
Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting


1
INSA—Centre for Vectors and Infectious Diseases Research, National Institute of Health Doutor Ricardo Jorge, Avenida da Liberdade n.-5, 2965-575 Águas de Moura, Portugal
2
ISAMB—Instituto de Saúde Ambiental, Faculdade de Medicina da Universidade de Lisboa, Av. Prof. Egas Moniz, Ed. Egas Moniz, 1649-028 Lisboa, Portugal
3
BioISI—Biosystems and Integrative Sciences Institute, Edificio TecLabs, Campus da FCUL, Campo Grande, 1749-016 Lisboa, Portugal
*
Author to whom correspondence should be addressed.
Viruses 2022, 14(8), 1768; https://0-doi-org.brum.beds.ac.uk/10.3390/v14081768
Submission received: 7 July 2022
/
Revised: 9 August 2022
/
Accepted: 11 August 2022
/
Published: 13 August 2022
(This article belongs to the Special Issue Sand Fly-Borne Phleboviruses, Volume II)
Abstract
:According to ICTV, there are currently 66 known phlebovirus species. More than 40 of these viruses were isolated or detected in phlebotomine sandflies and some of them are known pathogens. In Portugal, information about sandfly-borne phleboviruses is scarce and scattered sandfly-borne diseases are neglected and often not considered in differential diagnoses. The main objective of this work was to gather the existing information and to raise awareness about the circulating phleboviruses in this country. To date, Massilia and Alcube phleboviruses have been isolated from sandflies in southern Portugal. Human infections with Toscana and Sicilian phleboviruses have been reported, as well as seroprevalence in cats and dogs. More studies are needed in order to understand if the viruses isolated during the entomological surveys have an impact on human health and to fully understand the real importance of the already recognized pathogens in our country.
1. Introduction
The genus Phlebovirus is included in the Phenuiviridae family. According to ICTV, this genus currently comprises 66 viral species [1]. Phleboviruses, like all members of Phenuiviridae, are tri-segmented, with a negative-stranded RNA genome. In the case of phleboviruses, this genome includes a small segment (S) which encodes the nucleocapsid protein (N) and a smaller nonstructural protein (NS), a medium segment (M) responsible for encoding the non-structural protein (NSm) and two envelope glycoproteins (Gn and Gc) and finally, a large (L) segment which encodes the RNA-dependent RNA polymerase (RdRp) [2].
It is accepted that phlebovirus taxonomy is complex since it undertakes constant updates and rearrangements as new genomic data emerge due, not only to the high mutation rates because of the absence of error detection and correction activities by the viral polymerase, but also to the phenomenon of recombination. Presently, the phlebovirus species demarcation criteria determines that viruses with <95% identity in the amino acid sequence of RdRp, encoded by the L segment, are considered a unique species [3]. Nevertheless, reassortants may challenge this current definition of species, emphasizing the prospect that additional debate may be essential for determining when they may be considered as members of different species [3,4,5].
Nowadays, more than 40 phleboviruses species are known to have been isolated from or detected in phlebotomine sandflies (Diptera: Psychodidae, Phlebotominae) [5]. Phlebotomine sandflies can be found almost everywhere in the world and their distribution includes several countries with warm climate such as southern Europe, Asia, Africa, Australia and Central and South America [6]. In the subfamily, Phlebotominae, we can find the genera Phlebotomus and Sergentomyia, which are present in Eurasia and Africa, and the genus Lutzomyia dispersed through Central and South America. Sandflies in the genera Phlebotomus and Lutzomyia assume an important role in public health since they are recognized vectors, not only of pathogenic phleboviruses, but also (and mainly) Leishmania spp. Parasites which are responsible for an estimated 700,000 to 1 million new cases annually worldwide [7]. Bartonella bacilliformis, a bacterium which, to this day, is known to occur only in the Andes Mountains, in western South America, including Peru, Colombia, and Ecuador, is also transmitted by sandflies and responsible for Oroya fever and verruga Peruana (Peruvian warts) [8].
In Portugal, five species of sandflies are presently recognized: Phlebotomus ariasi, Ph. Papatasi, Ph. Perniciosus, Ph. Sergenti and Sergentomyia minuta. Phleboviruses have been isolated from all these five species in the Mediterranean region (e.g., Toscana phlebovirus from Ph. perniciosus, Ph sergenti and S. minuta; Sicilian phlebovirus from Ph. ariasi and Ph papatasi) [9,10]. Here we present a review of the sandfly-borne phleboviruses known to circulate in Portugal.
2. Sicilian phlebovirus
The Sicilian phlebovirus is a sandfly-borne virus only recently recognized as a species [11]. Nonetheless, the sandfly fever Sicilian virus (SFSV), belonging to the aforementioned species, was isolated by Albert Sabin in 1943, during World War II, from a serum sample collected from a soldier in the USA allied troops who fell ill after the Sicily landings, and again, in that same year during an outbreak of febrile disease in Egypt [12,13].
SFSV is mainly transmitted by Ph. Papatasi but other sandfly species, such as Ph. Ariasi, and other species of Larroussius group such as Ph. Perniciosus, Ph. Neglectus and Ph. longicuspis, may also transmit viruses from the Sicilian phlebovirus species [10,14,15].
After a 3–5-day incubation period, the SFSV infection is characterized by sudden and severe fever, accompanied by headaches, malaise, photophobia, myalgia and retro-orbital pain. This disease is often designated as pappataci fever, sandfly fever or three-day fever, since the febrile syndrome usually lasts two or three days [16]. In contrast with Toscana virus infections which can be neurotropic, SFSV infections in general are not believed to be associated with neurologic manifestations. Nevertheless, there is a report of a German 15-year-old girl who had been in Turkey, presenting severe meningitis after initial mild symptoms. According to the authors, ELISA and immunoblot confirmed infection with SFSV, and this was the first time SFSV was associated with neurological symptoms [17]. Another publication referring to central nervous system involvement of SFSV reported a 30-year-old man testing IgM positive in anti-SFV IgG and IgM detection immunoassays. Initially, the patient presented with acute gastroenteritis-like illness but, two days later, he developed severe encephalitis with status epilepticus [18]. In addition, there is a case, which remains as a probable infection of SFSV, where a patient who developed encephalitis with lethal consequences, was diagnosed through positive serum in ELISA IgM antibodies detection [19]. On the other hand, Ergunay and colleagues (2012) confirmed, through PCR and genome sequencing, a case of encephalitis in a 63-year-old woman infected with sandfly fever Turkey virus, a Sicilian phlebovirus species member [20]. Furthermore, and still referring to unusual manifestations, there is a report of skin lesions in two patients with PCR positive results for SFSV [21].
Human infections of SFSV or Sicilian-like phleboviruses have been reported in many countries of the Mediterranean region and the Middle East. Outbreaks or sporadic human cases have been described, for example, in Cyprus, Turkey, Iraq and Ethiopia [15,22,23,24,25,26,27,28,29,30]. In addition to those reported cases of acute disease, retrospective serological studies performed on humans presenting with febrile syndromes and/or compatible symptoms with SFSV infections, together with serosurveys of healthy individuals, indicate that Sicilian phlebovirus, or closely related viruses, circulate in three continents: Europe, Africa and Asia. In Europe, cases have been reported from countries such as France, Italy, Greece, Kosovo, Turkey and Cyprus [28,31,32,33,34,35,36,37,38,39]. In Africa, there are studies from Algeria, Sudan and Egypt [14,40,41], and in Asia, there are seroprevalence reports from Iran, Israel, Pakistan, Bangladesh and Afghanistan [42,43,44,45,46,47].
In 1976, Tesh and colleagues reported a very extensive study developed in 59 localities in Africa, the Mediterranean region, eastern Europe and Asia using neutralizing antibodies against eight phleboviruses and found out that SFSV circulated, among others, also in Croatia, Morocco, Somalia, Saudi Arabia, Moldova, Azerbaijan, Uzbekistan and Turkmenistan [48].
Regarding the circulation of Sicilian phlebovirus among vertebrates other than human, including wild and domestic mammals, several reports confirm the detection in different mammal species, including rodents, insectivores and carnivores in Tunisia, Morocco, Spain and Italy [49,50,51,52], bats in Spain [16], dogs in Greece, Cyprus and Tunisia [53,54] and livestock in Kosovo [55].
In Portugal, the first reference to the SFSV was made in 1974, when a survey for antibodies to arboviruses in human sera was carried out and hemagglutination inhibition assays showed four positive reactions (in 1690 tested samples) with the SFSV virus antigen [56]. Since then, no serological surveys have been performed and no human infections of SFSV were reported until the summer of 2017, when an eight-year-old boy presenting with fever, anorexia and mild headaches tested positive for antibodies against SFSV and then seroconverted [57,58]. This was the first time that SFSV infection was linked to a symptomatic infection in this country. The presence of the virus in Portugal is corroborated by several seroprevalence studies which found antibodies in 4.3% (17/400) of the studied human population in Setúbal district, southwest of Portugal, and in domestic animals such as cats (2.2%, 8/367) and dogs (50.8%, 581/1160; 56.3%, 327/581) [59,60,61,62].
3. Toscana phlebovirus
Toscana phlebovirus (TOSV) was isolated for the first time from Ph. perniciosus sandflies, collected in 1971 in Monte Argentario, Tuscany region, during an arbovirus study in Italy. Later, between 1980 and 1985, the virus was also isolated from Ph. perfiliewi in a study carried out in the same region whose objective was to determine the possible vectors and foci of TOSV and its importance with regard to human health [63].
It was only in the summer of 1983 that this virus was isolated for the first time from the cerebrospinal fluid (CSF) of a young woman with aseptic meningitis attending a hospital in the Tuscany region [64]. As such, it has now been recognized for a long time that TOSV is accountable, not only for asymptomatic or mild infections, but is also commonly associated with neurological disease, making it the most important sandfly-borne phlebovirus in terms of public health, and apparently, together with enteroviruses and herpesviruses, one of the three major viral pathogens responsible for aseptic meningitis reported during the warmer season in countries around the Mediterranean Basin [65,66].
A recent study estimated the incubation period of TOSV at 12 days [67]. Viremia persists only for two or three days, and cases can be diagnosed by direct isolation of the virus or RNA detection through reverse transcription PCR in CSF or blood [68]. In addition, there are records of TOSV RNA detection in urine samples and, more recently, of infectious TOSV in human semen which may indicate a potential for sexual transmission [69,70]. However, since the viremic period is short, the diagnosis of TOSV can be confirmed serologically, for example, through a four-fold or greater change in virus-specific quantitative antibody titers in paired sera or virus-specific IgM antibodies in serum with confirmatory virus-specific neutralizing antibodies in the same or a later specimen [71].
The most common clinical manifestations due to TOSV infections are fever, headache, nausea and vomiting, fatigue, photophobia, myalgia, febrile episodes, rash and stiff neck. Most infections, including the ones that lead to meningitis, encephalitis or meningoencephalitis, have a benign course and are self-limited; however, six fatal outcomes in elderly patients have been recorded: one in Italy and five in Romania [72,73]. Among the atypical clinical manifestations, with neurological involvement, reports of Guillain-Barré syndrome, hydrocephalus, hearing loss, speech problems, paresis, myositis and fasciitis, facial paralysis and personality changes can be found [74,75,76,77,78,79,80,81,82]. Testicular manifestations are also documented [76,83,84].
Until recently, TOSV was included in the sandfly fever Naples virus species but it is now considered a species of its own [11]. Furthermore, genetic studies have described the existence of diverse TOSV genotypes. In the latest review about TOSV, three lineages were confirmed to circulate in the Mediterranean area: lineage A strains in Italy, France, Turkey, Tunisia and Algeria; lineage B strains in Portugal, Spain, France, Morocco, Croatia and Turkey and lineage C in Croatia and Greece [85]. Co-circulation of strains A and B has been documented in France and Turkey and of strains B and C in Croatia. TOSV is also present in other Balkan countries such as Kosovo, Bosnia Herzegovina and Bulgaria, and in the Mediterranean islands of Elba, Baleares, Malta, Corsica, Sardinia, Cyprus and Crete [85]. Human seroprevalence of TOSV is usually around 10−24% but can reach 40% or higher within the endemic regions, as in the case of northern Tunisia where a microneutralization-based study detected a seroprevalence of 41% (522/1273) [86,87]. A seroprevalence as high 77.2% (278/360) was detected, for example, in sera samples collected in a high-risk population in Italy (Tuscany), and although the numbers varied significantly in the different regions, 37.5% (755/2016) of a studied resident population in Croatia presented IgM antibodies [88,89].
The natural cycle of TOSV remains unclear. Nevertheless, experimental infections of sandflies with the virus confirmed that both transovarian and venereal transmission may occur [90,91,92]. In addition, TOSV has also been isolated in nature from male and female sandflies [93]. The main vector is believed to be the sandfly Ph. Perniciosus, but whether there is a vertebrate reservoir is yet to be confirmed. The first isolation of TOSV from a non-human vertebrate was from a brain sample of a bat (Pipistrellus kuhlii) [63]. Additionally, TOSV sequences were identified in birds’ organs during the screening of avian specimens collected in the Mediterranean coast of the Anatolian peninsula, Turkey [94]. Viral isolation was not achieved but brain and kidney tissues from a greater flamingo (Phoenicopterus roseus), a great white pelican (Pelecanus onocrotalus) and a black stork (Ciconia nigra) tested positive for TOSV genotypes A and B in PCR assays [94]. TOSV has also been detected in other vertebrates, for example, in Granada (Spain), 48.3% (138/286) of dogs were seropositive for TOSV; seropositivity of 3.9% (9/231) was found in dogs on the eastern coast of Corsica and out of a total of 93 dogs, 4 were seropositive (4.3%) for TOSV in Kabylia [95,96,97].
To date, despite extensive efforts, TOSV has never been detected in entomological surveys conducted in wild-caught sandflies in Portugal and the first reference to the presence of this virus in the country was made in 1985 when its isolation from a tourist who became infected in Albufeira, in 1983, was reported in Sweden [98]. Therefore, Portugal was the second country, after Italy, to be considered endemic for TOSV [98]. Years later, in 1995, another tourist returned symptomatic to his country of origin, Germany, after being infected with TOSV in the region of Coimbra [99]. Subsequently, between 2002 and 2005, six more cases of TOSV were diagnosed, using molecular methods, in Portuguese patients with meningitis attending hospitals in the metropolitan area of Oporto, in the north of Portugal, but no genomic sequences were reported [100]. In a follow-up study, a seroprevalence of 3.9% (IgG) was found in 334 sera samples randomly collected from individuals who sought care in the hospitals in the same region [101].
In 2011, a report of a serological study performed at the National Reference Laboratory for vector-borne viruses (Centre for Vectors and Infectious Diseases Research, National Institute of Health, CEVDI/INSA) was published. This study included samples collected from 538 patients from all over the country, with and without neurological signs between 2004 and 2008 [102]. In house indirect immunofluorescence assay and commercial enzyme-linked immunosorbent assays were used. A prevalence of 4.2% (7/165) for IgG antibodies was found in the group of patients with neurological signs. Additionally, 3% (5/165) of the patients had IgG and IgM, revealing recent infections. In the group with no neurological signs, the IgG prevalence was 1.3% (5/373). In this study, only two patient samples were also confirmed with plaque reduction neutralization tests with the TOSV ISS. Phl.3 Italian strain, implying that different genotypes of TOSV virus may be circulating in Portugal.
In 2021, an update of TOSV cases in a total of 608 patients, whose samples were sent for laboratorial diagnosis at CEVDI/INSA between January 2008 and December 2018, referred to five acute TOSV infections. Three other patients presented serological evidence of previous contact with the virus [58]. Another recently published regional study (2022) involving only individuals from the Setúbal county, disclosed that out of 400 sera tested, 21 (5.3%) were positive for TOSV IgG [59].
Concerning seroprevalence studies in non-human vertebrates in Portugal, an investigation using in-house immunofluorescence assays performed in sera samples of wolves (Canis lupus signatus) and foxes (Vulpes vulpes) found that 1 in 49 wolves (2%) and 1 in 37 foxes (2.7%) presented IgG antibodies [103]. In another report using the same technique, 100 healthy military dogs were screened and one tested IgG positive for TOSV [104]. Other studies involving neutralization assays in pets detected seroprevalence for TOSV: 6.8%, (79/1160) and 6.2% (36/581) in dogs and 3.7% (7/189) and 4.9% (18/365) in cats. [60,61,62].
4. Massilia phlebovirus
Massilia phlebovirus was isolated for the first time from pools of Ph. perniciosus sandflies collected in the suburban area of Marseille, France, in July 2005 [105]. In order to assess the possible existence of human infections, a retrospective study aiming at the detection of Massilia phlebovirus RNA was performed in 477 CSF samples from local patients presenting central nervous system disease between 2002 and 2006. The results were all negative [105].
Sanchéz-Seco and colleagues, in 2010, reported the detection of Massilia-like virus in sandflies collected in Catalonia, Spain [106]. In the same country, during a survey in sandflies in Granada province, a phlebovirus was isolated, fully sequenced and tentatively named Granada virus (GRV) [107]. GRV was, by that time, described as a new phlebovirus, likely to be a natural reassortant of the Massilia virus [107]. Antibodies against GRV were investigated in sera from healthy individuals collected in the Granada province, in 2003. From a total of 248, 37 (14.9%) tested positive by immunofluorescence assay. To discard cross-reactivity with TOSV, the positive sera were tested for neutralizing antibodies against GRV, and five samples retrieved positive results [107].
In a subsequent study conducted in the same region, IgG detection through immunofluorescence assays was carried out in asymptomatic individuals and IgM detection was performed (with the same method) in patients attending healthcare facilities during the warmer months [108]. In total, 145 healthy individuals out of 920 (15.8%) were positive for anti-GRV antibodies, and from these, 17.9% (n = 26) were positive for anti-GRV neutralizing antibodies. In the group of symptomatic individuals, 36 out of 547 were IgM positive. The authors of the study concluded that GRV may infect humans in that region and, as expected, most cases would probably cause no symptoms [108].
Despite being initially regarded as a new virus, considering the new criteria for species demarcation, GRV seems, in fact, to belong to Massilia phlebovirus species [4]. The fact that human seroprevalence was found for GRV may suggest that Massilia phlebovirus can infect humans. However, it is not possible to assume that this species can cause human disease.
During a phlebotomine survey conducted in the south of Portugal during the summer in 2007 and 2008, a phlebovirus, tentatively named Arrábida virus, was isolated from sandflies collected in Arrábida region, Setúbal county. After whole genome sequencing, as in the case of GRV, it was believed that we were in the presence of a new reassortant phlebovirus [109]. However, once again, following the ICTV criteria, Arrábida virus must be considered a member of Massilia phlebovirus. In the same survey it was possible to detect and fully sequence two other Massilia phlebovirus variants (Massilia virus isolate 127 in Arrábida region and Massilia virus isolate 130 in eastern Algarve, Olhão county) and to isolate and fully sequence two other Massilia variants in sandflies collected, once again, in Arrábida: PoSFPhlebV/21/2007 and PoSFPhlebV/70/2007 [4,110].
In agreement with the gathered data, Massilia phlebovirus is known to circulate in Portugal, Spain and France. To date, no seroprevalence studies have been carried out in Portugal.
5. Alcube phlebovirus
Alcube phlebovirus was isolated from a pool of sandflies collected in Arrábida region, in July 2007 [110]. It was firstly described as a new phlebovirus species clustering with members of the Salehabad virus species complex and forming a distinct monophyletic lineage with Arbia, (isolated from sandflies in Italy), Salehabad (first isolated from sandflies in Iran) and Adana (isolated from sandflies in Turkey) viruses [64,109,111,112]. According to the new taxonomic arrangement [1], all these viruses are currently recognized as different species, except for Arbia virus which is currently included in Medjerda phlebovirus species [5].
In general, the species formerly included in the Salehabad group were not considered to be of significant medical or veterinary interest [113]. Nevertheless, high seroprevalence rates of Adana phlebovirus have been found in animals such as goats, sheep and dogs, and low seroprevalence rates have been identified in the human population [112]. On the other hand, genomic RNA of Adria virus (belonging to Salehabad phlebovirus species) was first detected in sandflies in Albania, and later in Greece in a child with febrile syndrome and convulsions, which may suggest its pathogenicity [114,115]. Further studies are needed in order to clarify whether Alcube phlebovirus infects humans.
6. Other Genome Detections of Phleboviruses in Sandfly Pools in Portugal
During the entomological surveys performed across two regions in the south of Portugal in the summers of 2007, 2008 and 2018, other RNA detections, unpublished until now, were achieved in 22 pools of sandflies in addition to the already fully sequenced and mentioned Alcube and Massilia phleboviruses strains.
For the additional detections, isolation attempts, performed as previously described [110], were not successful after at least three blind passages in Vero E6 cells, probably due to sample degradation. Thus, further investigation was not feasible. Nevertheless, it was possible to obtain partial sequences for the S segment of 13 phleboviruses strains [116,117], four for the M segment (this study) and five for the L segment [117] in this study. For the segment M sequence amplification, primers PhlebMF1 (5′-CTCKATTGAAKATKGCCATKGG-3′) and PhleboMR1 (5′-ATGCTTTGAGCAGAGYGGWGG-3′) were specifically designed to amplify a 494 bp partial sequence of viruses related to MASV, GRAV and TOSV. For pool 149/2008, amplification of a 1234 bp partial sequence of RdRp (segment L) was obtained using primers MASV_LF2 (5′-CTGACAAGGCTGACGGTTCT-3′) and MASV_LR2 (5′-TGTACCAACGCCACGATTGA-3′) designed to amplify Massilia virus genome. Briefly, for the primers designed in this study, 5 μL of RNA and 10 pmol of each primer were added to SuperScript® One-Step RT-PCR with Platinum® Taq (Invitrogen by Life Technologies, Carlsbad, CA, USA). Polymerase chain reaction (PCR) conditions were as follows: reverse transcription at 50 °C for 30 min, denaturation at 95 °C for 5 min, 45 cycles of 94 °C for 20 s, 55 °C for 60 s and 72 °C for 60 s, and a final extension at 72 °C for 5 min. To increase sensitivity, a second round PCR was performed using the same primers and high fidelity PCR master (Roche, Mannheim, Germany). The obtained amplicons were purified after gel agarose visualization, using JETquick PCR Product Purification Spin kit (GENOMED GmbH, Löhne, Germany) and sequenced bi-directionally using ABI Prism 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). Homology searches within the GenBank data set were performed using the BLASTn algorithm [118]. GenBank accession numbers of all new sequences reported in this study are presented in Table 1. Phleboviruses partial sequences were aligned with sequences available from GenBank using Clustal W within BioEdit version 7.2.5 [119] and manually edited whenever necessary (alignments are available in Supplementary Material). Phylogenetic analysis was performed using partial nucleotide sequences of nucleocapsid (n = 33), glycoprotein precursor (n = 20) and RNA-dependent RNA polymerase genes (n = 22 and n = 19). Maximum likelihood phylogenetic trees were estimated in Mega version X software [120] by using the best-fit model of nucleotide substitution as indicated by the Best DNA/Protein Model application (implemented in Mega X) [121]. The robustness of the inferred tree was tested by 1000 bootstrap replications.
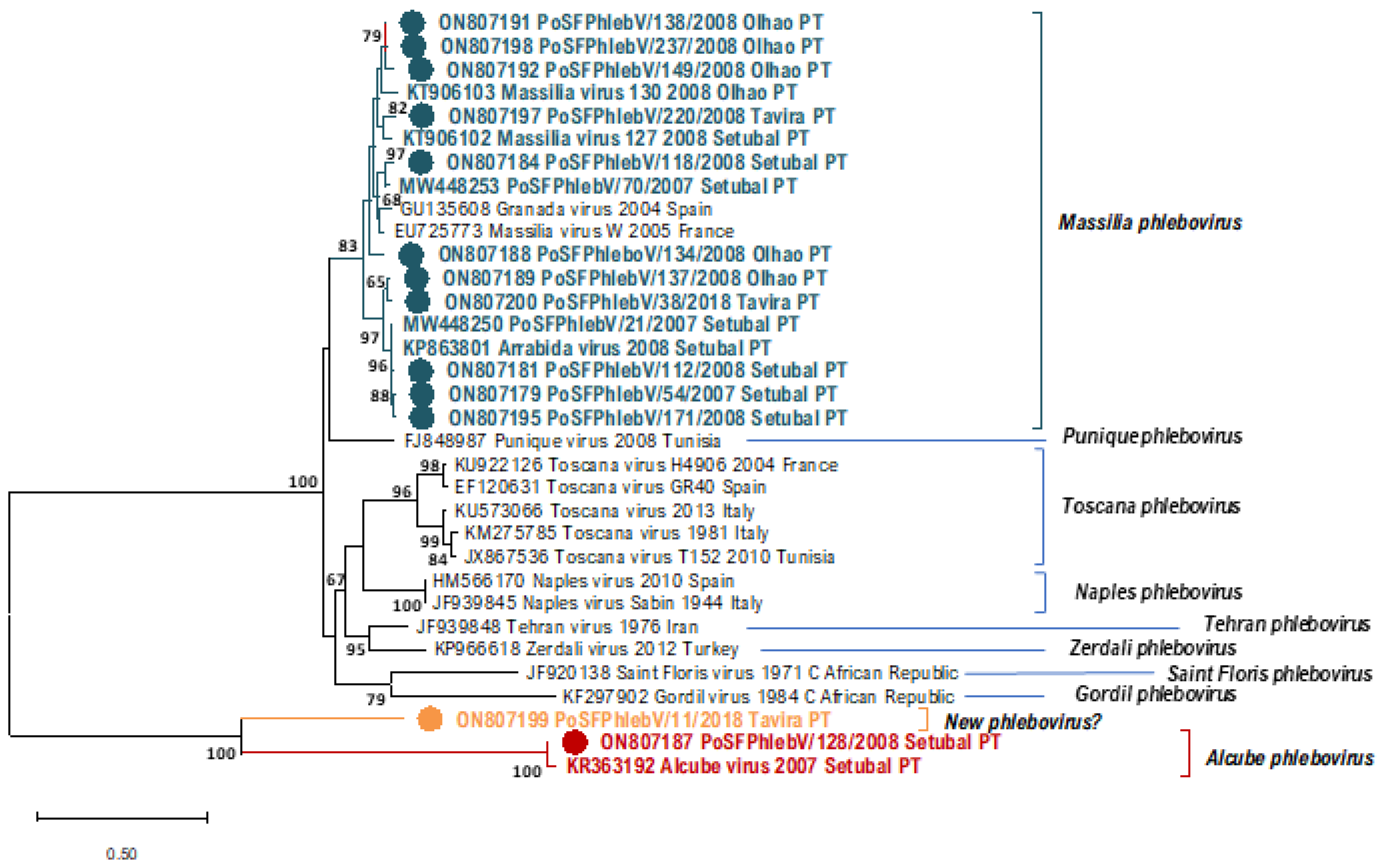
In Table 1, we can see the sequence data of all the phleboviruses detected/isolated from sandfly pools so far in Portugal. All the additionally detected phlebovirus sequences show a close relation to Massilia phlebovirus, except for PoSFPhlebV/128/2008 presumptively identified as Alcube phlebovirus through N gene partial sequence similarity (Table 1 and Figure 1, Figure 2 and Figure 3). The available data do not support a closer similarity of variants related to their geographic location detection (Setúbal versus Algarve region).
In the absence of complete genome sequences, it is not possible to draw firm conclusions about the genetic diversity of all detected viruses. Nevertheless, the similarity results obtained from BLASTn analysis (data not shown) and the branch changes in the trees for each genome segment (Figure 1, Figure 2 and Figure 3) indicate that some of these Massilia phlebovirus related viruses may represent reassortants. For instance, from the segment S partial sequence similarity analysis for PoSFPhleb/118/2008, it seems that this variant is closely related to Massilia phlebovirus PoSFPhleboV/70/2007 (Figure 1). However, using the segment M partial sequence, a closer similarity is observed with the GRV virus sequence (Figure 2). On the other hand, analyzing segment L partial sequences (Figure 3), no indication of a closely related ancestor of PoSFPhleb/118/2008 and the former Massilia phlebovirus variants is observed.
Branch changes are also clearly observed for PoSFPhlebV/112/2008 when analyzing different genome segments available sequences showing closer proximity to Massilia phlebovirus: Arrábida virus and PoSFPhleb/21/2007 in segment S, Massilia virus 127 and 130 in segment M and none of them in segment L sequences.
The partial gene N sequence detected in pool 11/2018 (PoSFPhlebV/11/2018; GenBank accession ON807199) shows a higher distance from all the other phleboviruses’ sequences. Although the available data are insufficient to enable the species identification, it seems to indicate potential circulation, in the Algarve region, of a new phlebovirus more closely related to Alcube Phlebovirus.
7. Discussion
In the last decades, entomological and virological studies have been expanding [122]. Concomitantly, the advances in laboratory techniques, particularly in molecular diagnostics, have enabled great strides in the identification and characterization of novel phleboviruses, resulting in a considerable increase in the number of recognized members of this genus. Phleboviruses may be responsible for more human diseases than previously thought [123]. However, as most native adults in endemic regions are immune, there are relatively few reports of clinical cases among indigenous people, in addition to the fact that mild symptoms do not frequently lead people to seek health care [23]. Often, reported cases of more severe infections are from travelers visiting endemic regions [124,125,126,127]. Yet, and surprisingly, some clinicians are still not considering TOSV and phleboviruses, in general, as causative agents of disease. Reports of the expansion of vector sandflies to more central countries in Europe, such as Germany, Switzerland and Austria, indicate that sandflies are expanding toward the north and raise awareness for the increasing risk for sandfly-borne diseases in territories not previously considered [128,129,130,131].
The first reference of a phlebovirus in Portugal was made in 1974, regarding human seroprevalence of Sicilian phlebovirus. This phlebovirus remained unnoticed in the human population until 2017. This species is distributed in three continents, and outbreaks caused by its members are known to occur from time to time in different parts of Eurasia and Africa. Cyprus phlebovirus (Sicilian-like) was recently indicated as the cause of a life-threatening condition in a 3.8-year-old child in Italy. It was the first time this variant was detected outside Cyprus [132]. This should be a warning sign that not only national reference centers, but also hospital microbiology laboratories should be equipped for systematic phleboviruses testing of patients presenting with febrile illness and central and peripheral nervous system febrile manifestations [53].
TOSV, which is also a widespread virus affecting two continents, causes infections leading to mild or neurological disease from north to south of Portugal. The fact that the only complete genomic sequences displayed in GenBank are from the S segment of the virus isolated in 1983 from a Swedish tourist corroborates that this virus is not being given its due importance in our country. For example, in Spain, in an update of TOSV neurological infection in Andalusia, from 1988 to 2020, TOSV was the second agent detected in CSF samples between April and November. In Granada province, considered as an hyperendemic area for TOSV, a seroprevalence of 25% was found [133]. Moreover, Collao and colleagues (2009) reported that sequences independently obtained in two laboratories from strains allegedly obtained from the same patient, infected in Portugal, clustered in different clades, genotypes A and B [134,135,136]. Indisputably, investigations of infections in the central nervous system and fever of unknown origin are needed in order to better understand the epidemiology of TOSV in Portugal and which genotypes of this virus are circulating. Also, other entomological investigations are necessary in Portugal.
The lack of genomic detection of phleboviruses in cats and dogs in Portugal is in line with studies from other regions. For instance, in an experimental infection of dogs with TOSV and Sicilian phlebovirus in Spain, it was demonstrated that healthy domestic dogs do not exhibit susceptibility to infection by these two viruses. The subjects did not show the development of noteworthy viremia nor expelled the viruses after being experimentally infected. Based on these results, dogs do not seem to be natural reservoir hosts of infection, nor play a meaningful part in phleboviruses’ transmission cycles [137]. As for wild mammals, a seroprevalence study of TOSV and Sicilian phleboviruses performed in bat colonies from southern Spain showed that, despite positive sera for both viruses found in different species, it is unlikely that these mammals play an important role in the biological cycle of the viruses in question [16].
MASV, with unknown pathogenic potential, was detected in entomological surveys in Setúbal and Faro, two districts in the south of Portugal. This fact leads us to believe that the virus may be widely spread in the country, or, at least, all over the south since the locations where this species was found are not contiguous. Even though complete genome sequences were not possible to achieve for the newly presented phleboviruses, analysis of their partial sequences, along with the complete genomes available within Massilia phebovirus species, highlights an impressive sequence diversity and potential for recombination/reassortment, which may explain the difficulty in detecting phleboviruses by PCR in surveillance studies. Further entomological surveys would be necessary to clarify this assumption, and human seroprevalence investigations could help to clarify if this virus is infectious to humans.
To date, Alcube phlebovirus has been detected only in Setúbal county, south of Portugal. Further investigation is needed to clarify its distribution and importance to public health which remains, as yet, undetermined.
8. Conclusions
Four phleboviruses are known to circulate in Portugal: Sicilian phlebovirus, Toscana phlebovirus, Alcube phlebovirus and Massilia phlebovirus. The first two are known pathogens in the Mediterranean Basin and responsible for cases of febrile illness or neurological disease in the summer months. The other two have not yet been classified as pathogens but further epidemiological studies must be performed in order to clarify this matter.
The possibility that a wider number of unidentified phleboviruses is circulating in Portugal should be considered. Furthermore, the fact that Alcube and Massilia viruses were isolated in 2007, at the very same location, meaning that co-circulating viruses can be found, raises concerns, especially due to the genomic characteristics of these viruses. Multiple infections in arthropods may favor reassortments of the genome segments and the major concern is that this process may modify the phleboviruses’ biological properties or originate viruses with unknown pathogen capacity. As such, the characterization of novel members of this genus and the competency to acknowledge the existence of reassortants is of utmost importance to prevent the occurrence of outbreaks of these emerging pathogens.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/v14081768/s1, File S1: Phleboviruses’ nucleotide partial sequence alignment of S segments, File S2: Phleboviruses’ nucleotide partial sequence alignment of M segments, File S3: (a) Phleboviruses’ nucleotide partial sequence alignment of L segments, File S4: (b) Phleboviruses’ nucleotide partial sequence alignment of L segments.
Author Contributions
Conceptualization, F.A.; methodology, F.A. and L.Z.-Z.; validation, F.A., L.Z.-Z. and M.J.A.; formal analysis, F.A. and L.Z.-Z.; writing—original draft preparation, F.A.; writing and editing, all authors.; funding acquisition, F.A. and M.J.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by the Portuguese Foundation for Science and Technology (FCT) through the project: “Phleboviruses in Portugal: vectors, pathogenesis and co-infections” (PTDC/DTP-SAP/0859/2014).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Natacha Milhano for the revision of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2021 Release. EC 53, Online, July 2021. Email Ratification March 2022 (MSL #37). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 5 April 2022).
- Bouloy, M. Molecular Biology of Phleboviruses. In Bunyaviridae Molecular and Cellular Biology; Plyusnin, A., Elliott, R.M., Eds.; Caister Academic Press: Norfolk, UK, 2011; pp. 95–128. [Google Scholar]
- International Committee on Taxonomy of Viruses. Virus Taxonomy Release: 2020 Release. Email Ratification March 2021 (MSL #36). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 30 April 2022).
- Amaro, F.; Zé-Zé, L.; Lourenço, J.; Giovanetti, M.; Becker, S.C.; Alves, M.J. Phylogenetic analysis of Massilia phlebovirus in Portugal. Viruses 2021, 13, 1412. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Calzolari, M. Taxonomy of phleboviruses, emphasizing those that are sandfly-borne. Viruses 2021, 13, 918. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. The biology and control of phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- World Health Organization. Leishmaniasis. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 13 June 2022).
- Centers for Disease Prevention and Control. Bartonella bacilliformis Infection. Available online: https://www.cdc.gov/bartonella/bartonella-bacilliformis/index.html (accessed on 25 May 2022).
- Ayhan, N.; Charrel, R.N. Sandfly-Borne Viruses of Demonstrated/Relevant Medical Importance. In Vectors and Vector-Borne Zoonotic Diseases; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Depaquit, J.; Grandadam, M.; Fouque, F.; Andry, P.E.; Peyrefitte, C. Arthropod-borne viruses transmitted by Phlebotomine sandflies in Europe: A review. Euro Surveill. 2010, 15, 19507. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Amarasingh, G.K.; Anthony, S.J.; Avšič-Županc, T.; Ayllón, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. 2020 taxonomic update for phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2020, 165, 3023–3072. [Google Scholar] [CrossRef] [PubMed]
- Moureau, G.; Bichaud, L.; Salez, N.; Ninove, L.; Hamrioui, B.; Belazzoug, S.; de Lamballerie, X.; Izri, A.; Charrel, R.N. Molecular and serological evidence for the presence of novel phleboviruses in sandflies from northern Algeria. Open. Virol. J. 2010, 4, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Bichaud, L.; de Lamballerie, X.; Alten, B.; Gould, E.A.; Charrel, R.N. Sandfly-borne phleboviruses of Eurasia and Africa: Epidemiology, genetic diversity, geographic range, control measures. Antiv. Res. 2013, 100, 54–74. [Google Scholar] [CrossRef]
- Izri, A.; Temmam, S.; Moureau, G.; Hamrioui, B.; de Lamballerie, X.; Charrel, R.N. Sandfly fever Sicilian virus, Algeria. Emerg. Infect. Dis. 2008, 14, 795–797. [Google Scholar] [CrossRef]
- Woyessa, A.B.; Omballa, V.; Wang, D.; Lambert, A.; Waiboci, L.; Ayele, W.; Ahmed, A.; Abera, N.A.; Cao, S.; Ochieng, M.; et al. An outbreak of acute febrile illness caused by Sandfly Fever Sicilian Virus in the Afar region of Ethiopia, 2011. Am. J. Trop. Med. Hyg. 2014, 91, 1250–1253. [Google Scholar] [CrossRef]
- Ayhan, N.; López-Roig, M.; Monastiri, A.; Charrel, R.N.; Serra-Cobo, J. Seroprevalence of Toscana Virus and Sandfly Fever Sicilian Virus in European bat colonies measured using a neutralization test. Viruses 2021, 13, 88. [Google Scholar] [CrossRef]
- Becker, M.; Zielen, S.; Schwarz, T.F.; Linde, R.; Hofmann, D. Pappataci-Fieber [Pappataci fever]. Klin. Padiatr. 1997, 209, 377–379. (In German) [Google Scholar] [CrossRef] [PubMed]
- Lesho, E.P.; Ludwig, G.V.; Wortmann, G. Encephalitis and Sandfly Fever (Sicilian) virus infection. Infect. Dis. Clin. Pract. 2004, 12, 352–354. [Google Scholar] [CrossRef]
- Selim, H.S.; El-Barrawy, M.A.; Rakha, M.E.; Yingst, S.L.; Baskharoun, M.F. Microbial study of meningitis and encephalitis cases. J. Egypt. Public Health Assoc. 2007, 82, 1–19. [Google Scholar]
- Ergunay, K.; Ismayilova, V.; Colpak, I.A.; Kansu, T.; Us, D. A case of central nervous system infection due to a novel Sandfly Fever Virus (SFV) variant: Sandfly Fever Turkey Virus (SFTV). J. Clin. Virol. 2012, 54, 79–82. [Google Scholar] [CrossRef]
- Temocin, F.; Sari, T.; Tulek, N. Sandfly fever with skin lesions: A case series from Turkey. J. Arthropod Borne Dis. 2016, 10, 608–612. [Google Scholar]
- Niklasson, B.; Eitrem, R. Sandfly fever among Swedish UN troops in Cyprus. Lancet 1985, 1, 1212. [Google Scholar] [CrossRef]
- Papa, A.; Konstantinou, G.; Pavlidou, V.; Antoniadis, A. Sandfly fever virus outbreak in Cyprus. Clin. Microbiol. Infect. 2006, 12, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Eitrem, R.; Niklasson, B.; Weiland, O. Sandfly fever among Swedish tourists. Scand. J. Infect. Dis. 1991, 23, 451–457. [Google Scholar] [CrossRef]
- Carhan, A.; Uyar, Y.; Ozkaya, E.; Ertek, M.; Dobler, G.; Dilcher, M.; Wang, Y.; Spiegel, M.; Hufert, F.; Weidmann, M. Characterization of a sandfly fever Sicilian virus isolated during a sandfly fever epidemic in Turkey. J. Clin. Virol. 2010, 48, 264–269. [Google Scholar] [CrossRef]
- Guler, S.; Guler, E.; Caglayik, D.Y.; Kokoglu, O.F.; Ucmak, H.; Bayrakdar, F.; Uyar, Y. A sandfly fever virus outbreak in the East Mediterranean region of Turkey. Int. J. Infect. Dis. 2012, 16, e244–e246. [Google Scholar] [CrossRef]
- Torun, E.C.; Yağçı, Ç.D.; Uyar, Y.; Korukluoğlu, G.; Ertek, M.K. Sandfly fever outbreak in a province at Central Anatolia, Turkey. Mikrobiyol. Bul. 2010, 44, 431–439. [Google Scholar]
- Tezcan, S.; Dinçer, E.; Ülger, M.; Özgür, D.; Erdoğan, S.; Özkul, A.; Emekdaş, G.; Ergünay, K. Serological investigation of phlebovirus exposure in blood donors from the Mediterranean Province of Mersin, Turkey. Mikrobiyol. Bul. 2015, 49, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Ellis, S.B.; Appenzeller, G.; Lee, H.; Mullen, K.; Swenness, R.; Pimentel, G.; Mohareb, E.; Warner, C. Outbreak of sandfly fever in central Iraq, September 2007. Mil. Med. 2008, 173, 949–953. [Google Scholar] [CrossRef]
- Riddle, M.S.; Althoff, J.M.; Earhart, K.; Monteville, M.R.; Yingst, S.L.; Mohareb, E.W.; Putnam, S.D.; Sanders, J.W. Serological evidence of arboviral infection and self-reported febrile illness among U.S. troops deployed to Al Asad, Iraq. Epidemiol. Infect. 2008, 136, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Piarroux, R.P.; Izri, A.; Ninove, L.; Mary, C.; De Lamballerie, X.; Charrel, R.N. Low seroprevalence of sandfly fever Sicilian virus antibodies in humans, Marseille, France. Clin. Microbiol. Infect. 2011, 17, 1189–1190. [Google Scholar] [CrossRef]
- Calamusa, G.; Valenti, R.M.; Vitale, F.; Mammina, C.; Romano, N.; Goedert, J.J.; Gori-Savellini, G.; Cusi, M.G.; Amodio, E. Seroprevalence of and risk factors for Toscana and Sicilian virus infection in a sample population of Sicily (Italy). J. Infect. 2012, 64, 212–217. [Google Scholar] [CrossRef]
- Marchi, S.; Trombetta, C.M.; Kistner, O.; Montomoli, E. Seroprevalence study of Toscana virus and viruses belonging to the Sandfly fever Naples antigenic complex in central and southern Italy. J. Infect. Public Health 2017, 10, 866–869. [Google Scholar] [CrossRef]
- Percivalle, E.; Cassaniti, I.; Calzolari, M.; Lelli, D.; Baldanti, F. Thirteen years of phleboviruses circulation in Lombardy, a Northern Italy region. Viruses 2021, 13, 209. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Papaevangelou, G. Effect of insecticide spraying for malaria control on the incidence of sandfly fever in Athens, Greece. Am. J. Trop. Med. Hyg. 1977, 26, 163–166. [Google Scholar] [CrossRef]
- Antoniadis, A.; Alexiou Daniel, S.; Malissiovas, N.; Doutsos, J.; Polyzoni, T.; LeDuc, J.W.; Peters, C.J.; Saviolakis, G. Seroepidemiological Survey for Antibodies to Arboviruses in Greece; In Hemorrhagic Fever with Renal Syndrome, Tick-and Mosquito-Borne Viruses; Springer: Vienna, Austria, 1990; pp. 277–285. [Google Scholar]
- Venturi, G.; Marchi, A.; Fiorentini, C.; Ramadani, N.; Quaglio, G.; Kalaveshi, A.; Bertinato, L.; Putoto, G.; Benedetti, E.; Rezza, G.; et al. Prevalence of antibodies to phleboviruses and flaviviruses in Peja, Kosovo. Clin. Microbiol. Infect. 2011, 17, 1180–1182. [Google Scholar] [CrossRef]
- Eitrem, R.; Stylianou, M.; Niklasson, B. High prevalence rates of antibody to three sandfly fever viruses (Sicilian, Naples, and Toscana) among Cypriots. Epidemiol. Infect. 1991, 107, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Billioud, G.; Tryfonos, C.; Richter, J. The Prevalence of Antibodies against Sandfly Fever Viruses and West Nile Virus in Cyprus. J. Arthropod. Borne. Dis. 2019, 13, 116–125, Published 30 March 2019. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.C.; Haberberger, R.L.; Salib, A.W.; Soliman, B.A.; El-Tigani, A.; Khalid, I.O.; Watts, D.M. Evaluation of arthropod-borne viruses and other infectious disease pathogens as the causes of febrile illnesses in the Khartoum Province of Sudan. J. Med. Virol. 1996, 48, 141–146. [Google Scholar] [CrossRef]
- Darwish, M.A.; Feinsod, F.M.; Scott, R.M.N.; Ksiazek, T.G.; Botros, B.A.M.; Farrag, I.H.; Said, S. Arboviral causes of non-specific fever and myalgia in a fever hospital patient population in Cairo, Egypt. Trans. R. Soc. Trop. Med. Hyg. 1987, 87, 1001–1003. [Google Scholar] [CrossRef]
- Saidi, S.; Tesh, R.B.; Javadian, E.; Sahabi, Z.; Nadim, A. Studies on the epidemiology of sandfly fever in Iran. II. The prevalence of human and animal infection with five phlebotomus fever virus serotypes in Isfahan province. Am. J. Trop. Med. Hyg. 1977, 26, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Shiraly, R.; Khosravi, A.; Farahangiz, S. Seroprevalence of sandfly fever virus infection in military personnel on the western border of Iran. J. Infect. Public Health 2017, 10, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Cohen, D.; Zaide, Y.; Karasenty, E.; Schwarz, M.; LeDuc, J.W.; Slepon, R.; Ksiazek, T.G.; Shemer, J.; Green, M.S. Prevalence of antibodies to West Nile fever, sandfly fever Sicilian, and sandfly fever Naples viruses in healthy adults in Israel. Public Health Rev. 1999, 27, 217–230. [Google Scholar]
- Darwish, M.A.; Hoogstraal, H.; Roberts, T.J.; Ghazi, R.; Amer, T. A seroepidemiological survey for Bunyaviridae and certain other arboviruses in Pakistan. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 446–450. [Google Scholar] [CrossRef]
- Gaidamovich, S.Y.; Baten, M.A.; Klisenko, G.A.; Melnikova, Y.E. Serological studies on sandfly fevers in the Republic of Bangladesh. Acta Virol. 1984, 28, 325–328. [Google Scholar]
- Nikolaev, V.P.; Perepelkin, V.S.; Raevskiĭ, K.K.; Prusakova, Z.M. [A natural focus of sandfly fever in the Republic of Afghanistan]. Zh. Mikrobiol. Epidemiol. Immunobiol. 1991, 3, 39–41. [Google Scholar]
- Tesh, R.B.; Saidi, S.; Gajdamovic, S.J.; Rodhain, F.; Vesenjak-Hirjan, J. Serological studies on the epidemiology of sandfly fever in the Old World. Bull. World Health Organ. 1976, 54, 663–674. [Google Scholar] [PubMed]
- Chaste, L.C.; Launay, H.; Bailly-Choumara, H.; Le Lay, G.; Beaucournu, J.C. Infections à arbovirus au Maroc: Sondage sérologique chez les petits mammifères du nord du pays [Arbovirus infections in Morocco: Serosurvey in small wild mammals in the northern part of the country]. Bull. Soc. Pathol. Exot. Filiales 1982, 75, 466–475. [Google Scholar]
- Chastel, C.; Bach-Hamba, D.; Launay, H.; Le Lay, G.; Hellal, H.; Beaucournu, J.C. Infections à arbovirus en Tunisie: Nouvelle enquête sérologique chez les petits mammifères sauvages [Arbovirus infections in Tunisia: New serological survey of small wild mammals]. Bull. Soc. Pathol. Exot. Filiales 1983, 76, 21–33. [Google Scholar] [PubMed]
- Chastel, C.; Launay, H.; Rogues, G.; Beaucournu, J.C. Infections à arbovirus en Espagne: Enquête sérologique chez les petits mammifères [Arbovirus infections in Spain: Serological survey on small mammals]. Bull. Soc. Pathol. Exot. Filiales 1980, 73, 384–390. [Google Scholar] [PubMed]
- Le Lay-Roguès, G.; Valle, M.; Chastel, C.; Beaucournu, J.C. Petits mammifères sauvages et arbovirus en Italie [Small wild mammals and arboviruses in Italy]. Bull Soc Pathol Exot Filiales 1983, 76, 333–345. [Google Scholar] [PubMed]
- Alwassouf, S.; Christodoulou, V.; Bichaud, L.; Ntais, P.; Mazeris, A.; Antoniou, M.; Charrel, R.N. Seroprevalence of Sandfly-Borne Phleboviruses Belonging to Three Serocomplexes (Sandfly fever Naples, Sandfly fever Sicilian and Salehabad) in Dogs from Greece and Cyprus Using Neutralization Test. PLoS Negl. Trop. Dis. 2016, 10, e0005063. [Google Scholar] [CrossRef] [PubMed]
- Sakhria, S.; Alwassouf, S.; Fares, W.; Bichaud, L.; Dachraoui, K.; Alkan, C.; Zoghlami, Z.; de Lamballerie, X.; Zhioua, E.; Charrel, R.N. Presence of sandfly-borne phleboviruses of two antigenic complexes (Sandfly fever Naples virus and Sandfly fever Sicilian virus) in two different bio-geographical regions of Tunisia demonstrated by a microneutralisation-based seroprevalence study in dogs. Parasit. Vectors 2014, 7, 476. [Google Scholar] [CrossRef]
- Ayhan, N.; Sherifi, K.; Taraku, A.; Bërxholi, K.; Charrel, R.N. High Rates of Neutralizing Antibodies to Toscana and Sandfly Fever Sicilian Viruses in Livestock, Kosovo. Emerg. Infect. Dis. 2017, 23, 989–992. [Google Scholar] [CrossRef]
- Filipe, A.R. Serological survey for antibodies to arboviruses in the human population of Portugal. Trans. R. Soc. Trop. Med. Hyg. 1974, 68, 311–314. [Google Scholar] [CrossRef]
- Guerra, A.B.; Gouveia, C.; Zé, L.; Amaro, F.; Ferreira, G.C.; Brito, M.J. Prolonged Febrile Illness Caused by Sicilian Virus Infection in Portugal. In Proceedings of the 36th Annual meeting of the European Society for Paediatric Infectious Diseases, Malmo, Sweden, 28 May–2 June 2018. [Google Scholar]
- Amaro, F.; Zé-Zé, L.; Luz, M.T.; Alves, M.J. Toscana Virus: Ten Years of Diagnostics in Portugal. Acta Med. Port. 2021, 34, 677–681. [Google Scholar] [CrossRef]
- Maia, C.; Ayhan, N.; Cristovão, J.M.; Pereira, A.; Charrel, R. Human seroprevalence of Toscana virus and Sicilian phlebovirus in the southwest of Portugal. Eur. J. Clin. Microbio.l Infect. Dis. 2022, 41, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Alwassouf, S.; Maia, C.; Ayhan, N.; Coimbra, M.; Cristovão, J.M.; Richet, H.; Bichaud, L.; Campino, L.; Charrel, R.N. Neutralization-based seroprevalence of Toscana virus and sandfly fever Sicilian virus in dogs and cats from Portugal. J. Gen. Virol. 2016, 97, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Ayhan, N.; Cristóvão, J.M.; Vilhena, H.; Martins, Â.; Cachola, P.; Henriques, J.; Coimbra, M.; Catarino, A.; Lestinova, T.; et al. Antibody response to Toscana Virus and Sandfly Fever Sicilian Virus in cats naturally exposed to phlebotomine Sand Fly bites in Portugal. Microorganisms 2019, 7, 339. [Google Scholar] [CrossRef]
- Maia, C.; Alwassouf, S.; Cristóvão, J.M.; Ayhan, N.; Pereira, A.; Charrel, R.N.; Campino, L. Serological association between Leishmania infantum and sand fly fever Sicilian (but not Toscana) virus in sheltered dogs from southern Portugal. Parasit. Vectors 2017, 10, 92. [Google Scholar] [CrossRef]
- Verani, P.; Ciufolini, M.G.; Caciolli, S.; Renzi, A.; Nicoletti, L.; Sabatinelli, G.; Bartolozzi, D.; Volpi, G.K.; Amaducci, L.; Coluzzi, M. Ecology of viruses isolated from sand flies in Italy and characterized of a new Phlebovirus (Arbia virus). Am. J. Trop. Med. Hyg. 1988, 38, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Leoncini, F.; Bartolozzi, D.; Banchi, S. Il virus Toscana: Un nuovo Phlebovirus causa di malattie infiammatorie acute del SNC nell’uomo. Giorn. Mal. Inf. Parass. 1986, 38, 649–652. [Google Scholar]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sánchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emer. Infec. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef]
- Charrel, R.N.; Bichaud, L.; de Lamballerie, X. Emergence of Toscana virus in the mediterranean area. World J. Virol. 2012, 1, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Laroche, L.; Jourdain, F.; Ayhan, N.; Bañuls, A.L.; Charrel, R.; Prudhomme, J. Incubation Period for Neuroinvasive Toscana Virus Infections. Emerg. Infect. Dis. 2021, 27, 3147–3150. [Google Scholar] [CrossRef]
- Pierro, A.; Ficarelli, S.; Ayhan, N.; Morini, S.; Raumer, L.; Bartoletti, M.; Mastroianni, A.; Prati, F.; Schivazappa, S.; Cenni, P.; et al. Characterization of antibody response in neuroinvasive infection caused by Toscana virus. Clin. Microbiol. Infect. 2017, 23, 868–873. [Google Scholar] [CrossRef]
- Ergunay, K.; Kaplan, B.; Okar, S.; Akkutay-Yoldar, Z.; Kurne, A.; Arsava, E.M.; Ozkul, A. Urinary detection of toscana virus nucleic acids in neuroinvasive infections. J. Clin. Virol. 2015, 70, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Matusali, G.; D’Abramo, A.; Terrosi, C.; Carletti, F.; Colavita, F.; Vairo, F.; Savellini, G.G.; Gandolfo, C.; Anichini, G.; Lalle, E.; et al. Infectious Toscana Virus in Seminal Fluid of Young Man Returning from Elba Island, Italy. Emer. Infect. Dis. 2022, 28, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Arboviral Diseases, Neuroinvasive and Non-Neuroinvasive. 2015 Case Definition. Available online: https://ndc.services.cdc.gov/case-definitions/arboviral-diseases-neuroinvasive-and-non-neuroinvasive-2015/ (accessed on 8 June 2022).
- Bartels, S.; de Boni, L.; Kretzschmar, H.A.; Heckmann, J.G. Lethal encephalitis caused by the Toscana virus in an elderly patient. J. Neurol. 2012, 259, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Popescu, C.P.; Cotar, A.I.; Dinu, S.; Zaharia, M.; Tardei, G.; Ceausu, E.; Badescu, D.; Ruta, S.; Ceianu, C.S.; Florescu, S.A. Emergence of Toscana Virus, Romania, 2017–2018. Emerg. Infect. Dis. 2021, 27, 1482–1485. [Google Scholar] [CrossRef]
- Rota, E.; Morelli, N.; Immovilli, P.; De Mitri, P.; Guidetti, D. Guillain-Barré-like axonal polyneuropathy associated with Toscana virus infection: A case report. Medicine (Baltimore). 2017, 96, e8081. [Google Scholar] [CrossRef]
- Okar, S.V.; Bekircan-Kurt, C.E.; Hacıoğlu, S.; Erdem-Özdamar, S.; Özkul, A.; Ergünay, K. Toscana virus associated with Guillain-Barré syndrome: A case-control study. Acta Neurol. Belg. 2021, 121, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, F.; Ciufolini, M.G.; Francisci, D.; Marchi, A.; Venturi, G.; Fiorentini, C.; Luchetta, M.L.; Bruto, L.; Pauluzzi, S. Unusual presentation of life-threatening Toscana virus meningoencephalitis. Clin. Infect. Dis. 2004, 38, 515–520. [Google Scholar] [CrossRef]
- Oechtering, J.; Petzold, G.C. Acute hydrocephalus due to impaired CSF resorption in Toscana virus meningoencephalitis. Neurology 2012, 79, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, F.A.; Moreno-Docón, A.; Segovia-Hernández, M.; Fernández-Barreiro, A. Sordera como secuela de meningitis por el virus Toscana [Deafness as a sequela of Toscana virus meningitis]. Med. Clin. 2008, 130, 639. (In Spanish) [Google Scholar] [CrossRef]
- Sanbonmatsu-Gámez, S.; Pérez-Ruiz, M.; Palop-Borrás, B.; Navarro-Marí, J.M. Unusual manifestation of toscana virus infection, Spain. Emerg. Infect. Dis. 2009, 15, 347–348. [Google Scholar] [CrossRef]
- Mosnier, E.; Charrel, R.; Vidal, B.; Ninove, L.; Schleinitz, N.; Harlé, J.R.; Bernit, E. Toscana virus myositis and fasciitis. Med. Mal. Infect. 2013, 43, 208–210. [Google Scholar] [CrossRef]
- Marlinge, M.; Crespy, L.; Zandotti, C.; Piorkowski, G.; Kaphan, E.; Charrel, R.N.; Ninove, L. Afebrile meningoencephalitis with transient central facial paralysis due to Toscana virus infection, southeastern France, 2014 [corrected]. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. Eur. Commun. Dis. Bull. 2014, 19, 20974. [Google Scholar] [CrossRef]
- Serata, D.; Rapinesi, C.; Del Casale, A.; Simonetti, A.; Mazzarini, L.; Ambrosi, E.; Kotzalidis, G.D.; Fensore, C.; Girardi, P.; Tatarelli, R. Personality changes after Toscana virus (TOSV) encephalitis in a 49-year-old man: A case report. Intern. J. Neurosc. 2011, 121, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Zanelli, G.; Bianco, C.; Cusi, M.G. Testicular involvement during Toscana virus infection: An unusual manifestation? Infection 2013, 41, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Mascitti, H.; Calin, R.; Dinh, A.; Makhloufi, S.; Davido, B. Testicular pain associated with clear fluid meningitis: How many cases of Toscana virus are we missing? Int. J. Infect. Dis. 2020, 93, 198–200. [Google Scholar] [CrossRef]
- Ayhan, N.; Charrel, R.N. An update on Toscana virus distribution, genetics, medical and diagnostic aspects. Clin. Microbiol Infect. 2020, 26, 1017–1023. [Google Scholar] [CrossRef]
- Sun, M.H.; Ji, Y.F.; Li, G.H.; Shao, J.W.; Chen, R.X.; Gong, H.Y.; Chen, S.Y.; Chen, J.M. Highly adaptive Phenuiviridae with biomedical importance in multiple fields. J. Med. Virol. 2022, 94, 2388–2401. [Google Scholar] [CrossRef]
- Sakhria, S.; Bichaud, L.; Mensi, M.; Salez, N.; Dachraoui, K.; Thirion, L.; Cherni, S.; Chelbi, I.; De Lamballerie, X.; Zhioua, E.; et al. Co-circulation of Toscana virus and Punique virus in northern Tunisia: A microneutralisation-based seroprevalence study. PLoS Negl. Trop. Dis. 2013, 7, e2429. [Google Scholar] [CrossRef]
- Valassina, M.; Valentini, M.; Pugliese, A.; Valensin, P.E.; Cusi, M.G. Serological survey of Toscana virus infections in a high-risk population in Italy. Clin. Diagn. Lab. Immunol. 2003, 10, 483–484. [Google Scholar] [CrossRef]
- Punda-Polić, V.; Jerončić, A.; Mohar, B.; Šiško Kraljević, K. Prevalence of Toscana virus antibodies in residents of Croatia. Clin. Microbiol. Infect. 2012, 18, E200–E203. [Google Scholar] [CrossRef]
- Tesh, R.B.; Lubroth, J.; Guzman, H. Simulation of arbovirus overwintering: Survival of Toscana virus (Bunyaviridae:Phlebovirus) in its natural sand fly vector Phlebotomus perniciosus. Am. J. Trop. Med. Hyg. 1992, 47, 574–581. [Google Scholar] [CrossRef]
- Ciufolini, M.G.; Maroli, M.; Guandalini, E.; Marchi, A.; Verani, P. Experimental studies on the maintenance of Toscana and Arbia viruses (Bunyaviridae: Phlebovirus). Am. J. Trop. Med. Hyg. 1989, 40, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Modi, G.B. Maintenance of Toscana virus in Phlebotomus perniciosus by vertical transmission. Am. J. Trop. Med. Hyg. 1987, 36, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Dachraoui, K.; Piorkowski, G.; Chelbi, I.; Moureau, G.; Cherni, S.; De Lamballerie, X.; Sakhria, S.; Charrel, R.N.; Zhioua, E. Toscana virus isolated from sandflies, Tunisia. Emerg. Infect. Dis. 2013, 19, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Hacioglu, S.; Dincer, E.; Isler, C.T.; Karapinar, Z.; Ataseven, V.S.; Ozkul, A.; Ergunay, K. A Snapshot Avian Surveillance Reveals West Nile Virus and Evidence of Wild Birds Participating in Toscana Virus Circulation. Vector Borne Zoonotic Dis. 2017, 17, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Marí, J.M.; Palop-Borrás, B.; Pérez-Ruiz, M.; Sanbonmatsu-Gámez, S. Serosurvey study of Toscana virus in domestic animals, Granada, Spain. Vector Borne Zoonotic Dis. 2011, 11, 583–587. [Google Scholar] [CrossRef]
- Dahmani, M.; Alwassouf, S.; Grech-Angelini, S.; Marié, J.L.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Corsica, France. Parasit. Vectors 2016, 9, 381. [Google Scholar] [CrossRef]
- Tahir, D.; Alwassouf, S.; Loudahi, A.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Kabylia (Algeria). Clin. Microbiol. Infect. 2016, 22, e16–e17. [Google Scholar] [CrossRef]
- Ehrnst, A.; Peters, C.J.; Niklasson, B.; Svedmyr, A.; Holmgren, B. Neurovirulent Toscana virus (a sandfly fever virus) in Swedish man after visit to Portugal. Lancet 1985, 1, 1212–1213. [Google Scholar] [CrossRef]
- Schwarz, T.F.; Jäger, G.; Gilch, S.; Pauli, C. Serosurvey and laboratory diagnosis of imported sandfly fever virus, serotype Toscana, infection in Germany. Epidemiol. Infect. 1995, 114, 501–510. [Google Scholar] [CrossRef]
- Santos, L.; Simões, J.; Costa, R.; Martins, S.; Lecour, H. Toscana virus meningitis in Portugal, 2002-2005. Euro Surveill. 2007, 12, E3–E4. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Cardoso, M.J.; Marinho, A.S.; Guimarães, T.; Sarmento, A. Seroprevalence survey of Toscana virus infection in Oporto region. Acta Med. Port. 2011, 24, 479–482. (In Portuguese) [Google Scholar]
- Amaro, F.; Luz, T.; Parreira, P.; Ciufolini, M.G.; Marchi, A.; Janeiro, N.; Zagalo, A.; Proença, P.; Ramos, M.I.; Alves, M.J. Toscana virus in the Portuguese population: Serosurvey and clinical cases. Acta Med. Port. 2011, 24 (Suppl. 2), 503–508. (In Portuguese) [Google Scholar] [PubMed]
- Amaro, F. Phlebovirus, Importância em Saúde Pública em Portugal. Ph.D. Thesis, Faculty of Sciences, University of Lisbon, Lisbon, Portugal, 14 December 2010. (In Portuguese). [Google Scholar]
- Alho, A.M.; Pita, J.; Amaro, A.; Amaro, F.; Schnyder, M.; Grimm, F.; Custódio, A.C.; Cardoso, L.; Deplazes, P.; de Carvalho, L.M. Seroprevalence of vector-borne pathogens and molecular detection of Borrelia afzelii in military dogs from Portugal. Parasit. Vectors 2016, 9, 225. [Google Scholar] [CrossRef]
- Charrel, R.N.; Moureau, G.; Temmam, S.; Izri, A.; Marty, P.; Parola, P.; da Rosa, A.T.; Tesh, R.B.; de Lamballerie, X. Massilia virus, a novel Phlebovirus (Bunyaviridae) isolated from sandflies in the Mediterranean. Vector Borne Zoonotic Dis. 2009, 9, 519–530. [Google Scholar] [CrossRef]
- Sanchez-Seco, M.P.; Vazquez, A.; Collao, X.; Hernandez, L.; Aranda, C.; Ruiz, S.; Escosa, R.; Marques, E.; Bustillo, M.A.; Molero, F.; et al. Surveillance of arboviruses in Spanish wetlands: Detection of new flavi- and phleboviruses. Vector Borne Zoonotic Dis. 2010, 10, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Collao, X.; Palacios, G.; de Ory, F.; Sanbonmatsu, S.; Pérez-Ruiz, M.; Navarro, J.M.; Molina, R.; Hutchison, S.K.; Lipkin, W.I.; Tenorio, A.; et al. Granada virus: A natural phlebovirus reassortant of the sandfly fever Naples serocomplex with low seroprevalence in humans. Am. J. Trop. Med. Hyg. 2010, 83, 760–765. [Google Scholar] [CrossRef]
- Navarro-Marí, J.M.; Gómez-Camarasa, C.; Pérez-Ruiz, M.; Sanbonmatsu-Gámez, S.; Pedrosa-Corral, I.; Jiménez-Valera, M. Clinic-epidemiologic study of human infection by Granada virus, a new phlebovirus within the sandfly fever Naples serocomplex. Am. J. Trop Med. Hyg. 2013, 88, 1003–1006. [Google Scholar] [CrossRef]
- Amaro, F.; Hanke, D.; Zé-Zé, L.; Alves, M.J.; Becker, S.C.; Höper, D. Genetic characterization of Arrabida virus, a novel phlebovirus isolated in South Portugal. Virus Res. 2016, 214, 19–25. [Google Scholar] [CrossRef]
- Amaro, F.; Zé-Zé, L.; Alves, M.J.; Börstler, J.; Clos, J.; Lorenzen, S.; Becker, S.C.; Schmidt-Chanasit, J.; Cadar, D. Co-circulation of a novel phlebovirus and Massilia virus in sandflies, Portugal. Virol. J. 2015, 12, 174. [Google Scholar] [CrossRef]
- Palacios, G.; Savji, N.; Travassos da Rosa, A.; Desai, A.; Sanchez-Seco, M.P.; Guzman, H.; Lipkin, W.I.; Tesh, R. Characterization of the Salehabad virus species complex of the genus Phlebovirus (Bunyaviridae). J. Gen. Virol. 2013, 94 (Pt 4), 837–842. [Google Scholar] [CrossRef]
- Alkan, C.; Alwassouf, S.; Piorkowski, G.; Bichaud, L.; Tezcan, S.; Dincer, E.; Ergunay, K.; Ozbel, Y.; Alten, B.; de Lamballerie, X.; et al. Isolation, genetic characterization, and seroprevalence of Adana virus, a novel phlebovirus belonging to the Salehabad virus complex, in Turkey. J. Virol. 2015, 89, 4080–4091. [Google Scholar] [CrossRef] [PubMed]
- Bichaud, L.; Dachraoui, K.; Alwassouf, S.; Alkan, C.; Mensi, M.; Piorkowski, G.; Sakhria, S.; Seston, M.; Fares, W.; De Lamballerie, X.; et al. Isolation, full genomic characterization and neutralization-based human seroprevalence of Medjerda Valley virus, a novel sandfly-borne phlebovirus belonging to the Salehabad virus complex in northern Tunisia. J. Gen. Virol. 2016, 97, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Velo, E.; Bino, S. A novel phlebovirus in Albanian sandflies. Clin. Microbiol. Infect. 2011, 17, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Anagnostou, V.; Pardalos, G.; Athanasiou-Metaxa, M.; Papa, A. Novel phlebovirus in febrile child, Greece. Emerg. Infect. Dis. 2011, 17, 940–941. [Google Scholar] [CrossRef]
- Lambert, A.J.; Lanciotti, R.S. Consensus amplification and novel multiplex sequencing method for S segment species identification of 47 viruses of the Orthobunyavirus, Phlebovirus, and Nairovirus genera of the family Bunyaviridae. J. Clin. Microbiol. 2009, 47, 2398–2404. [Google Scholar] [CrossRef]
- Sánchez-Seco, M.P.; Echevarría, J.M.; Hernández, L.; Estévez, D.; Navarro-Marí, J.M.; Tenorio, A. Detection and identification of Toscana and other phleboviruses by RT-nested-PCR assays with degenerated primers. J. Med. Virol. 2003, 71, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2020. [Google Scholar]
- Ayhan, N.; Charrel, R.N. Of phlebotomines (sandflies) and viruses: A comprehensive perspective on a complex situation. Curr. Opin. Insect. Sci. 2017, 22, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Ergunay, K.; Ayhan, N.; Charrel, R.N. Novel and emergent sandfly-borne phleboviruses in Asia Minor: A systematic review. Rev. Med. Virol. 2017, 27, e1898. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Weinberg, A.N.; Muth, D.J.; Lazuick, J.S. Toscana virus infection in United States citizen returning from Italy. Lancet 1987, 329, 165–166. [Google Scholar] [CrossRef]
- Sonderegger, B.; Hachler, H.; Dobler, G.; Frei, M. Imported aseptic meningitis due to Toscana virus acquired on the island of Elba, Italy, August 2008. Euro Surveill. 2009, 14, 19079. [Google Scholar] [CrossRef]
- Schultze, D.; Korte, W.; Rafeiner, P.; Niedrig, M. First report of sandfly fever virus infection imported from Malta into Switzerland, October 2011. Euro Surveill. 2012, 17, 20209. [Google Scholar] [CrossRef]
- Arden, K.E.; Heney, C.; Shaban, B.; Nimmo, G.R.; Nissen, M.D.; Sloots, T.P.; Mackay, I.M. Detection of Toscana virus from an adult traveler returning to Australia with encephalitis. J. Med. Virol. 2017, 89, 1861–1864. [Google Scholar] [CrossRef]
- Oerther, S.; Jöst, H.; Heitmann, A.; Lühken, R.; Krüger, A.; Steinhausen, I.; Brinker, C.; Lorentz, S.; Marx, M.; Schmidt-Chanasit, J.; et al. Phlebotomine sand flies in Southwest Germany: An update with records in new locations. Parasit. Vectors 2020, 13, 173. [Google Scholar] [CrossRef]
- Grimm, F.; Gessler, M.; Jenni, L. Aspects of sandfly biology in southern Switzerland. Med. Vet. Entomol. 1993, 7, 170–176. [Google Scholar] [CrossRef]
- Poeppl, W.; Obwaller, A.G.; Weiler, M.; Burgmann, H.; Mooseder, G.; Lorentz, S.; Rauchenwald, F.; Aspöck, H.; Walochnik, J.; Naucke, T.J. Emergence of sandflies (Phlebotominae) in Austria, a Central European country. Parasitol. Res. 2013, 112, 4231–4237. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Van Bortel, W.; Zeller, H.; Alten, B. A summary of the evidence for the change in European distribution of phlebotomine sand flies (Diptera: Psychodidae) of public health importance. J. Vector Ecol. 2014, 39, 72–77. [Google Scholar] [CrossRef]
- La Bella, S.; Scorrano, G.; Greco, R.; Trotta, D.; Aricò, M. Cyprus virus infection: A rare but life-threatening diagnosis in children. Pediatr. Infect. Dis. J. 2022, 41, e299–e300. [Google Scholar] [CrossRef] [PubMed]
- Sanbonmatsu-Gámez, S.; Pedrosa-Corral, I.; Navarro-Marí, J.M.; Pérez-Ruiz, M. Update in Diagnostics of Toscana Virus Infection in a Hyperendemic Region (Southern Spain). Viruses 2021, 13, 1438. [Google Scholar] [CrossRef] [PubMed]
- Collao, X.; Palacios, G.; Sanbonmatsu-Gámez, S.; Pérez-Ruiz, M.; Negredo, A.I.; Navarro-Marí, J.M.; Grandadam, M.; Aransay, A.M.; Lipkin, W.I.; Tenorio, A.; et al. Genetic diversity of Toscana virus. Emerg. Infect. Dis. 2009, 15, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.Y.; Tesh, R.B.; Travassos da Rosa, A.P.A.; Peters, C.J.; Yang, Z.; Guzman, H.; Xiao, S.Y. Phylogenetic relationships among members of the genus Phlebovirus (Bunyaviridae) based on partial M segment sequence analyses. J. Gen. Virol. 2003, 84 (Pt 2), 465–473. [Google Scholar] [CrossRef]
- Venturi, G.; Madeddu, G.; Rezza, G.; Ciccozzi, M.; Pettinato, M.L.; Cilliano, M.; Fiorentini, C.; Mura, M.S.; Ciufolini, M.G. Detection of Toscana virus central nervous system infections in Sardinia Island, Italy. J. Clin. Virol. 2007, 40, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.; Ayhan, N.; Ortuño, M.; Ortiz, J.; Gould, E.A.; Maia, C.; Berriatua, E.; Charrel, R.N. Experimental infection of dogs with Toscana Virus and Sandfly Fever Sicilian Virus to determine their potential as possible vertebrate Hosts. Microorganisms 2020, 8, 596. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Maximum likelihood phylogenetic tree of phleboviruses’ partial nucleocapsid nucleotide sequences (segment S, 674 positions in the final dataset) using Kimura 2-parameter model and discrete gamma distribution. Sequence names in blue (Massilia phlebovirus), in red (Alcube phlebovirus), and yellow (New phlebovirus?) were detected in Portugal (PT). Sequence names marked with a colored bullet have been sequenced in this study. Bootstrap support over 60% is presented on branches. Used sequence alignment is available in Supplementary File S1: Phleboviruses’ nucleotide partial sequence alignment of S segments. Names of the sequences in the tree match the IDs in the alignment made available as a Supplementary File.
Figure 1.
Maximum likelihood phylogenetic tree of phleboviruses’ partial nucleocapsid nucleotide sequences (segment S, 674 positions in the final dataset) using Kimura 2-parameter model and discrete gamma distribution. Sequence names in blue (Massilia phlebovirus), in red (Alcube phlebovirus), and yellow (New phlebovirus?) were detected in Portugal (PT). Sequence names marked with a colored bullet have been sequenced in this study. Bootstrap support over 60% is presented on branches. Used sequence alignment is available in Supplementary File S1: Phleboviruses’ nucleotide partial sequence alignment of S segments. Names of the sequences in the tree match the IDs in the alignment made available as a Supplementary File.

Figure 2.
Maximum likelihood phylogenetic tree of phleboviruses partial glycoprotein precursor nucleotide sequences (segment M, 461 positions in the final dataset) using Tamura 3-parameter model and discrete gamma distribution. Sequence names in blue (Massilia phlebovirus) were detected in Portugal (PT). Sequence names marked with a colored bullet have been sequenced in this study. Bootstrap support over 60% is presented on branches. Used sequence alignment is available in Supplementary File S2: Phleboviruses’ nucleotide partial sequence alignment of M segments. Names of the sequences in the tree match the IDs in the alignment made available as a Supplementary File.
Figure 2.
Maximum likelihood phylogenetic tree of phleboviruses partial glycoprotein precursor nucleotide sequences (segment M, 461 positions in the final dataset) using Tamura 3-parameter model and discrete gamma distribution. Sequence names in blue (Massilia phlebovirus) were detected in Portugal (PT). Sequence names marked with a colored bullet have been sequenced in this study. Bootstrap support over 60% is presented on branches. Used sequence alignment is available in Supplementary File S2: Phleboviruses’ nucleotide partial sequence alignment of M segments. Names of the sequences in the tree match the IDs in the alignment made available as a Supplementary File.

Figure 3.
Maximum likelihood phylogenetic trees of phleboviruses partial RNA-dependent RNA polymerase nucleotide sequences (segment L). Sequence names in blue (Massilia phlebovirus) and in red (Alcube phlebovirus) were detected in Portugal (PT). Sequence names marked with a colored bullet have been sequenced in this study. Bootstrap support over 60% is presented on branches. Names of the sequences in the tree match the IDs in the alignment made available as a Supplementary File. (a) Maximum likelihood phylogenetic tree of phleboviruses partial RNA-dependent RNA polymerase nucleotide sequences (segment L, 227 positions in the final dataset) using Tamura 3-parameter model and discrete gamma distribution. Used sequence alignment is available in Supplementary File S3: (a) Phleboviruses’ nucleotide partial sequence alignment of L segments. (b) Maximum likelihood phylogenetic tree of phleboviruses partial RNA-dependent RNA polymerase nucleotide sequences (segment L, 1065 positions in the final dataset) using general time reversible model and discrete gamma distribution. Used sequence alignment is available in Supplementary File S4: (b) Phleboviruses’ nucleotide partial sequence alignment of L segments.
Figure 3.
Maximum likelihood phylogenetic trees of phleboviruses partial RNA-dependent RNA polymerase nucleotide sequences (segment L). Sequence names in blue (Massilia phlebovirus) and in red (Alcube phlebovirus) were detected in Portugal (PT). Sequence names marked with a colored bullet have been sequenced in this study. Bootstrap support over 60% is presented on branches. Names of the sequences in the tree match the IDs in the alignment made available as a Supplementary File. (a) Maximum likelihood phylogenetic tree of phleboviruses partial RNA-dependent RNA polymerase nucleotide sequences (segment L, 227 positions in the final dataset) using Tamura 3-parameter model and discrete gamma distribution. Used sequence alignment is available in Supplementary File S3: (a) Phleboviruses’ nucleotide partial sequence alignment of L segments. (b) Maximum likelihood phylogenetic tree of phleboviruses partial RNA-dependent RNA polymerase nucleotide sequences (segment L, 1065 positions in the final dataset) using general time reversible model and discrete gamma distribution. Used sequence alignment is available in Supplementary File S4: (b) Phleboviruses’ nucleotide partial sequence alignment of L segments.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Phleboviruses detected and/or isolated from sandfly pools in Portugal.
| Designation in GenBank | Collection Date | Collection County | S Segment (nr of bp/) | M Segment (nr of bp) | L Segment (nr of bp) | Accession Number | Species ID | Reference |
|---|---|---|---|---|---|---|---|---|
| Alcube virus strain S20 1 | July 2007 | Setúbal | 1758 | 4164 | 6405 | KR363190-192 | Alcube phlebovirus | [110] |
| PoSFPhlebV/21/2007 1 | July 2007 | Setúbal | 1854 | 4221 | 6404 | MW448250-252 | Massilia phlebovirus | [4] |
| PoSFPhlebV/54/2007 1 | August 2007 | Setúbal | 608 | - | 227 | ON807179-180 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/70/2007 2 | September 2007 | Setúbal | 1873 | 4229 | 6386 | MW448253-255 | Massilia phlebovirus | [4] |
| PoSFPhlebV/112/2008 2 | May 2008 | Setúbal | 547 | 461 | 224 | ON807181-183 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/118/2008 2 | June 2008 | Setúbal | 670 | 461 | 227 | ON807184 | Massilia phlebovirus * | This publication |
| Arrabida virus strain PoSFPhlebV/126/2008 1 | June 2008 | Setúbal | 1840 | 4198 | 6391 | KP863799-801 | Massilia phlebovirus | [109] |
| Massilia virus strain 127 1 | June 2008 | Setúbal | 1864 | 4225 | 6404 | KT906098, 100, 102 | Massilia phlebovirus | [110] |
| PoSFPhlebV/128/2008 1 | June 2008 | Setúbal | 310 | - | - | ON807187 | Alcube phlebovirus * | This publication |
| Massilia virus strain 130 3 | June 2008 | Olhão | 1864 | 4225 | 6404 | KT906099, 101, 103 | Massilia phlebovirus | [110] |
| PoSFPhlebV/134/2008 3 | June 2008 | Olhão | 338 | - | - | ON807188 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/137/2008 3 | June 2008 | Olhão | 653 | - | 212 | ON807189-190 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/138/2008 3 | June 2008 | Olhão | 670 | - | - | ON807191 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/149/2008 4 | June 2008 | Olhão | 392 | 461 | 1041 | ON807192-194 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/171/2008 5 | July 2008 | Setúbal | 333 | 461 | - | ON807195-196 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/220/2008 6 | August 2008 | Tavira | 631 | - | - | ON807197 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/237/2008 4 | September 2008 | Olhão | 452 | - | - | ON807198 | Massilia phlebovirus * | This publication |
| PoSFPhlebV/11/2018 7 | May 2018 | Tavira | 381 | - | - | ON807199 | New phlebovirus? | This publication |
| PoSFPhlebV/38/2018 7 | June 2018 | Tavira | 530 | - | - | ON807200 | Massilia phlebovirus * | This publication |
1 Sheep pen in a farm; 2 hennery near a country house; 3 pigeonry near a country house; 4 hennery, near a country house; 5 kennel in a dog shelter; 6 hennery in a farm and 7 hennery near a country house. Strains with the same superscript number were collected at the same location. * Presumptive species ID based on available sequence analysis.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Amaro, F.; Zé-Zé, L.; Alves, M.J. Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting. Viruses 2022, 14, 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081768
AMA Style
Amaro F, Zé-Zé L, Alves MJ. Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting. Viruses. 2022; 14(8):1768. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081768
Chicago/Turabian StyleAmaro, Fátima, Líbia Zé-Zé, and Maria João Alves. 2022. "Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting" Viruses 14, no. 8: 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081768
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.