

A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protect Pigs against a Virulent CSFV Challenge
 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Medium, Virus, and Titration
2.2. Viruses
2.3. Animals
2.4. Vaccination and Challenge
2.4.1. Clinical Examination and Sample Collection from Pigs Following Vaccination and Challenge
2.4.2. Total and Differential White Blood Cell Counts
2.5. Detection of CSFV Viremia

2.6. Detection of Neutralizing Antibodies
2.7. Statistical Analysis
3. Results
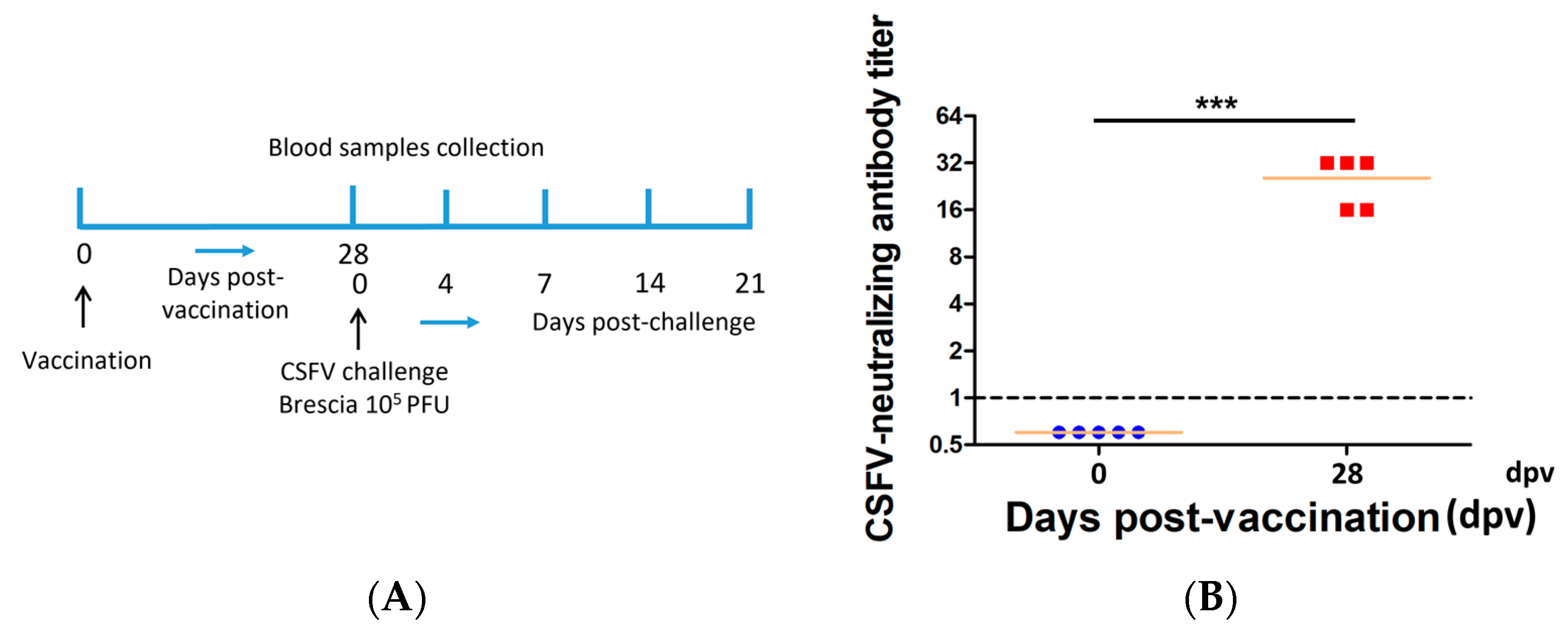
3.1. PRVtmv+ Vaccinated Pigs Remained Clinically Normal and Generated Moderate Levels of CSFV-Specific Neutralizing Antibody Titers
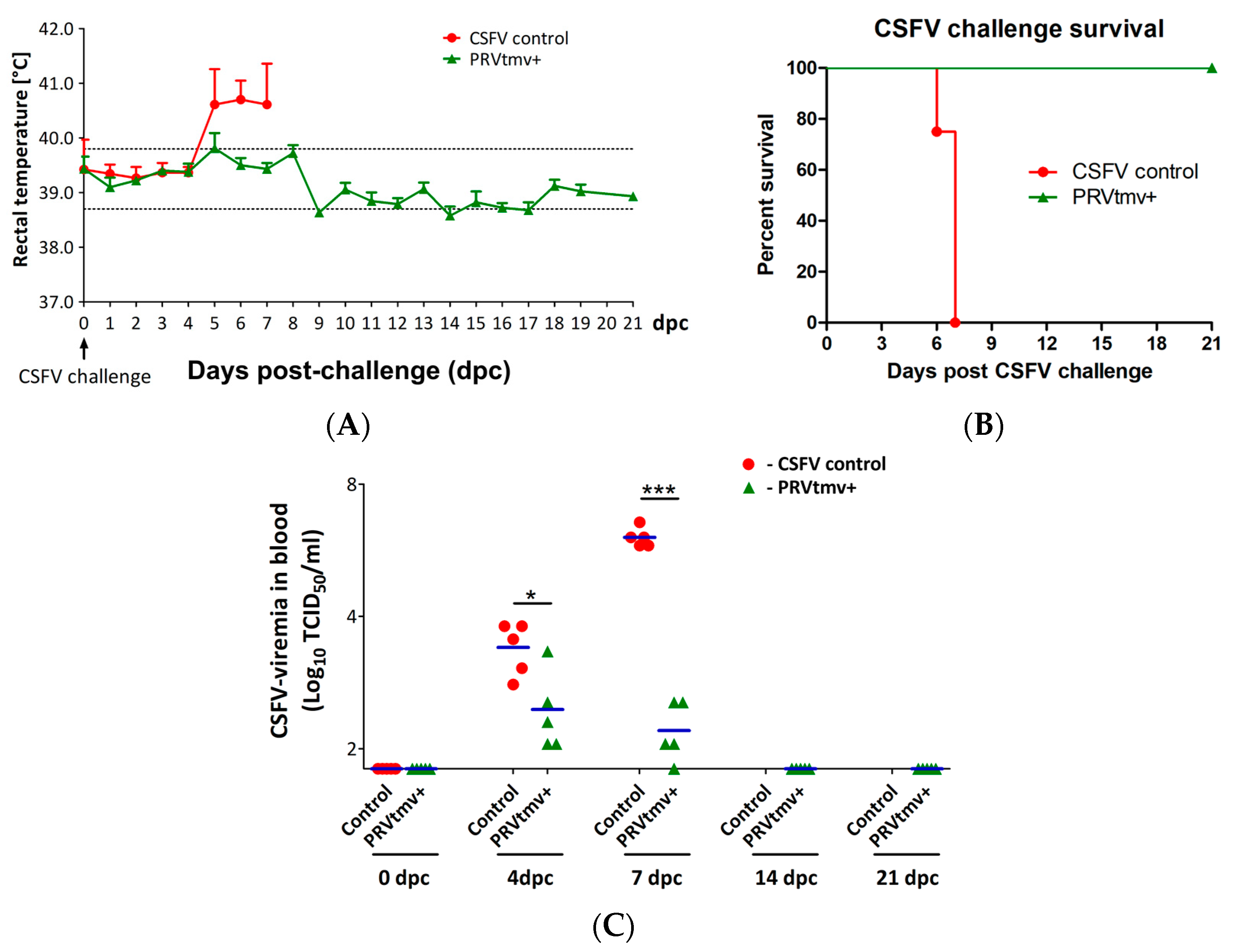
3.2. PRVtmv+ Vaccinated Pigs Were Protected against Severe CSFV Clinical Disease
3.3. PRVtmv+ Vaccinated Pigs Had a Three-Fold Lower CSFV Viremia in Blood
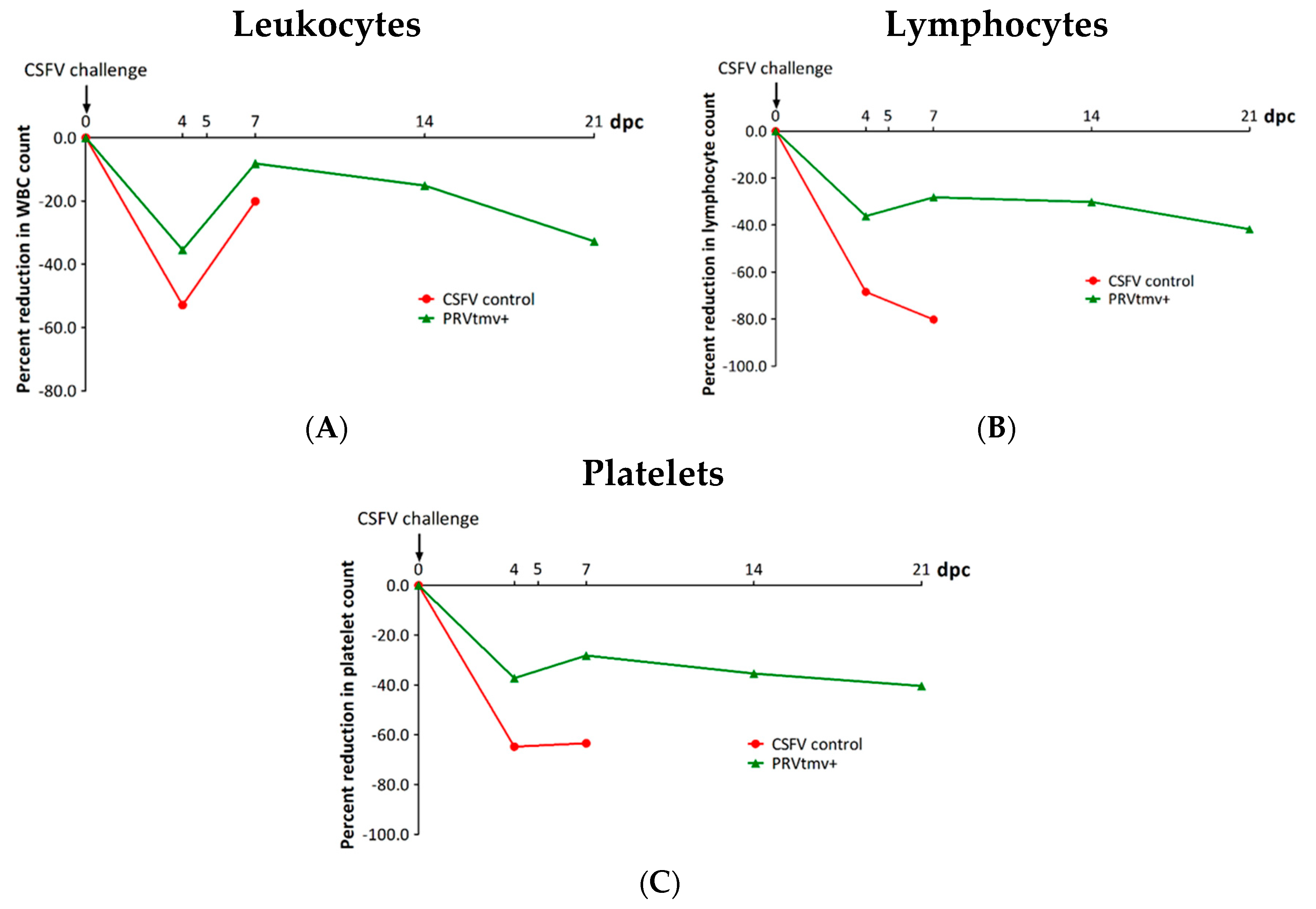
3.4. PRVtmv+ Vaccinated Pigs Had Significantly Milder Leukopenia and Thrombocytopenia after the CSFV Challenge
3.5. PRVtmv+ Vaccinated Pigs Did Not Have Any CSFV-Specific Gross Lesions upon CSFV Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical Swine Fever-An Updated Review. Viruses 2017, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Postel, A.; Austermann-Busch, S.; Petrov, A.; Moennig, V.; Becher, P. Epidemiology, diagnosis and control of classical swine fever: Recent developments and future challenges. Transbound. Emerg. Dis. 2018, 65 (Suppl. S1), 248–261. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, M.P.; Horst, S.H.; Huirne, R.B.; Dijkhuizen, A.A. A model to estimate the financial consequences of classical swine fever outbreaks: Principles and outcomes. Prev. Vet. Med. 1999, 42, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Stegeman, A.; Elbers, A.; de Smit, H.; Moser, H.; Smak, J.; Pluimers, F. The 1997-1998 epidemic of classical swine fever in the Netherlands. Vet. Microbiol. 2000, 73, 183–196. [Google Scholar] [CrossRef]
- KRAMER-SCHADT, S.; FERNÁNDEZ, N.; THULKE, H.-H. Potential ecological and epidemiological factors affecting the persistence of classical swine fever in wild boar Sus scrofa populations. Mammal. Rev. 2007, 37, 1–20. [Google Scholar] [CrossRef]
- Ganges, L.; Crooke, H.R.; Bohorquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef]
- Brown, V.R.; Bevins, S.N. A Review of Classical Swine Fever Virus and Routes of Introduction into the United States and the Potential for Virus Establishment. Front. Vet. Sci. 2018, 5, 31. [Google Scholar] [CrossRef]
- Coronado, L.; Perera, C.L.; Rios, L.; Frías, M.T.; Pérez, L.J. A Critical Review about Different Vaccines against Classical Swine Fever Virus and Their Repercussions in Endemic Regions. Vaccines 2021, 9, 154. [Google Scholar] [CrossRef]
- Kaden, V.; Lange, E.; Kuster, H.; Muller, T.; Lange, B. An update on safety studies on the attenuated "RIEMSER Schweinepestoralvakzine" for vaccination of wild boar against classical swine fever. Vet. Microbiol. 2010, 143, 133–138. [Google Scholar] [CrossRef]
- Rossi, S.; Staubach, C.; Blome, S.; Guberti, V.; Thulke, H.H.; Vos, A.; Koenen, F.; Le Potier, M.F. Controlling of CSFV in European wild boar using oral vaccination: A review. Front. Microbiol. 2015, 6, 1141. [Google Scholar] [CrossRef]
- de Smit, A.J.; van Gennip, H.G.; Miedema, G.K.; van Rijn, P.A.; Terpstra, C.; Moormann, R.J. Recombinant classical swine fever (CSF) viruses derived from the Chinese vaccine strain (C-strain) of CSF virus retain their avirulent and immunogenic characteristics. Vaccine 2000, 18, 2351–2358. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Tautz, N.; Becher, P.; Thiel, H.J.; Kummerer, B.M. Recovery of cytopathogenic and noncytopathogenic bovine viral diarrhea viruses from cDNA constructs. J. Virol. 1997, 71, 1735. [Google Scholar] [CrossRef] [PubMed]
- Reimann, I.; Depner, K.; Trapp, S.; Beer, M. An avirulent chimeric Pestivirus with altered cell tropism protects pigs against lethal infection with classical swine fever virus. Virology 2004, 322, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Konig, P.; Blome, S.; Gabriel, C.; Reimann, I.; Beer, M. Innocuousness and safety of classical swine fever marker vaccine candidate CP7_E2alf in non-target and target species. Vaccine 2011, 30, 5–8. [Google Scholar] [CrossRef]
- MacDiarmid, S.C. Aujeszky’s disease eradication in New Zealand. Aust. Vet. J. 2000, 78, 470–471. [Google Scholar] [CrossRef]
- Muller, T.; Batza, H.J.; Schluter, H.; Conraths, F.J.; Mettenleiter, T.C. Eradication of Aujeszky’s disease in Germany. J. Vet. Med. B Infect. Dis. Vet. Public. Health 2003, 50, 207–213. [Google Scholar] [CrossRef]
- OIE. OIE World Animal Health Information System. Available online: https://wahis.woah.org/#/dashboards/country-or-disease-dashboard (accessed on 12 October 2023).
- Tan, L.; Yao, J.; Yang, Y.; Luo, W.; Yuan, X.; Yang, L.; Wang, A. Current Status and Challenge of Pseudorabies Virus Infection in China. Virol. Sin. 2021, 36, 588–607. [Google Scholar] [CrossRef]
- Liu, A.; Xue, T.; Zhao, X.; Zou, J.; Pu, H.; Hu, X.; Tian, Z. Pseudorabies Virus Associations in Wild Animals: Review of Potential Reservoirs for Cross-Host Transmission. Viruses 2022, 14, 2254. [Google Scholar] [CrossRef]
- Pacini, M.I.; Forzan, M.; Cilia, G.; Bernardini, L.; Marzoli, F.; Pedonese, F.; Bandecchi, P.; Fratini, F.; Mazzei, M. Detection of Pseudorabies Virus in Wild Boar Foetus. Animals 2020, 10, 366. [Google Scholar] [CrossRef]
- Pedersen, K.; Turnage, C.T.; Gaston, W.D.; Arruda, P.; Alls, S.A.; Gidlewski, T. Pseudorabies detected in hunting dogs in Alabama and Arkansas after close contact with feral swine (Sus scrofa). BMC Vet. Res. 2018, 14, 388. [Google Scholar] [CrossRef]
- Zimmerman, J.J. Diseases of Swine, 10th ed.; Wiley-Blackwell: Ames, IA, USA, 2012. [Google Scholar]
- Zhou, H.; Pan, Y.; Liu, M.; Han, Z. Prevalence of Porcine Pseudorabies Virus and Its Coinfection Rate in Heilongjiang Province in China from 2013 to 2018. Viral Immunol. 2020, 33, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Bai, C.; Sun, J.; Chang, S.; Zhang, X. Emergence of virulent pseudorabies virus infection in northern China. J. Vet. Sci. 2013, 14, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Bo, Z.; Miao, Y.; Xi, R.; Gao, X.; Miao, D.; Chen, H.; Jung, Y.S.; Qian, Y.; Dai, J. Emergence of a novel pathogenic recombinant virus from Bartha vaccine and variant pseudorabies virus in China. Transbound. Emerg. Dis. 2021, 68, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Delva, J.L.; Nauwynck, H.J.; Mettenleiter, T.C.; Favoreel, H.W. The Attenuated Pseudorabies Virus Vaccine Strain Bartha K61: A Brief Review on the Knowledge Gathered During 60 Years of Research. Pathogens 2020, 9, 897. [Google Scholar] [CrossRef] [PubMed]
- Pavulraj, S.; Pannhorst, K.; Stout, R.W.; Paulsen, D.B.; Carossino, M.; Meyer, D.; Becher, P.; Chowdhury, S.I. A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protects Pigs against PCV2b Challenge and Induces Serum Neutralizing Antibody Response against CSFV. Vaccines 2022, 10, 305. [Google Scholar] [CrossRef]
- Risatti, G.R.; Holinka, L.G.; Lu, Z.; Kutish, G.F.; Tulman, E.R.; French, R.A.; Sur, J.H.; Rock, D.L.; Borca, M.V. Mutation of E1 glycoprotein of classical swine fever virus affects viral virulence in swine. Virology 2005, 343, 116–127. [Google Scholar] [CrossRef]
- Risatti, G.R.; Borca, M.V.; Kutish, G.F.; Lu, Z.; Holinka, L.G.; French, R.A.; Tulman, E.R.; Rock, D.L. The E2 glycoprotein of classical swine fever virus is a virulence determinant in swine. J. Virol. 2005, 79, 3787–3796. [Google Scholar] [CrossRef]
- Holinka, L.G.; Fernandez-Sainz, I.; O’Donnell, V.; Prarat, M.V.; Gladue, D.P.; Lu, Z.; Risatti, G.R.; Borca, M.V. Development of a live attenuated antigenic marker classical swine fever vaccine. Virology 2009, 384, 106–113. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Edwards, S.; Moennig, V.; Wensvoort, G. The development of an international reference panel of monoclonal antibodies for the differentiation of hog cholera virus from other pestiviruses. Vet. Microbiol. 1991, 29, 101–108. [Google Scholar] [CrossRef]
- OIE. Classical Swine Fever (Infection with Classical Swine Fever Virus). Available online: https://www.woah.org/fileadmin/Home/fr/Health_standards/tahm/3.09.03_CSF.pdf (accessed on 24 March 2023).
- Koomkrong, N.; Boonkaewwan, C.; Laenoi, W.; Kayan, A. Blood haematology, muscle pH and serum cortisol changes in pigs with different levels of drip loss. Asian-Australas. J. Anim. Sci. 2017, 30, 1751–1755. [Google Scholar] [CrossRef] [PubMed]
- Pavulraj, S.; Stout, R.W.; Paulsen, D.B.; Chowdhury, S.I. Live Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Undergoes an Abortive Replication Cycle in the TG Neurons following Latency Reactivation. Viruses 2023, 15, 473. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.; Fukusho, A.; Lefevre, P.C.; Lipowski, A.; Pejsak, Z.; Roehe, P.; Westergaard, J. Classical swine fever: The global situation. Vet. Microbiol. 2000, 73, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Fritzemeier, J.; Teuffert, J.; Greiser-Wilke, I.; Staubach, C.; Schluter, H.; Moennig, V. Epidemiology of classical swine fever in Germany in the 1990s. Vet Microbiol 2000, 77, 29–41. [Google Scholar] [CrossRef]
- Leifer, I.; Hoffmann, B.; Hoper, D.; Bruun Rasmussen, T.; Blome, S.; Strebelow, G.; Horeth-Bontgen, D.; Staubach, C.; Beer, M. Molecular epidemiology of current classical swine fever virus isolates of wild boar in Germany. J. Gen. Virol. 2010, 91, 2687–2697. [Google Scholar] [CrossRef]
- de Oliveira, L.G.; Gatto, I.R.H.; Mechler-Dreibi, M.L.; Almeida, H.M.S.; Sonalio, K.; Storino, G.Y. Achievements and Challenges of Classical Swine Fever Eradication in Brazil. Viruses 2020, 12, 1327. [Google Scholar] [CrossRef]
- Graham, S.P.; Everett, H.E.; Haines, F.J.; Johns, H.L.; Sosan, O.A.; Salguero, F.J.; Clifford, D.J.; Steinbach, F.; Drew, T.W.; Crooke, H.R. Challenge of Pigs with Classical Swine Fever Viruses after C-Strain Vaccination Reveals Remarkably Rapid Protection and Insights into Early Immunity. PLoS ONE 2012, 7, e29310. [Google Scholar] [CrossRef]
- Schang, L.M.; Kutish, G.F.; Osorio, F.A. Correlation between precolonization of trigeminal ganglia by attenuated strains of pseudorabies virus and resistance to wild-type virus latency. J. Virol. 1994, 68, 8470–8476. [Google Scholar] [CrossRef]
- Nelson, S.A.; Sant, A.J. Potentiating Lung Mucosal Immunity Through Intranasal Vaccination. Front. Immunol. 2021, 12, 808527. [Google Scholar] [CrossRef]
- Zhang, F.; Peng, B.; Chang, H.; Zhang, R.; Lu, F.; Wang, F.; Fang, F.; Chen, Z. Intranasal Immunization of Mice to Avoid Interference of Maternal Antibody against H5N1 Infection. PLoS ONE 2016, 11, e0157041. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, E.; Medina-Ramirez, E.; Pavulraj, S.; Gladue, D.P.; Borca, M.; Chowdhury, S.I. A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protect Pigs against a Virulent CSFV Challenge. Viruses 2023, 15, 2143. https://0-doi-org.brum.beds.ac.uk/10.3390/v15112143
Silva E, Medina-Ramirez E, Pavulraj S, Gladue DP, Borca M, Chowdhury SI. A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protect Pigs against a Virulent CSFV Challenge. Viruses. 2023; 15(11):2143. https://0-doi-org.brum.beds.ac.uk/10.3390/v15112143
Chicago/Turabian StyleSilva, Ediane, Elizabeth Medina-Ramirez, Selvaraj Pavulraj, Douglas P. Gladue, Manuel Borca, and Shafiqul I. Chowdhury. 2023. "A Triple Gene-Deleted Pseudorabies Virus-Vectored Subunit PCV2b and CSFV Vaccine Protect Pigs against a Virulent CSFV Challenge" Viruses 15, no. 11: 2143. https://0-doi-org.brum.beds.ac.uk/10.3390/v15112143