
A Proteomic Approach to Study the Biological Role of Hepatitis C Virus Protein Core+1/ARFP

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Sample Preparation for MS Analysis
2.3. LC-MS/MS Analysis
2.4. MS Data Processing and Quantification
2.5. MS Data Analysis
2.6. Functional Network Enrichment Analysis
2.7. GSEA Analysis
2.8. Bibliographic Searches
2.9. Visualisation of Proteomics Data
2.10. Cellular Pathway Analysis Using KEGG Annotations
2.11. RNA Isolation, cDNA Synthesis, and qPCR
2.12. Wound-Healing Assay
3. Results
3.1. Proteomic Analysis of the Huh 7.5/ARFP Expressing Cell Lines
3.2. The Expression of Core+1/ARFP Changes the Proteome of the Huh7.5 Cells
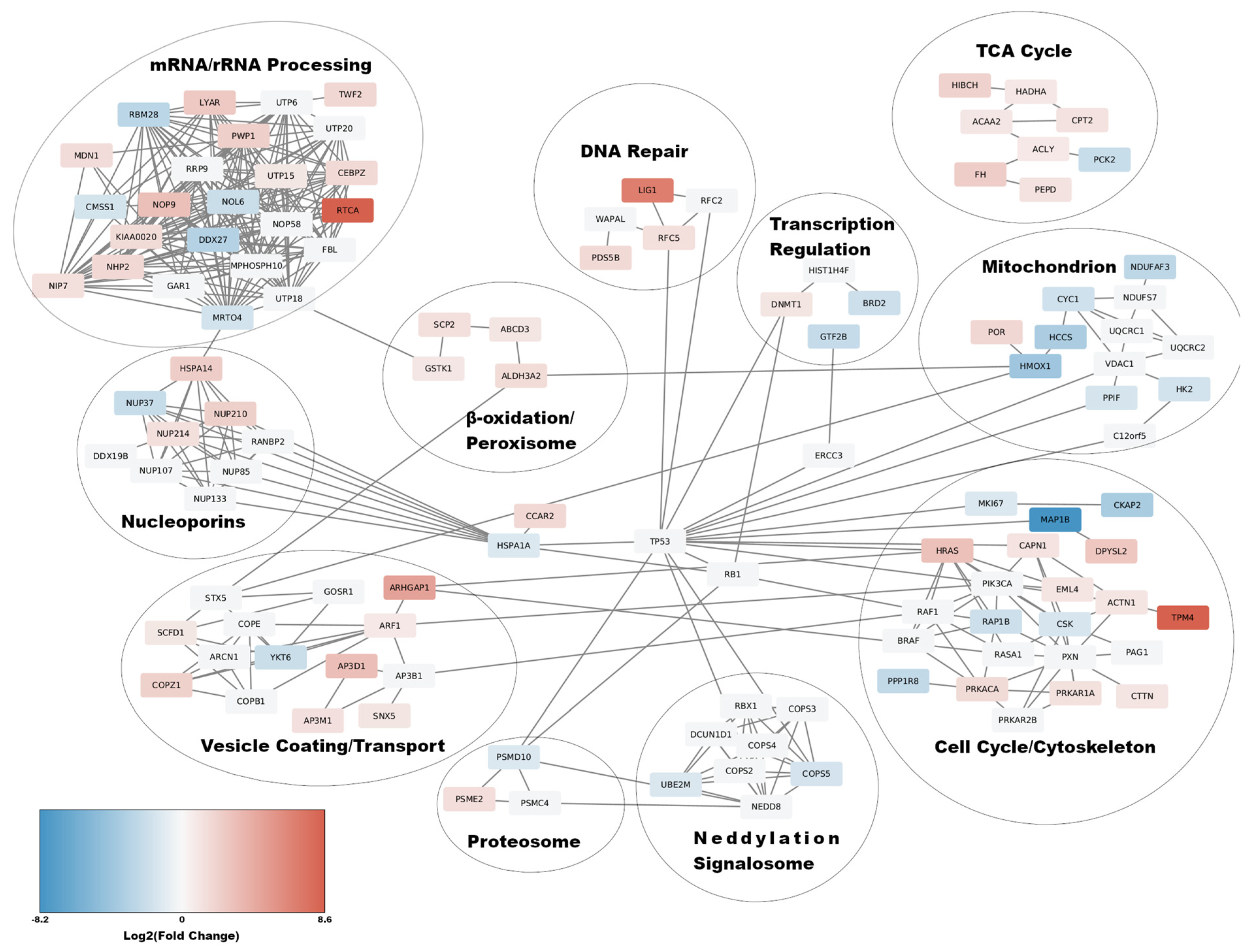
3.3. Functional Network Enrichment Analysis Using the STRINGDB
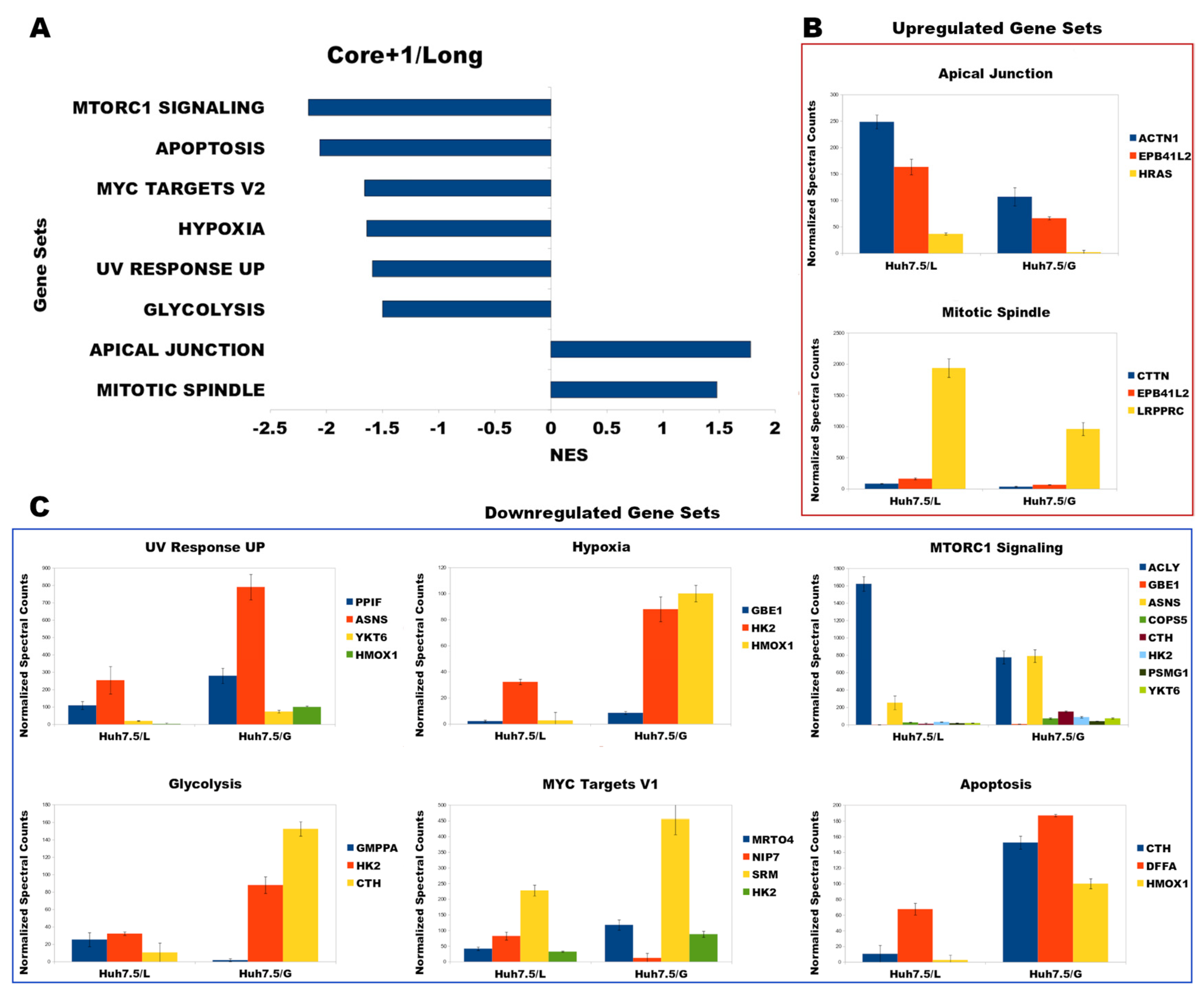
3.4. Functional Analysis Using Gene Set Enrichment Analysis (GSEA)
3.5. Correlation of GSEA Results with Liver Pathology
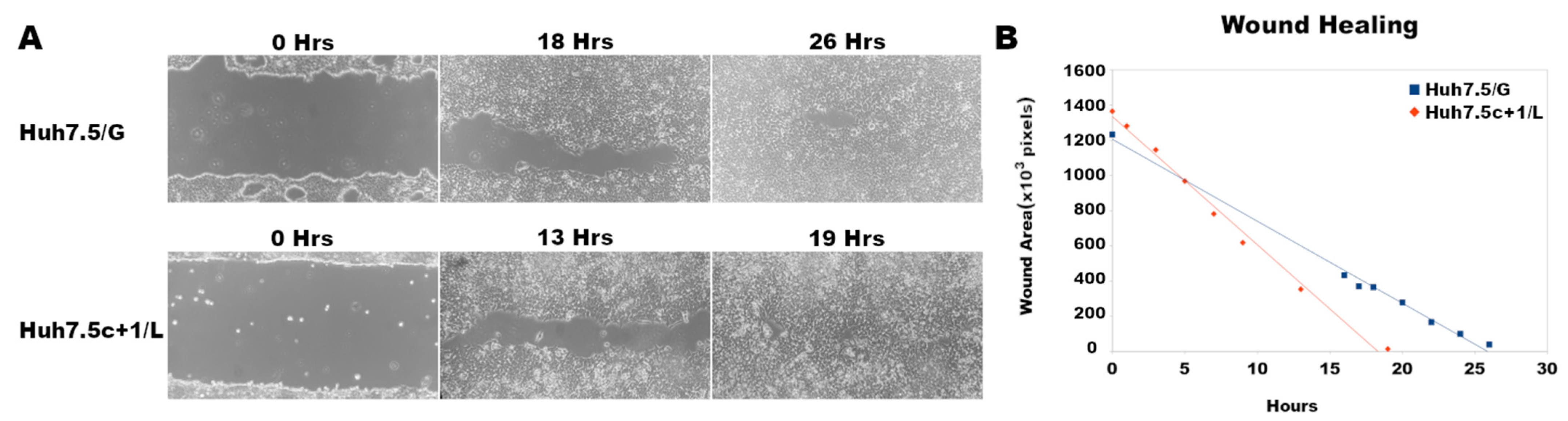
3.6. Core+1/Long Stable Expression in Huh7.5 Cells Increases Cell Migration
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pietschmann, T.; Brown, R.J.P. Hepatitis C Virus. Trends Microbiol. 2019, 27, 379–380. [Google Scholar] [CrossRef]
- World Health Organization. Global Hepatitis Report; World Health Organization: Geneve, Switzerland, 2017.
- Lin, M.V.; King, L.Y.; Chung, R.T. Hepatitis C virus-associated cancer. Annu. Rev. Pathol. 2015, 10, 345–370. [Google Scholar] [CrossRef] [PubMed]
- Mesri, E.A.; Feitelson, M.A.; Munger, K. Human viral oncogenesis: A cancer hallmarks analysis. Cell Host Microbe 2014, 15, 266–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burlone, M.E.; Budkowska, A. Hepatitis C virus cell entry: Role of lipoproteins and cellular receptors. J. Gen. Virol. 2009, 90, 1055–1070. [Google Scholar] [CrossRef]
- Blight, K.J.; Kolykhalov, A.A.; Rice, C.M. Efficient initiation of HCV RNA replication in cell culture. Science 2000, 290, 1972–1974. [Google Scholar] [CrossRef]
- Moradpour, D.; Penin, F. Hepatitis C virus proteins: From structure to function. Curr. Top. Microbiol. Immunol. 2013, 369, 113–142. [Google Scholar] [CrossRef] [PubMed]
- Bartenschlager, R.; Penin, F.; Lohmann, V.; André, P. Assembly of infectious hepatitis C virus particles. Trends Microbiol. 2011, 19, 95–103. [Google Scholar] [CrossRef]
- Alvisi, G.; Madan, V.; Bartenschlager, R. Hepatitis C virus and host cell lipids: An intimate connection. RNA Biol. 2011, 8, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Shimotohno, K. HCV Assembly and Egress via Modifications in Host Lipid Metabolic Systems. Cold Spring Harb. Perspect. Med. 2021, 11, a036814. [Google Scholar] [CrossRef]
- Walsh, D.; Mathews, M.B.; Mohr, I. Tinkering with translation: Protein synthesis in virus–infected cells. Cold Spring Harb. Perspect. Biol. 2013, 5, a012351. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, Z.A.; Kieft, J.S. Viral RNA structure-based strategies to manipulate translation. Nat. Rev. Microbiol. 2019, 17, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Mohamadi, M.; Azarbayjani, K.; Mozhgani, S.H.; Bamdad, T.; Alamdary, A.; Nikoo, H.R.; Hashempour, T.; Hedayat Yaghoobi, M.; Ajorloo, M. Hepatitis C virus alternative reading frame protein (ARFP): Production, features, and pathogenesis. J. Med. Virol. 2020, 92, 2930–2937. [Google Scholar] [CrossRef] [PubMed]
- Branch, A.D.; Stump, D.D.; Gutierrez, J.A.; Eng, F.; Walewski, J.L. The hepatitis C virus alternate reading frame (ARF) and its family of novel products: The alternate reading frame protein/F-protein, the double-frameshift protein, and others. Semin. Liver. Dis. 2005, 25, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Varaklioti, A.; Vassilaki, N.; Georgopoulou, U.; Mavromara, P. Alternate translation occurs within the core coding region of the hepatitis C viral genome. J. Biol. Chem. 2002, 277, 17713–17721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Choi, J.; Yen, T.S.B.; Lu, W.; Strohecker, A.; Govindarajan, S.; Chien, D.; Selby, M.J.; Ou, J.H. Synthesis of a novel hepatitis C virus protein by ribosomal frameshift. EMBO J. 2001, 20, 3840–3848. [Google Scholar] [CrossRef] [Green Version]
- Walewski, J.L.; Keller, T.R.; Stump, D.D.; Branch, A.D. Evidence for a new hepatitis C virus antigen encoded in an overlapping reading frame. RNA 2001, 7, 710–721. [Google Scholar] [CrossRef] [Green Version]
- Vassilaki, N.; Mavromara, P. The HCV ARFP/F/core+1 protein: Production and functional analysis of an unconventional viral product. IUBMB Life 2009, 61, 739–752. [Google Scholar] [CrossRef]
- Vassilaki, N.; Mavromara, P. Two alternative translation mechanisms are responsible for the expression of the HCV ARFP/F/core+1 coding open reading frame. J. Biol. Chem. 2003, 278, 40503–40513. [Google Scholar] [CrossRef] [Green Version]
- Baril, M.; Brakier-Gingras, L. Translation of the F protein of hepatitis C virus is initiated at a non-AUG codon in a +1 reading frame relative to the polyprotein. Nucleic Acids Res. 2005, 33, 1474–1486. [Google Scholar] [CrossRef] [Green Version]
- Vassilaki, N.; Friebe, P.; Meuleman, P.; Kallis, S.; Kaul, A.; Paranhos-Baccalà, G.; Leroux-Roels, G.; Mavromara, P.; Bartenschlager, R. Role of the hepatitis C virus core+1 open reading frame and core cis-acting RNA elements in viral RNA translation and replication. J. Virol. 2008, 82, 11503–11515. [Google Scholar] [CrossRef] [Green Version]
- Kotta-Loizou, I.; Karakasiliotis, I.; Vassilaki, N.; Sakellariou, P.; Bartenschlager, R.; Mavromara, P. Expression of the novel hepatitis C virus core+1/ARF protein in the context of JFH1-based replicons. J. Virol. 2015, 89, 5164–5170. [Google Scholar] [CrossRef] [Green Version]
- Kassela, K.; Karakasiliotis, I.; Charpantidis, S.; Koskinas, J.; Mylopoulou, T.; Mimidis, K.; Sarrazin, C.; Grammatikos, G.; Mavromara, P. High prevalence of antibodies to core+1/ARF protein in HCV-infected patients with advanced cirrhosis. J. Gen. Virol. 2017, 98, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Ajorloo, M.; Bamdad, T.; Hashempour, T.; Alborzi, A.M.; Mozhgani, S.H.; Asadi, R.; Haj-sheykholeslami, A.; Merat, S. Detection of Specific Antibodies to HCV-ARF/CORE+1 Protein in Cirrhotic and Non-Cirrhotic Patients with Hepatitis C: A Possible Association with Progressive Fibrosis. Arch. Iran. Med. 2015, 18, 304–307. [Google Scholar] [PubMed]
- Dalagiorgou, G.; Vassilaki, N.; Foka, P.; Boumlic, A.; Kakkanas, A.; Kochlios, E.; Khalili, S.; Aslanoglou, E.; Veletza, S.; Orfanoudakis, G.; et al. High levels of HCV core+1 antibodies in HCV patients with hepatocellular carcinoma. J. Gen. Virol. 2011, 92, 1343–1351. [Google Scholar] [CrossRef]
- Karamitros, T.; Kakkanas, A.; Katsoulidou, A.; Sypsa, V.; Dalagiorgou, G.; Mavromara, P.; Hatzakis, A. Detection of specific antibodies to HCV-ARF/CORE+1 protein in patients treated with pegylated interferon plus ribavirin. J. Viral Hepat. 2012, 19, 182–188. [Google Scholar] [CrossRef] [PubMed]
- McMullan, L.K.; Grakoui, A.; Evans, M.J.; Mihalik, K.; Puig, M.; Branch, A.D.; Feinstone, S.M.; Rice, C.M. Evidence for a functional RNA element in the hepatitis C virus core gene. Proc. Natl. Acad. Sci. USA 2007, 104, 2879–2884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, J.; Eng, F.J.; Branch, A.D. HCV and HCC: Clinical update and a review of HCC-associated viral mutations in the core gene. Semin. Liver. Dis. 2011, 31, 347–355. [Google Scholar] [CrossRef]
- Sobesky, R.; Feray, C.; Rimlinger, F.; Derian, N.; Dos Santos, A.; Roque-Afonso, A.M.; Samuel, D.; Bréchot, C.; Thiers, V. Distinct hepatitis C virus core and F protein quasispecies in tumoral and nontumoral hepatocytes isolated via microdissection. Hepatology 2007, 46, 1704–1712. [Google Scholar] [CrossRef]
- Samrat, S.K.; Li, W.; Singh, S.; Kumar, R.; Agrawal, B. Alternate Reading Frame Protein (F Protein) of Hepatitis C Virus: Paradoxical Effects of Activation and Apoptosis on Human Dendritic Cells Lead to Stimulation of T Cells. PLoS ONE 2014, 9, e86567. [Google Scholar] [CrossRef] [PubMed]
- Fiorucci, M.; Boulant, S.; Fournillier, A.; Abraham, J.D.; Lavergne, J.P.; Paranhos-Baccala, G.; Inchauspé, G.; Bain, C. Expression of the alternative reading frame protein of Hepatitis C virus induces cytokines involved in hepatic injuries. J. Gen. Virol. 2007, 88, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, X.; Deng, X.; Yue, M.; Zhang, J.; Zhu, D.; Zhou, Z.; Zhai, X.; Xu, K.; Zhang, Y. Hepatitis C virus alternate reading frame protein decreases interferon-α secretion in peripheral blood mononuclear cells. Mol. Med. Rep. 2014, 9, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Seronello, S.; Mayer, W.; Ojcius, D.M. Hepatitis C Virus Frameshift/Alternate Reading Frame Protein Suppresses Interferon Responses Mediated by Pattern Recognition Receptor Retinoic-Acid-Inducible Gene-I. PLoS ONE 2016, 11, e0158419. [Google Scholar] [CrossRef]
- Mylopoulou, T.; Papadopoulos, V.; Kassela, K.; Karakasiliotis, I.; Souvalidou, F.; Mimidis, P.; Veletza, S.; Mavromara, P.; Mimidis, K. Relationship between antibodies to hepatitis C virus core+1 protein and treatment outcome. Ann. Gastroenterol. 2018, 31, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Douam, F.; PLoSs, A. Proteomic approaches to analyzing hepatitis C virus biology. Proteomics 2015, 15, 2051–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBride, A.A. The Promise of Proteomics in the Study of Oncogenic Viruses. Mol. Cell. Proteomics 2017, 16, S65–S74. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, S.; Karakasiliotis, I.; Mavromara, P. Hepatitis C Virus core+1/ARF Protein Modulates the Cyclin D1/pRb Pathway and Promotes Carcinogenesis. J. Virol. 2018, 92, e02036-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, P.E.; Efentakis, P.; Abu Qourah, F.; Femminò, S.; Makridakis, M.; Kanaki, Z.; Varela, A.; Tsoumani, M.; Davos, C.H.; Dimitriou, C.A.; et al. Chronic Empagliflozin Treatment Reduces Myocardial Infarct Size in Nondiabetic Mice Through STAT-3-Mediated Protection on Microvascular Endothelial Cells and Reduction of Oxidative Stress. Antioxid. Redox Signal. 2021, 34, 551–571. [Google Scholar] [CrossRef] [PubMed]
- Aguilan, J.T.; Kulej, K.; Sidoli, S. Guide for protein fold change and p-value calculation for non-experts in proteomics. Mol. Omics 2020, 16, 573–582. [Google Scholar] [CrossRef]
- Levin, Y. The role of statistical power analysis in quantitative proteomics. PROTEOMICS 2011, 11, 2565–2567. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Jonkman, J.E.N.; Cathcart, J.A.; Xu, F.; Bartolini, M.E.; Amon, J.E.; Stevens, K.M.; Colarusso, P. An introduction to the wound healing assay using live-cell microscopy. Cell Adh. Migr. 2014, 8, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbriano, C.; Bolognese, F.; Gurtner, A.; Piaggio, G.; Mantovani, R. HSP-CBF is an NF-Y-dependent coactivator of the heat shock promoters CCAAT boxes. J. Biol. Chem. 2001, 276, 26332–26339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, M.; Ebstein, F.; Bürger, E.; Textoris-Taube, K.; Gorny, X.; Urban, S.; Zhao, F.; Dannenberg, T.; Sucker, A. The proteasome immunosubunits, PA28 and ER-aminopeptidase 1 protect melanoma cells from efficient MART-126-35 -specific T-cell recognition. Eur. J. Immunol. 2015, 45, 3257–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, P.; Freese, K.; Mahli, A.; Thasler, W.E.; Hellerbrand, C.; Bosserhoff, A.K. Combined effects of PLK1 and RAS in hepatocellular carcinoma reveal rigosertib as promising novel therapeutic “dual-hit” option. Oncotarget 2018, 9, 3605–3618. [Google Scholar] [CrossRef] [Green Version]
- Naydenov, N.G.; Joshi, S.; Feygin, A.; Saini, S.; Litovchick, L.; Ivanov, A.I. A membrane fusion protein, Ykt6, regulates epithelial cell migration via microRNA-mediated suppression of Junctional Adhesion Molecule A. Cell Cycle 2018, 17, 1812–1831. [Google Scholar] [CrossRef]
- Konishi, S.; Ishiguro, H.; Shibata, Y.; Kudo, J.; Terashita, Y.; Sugiura, H.; Koyama, H.; Kimura, M.; Sato, A.; Shinoda, N.; et al. Decreased expression of DFF45/ICAD is correlated with a poor prognosis in patients with esophageal carcinoma. Cancer 2002, 95, 2473–2478. [Google Scholar] [CrossRef] [PubMed]
- Stock, K.; Borrink, R.; Mikesch, J.H.; Hansmeier, A.; Rehkämper, J.; Trautmann, M.; Wardelmann, E.; Hartmann, W.; Sperveslage, J.; Steinestel, K. Overexpression and Tyr421-phosphorylation of cortactin is induced by three-dimensional spheroid culturing and contributes to migration and invasion of pancreatic ductal adenocarcinoma (PDAC) cells. Cancer Cell Int. 2019, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Du, H.; Shao, S.; Bo, T.; Yu, C.; Chen, W.; Zhao, L.; Li, Q.; Wang, L.; Liu, X.; et al. Cyclophilin D deficiency attenuates mitochondrial perturbation and ameliorates hepatic steatosis. Hepatology 2018, 68, 62–77. [Google Scholar] [CrossRef] [PubMed]
- AbdulHameed, M.D.; Tawa, G.J.; Kumar, K.; Ippolito, D.L.; Lewis, J.A.; Stallings, J.D.; Wallqvist, A. Systems level analysis and identification of pathways and networks associated with liver fibrosis. PLoS ONE 2014, 9, e112193. [Google Scholar] [CrossRef]
- Tsui, T.Y.; Lau, C.K.; Ma, J.; Fan, S.T.; Glockzin, G.; Obed, A.; Schlitt, H.J. Adeno-associated virus-mediated heme oxygenase-1 gene transfer suppresses the progression of micronodular cirrhosis in rats. World J. Gastroenterol. 2006, 12, 2016–2023. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef]
- Franks, A.; Airoldi, E.; Slavov, N. Post-transcriptional regulation across human tissues. PLoS Comput. Biol. 2017, 13, e1005535. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.Y.; Suthar, M.S. Mechanisms of innate immune evasion in re-emerging RNA viruses. Curr. Opin. Virol. 2015, 12, 26–37. [Google Scholar] [CrossRef]
- Yarbrough, M.L.; Mata, M.A.; Sakthivel, R.; Fontoura, B.M.A. Viral subversion of nucleocytoplasmic trafficking. Traffic 2014, 15, 127–140. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, Y.E.; Palazzo, A.F. Crosstalk between nucleocytoplasmic trafficking and the innate immune response to viral infection. J. Biol. Chem. 2021, 297, 297–298. [Google Scholar] [CrossRef]
- Bonamassa, B.; Ciccarese, F.; Di Antonio, V.; Contarini, A.; Palù, G.; Alvisi, G. Hepatitis C virus and host cell nuclear transport machinery: A clandestine affair. Front. Microbiol. 2015, 6, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Liu, Y.; Feng, W.; Lei, L.; Du, Y.; Wu, J.; Wang, S. NUP37, a positive regulator of YAP/TEAD signaling, promotes the progression of hepatocellular carcinoma. Oncotarget 2017, 8, 98004–98013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Wang, T.; Wang, F.; Hu, X.; Zhan, G.; Jin, X.; Zhang, L.; Li, Y. NUP37 silencing induces inhibition of cell proliferation, G1 phase cell cycle arrest and apoptosis in non–small cell lung cancer cells. Pathol. Res. Pract. 2020, 216, 152836. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Cigdem, S.; Okuwaki, M.; Nagata, K. Leukemia–Associated Nup214 Fusion Proteins Disturb the XPO1–Mediated Nuclear–Cytoplasmic Transport Pathway and Thereby the NF–κB Signaling Pathway. Mol. Cell. Biol. 2016, 36, 1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, M.; Rebensburg, S.V.; Takata, M.A.; Zang, T.M.; Yamashita, M.; Kvaratskhelia, M.; Bieniasz, P.D. Nuclear pore heterogeneity influences HIV-1 infection and the antiviral activity of MX2. eLife 2018, 7, e35738. [Google Scholar] [CrossRef]
- Gupta, P.; Sharma, A.; Han, J.; Yang, A.; Bhomia, M.; Knollmann-Ritschel, B.; Puri, R.K.; Maheshwari, R.K. Differential host gene responses from infection with neurovirulent and partially-neurovirulent strains of Venezuelan equine encephalitis virus. BMC Infect. Dis. 2017, 17, 309. [Google Scholar] [CrossRef] [Green Version]
- Versteeg, K.; Menicucci, A.R.; Woolsey, C.; Mire, C.E.; Geisbert, J.B.; Cross, R.W.; Agans, K.N.; Jeske, D.; Messaoudi, I.; Geisbert, T.W. Infection with the Makona variant results in a delayed and distinct host immune response compared to previous Ebola virus variants. Sci. Rep. 2019, 7, 9730. [Google Scholar] [CrossRef]
- Lam, Y.W.; Evans, V.C.; Heesom, K.J.; Lamond, A.I.; Matthews, D.A. Proteomics analysis of the nucleolus in adenovirus-infected cells. Mol. Cell. Proteom. 2010, 9, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Chiu, H.C.; Hannemann, H.; Heesom, K.J.; Matthews, D.A.; Davidson, A.D. High-Throughput Quantitative Proteomic Analysis of Dengue Virus Type 2 Infected A549 Cells. PLoS ONE 2014, 9, e93305. [Google Scholar] [CrossRef]
- Gerresheim, G.K.; Roeb, E.; Michel, A.M.; Niepmann, M. Hepatitis C Virus Downregulates Core Subunits of Oxidative Phosphorylation, Reminiscent of the Warburg Effect in Cancer Cells. Cells 2019, 8, 1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, D.N.; Pu, C.H.; Lewis, J.T.; Bhat, R.; Anwar-Mohamed, A.; Logan, M.; Lund, G.; Addison, W.R.; Lehner, R.; Kneteman, N.M. Oxidative Stress Attenuates Lipid Synthesis and Increases Mitochondrial Fatty Acid Oxidation in Hepatoma Cells Infected with Hepatitis C Virus. J. Biol. Chem. 2016, 291, 1974–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassilaki, N.; Boleti, H.; Mavromara, P. Expression studies of the core+1 protein of the hepatitis C virus 1a in mammalian cells. The influence of the core protein and proteasomes on the intracellular levels of core+1. FEBS J. 2007, 274, 4057–4074. [Google Scholar] [CrossRef] [PubMed]
- Tai, A.W.; Benita, Y.; Peng, L.F.; Kim, S.S.; Sakamoto, N.; Xavier, R.J.; Chung, R.T. A functional genomic screen identifies cellular cofactors of hepatitis C virus replication. Cell Host Microbe 2009, 5, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germain, M.A.; Chatel-Chaix, L.; Gagné, B.; Bonneil, É.; Thibault, P.; Pradezynski, F.; de Chassey, B.; Meyniel-Schicklin, L.; Lotteau, V.; Baril, M.; et al. Elucidating novel hepatitis C virus-host interactions using combined mass spectrometry and functional genomics approaches. Mol. Cell. Proteomics. 2014, 13, 184–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, G.H.; Khan, M.; Yang, S.; Siddiqui, A. Hepatitis C Virus Lipoviroparticles Assemble in the Endoplasmic Reticulum (ER) and Bud off from the ER to the Golgi Compartment in COPII Vesicles. J. Virol. 2017, 91, e00499-17. [Google Scholar] [CrossRef] [Green Version]
- Vassilaki, N.; Kalliampakou, K.I.; Mavromara, P. Differences in the expression of the hepatitis C virus core+1 open reading frame between a nuclear and a cytoplasmic expression system. J. Gen. Viroloy 2008, 89, 222–231. [Google Scholar] [CrossRef]
- McGivern, D.R.; Lemon, S.M. Virus-specific mechanisms of carcinogenesis in hepatitis C virus associated liver cancer. Oncogene 2011, 30, 1969–1983. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.B.; Shao, S.W.; Zhao, L.J.; Luan, J.; Cao, J.; Gao, J.; Zhu, S.Y.; Qi, Z.T. Hepatitis C Virus F Protein Up-Regulates c-myc and Down-Regulates p53 in Human Hepatoma HepG2 Cells. Intervirology 2007, 50, 341–346. [Google Scholar] [CrossRef]
- Alisi, A.; Arciello, M.; Petrini, S.; Conti, B.; Missale, G.; Balsano, C. Focal adhesion kinase (FAK) mediates the induction of pro-oncogenic and fibrogenic phenotypes in hepatitis C virus (HCV)-infected cells. PLoS ONE 2012, 7, e44147. [Google Scholar] [CrossRef]
- Elsner, L.; Flügge, P.F.; Lozano, J.; Muppala, V.; Eiz-Vesper, B.; Demiroglu, S.Y.; Malzahn, D.; Herrmann, T.; Brunner, E.; Bickeböller, H.; et al. The endogenous danger signals HSP70 and MICA cooperate in the activation of cytotoxic effector functions of NK cells. J. Cell Mol. Med. 2010, 14, 992–1002. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, T.; Nishimura, Y.; Ichimura, T.; Suzuki, K.; Miyamura, T.; Suzuki, T.; Moriishi, K.; Matsuura, Y. Hepatitis C virus RNA replication is regulated by FKBP8 and Hsp90. EMBO J. 2006, 25, 5015–5025. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG PATHWAY (Pathway ID) | Gene Names | ||||
|---|---|---|---|---|---|
| Tight junction (hsa04530) | CTTN | ACTN1 | PKA | ||
| Actin Cytoskeleton (hsa04810) | ACTN1 | HRAS | - | - | - |
| MAPK Signaling (hsa04010) | HRAS | PKA | RAP1 | HSP72 | - |
| Antigen processing and Presentation (hsa04612) | PA28 | HSP70 | - | - | - |
| Endocytosis(hsa04144) | ARF | HRAS | HSC70 | SNX5 | - |
| Fatty acid degradation (hsa00071) | MFP | HADHA | ACAA | CTP2 | ALDH |
| Peroxisome(hsa04146) | SPX | ABCD | GSTK1 | PMP70 | - |
| Regulation | Hallmarks | Enriched Genes | Protein Names |
|---|---|---|---|
| Up regulated | APICAL JUNCTION | ACTN1 | Alpha-actinin-1 |
| EPB41L2 | Band 4.1-like protein 2 | ||
| HRAS | GTPase Hras | ||
| MITOTIC SPINDLE | CTTN | Src substrate cortactin | |
| EPB41L2 | Band 4.1-like protein 2 | ||
| LRPPRC | Leucine-rich PPR motif-containing protein | ||
| Down-regulated | MTORC1 SIGNALING | ACLY | ATP-citrate synthase |
| GBE1 | 1,4-alpha-glucan-branching enzyme | ||
| ASNS | Asparagine synthetase | ||
| COPS5 | COP9 signalosome complex subunit 5 | ||
| CTH | Cystathionine gamma-lyase | ||
| HK2 | Hexokinase-2 | ||
| PSMG1 | Proteasome assembly chaperone 1 | ||
| YKT6 | Synaptobrevin homolog YKT6 | ||
| APOPTOSIS | CTH | Cystathionine gamma-lyase | |
| DFFA | DNA fragmentation factor subunit alpha | ||
| HMOX1 | Heme oxygenase 1 | ||
| MYC TARGETS V1 | MRTO4 | mRNA turnover protein 4 homolog | |
| NIP7 | 60S ribosome subunit biogenesis protein homolog | ||
| SRM | Pyrimidine/purine nucleoside phosphorylase | ||
| HK2 | Hexokinase-2 | ||
| HYPOXIA | GBE1 | 1,4-alpha-glucan-branching enzyme | |
| HK2 | Hexokinase-2 | ||
| HMOX1 | Heme oxygenase 1 | ||
| UV RESPONSE UP | PPIF | Peptidyl-prolyl cis-trans isomerase F | |
| ASNS | Asparagine synthetase | ||
| YKT6 | Synaptobrevin homolog YKT6 | ||
| HMOX1 | Heme oxygenase 1 | ||
| GLYCOLYSIS | GMPPA | Mannose-1-phosphate guanyltransferase alpha | |
| HK2 | Hexokinase-2 | ||
| CTH | Cystathionine gamma-lyase |
| Gene Symbol | Phenotype | References |
|---|---|---|
| HRas | Carcinoma | [50] |
| CTTN | Tumor cell invasion | [53] |
| DFFA | Tumor cell invasion | [52] |
| YKT6 | Tumor cell invasion | [51] |
| PPIF | Steatosis | [54] |
| ACTN1 | Cirrhosis | [55] |
| EPB41L2 | Cirrhosis | [55] |
| HMOX | Cirrhosis | [56] |
| ASNS | Cirrhosis | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrazas, V.; Moustafa, S.; Makridakis, M.; Karakasiliotis, I.; Vlahou, A.; Mavromara, P.; Katsani, K.R. A Proteomic Approach to Study the Biological Role of Hepatitis C Virus Protein Core+1/ARFP. Viruses 2022, 14, 1694. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081694
Vrazas V, Moustafa S, Makridakis M, Karakasiliotis I, Vlahou A, Mavromara P, Katsani KR. A Proteomic Approach to Study the Biological Role of Hepatitis C Virus Protein Core+1/ARFP. Viruses. 2022; 14(8):1694. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081694
Chicago/Turabian StyleVrazas, Vasileios, Savvina Moustafa, Manousos Makridakis, Ioannis Karakasiliotis, Antonia Vlahou, Penelope Mavromara, and Katerina R. Katsani. 2022. "A Proteomic Approach to Study the Biological Role of Hepatitis C Virus Protein Core+1/ARFP" Viruses 14, no. 8: 1694. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081694