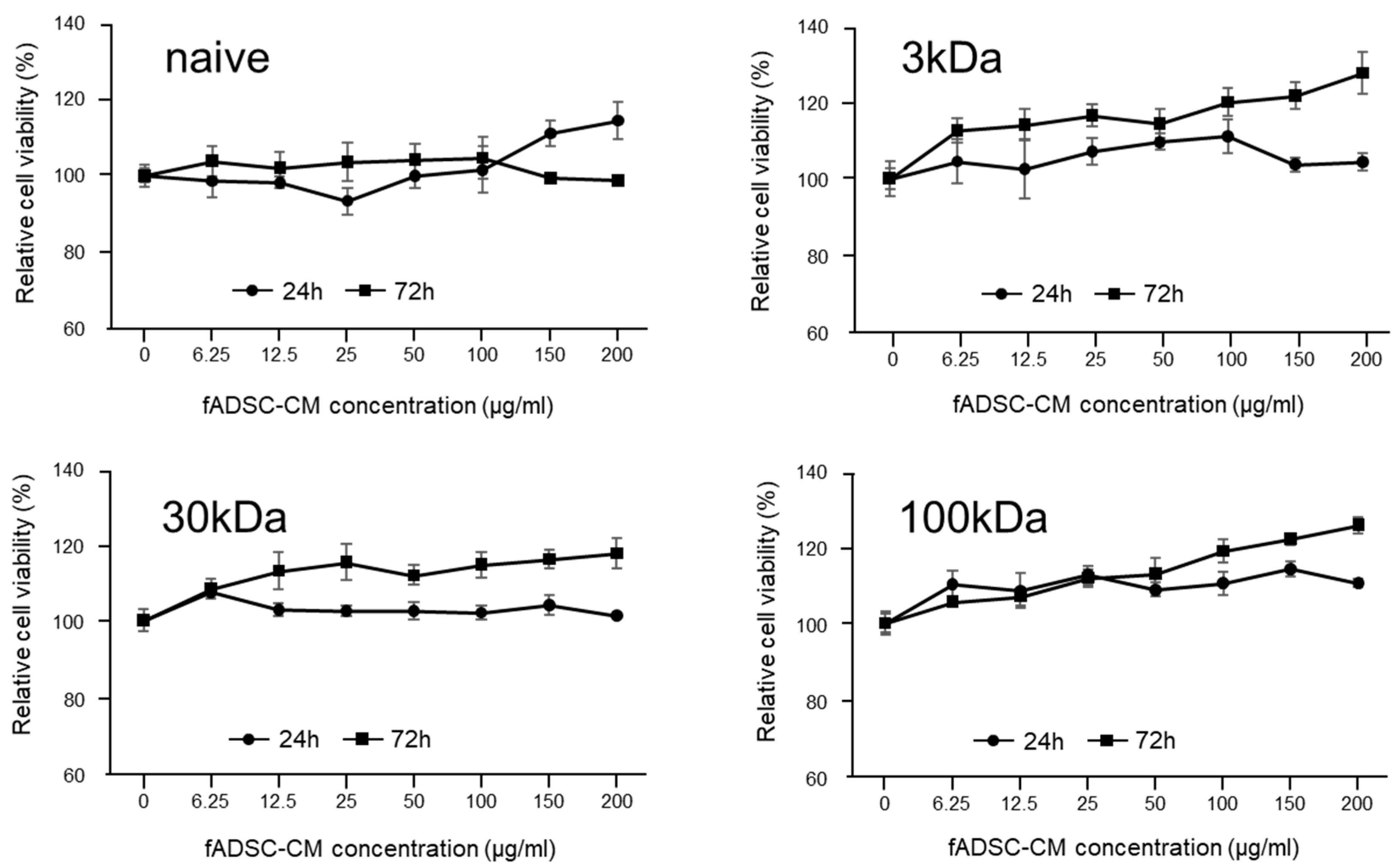
In this study, feline AD-MSC secretome was collected as conditioned medium (CM). To prepare the feline AD-MSC CM (fADSC-CM), passage 2 feline AD-MSCs were seeded (3.0 × 104 cells/cm2) individually into DMEM supplemented with 10% FBS and 1% antibiotic–antimycotic solution and incubated overnight. Adherent cells were washed and further incubated in FBS-free DMEM for 36 h, then the medium was centrifuged at 300× g for 5 min and 1200× g for 20 min at 4 °C to remove cells and debris. The naïve CM was concentrated using molecular weight cut-off (MWCO) ultrafiltration with membranes of 3 kDa, 30 kDa, and 100 kDa. The total protein concentrations of naïve and MWCO-concentrated supernatants were measured with a BCA assay kit, then stored at −80 °C until further assay.
2.4. Cells and Viruses
The CRFK cells were obtained from the American Type Culture Collection. The cells were cultured in DMEM containing 10% FBS, 100 U/mL penicillin, and 100 µg/mL streptomycin (complete medium). FCV strain F4 and FHV1 strain C7301 were used in this study. The viral titers of FCV and FHV1 were determined by median tissue culture infectious dose (TCID50) assay.
2.5. Cytotoxicity Assay for fADSC-CM
Prior to the assay, the CRFK cells (2.0 × 104 cells/well, 100 µL/well) were seeded into flat-bottom 96-well plates and incubated overnight with 5% CO2 at 37 °C. The cell monolayers were then treated with a series of protein concentrations (6.25, 12.5, 25, 50, 100, 150, 200 µg/mL) of fADSC-CM for 24 h and 72 h. Cytotoxicity assays were performed using a Cell Counting Kit-8 according (Dojindo) to the manufacturer’s instructions.
2.6. Effect of fADSC-CM on Viral Replication
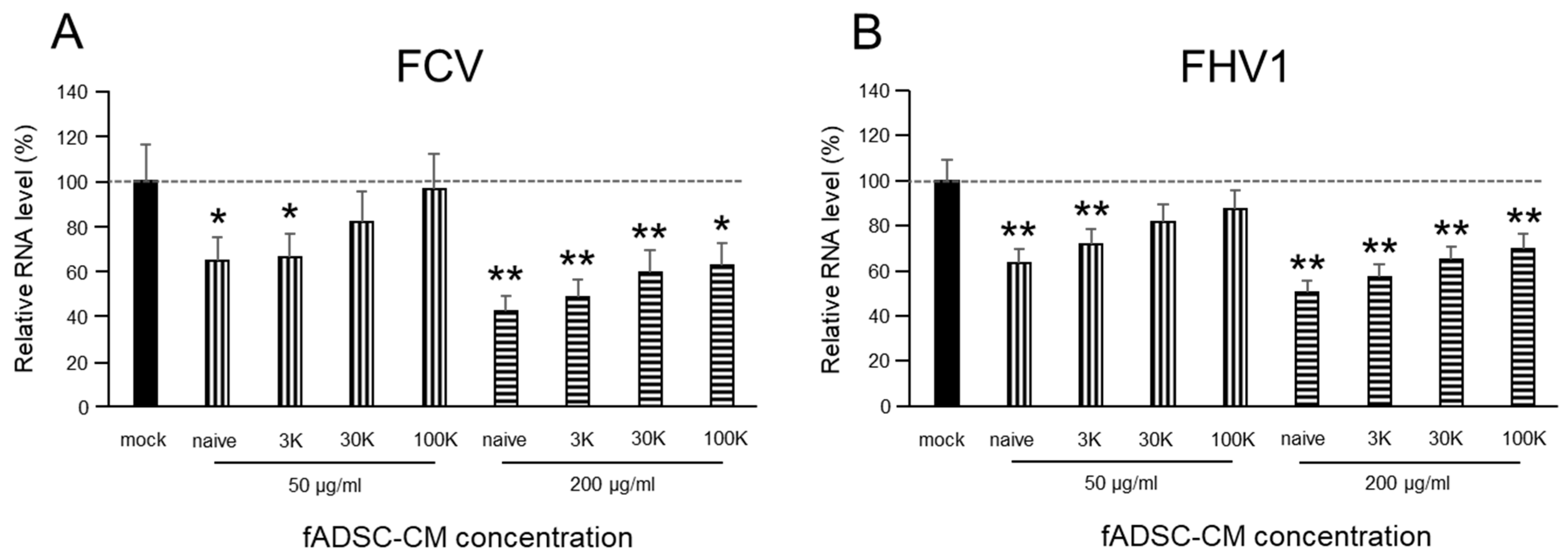
To evaluate whether fADSC-CM would affect the replication of FCV and FHV1 in CRFK cells, cells (2.0 × 104 cells/well) were incubated overnight in 24-well plates at 37 °C and 5% CO2 in the complete medium. The complete medium was removed, and the monolayer was infected with FCV and FHV1 at a multiplicity of infection (MOI) of 0.1 at 37 °C for 1 h in DMEM. After removing DMEM containing viruses and washing three times with PBS, the infected cells were incubated with fADSC-CM (50 and 200 µg/mL) for 24 h. After incubation, the total RNA was extracted from the cells, and the levels of vmRNA were determined by real-time reverse-transcription (RT) PCR.
2.7. Effect of fADSC-CM on Viral Entry
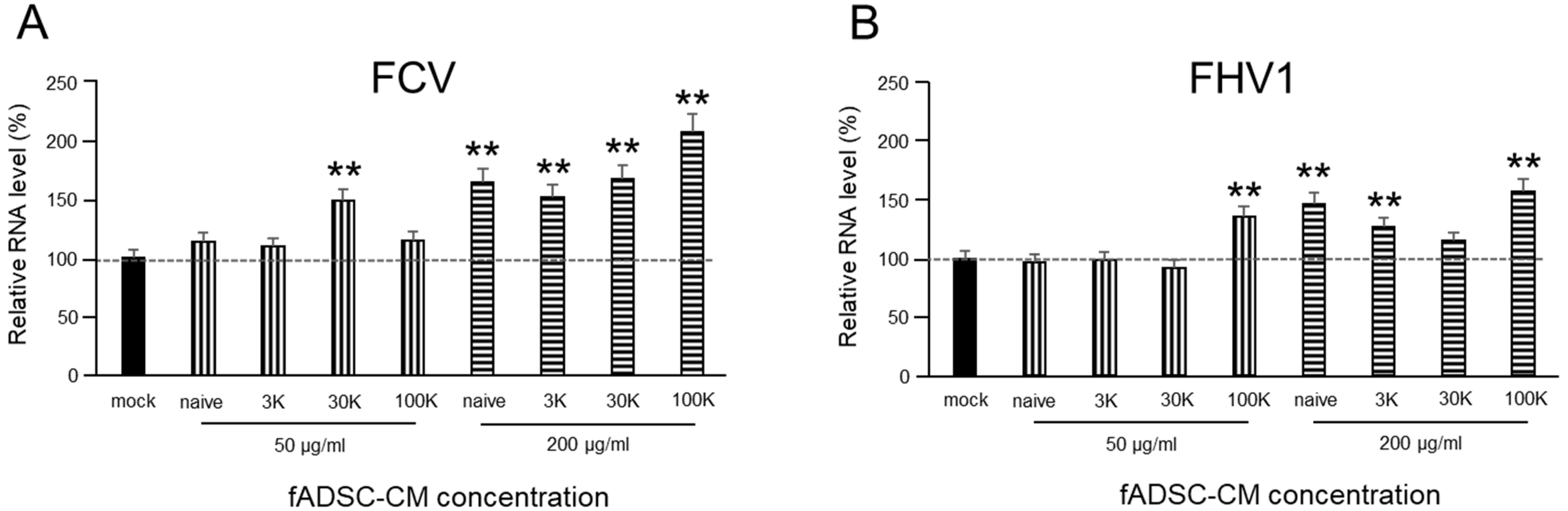
To evaluate whether fADSC-CM would affect the entry of FCV and FHV1 into CRFK cells, cells were incubated overnight in 24-well plates at 37 °C in 5% CO2 in the complete medium. The complete medium was removed, and the monolayer was infected at an MOI of 0.1 with FCV and FHV1 solutions containing different concentrations of fADSC-CM (50 µg/mL and 200 µg/mL) at 37 °C for 1 h. After removing the virus solutions and washing three times with PBS, the cells were incubated in the complete medium for an additional 1 h. After incubation, the total RNA was extracted from the cells to determine the levels of vmRNA by real-time RT-PCR.
2.8. Effects of Different Amounts of fADSC-CM on Viral Replication
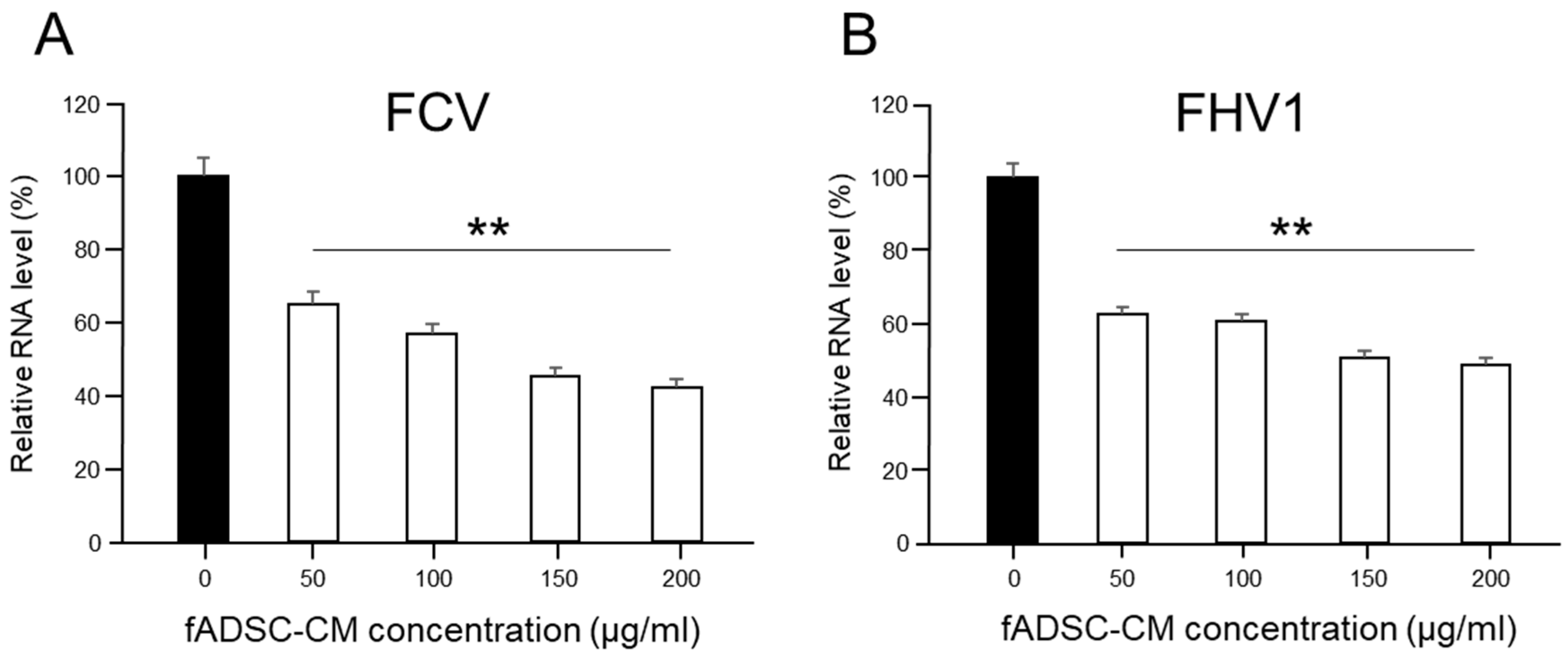
To determine whether the antiviral effect of fADSC-CM on the CRFK cells was dose-dependent, different concentrations of naïve fADSC-CM (50 µg/mL, 100 µg/mL, 150 µg/mL, and 200 µg/mL) were administrated after FCV and FHV1 infection. After incubation for 24 h at 37 °C in 5% CO2, the total RNA was extracted from the cells, and the levels of vmRNA were determined by real-time RT-PCR.
2.9. Effect of fADSC-CM on Different Phases of CRFK Cells under Infection Conditions
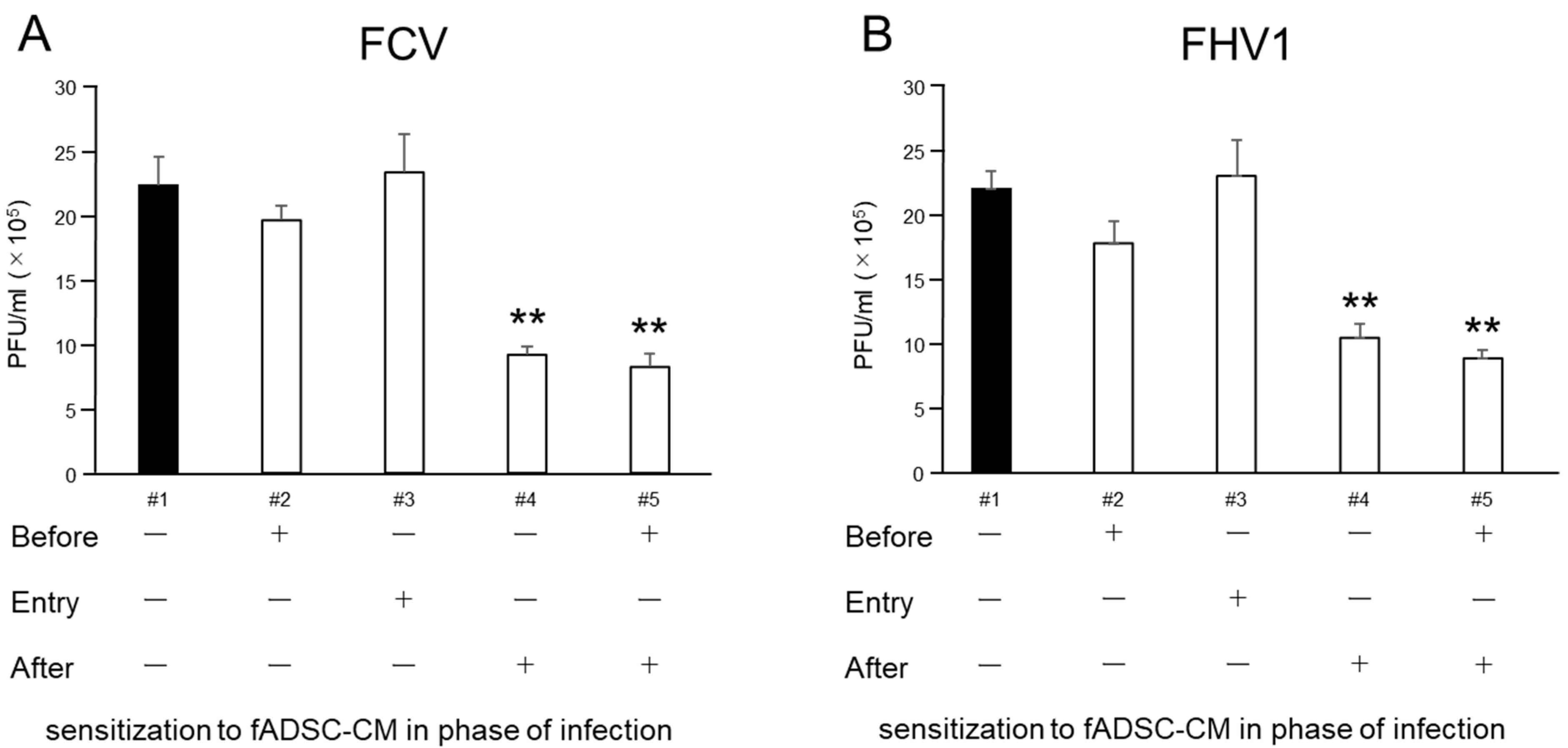
To compare the antiviral activity of fADSC-CM on the CRFK cells challenged with infection, the CRFK cells were exposed to naïve fADSC-CM before infection, at the time of infection, and/or after infection and incubated in 24-well plates or 12 h at 37 °C in the complete medium. For the before the infection group, the monolayer was incubated with 200 µg/mL of naïve fADSC-CM at 37 °C for 12 h, then infected with FCV and FHV1 at an MOI of 0.1 for 1 h. After infection, the CRFK cells were washed three times with PBS and incubated in 2% FCS DMEM for 24 h. For the group representing the duration of the infection, the monolayer was incubated in the complete medium for 12 h, then exposed to FCV and FHV1 solutions at an MOI of 0.1 that included 200 µg/mL of fADSC-CM for 1 h. For the after the viral infection group, the CRFK cells were washed three times with PBS, then incubated in 2% FCS DMEM for 24 h; then the monolayer was incubated with the complete medium for 12 h and the cells infected with FCV and FHV1 at an MOI of 0.1 for 1 h. After infection, the CRFK cells were washed three times in PBS and incubated in 200 µg/mL of naïve fADSC-CM for 24 h. After incubation for 24 h, the total RNA was extracted from the cells, and the levels of vmRNA were determined by real-time RT-PCR. After 24 h incubation of all the groups, cell supernatants were collected for using the plaque assay.
2.10. Plaque Assay
When the CRFK cells in the 10 cm-dish formed a monolayer, the cell supernatants were diluted serially 10-fold and added to the cells after removing the complete medium and washing with PBS. After incubating at 37 °C for 1 h, 2× DMEM and 3% carboxymethyl-cellulose were mixed at a 1:1 ratio and added to the plates, followed by culture at 37 °C for 36 h [
24]. The cells were fixed with 4% formaldehyde and stained with 1% crystal violet to calculate the number of plaques.
2.11. Real-Time RT-PCR
The total RNA from FCV- and FHV1-infected CRFK cells was isolated using NucleoSpin RNA (Takara) according to the manufacturer’s instructions. cDNA was synthesized from 0.5 µg of the total RNA using random primers and the GoScript Reverse Transcriptase system (Promega), according to the manufacturer’s instructions. Real-time RT-PCR analyses were performed using GoTaq Probe qPCR Master Mix (Promega) to determine the mRNA levels of FCV strain F4 and the glycoprotein C gene for FHV1 strain F7301. mRNA levels of the housekeeping gene β-glucuronidase (
GUSB) were used for normalization. The primers and probe sequences are shown in
Table 1 [
25,
26,
27]. Amplification conditions were 95 °C for 2 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 60 s. After 40 cycles, a dissociation curve was generated to verify the specificity of each primer. All reactions were performed in duplicate. Expression levels of target genes were normalized to the level of GUSB and quantified by the ΔΔCt method.
2.12. RNA Sequencing
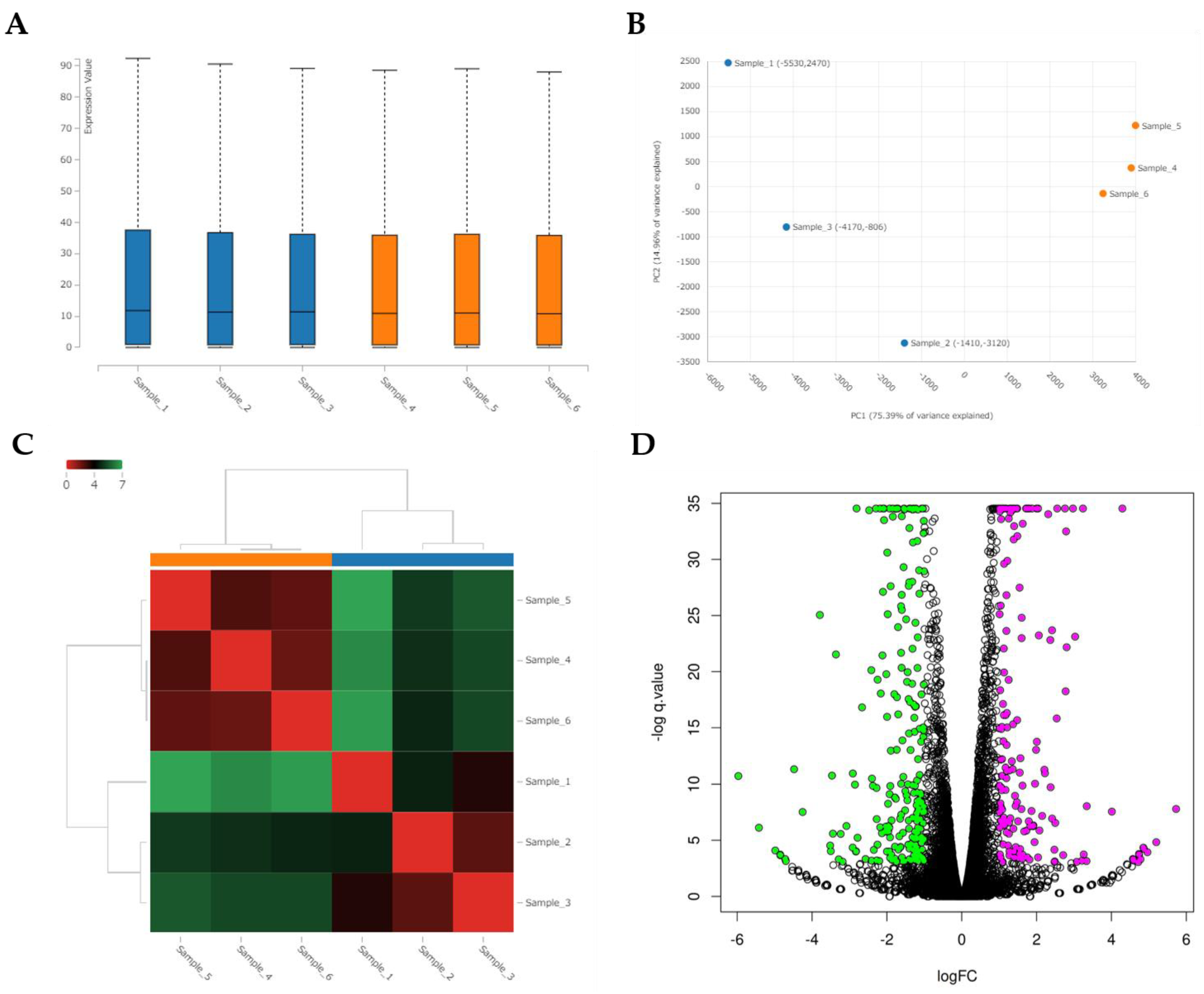
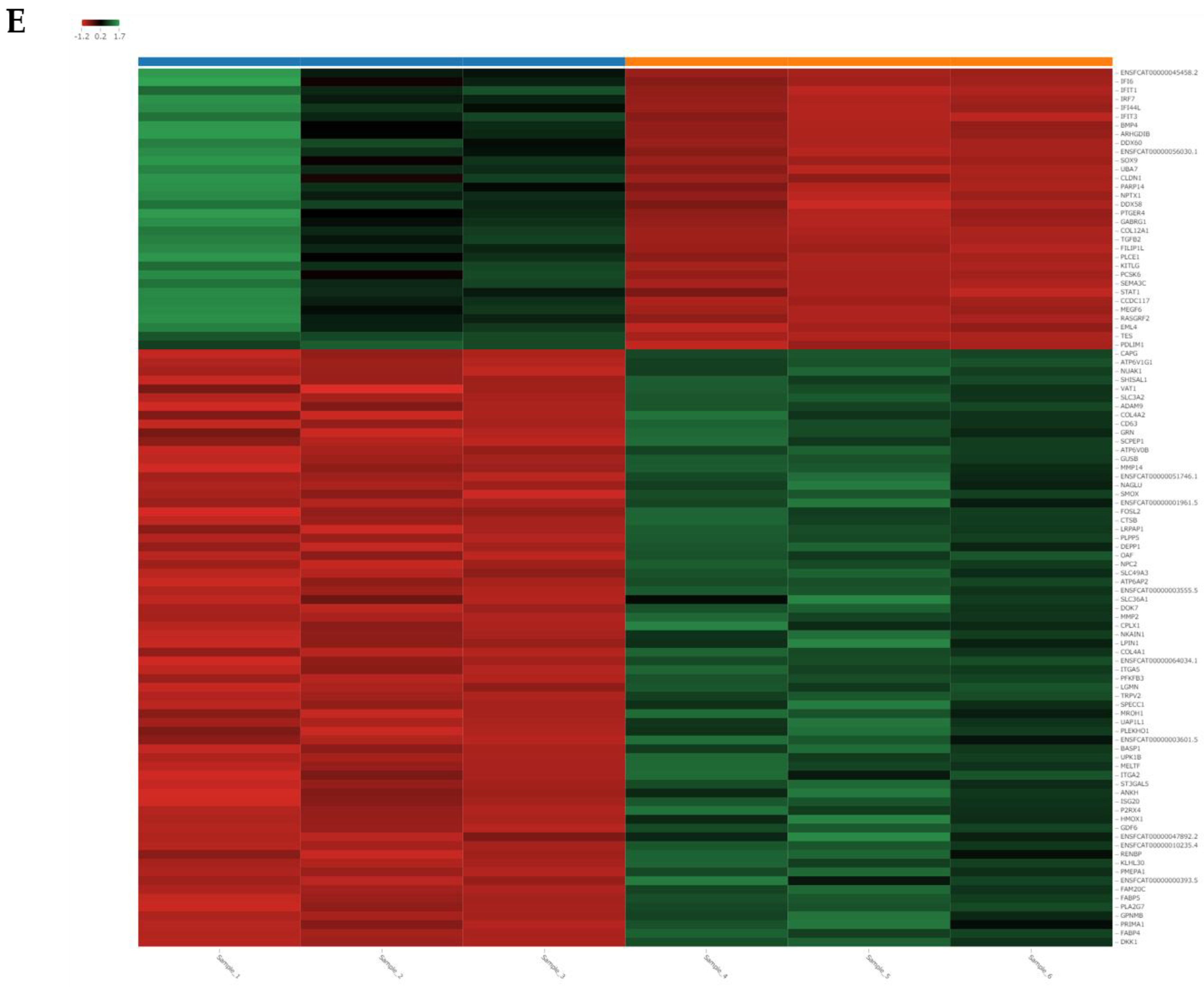
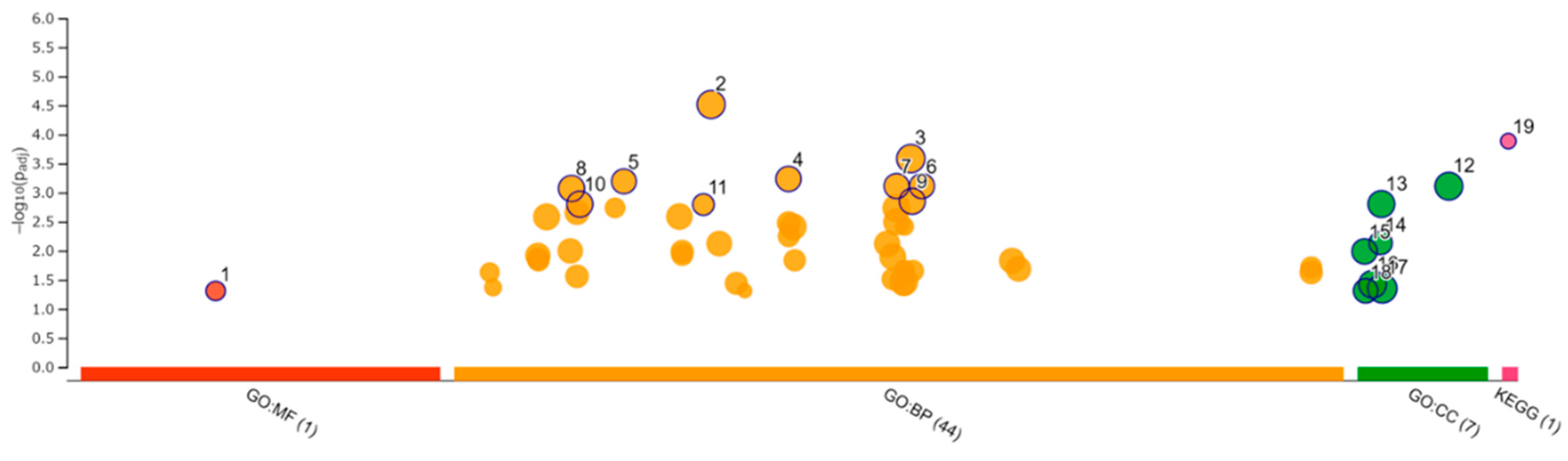
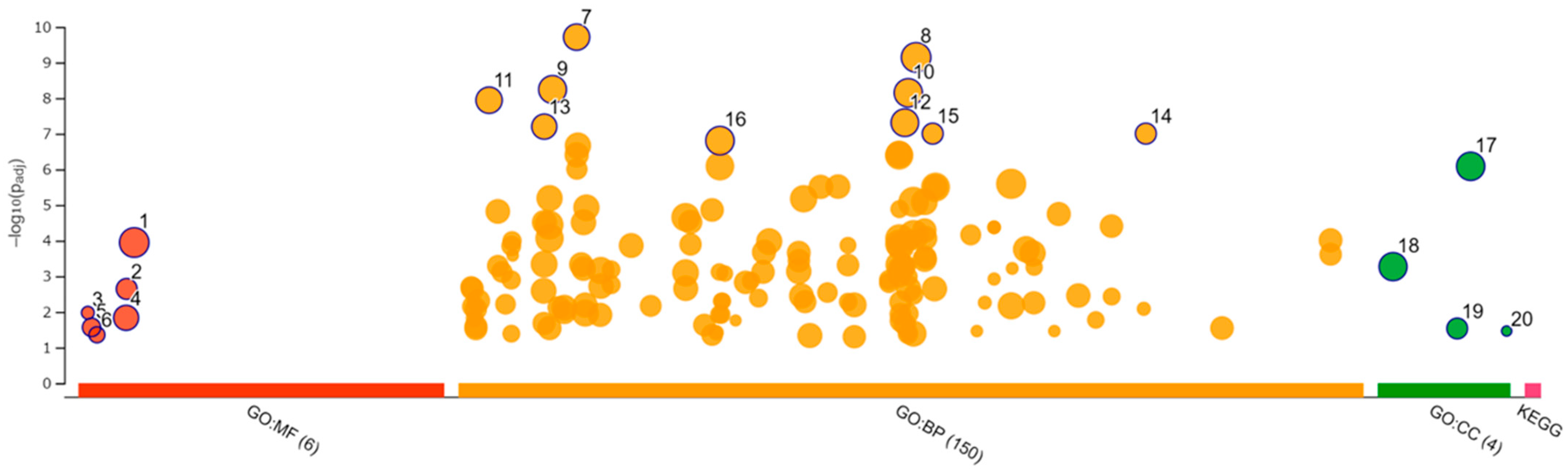
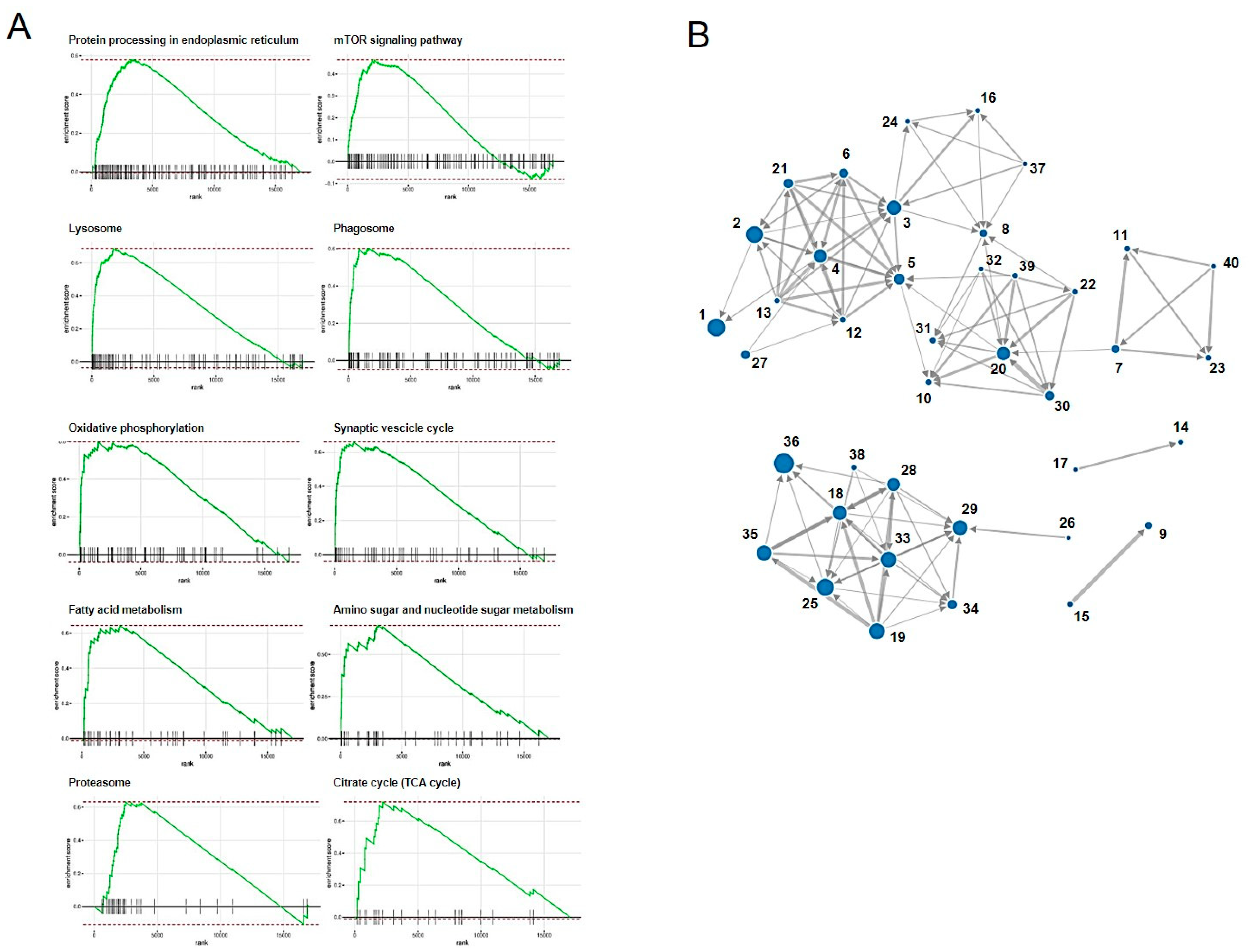
We performed RNA-seq of the total RNA samples isolated from the CRFK cells after 12 h of incubation in the complete medium (control, n = 3) and with 200 µg/mL of naïve fADSC-CM (n = 3). cDNA library construction was carried out with 1 µg of the total RNA using NEBNext Ultra II RNA Library Prep Kit for Illumina, according to the manufacturer’s instructions, followed by paired-end sequencing (2 × 150 bp) using the Novaseq6000. For each library, an average of 16–20 million read pairs were generated. Quality control checks of the sequencing raw data were conducted with FastQC, while adapter-trimming was performed with Trim Galore. The relative expression of transcripts was quantified for each sample using featureCount. Fastq files were mapped to the Felis_Catus_9.0 reference genome using STAR software (ver2.7). Differentially expressed genes (upregulated or downregulated genes) were determined using edgeR (ver. 3.22.3) based on an adjusted p-value of <0.05 and fold change of >2 or <0.5. Biological function Gene Ontology (GO) analysis was performed by g:Profiler, and gene set enrichment analysis (GSEA) was performed by RaNA-seq.
2.13. Statistical Analysis
All experiments were performed in triplicate and repeated in three independent experiments except for RNA-seq, and all data are presented as the mean ± standard deviation. Differences among multiple groups were assessed by one-way analysis of variance, and differences were compared using the Tukey–Kramer post hoc test. p < 0.05 was considered statistically significant. Statistical analyses were performed using Excel 2019 with add-in software Statcel 3.
4. Discussion
MSCs and/or their secretomes have been investigated as methods of therapy used in several animal models of viral disease [
9]. These studies have demonstrated that MSC treatment effectively suppresses inflammation and enhances immunity. Moreover, some studies have shown that microRNAs (miRNAs), including those in exosomes secreted by MSCs, directly inhibit viral expression and replication [
28,
29]. In the veterinary field, FCV and FHV1 are associated with the onset and severity of feline chronic gingivostomatitis and upper respiratory infections, and some studies have shown the usefulness of MSCs for therapy against these conditions [
21,
22,
23]. These studies primarily discussed improving clinical symptoms dampening inflammation and did not examine the effects on the viruses themselves. Therefore, we investigated the effects of MSC secretome at different protein concentrations and molecular weights on FCV and FHV1 in this study.
Most studies have focused on the antiviral effects of miRNAs contained in exosomes secreted from MSCs [
9,
28,
29,
30], but the antiviral effects of MSC secretomes have not been investigated. Therefore, naïve feline AD-MSC secretome were collected as CM in this study. Based on the protein concentrations of the naïve fADSC-CM, the highest concentration used in the comparison experiments was 200 µg/mL. Several methods of purifying exosomes from CM have been published, and the use of MWCO ultrafiltration has been reported to increase exosome recovery [
7,
31,
32]. In this study, vmRNA expression in the CRFK cells sensitized to naïve fADSC-CM showed the greatest inhibition of both FCV and FHV1 compared with the expression in the CRFK cells sensitized to 3 kDa, 30 kDa, and 100 kDa MWCO-concentrated fADSC-CM. We performed our experiments based on naïve fADSC-CM. Therefore, it is possible that the number of exosomes was not so different from that in naïve fADSC-CM. From the results of the dose-dependent suppression of viral replication, therefore, FCV and FHV1 may also be inhibited by exosomes secreted from fAD-MSCs. To clarify the antiviral effects of feline AD-MSCs exosomes on FCV and FHV1, further comparative experiments using purified feline AD-MSCs exosomes and CM excluded exosomes are needed.
The results of sensitizing cells to CM during different phases of the viral infection process showed that CM did not inhibit viral entry but did inhibit both FCV and FHV1 replication. These effects were similar to the antiviral effects of exosomal miRNAs demonstrated on hepatitis C virus, which showed inhibited viral replication after infection of the cells [
28]. In both FCV and FHV1, vmRNA expressions were increased in the CRFK cells sensitized at the virus attachment and entry phases. Feline junctional adhesion molecule (JAM)-1, an immunoglobulin-like protein present in tight junctions, was identified as a cellular-binding molecule of the FCV F4 strain [
33]. In our RNA-seq results, we detected no
JAM-1 expression in the CRFK cells, but there was no difference in
JAM-2 or
JAM-3 expression between the mock CRFK cells and the CRFK cells sensitized with fADSC-CM. In FHV1, envelope glycoproteins play important roles in cell attachment and entry, and heparin sulfate on the host cell surface serves as a receptor [
34,
35]. RNA-seq of the CRFK cells detected some heparin-sulfate-related genes, but there were no changes in the expression of these in the CRFK cells sensitized with fADSC-CM compared with the expression of these in mock CRFK cells. It is unclear why FCV and FHV1 entry into the CRFK cells was facilitated, but we speculated that factors other than host cell receptors may have been involved.
GSEA showed that fADSC-CM altered various functions of CRFK cells, such as metabolic pathways, signaling pathways, the cell cycle, the lysosome, oxidative phosphorylation, and the phagosome. Although RNA-seq did not determine the mechanisms involved in the inhibition of viral replication, the observation that fADSC-CM inhibited the viral replication of two different type of viruses, FCV, which is a single-positive-stranded RNA non-enveloped virus, and FHV1, which is a double-stranded DNA enveloped virus, was very interesting.
In conclusion, our findings revealed that fADSC-CM had antiviral effects by inhibiting FCV and FHV1 replication. It is suggested that AD-MSC therapy for feline chronic gingivostomatitis and upper respiratory infection may be effective due not only to the anti-inflammatory effects, as already reported, but also effects that prevent viral replication. Further studies are needed to clarify the mechanisms by which fADSC-CM prevents viral replication.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}