
Baculovirus Display of Varicella–Zoster Virus Glycoprotein E Induces Robust Humoral and Cellular Immune Responses in Mice

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell, Virus Stock, Reagents and Antibodies
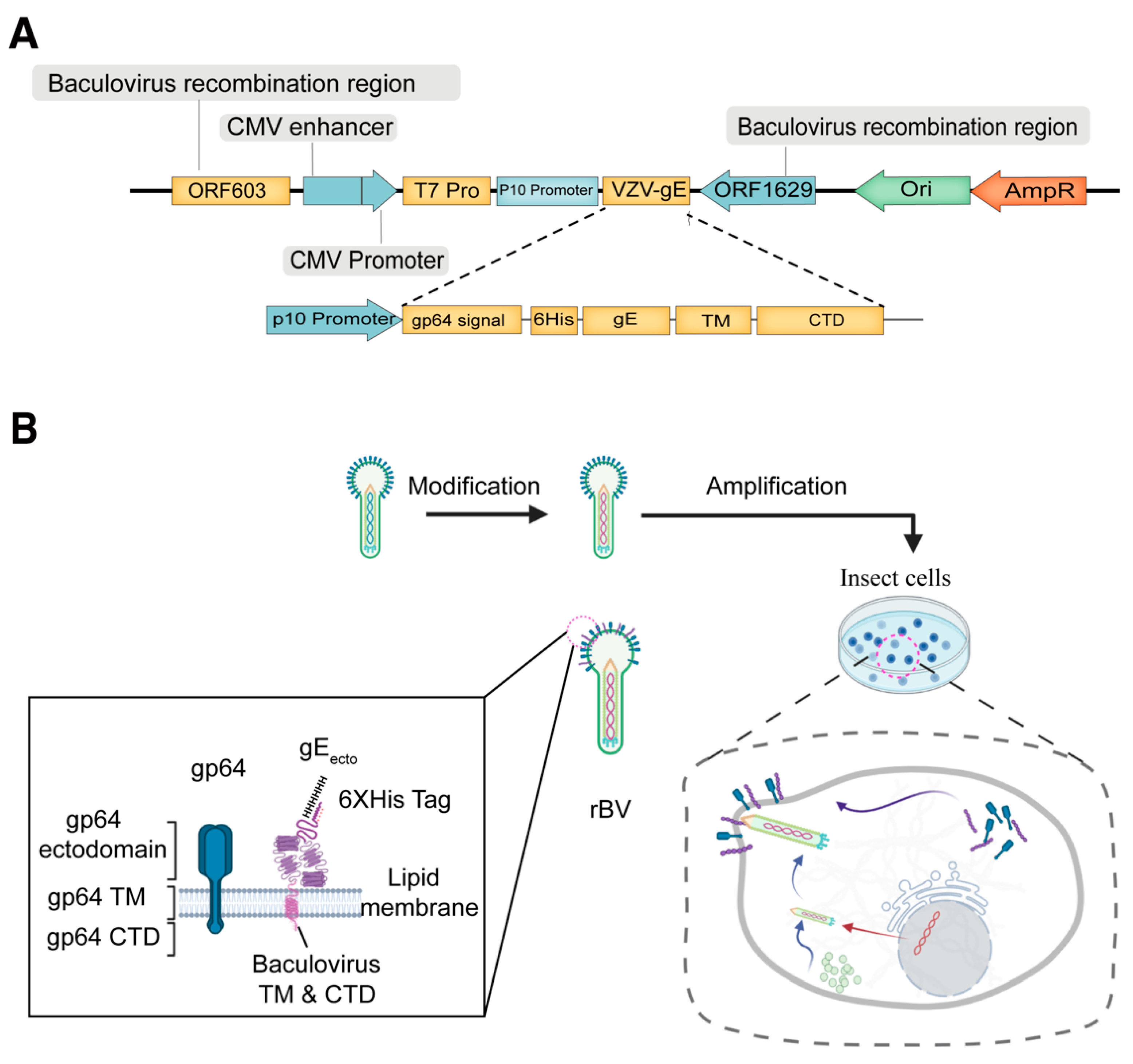
2.2. Construction of Recombinant Baculovirus
2.3. gE Plasmids Construction and Protein Purification
2.4. Purification and Titration of the Recombinant Baculovirus
2.5. Western Blotting
2.6. Immunofluorescence Assay (IFA)
2.7. Flow Cytometry
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Immune Electron Microscopy (IEM)
2.10. Immunization of BALB/c Mice
2.11. Neutralization Assay
2.12. Enzyme-Linked Immunospot Assay (ELISPOT)
2.13. Intracellular Cytokine Staining (ICS) and Flow Cytometry
2.14. Statistical Analysis
3. Results
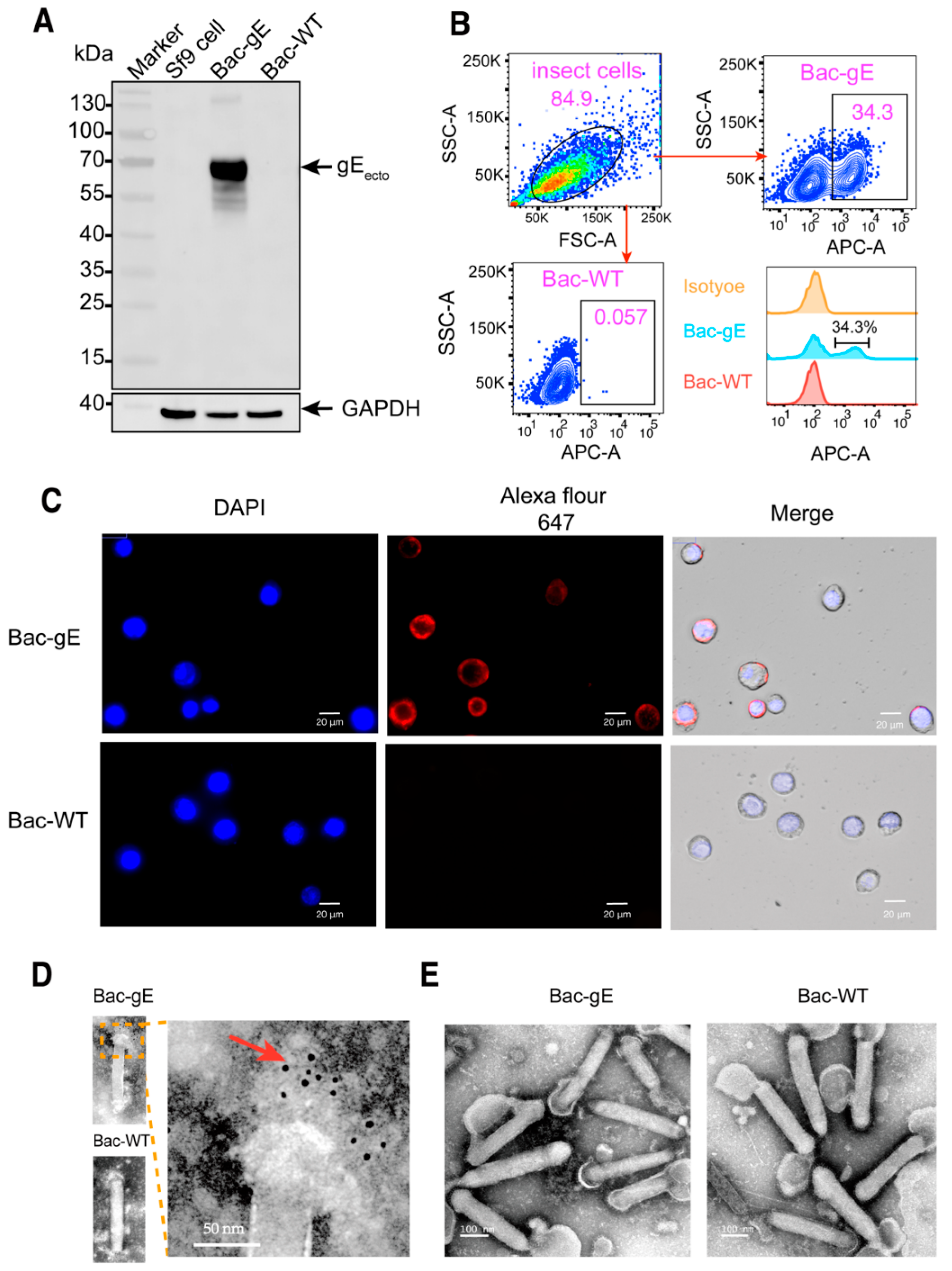
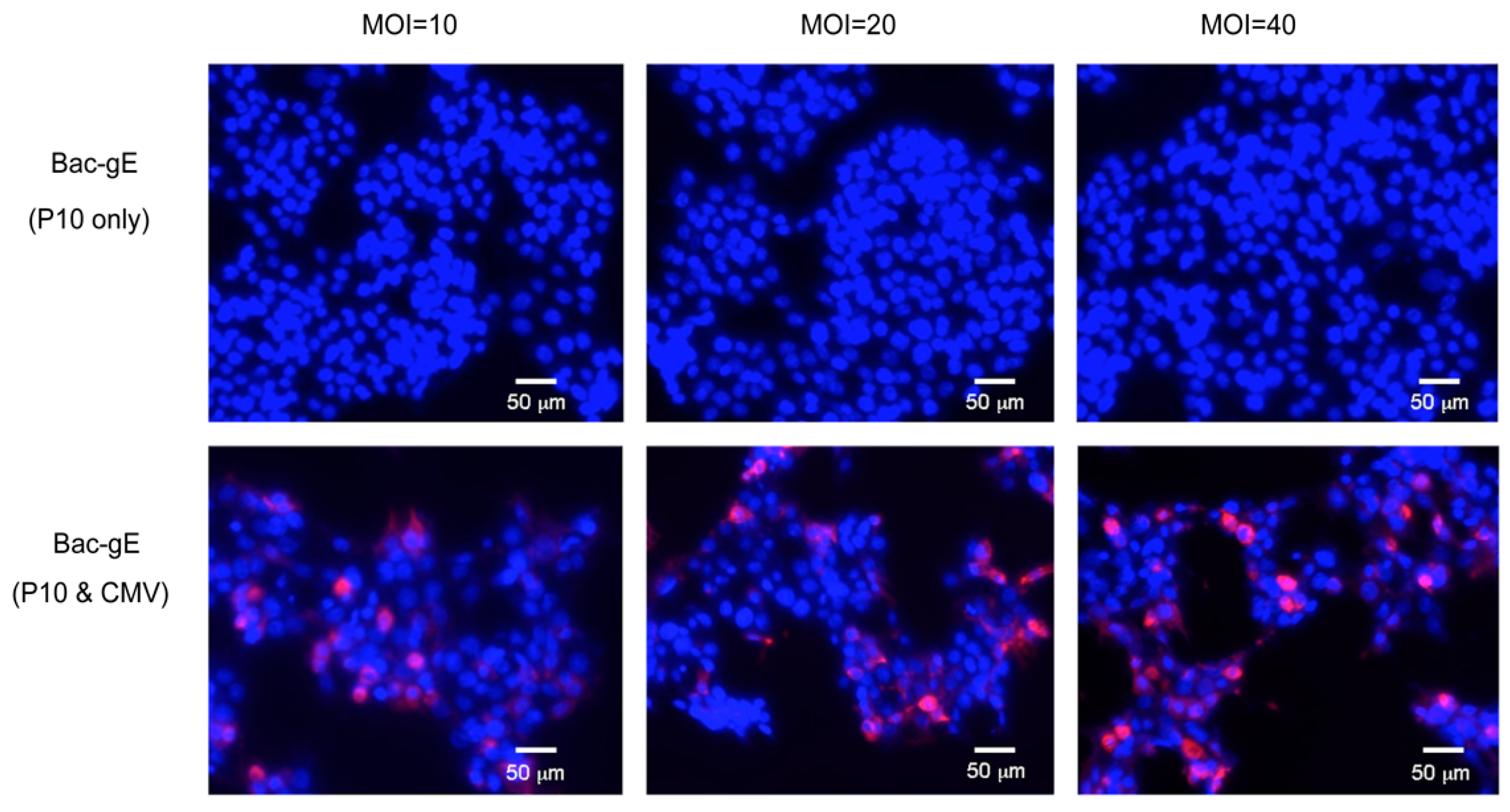
3.1. Generation of a Recombinant Baculovirus Displaying the gE Protein
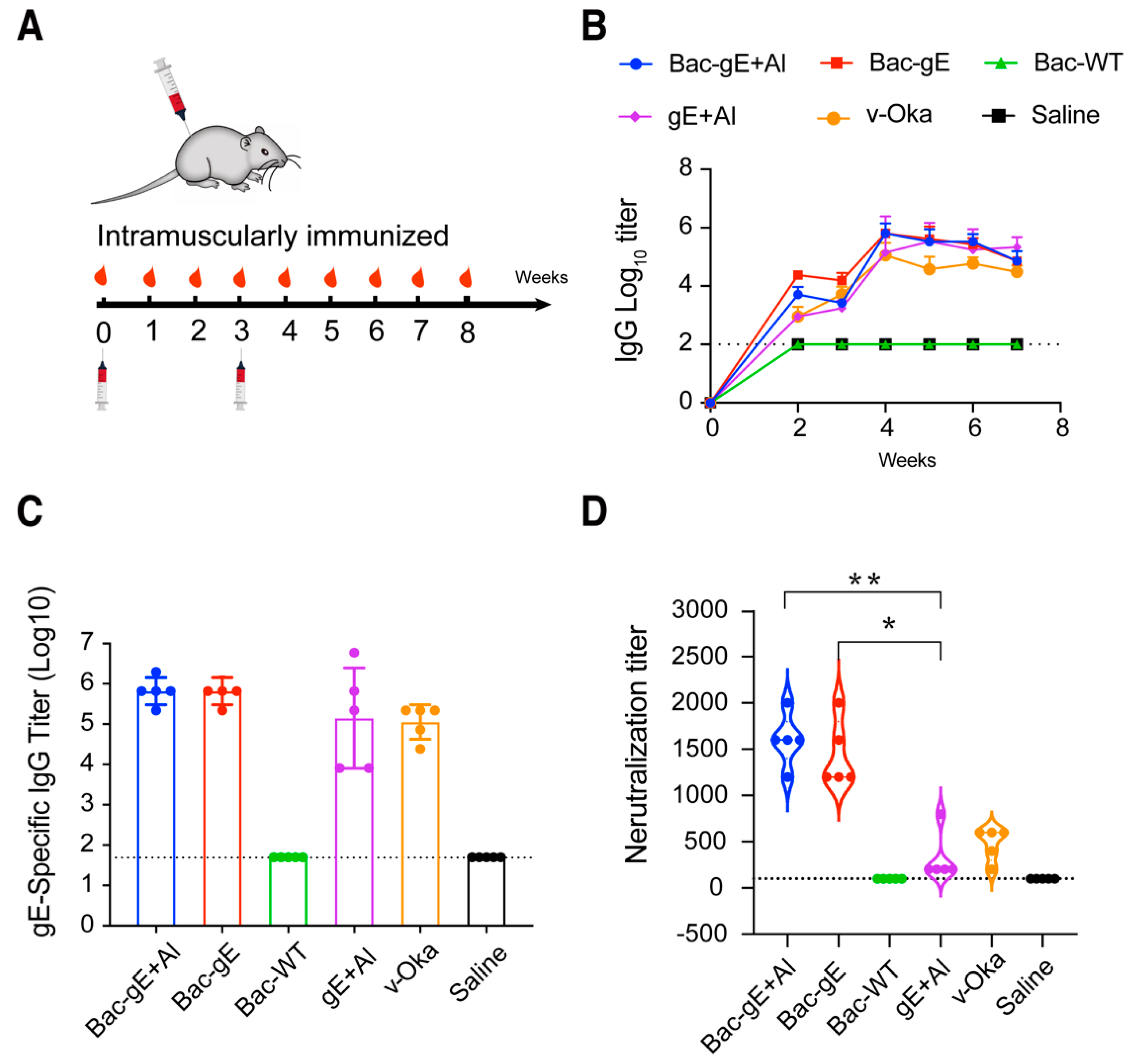
3.2. Bac–gE Elicits a Robust Humoral Immune Response in Mice
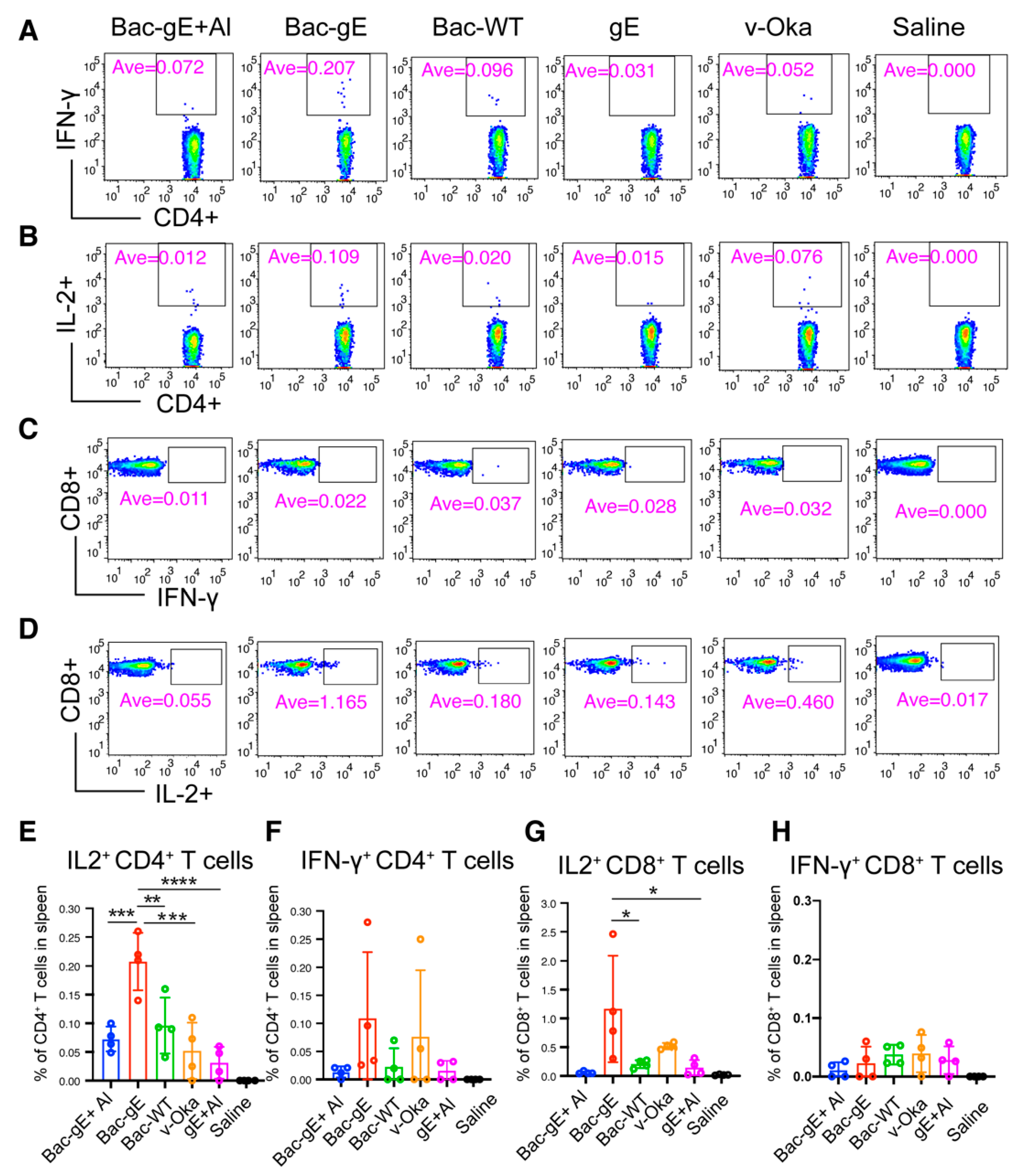
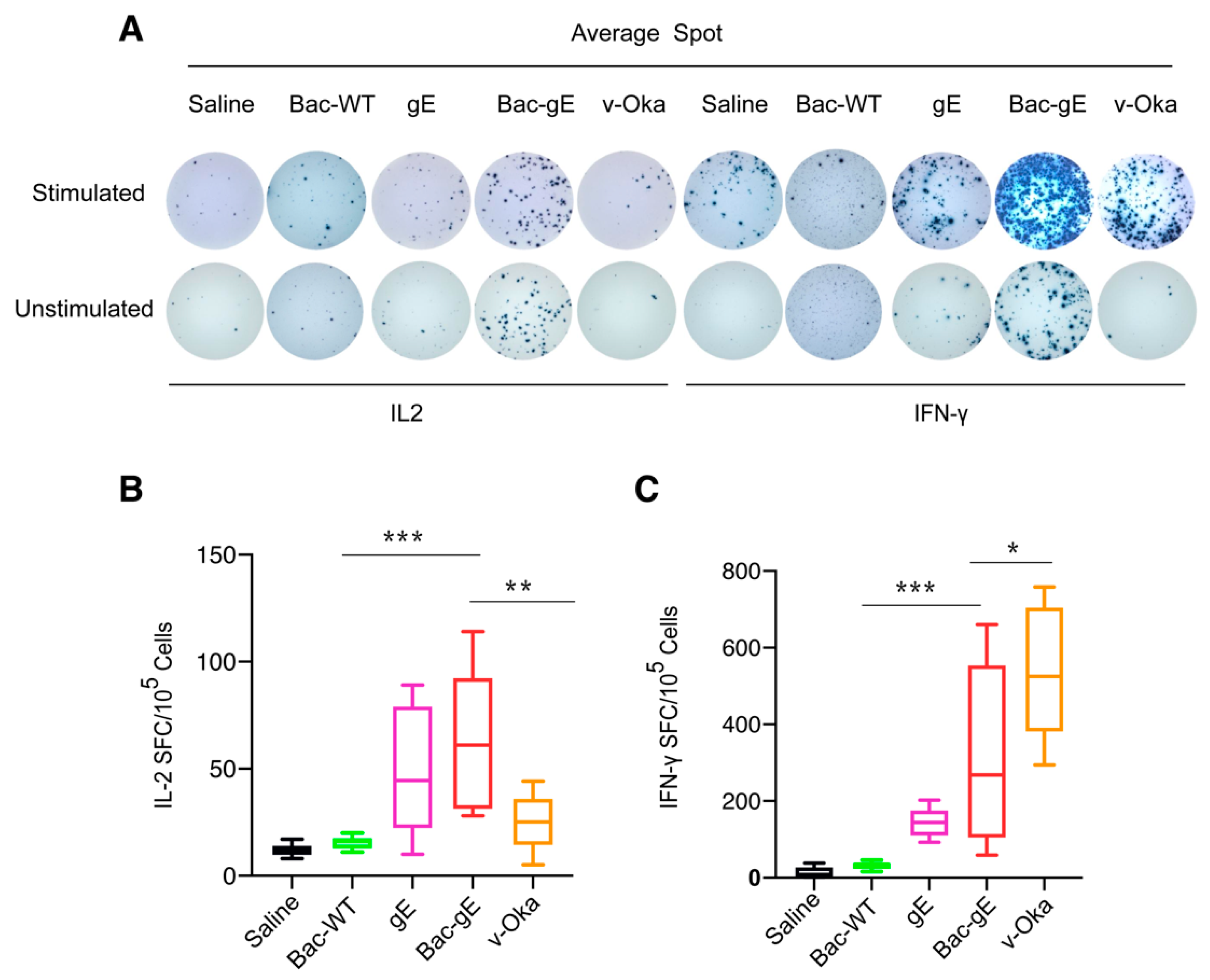
3.3. Bac–gE Induces a Strong Cellular Immune Response in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kennedy, P.G.E.; Gershon, A.A. Clinical Features of Varicella–zoster Virus Infection. Viruses 2018, 10, 609. [Google Scholar] [CrossRef] [PubMed]
- Kramer, P.R.; Stinson, C.; Umorin, M.; Deng, M.; Rao, M.; Bellinger, L.L.; Yee, M.B.; Kinchington, P.R. Lateral thalamic control of nociceptive response after whisker pad injection of varicella zoster virus. Neuroscience 2017, 356, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.; Lin, Y.C.; Lao, H.C.; Chen, C.C. Interventional Treatments for Postherpetic Neuralgia: A Systematic Review. Pain Physician 2019, 22, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Batram, M.; Witte, J.; Schwarz, M.; Hain, J.; Ultsch, B.; Steinmann, M.; Bhavsar, A.; Wutzler, P.; Criée, C.-P.; Hermann, C.; et al. Burden of Herpes Zoster in Adult Patients with Underlying Conditions: Analysis of German Claims Data, 2007-2018. Dermatol. Ther. 2021, 11, 1009–1026. [Google Scholar] [CrossRef]
- Insinga, R.P.; Itzler, R.F.; Pellissier, J.M.; Saddier, P.; Nikas, A.A. The incidence of herpes zoster in a United States administrative database. J. Gen. Intern. Med. 2005, 20, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Suo, L.; Li, J.; Pang, X. A retrospective survey on herpes zoster disease burden and characteristics in Beijing, China. Hum. Vaccines Immunother. 2018, 14, 2632–2635. [Google Scholar] [CrossRef]
- Sato, T.; Yamamoto, W.; Tanaka, A.; Shimazaki, H.; Sugita, S.; Kaburaki, T.; Takeuchi, M. Viral Loads in Ocular Fluids of Acute Retinal Necrosis Eyes Infected by Varicella–zoster Virus Treated with Intravenous Acyclovir Treatment. J. Clin. Med. 2020, 9, 1204. [Google Scholar] [CrossRef]
- Inaba, I.; Kondo, Y.; Iwasaki, S.; Tsuruhashi, S.; Akaishi, A.; Morita, K.; Oniki, K.; Saruwatari, J.; Ishitsuka, Y.; Irie, T. Risk Evaluation for Acute Kidney Injury Induced by the Concomitant Use of Valacyclovir, Analgesics, and Renin–Angiotensin System Inhibitors: The Detection of Signals of Drug–Drug Interactions. Front. Pharmacol. 2019, 10, 874. [Google Scholar] [CrossRef]
- Gershon, A.A.; Gershon, M.D.; Breuer, J.; Levin, M.J.; Oaklander, A.L.; Griffiths, P.D. Advances in the understanding of the pathogenesis and epidemiology of herpes zoster. J. Clin. Virol. 2010, 48 (Suppl. S1), S2–S7. [Google Scholar] [CrossRef]
- Crooke, S.N.; Ovsyannikova, I.G.; Poland, G.A.; Kennedy, R.B. Immunosenescence and human vaccine immune responses. Immun. Ageing 2019, 16, 25. [Google Scholar] [CrossRef]
- Harpaz, R.; Ortega-Sanchez, I.R.; Seward, J.F. Prevention of herpes zoster: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2008, 57, 1–16. [Google Scholar] [PubMed]
- Takahashi, M.; Otsuka, T.; Okuno, Y.; Asano, Y.; Yazaki, T.; Isomura, S. Live vaccine used to prevent the spread of varicella in children in hospital. Lancet 1974, 304, 1288–1290. [Google Scholar] [CrossRef]
- Dagnew, A.F.; Ilhan, O.; Lee, W.-S.; Woszczyk, D.; Kwak, J.-Y.; Bowcock, S.; Sohn, S.K.; Rodriguez Macías, G.; Chiou, T.-J.; Quiel, D.; et al. Immunogenicity and safety of the adjuvanted recombinant zoster vaccine in adults with haematological malignancies: A phase 3, randomised, clinical trial and post-hoc efficacy analysis. Lancet Infect. Dis. 2019, 19, 988–1000. [Google Scholar] [CrossRef]
- James, S.F.; Chahine, E.B.; Sucher, A.J.; Hanna, C. Shingrix: The New Adjuvanted Recombinant Herpes Zoster Vaccine. Ann. Pharmacother. 2018, 52, 673–680. [Google Scholar] [CrossRef]
- Vink, P.; Shiramoto, M.; Ogawa, M.; Eda, M.; Douha, M.; Heineman, T.; Lal, H. Safety and immunogenicity of a Herpes Zoster subunit vaccine in Japanese population aged ≥50 years when administered subcutaneously vs. intramuscularly. Hum. Vaccines Immunother. 2017, 13, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Zarin, W.; Cardoso, R.; Veroniki, A.-A.; Khan, P.A.; Nincic, V.; Ghassemi, M.; Warren, R.; Sharpe, J.P.; Page, A.V. Efficacy, effectiveness, and safety of herpes zoster vaccines in adults aged 50 and older: Systematic review and network meta-analysis. bmj 2018, 363, k4029. [Google Scholar] [CrossRef] [PubMed]
- Lacaille-Dubois, M.A. Updated insights into the mechanism of action and clinical profile of the immunoadjuvant QS-21: A review. Phytomedicine 2019, 60, 152905. [Google Scholar] [CrossRef]
- Kost, T.A.; Kemp, C.W. Fundamentals of Baculovirus Expression and Applications. In Advanced Technologies for Protein Complex Production and Characterization; Vega, M.C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 187–197. [Google Scholar]
- Felberbaum, R.S. The baculovirus expression vector system: A commercial manufacturing platform for viral vaccines and gene therapy vectors. Biotechnol. J. 2015, 10, 702–714. [Google Scholar] [CrossRef]
- Wang, R.; Deng, F.; Hou, D.; Zhao, Y.; Guo, L.; Wang, H.; Hu, Z. Proteomics of the Autographa californica nucleopolyhedrovirus budded virions. J. Virol. 2010, 84, 7233–7242. [Google Scholar] [CrossRef]
- Boldogkői, Z.; Moldován, N.; Szűcs, A.; Tombácz, D. Transcriptome-wide analysis of a baculovirus using nanopore sequencing. Sci. Data 2018, 5, 180276. [Google Scholar] [CrossRef]
- Luo, D.; Miao, Y.; Ke, X.; Tan, Z.; Hu, C.; Li, P.; Wang, T.; Zhang, Y.; Sun, J.; Liu, Y.; et al. Baculovirus Surface Display of Zika Virus Envelope Protein Protects against Virus Challenge in Mouse Model. Virol. Sin. 2020, 35, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Tung, M.-C.; Lu, H.-Y.; Chang, Y.-K.; Huang, W.-R.; Liao, T.-L.; Wu, H.-Y.; Chang, C.-D.; Fan, H.-C.; Nielsen, B.L.; Liu, H.-J. Baculovirus surface display of the HA protein of H5N2 avian influenza virus and its immunogenicity against a lethal challenge with H5N1 virus in chickens. Vet. Microbiol. 2020, 243, 108640. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Yang, W.-C.; Chang, Y.-K.; Wang, C.-Y.; Huang, W.-R.; Li, J.-Y.; Chuang, K.-P.; Wu, H.-Y.; Chang, C.-D.; Nielsen, B.L.; et al. Construction of polycistronic baculovirus surface display vectors to express the PCV2 Cap(d41) protein and analysis of its immunogenicity in mice and swine. Vet. Res. 2020, 51, 112. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Liu, Y.; Qu, X.; Deng, H.; Ding, M.; Lau, T.L.T.; Yu, A.C.-H.; Chen, J. Baculovirus Surface Display of SARS Coronavirus (SARS-CoV) Spike Protein and Immunogenicity of the Displayed Protein in Mice Models. DNA Cell Biol. 2006, 25, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Ono, C.; Ninomiya, A.; Yamamoto, S.; Abe, T.; Wen, X.; Fukuhara, T.; Sasai, M.; Yamamoto, M.; Saitoh, T.; Satoh, T. Innate immune response induced by baculovirus attenuates transgene expression in mammalian cells. J. Virol. 2014, 88, 2157–2167. [Google Scholar] [CrossRef]
- Yeh, D.-W.; Lai, C.-Y.; Liu, Y.-L.; Lu, C.-H.; Tseng, P.-H.; Yuh, C.-H.; Yu, G.-Y.; Liu, S.-J.; Leng, C.-H.; Chuang, T.-H. CpG-oligodeoxynucleotides developed for grouper toll-like receptor (TLR) 21s effectively activate mouse and human TLR9s mediated immune responses. Sci. Rep. 2017, 7, 17297. [Google Scholar] [CrossRef]
- Premanand, B.; Zhong Wee, P.; Prabakaran, M. Baculovirus Surface Display of Immunogenic Proteins for Vaccine Development. Viruses 2018, 10, 298. [Google Scholar] [CrossRef]
- Pan, Y.; Zhao, Q.; Fang, L.; Luo, R.; Chen, H.; Xiao, S. Efficient gene delivery into mammalian cells by recombinant baculovirus containing a hybrid cytomegalovirus promoter/Semliki Forest virus replicon. J. Gene Med. 2009, 11, 1030–1038. [Google Scholar] [CrossRef]
- Liu, J.; Ye, X.; Jia, J.; Zhu, R.; Wang, L.; Chen, C.; Yang, L.; Wang, Y.; Wang, W.; Ye, J.; et al. Serological Evaluation of Immunity to the Varicella–zoster Virus Based on a Novel Competitive Enzyme-Linked Immunosorbent Assay. Sci. Rep. 2016, 6, 20577. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Z.; Zhang, Z.; Qiao, J.; Rong, R.; Zhang, Y.; Yao, Q.; Li, Z.; Shen, H.; Huang, F.; et al. Characterization of native-like HIV-1 gp140 glycoprotein expressed in insect cells. Vaccine 2019, 37, 1418–1427. [Google Scholar] [CrossRef]
- Guo, G.; Gao, M.; Gao, X.; Zhu, B.; Huang, J.; Tu, X.; Kim, W.; Zhao, F.; Zhou, Q.; Zhu, S.; et al. Reciprocal regulation of RIG-I and XRCC4 connects DNA repair with RIG-I immune signaling. Nat. Commun. 2021, 12, 2187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qiao, L.; Hu, X.; Zhao, K.; Zhang, Y.; Chai, F.; Pan, Z. Baculovirus vectors expressing F proteins in combination with virus-induced signaling adaptor (VISA) molecules confer protection against respiratory syncytial virus infection. Vaccine 2016, 34, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Maximova, O.A.; Liu, G.; Kenney, H.; Teterina, N.; Bloom, M.E.; Grabowski, J.M.; Mlera, L.; Nagata, B.M.; Moore, I.; et al. Routes of Zika virus dissemination in the testis and epididymis of immunodeficient mice. Nat. Commun. 2018, 9, 5350. [Google Scholar] [CrossRef] [PubMed]
- Kisuya, J.; Chemtai, A.; Raballah, E.; Keter, A.; Ouma, C. The diagnostic accuracy of Th1 (IFN-γ, TNF-α, and IL-2) and Th2 (IL-4, IL-6 and IL-10) cytokines response in AFB microscopy smear negative PTB-HIV co-infected patients. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Arvin, A.M.; Wolinsky, J.S.; Kappos, L.; Morris, M.I.; Reder, A.T.; Tornatore, C.; Gershon, A.; Gershon, M.; Levin, M.J.; Bezuidenhoudt, M.; et al. Varicella–zoster virus infections in patients treated with fingolimod: Risk assessment and consensus recommendations for management. JAMA Neurol. 2015, 72, 31–39. [Google Scholar] [CrossRef]
- Sauboin, C.; Holl, K.; Bonanni, P.; Gershon, A.A.; Benninghoff, B.; Carryn, S.; Burgess, M.A.; Wutzler, P. The impact of childhood varicella vaccination on the incidence of herpes zoster in the general population: Modelling the effect of exogenous and endogenous Varicella–zoster virus immunity boosting. BMC Infect. Dis. 2019, 19, 126. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Pasquale, A.D.; Preiss, S.; Silva, F.T.; Garçon, N. Vaccine Adjuvants: From 1920 to 2015 and Beyond. Vaccines 2015, 3, 320. [Google Scholar] [CrossRef]
- Alving, C.R.; Beck, Z.; Matyas, G.R.; Rao, M. Liposomal adjuvants for human vaccines. Expert Opin. Drug Deliv. 2016, 13, 807–816. [Google Scholar] [CrossRef]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like receptors activation, signaling, and targeting: An overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef]
- Mu, L.M.; Liu, L.; Liu, R.; Du, Y.F.; Luo, Q.; Xu, J.R.; Xie, Y.; Lu, W.L. Nanostructured SL9-CpG Lipovaccines Elicit Immune Response for the Treatment of Melanoma. Int. J. Mol. Sci. 2019, 20, 2207. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Hemmi, H.; Miyamoto, H.; Moriishi, K.; Tamura, S.; Takaku, H.; Akira, S.; Matsuura, Y. Involvement of the Toll-like receptor 9 signaling pathway in the induction of innate immunity by baculovirus. J. Virol. 2005, 79, 2847–2858. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Du, M.; Liu, X.; Zhang, Z.; Zhang, Z.; Meng, X.; Li, Y. Interaction with the Receptor SLAM and Baculovirus Surface Display of Peste des petits ruminants Virus Hemagglutinin. DNA Cell Biol. 2020, 39, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-Y.; Hsu, W.-T.; Chao, Y.-C.; Chang, H.-W. Display of porcine epidemic diarrhea virus spike protein on baculovirus to improve immunogenicity and protective efficacy. Viruses 2018, 10, 346. [Google Scholar] [CrossRef]
- Pidre, M.L.; Ferrelli, M.L.; Haase, S.; Romanowski, V. Baculovirus Display: A Novel Tool for Vaccination. In Current Issues in Molecular Virology-Viral Genetics and Biotechnological Applications; InTech Open: London, UK, 2013; pp. 137–164. [Google Scholar]
- Hosseini, A.; Hashemi, V.; Shomali, N.; Asghari, F.; Gharibi, T.; Akbari, M.; Gholizadeh, S.; Jafari, A. Innate and adaptive immune responses against coronavirus. Biomed. Pharmacother. 2020, 132, 110859. [Google Scholar] [CrossRef]
- Dai, H.; Lan, P.; Zhao, D.; Abou-Daya, K.; Liu, W.; Chen, W.; Friday, A.J.; Williams, A.L.; Sun, T.; Chen, J. PIRs mediate innate myeloid cell memory to nonself MHC molecules. Science 2020, 368, 1122–1127. [Google Scholar] [CrossRef]
- Cao, Y.; Zhu, X.; Hossen, M.N.; Kakar, P.; Zhao, Y.; Chen, X. Augmentation of vaccine-induced humoral and cellular immunity by a physical radiofrequency adjuvant. Nat. Commun. 2018, 9, 3695. [Google Scholar] [CrossRef]
- Hayward, A.C.; Wang, L.; Goonetilleke, N.; Fragaszy, E.B.; Bermingham, A.; Copas, A.; Dukes, O.; Millett, E.R.C.; Nazareth, I.; Nguyen-Van-Tam, J.S.; et al. Natural T Cell–mediated Protection against Seasonal and Pandemic Influenza. Results of the Flu Watch Cohort Study. Am. J. Respir. Crit. Care Med. 2015, 191, 1422–1431. [Google Scholar] [CrossRef]
- Hong, X.; Zhong, X.; Du, G.; Hou, Y.; Zhang, Y.; Zhang, Z.; Gong, T.; Zhang, L.; Sun, X. The pore size of mesoporous silica nanoparticles regulates their antigen delivery efficiency. Sci. Adv. 2020, 6, eaaz4462. [Google Scholar] [CrossRef]
- Hu, Y.C. Baculovirus Vectors for Gene Therapy. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2006; Volume 68, pp. 287–320. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine | Dose (PFU) | Dose (ug) | Bleeding (Week) | Timepoint Spleen Removal (Week) |
|---|---|---|---|---|
| Bac–gE +Al | 1 × 108 | 0.3 | 1–7 | 5 |
| Bac–gE | 1 × 108 | 0.3 | 1–7 | 5 |
| Bac–WT | 1 × 108 | - | 1–7 | 5 |
| gE+Al | - | 0.3 | 1–7 | 5 |
| Voka | 5 × 106 | 0.3 | 1–7 | 5 |
| PBS | - | - | 1–7 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, W.; Li, T.; Zhang, S.; Wang, Y.; Hong, M.; Cui, L.; Wang, H.; Zhang, Y.; Chen, T.; Zhu, R.; et al. Baculovirus Display of Varicella–Zoster Virus Glycoprotein E Induces Robust Humoral and Cellular Immune Responses in Mice. Viruses 2022, 14, 1785. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081785
Xue W, Li T, Zhang S, Wang Y, Hong M, Cui L, Wang H, Zhang Y, Chen T, Zhu R, et al. Baculovirus Display of Varicella–Zoster Virus Glycoprotein E Induces Robust Humoral and Cellular Immune Responses in Mice. Viruses. 2022; 14(8):1785. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081785
Chicago/Turabian StyleXue, Wenhui, Tingting Li, Sibo Zhang, Yingbin Wang, Minqing Hong, Lingyan Cui, Hong Wang, Yuyun Zhang, Tingting Chen, Rui Zhu, and et al. 2022. "Baculovirus Display of Varicella–Zoster Virus Glycoprotein E Induces Robust Humoral and Cellular Immune Responses in Mice" Viruses 14, no. 8: 1785. https://0-doi-org.brum.beds.ac.uk/10.3390/v14081785