
Caboxamycin Inhibits Heart Inflammation in a Coxsackievirus B3-Induced Myocarditis Mouse Model
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedure
2.2. Collection and Phylogenetic Analysis of Strain SC0774
2.3. Fermentation, Extraction, and Isolation
2.4. Viruses and Cells
2.5. Screening and Optimization of Antiviral Compounds
2.6. Myocarditis Mouse Model and Drug Administration
2.7. Quantitative Real-Time PCR
2.8. Immunoblot Analysis
2.9. Histologic Findings
2.10. Statistical Analysis
3. Results
3.1. Compound 1 Characterization
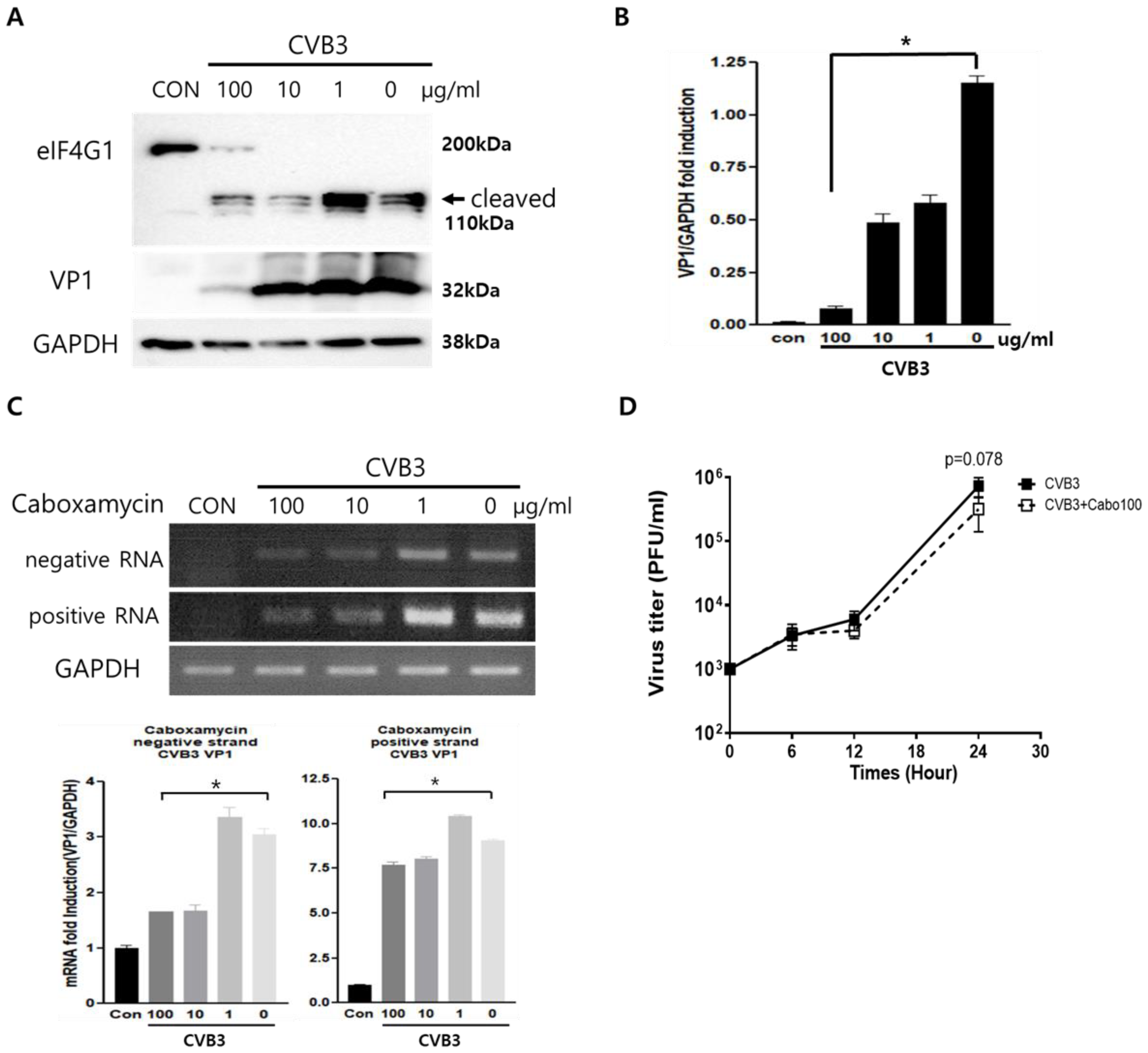
3.2. Caboxamycin Strongly Inhibited Coxsackievirus B3 Replication
3.3. Caboxamycin Treatment Inhibited Apoptosis and Improved the Cell Survival Signaling Pathway
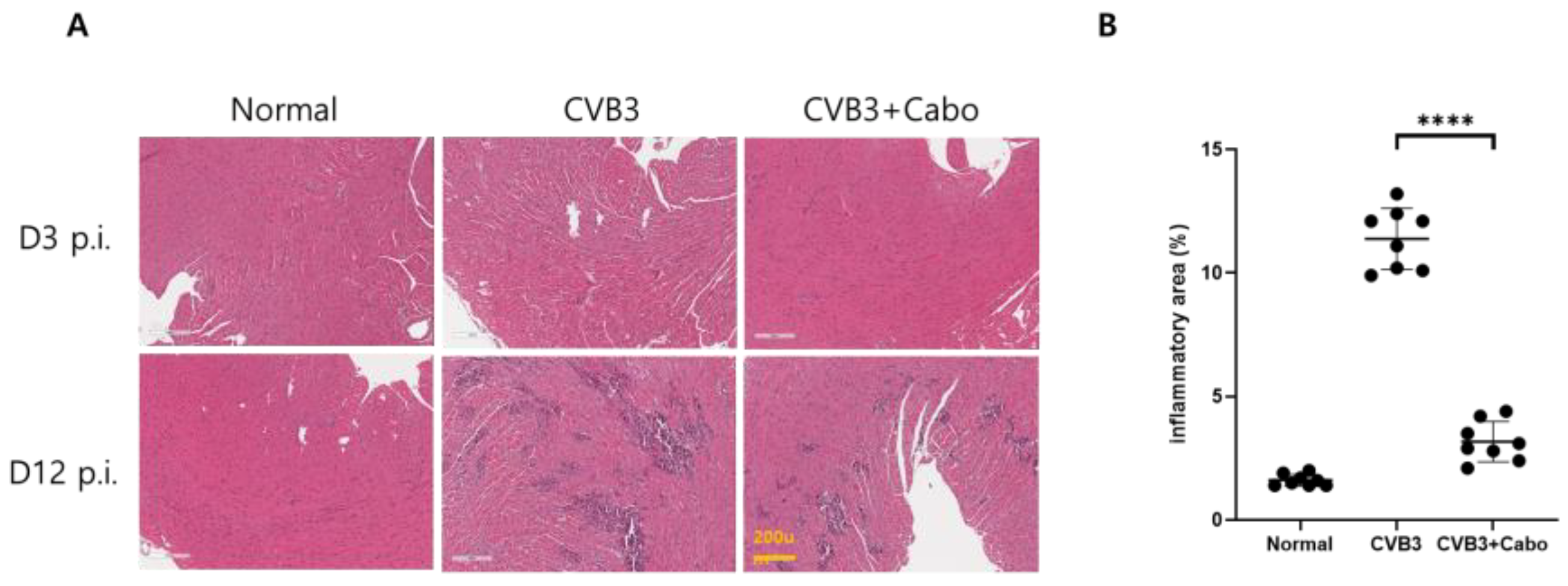
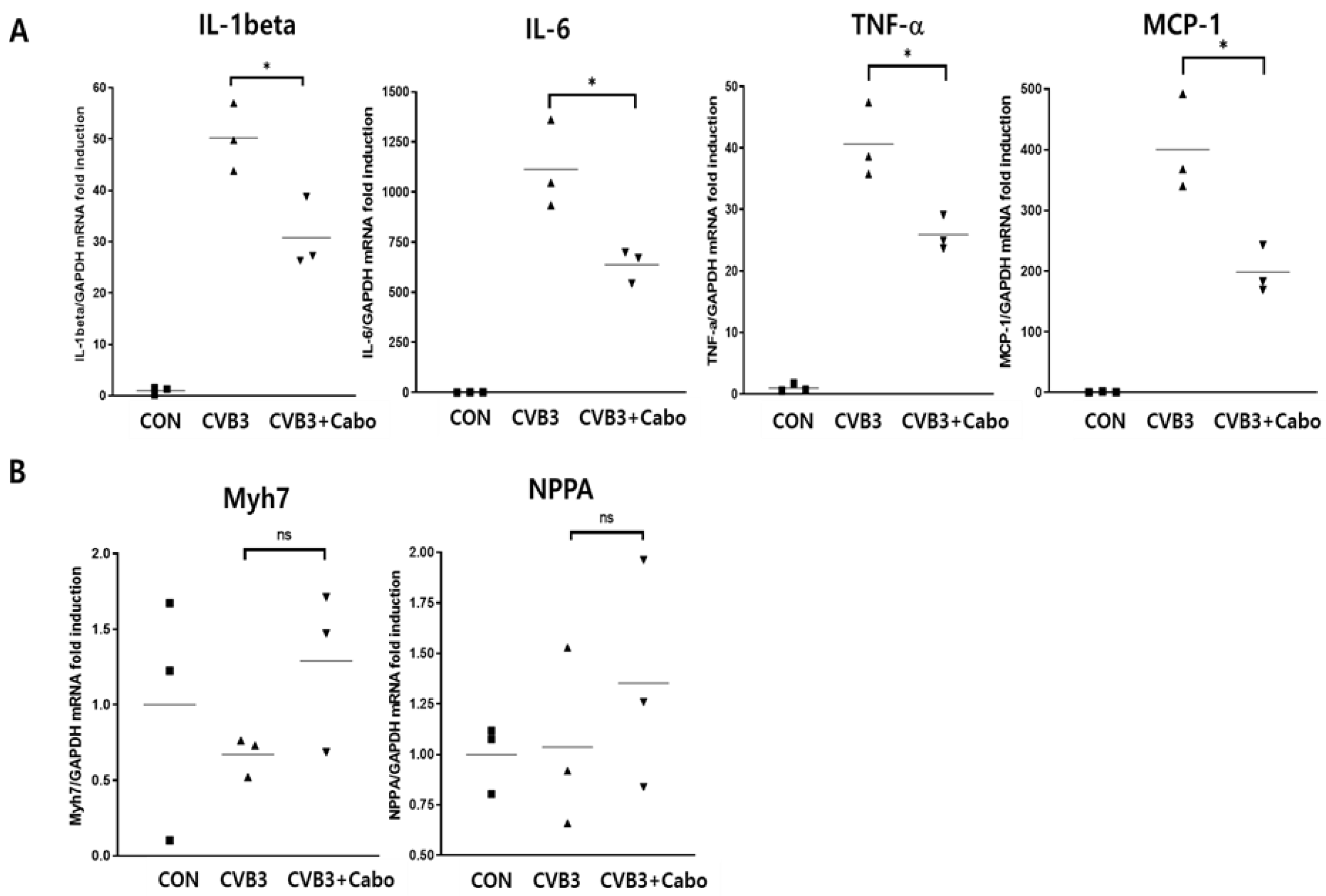
3.4. Caboxamycin Reduced Cardiac Inflammation and Virus Replication in a CVB3-Induced Myocarditis Mouse Model
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huber, S.A.; Lodge, P.A. Coxsackievirus B-3 myocarditis. Identification of different pathogenic mechanisms in DBA/2 and Balb/c mice. Am. J. Pathol. 1986, 122, 284–291. [Google Scholar] [PubMed]
- Feldman, A.M.; McNamara, D. Myocarditis. N. Engl. J. Med. 2000, 343, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, J.F. Viral myocarditis. A review. Am. J. Pathol. 1980, 101, 425–484. [Google Scholar] [PubMed]
- Knowlton, K.U. Myocarditis: An Intersection Between Genetic and Acquired Causes of Human Cardiomyopathy. J. Am. Coll. Cardiol. 2017, 69, 1666–1668. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.M.; Mulkey, S.B.; Campos, J.M.; DeBiasi, R.L. Laboratory diagnosis of CNS infections in children due to emerging and re-emerging neurotropic viruses. Pediatr. Res. 2024, 95, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Abzug, M.J.; Cloud, G.; Bradley, J.; Sanchez, P.J.; Romero, J.; Powell, D.; Lepow, M.; Mani, C.; Capparelli, E.V.; Blount, S.; et al. Double blind placebo-controlled trial of pleconaril in infants with enterovirus meningitis. Pediatr. Infect. Dis. J. 2003, 22, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Setia, A.; Bhatia, J.; Bhattacharya, S. An Overview of Acute Flaccid Myelitis. CNS Neurol. Disord. Drug Targets 2022, 21, 774–794. [Google Scholar] [CrossRef] [PubMed]
- Nayak, G.; Bhuyan, S.K.; Bhuyan, R.; Sahu, A.; Kar, D.; Kuanar, A. Global emergence of Enterovirus 71: A systematic review. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 78. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Qi, J.; Chen, Z.; Xu, X.; Gao, F.; Lin, D.; Qian, W.; Liu, H.; Jiang, H.; Yan, J.; et al. Enterovirus 71 and coxsackievirus A16 3C proteases: Binding to rupintrivir and their substrates and anti-hand, foot, and mouth disease virus drug design. J. Virol. 2011, 85, 10319–10331. [Google Scholar] [CrossRef]
- Zhao, Z.; Cai, T.Z.; Lu, Y.; Liu, W.J.; Cheng, M.L.; Ji, Y.Q. Coxsackievirus B3 induces viral myocarditis by upregulating toll-like receptor 4 expression. Biochemistry 2015, 80, 455–462. [Google Scholar] [CrossRef]
- Herskowitz, A.; Beisel, K.W.; Wolfgram, L.J.; Rose, N.R. Coxsackievirus B3 murine myocarditis: Wide pathologic spectrum in genetically defined inbred strains. Hum. Pathol. 1985, 16, 671–673. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Yajima, T.; Lim, B.K.; Stenbit, A.; Dublin, A.; Dalton, N.D.; Summers-Torres, D.; Molkentin, J.D.; Duplain, H.; Wessely, R.; et al. Inducible cardiac-restricted expression of enteroviral protease 2A is sufficient to induce dilated cardiomyopathy. Circulation 2007, 115, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.K.; Peter, A.K.; Xiong, D.; Narezkina, A.; Yung, A.; Dalton, N.D.; Hwang, K.K.; Yajima, T.; Chen, J.; Knowlton, K.U. Inhibition of Coxsackievirus-associated dystrophin cleavage prevents cardiomyopathy. J. Clin. Investig. 2013, 123, 5146–5151. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.H.; Lee, W.G.; Kim, Y.C.; Ju, E.S.; Lim, B.K.; Choi, J.O.; Kim, D.K.; Jeon, E.S. Antiviral activity of coxsackievirus B3 3C protease inhibitor in experimental murine myocarditis. J. Infect. Dis. 2012, 205, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.T.; Hsu, W.C.; Lin, C.C. Antiviral natural products and herbal medicines. J. Tradit. Complement. Med. 2014, 4, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Opavsky, M.A.; Martino, T.; Rabinovitch, M.; Penninger, J.; Richardson, C.; Petric, M.; Trinidad, C.; Butcher, L.; Chan, J.; Liu, P.P. Enhanced ERK-1/2 activation in mice susceptible to coxsackievirus-induced myocarditis. J. Clin. Investig. 2002, 109, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, T.; Matsui, T.; Aoyama, T.; Leri, A.; Anversa, P.; Li, L.; Ogawa, W.; del Monte, F.; Gwathmey, J.K.; Grazette, L.; et al. PI3K rescues the detrimental effects of chronic Akt activation in the heart during ischemia/reperfusion injury. J. Clin. Investig. 2005, 115, 2128–2138. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, X.; Xiong, L.; Ren, J. Ablation of Akt2 prevents paraquat-induced myocardial mitochondrial injury and contractile dysfunction: Role of Nrf2. Toxicol. Lett. 2017, 269, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Park, J.H.; Jeon, E.S.; Kim, J.H.; Lim, B.K. Fructus Amomi Cardamomi Extract Inhibit Coxsackievirus-B3 Induced Myocarditis in Murine Myocarditis Model. J. Microbiol. Biotechnol. 2016, 26, 2012–2018. [Google Scholar] [CrossRef]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Lam, K.S. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol. 2006, 9, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Losada, A.A.; Cano-Prieto, C.; Garcia-Salcedo, R.; Brana, A.F.; Mendez, C.; Salas, J.A.; Olano, C. Caboxamycin biosynthesis pathway and identification of novel benzoxazoles produced by cross-talk in Streptomyces sp. NTK 937. Microb. Biotechnol. 2017, 10, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, C.; Schneider, K.; Bruntner, C.; Irran, E.; Nicholson, G.; Bull, A.T.; Jones, A.L.; Brown, R.; Stach, J.E.; Goodfellow, M.; et al. Caboxamycin, a new antibiotic of the benzoxazole family produced by the deep-sea strain Streptomyces sp. NTK 937. J. Antibiot. 2009, 62, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Lee, G.H.; Badorff, C.; Dorner, A.; Lee, S.; Wolf, P.; Knowlton, K.U. Dystrophin deficiency markedly increases enterovirus-induced cardiomyopathy: A genetic predisposition to viral heart disease. Nat. Med. 2002, 8, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, K.U.; Jeon, E.S.; Berkley, N.; Wessely, R.; Huber, S. A mutation in the puff region of VP2 attenuates the myocarditic phenotype of an infectious cDNA of the Woodruff variant of coxsackievirus B3. J. Virol. 1996, 70, 7811–7818. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.H.; Jeon, E.S.; Lim, B.K. Macrophage-Specific Coxsackievirus and Adenovirus Receptor Deletion Enhances Macrophage M1 Polarity in CVB3-Induced Myocarditis. Int. J. Mol. Sci. 2023, 24, 5309. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.T., Jr. Myocarditis. N. Engl. J. Med. 2009, 360, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, D.; Yusung, S.; Frisancho, S.; Barrett, M.; Gatewood, S.; Steele, R.; Rose, N.R. IL-12 receptor beta 1 and Toll-like receptor 4 increase IL-1 beta- and IL-18-associated myocarditis and coxsackievirus replication. J. Immunol. 2003, 170, 4731–4737. [Google Scholar] [CrossRef]
- Chen, Y.C.; Yu, C.K.; Wang, Y.F.; Liu, C.C.; Su, I.J.; Lei, H.Y. A murine oral enterovirus 71 infection model with central nervous system involvement. J. Gen. Virol. 2004, 85, 69–77. [Google Scholar] [CrossRef]
- Klingel, K.; Kandolf, R. The role of enterovirus replication in the development of acute and chronic heart muscle disease in different immunocompetent mouse strains. Scand. J. Infect. Dis. Suppl. 1993, 88, 79–85. [Google Scholar]
- Castello, A.; Alvarez, E.; Carrasco, L. Differential cleavage of eIF4GI and eIF4GII in mammalian cells. Effects on translation. J. Biol. Chem. 2006, 281, 33206–33216. [Google Scholar] [CrossRef] [PubMed]
- Mohamud, Y.; Fu, C.; Fan, Y.M.; Zhang, Y.L.; Lin, J.F.C.; Hwang, S.W.; Wang, Z.C.; Luo, H. Activation of cGAS-STING suppresses coxsackievirus replication via interferon-dependent signaling. Antiviral Res. 2024, 222, 105811. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sense (5′ → 3′) | Antisense (5′ → 3′) |
|---|---|---|
| IL-1β | TTG ACG GAC CCC AAA GAG TG | ACT CCT GTA CTC GTG GAA GA |
| IL-6 | GTA CTC CAG AAG ACC AGA GG | TGC TGG TGA CAA CCA CGG CC |
| TNF-α | TTG ACC TCA GCG CTG AGT TG | CCT GTA GCC CAC GTC GTA GC |
| MCP-1 | ACC TGG ATC GGA ACC AAA TG | CCT TAG GGC AGA TGC AGT TTT AA |
| NPPA | TACAGTGCGGTGTCCAACACAG | TGCTTCCTCAGTCTGCTCACTC |
| Myh7 | GCTGAAAGCAGAAAGAGATTATC | TGGAGTTCTTCTCTTCTGGAG |
| GAPDH | ATC AAC GAC CCC TTC ATT GAC C | CCA GTA GAC TCC ACG ACA TAC TCA GC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-G.; Hillman, P.F.; Lee, Y.-J.; Jeon, H.-E.; Lim, B.-K.; Nam, S.-J. Caboxamycin Inhibits Heart Inflammation in a Coxsackievirus B3-Induced Myocarditis Mouse Model. Viruses 2024, 16, 677. https://0-doi-org.brum.beds.ac.uk/10.3390/v16050677
Kim H-G, Hillman PF, Lee Y-J, Jeon H-E, Lim B-K, Nam S-J. Caboxamycin Inhibits Heart Inflammation in a Coxsackievirus B3-Induced Myocarditis Mouse Model. Viruses. 2024; 16(5):677. https://0-doi-org.brum.beds.ac.uk/10.3390/v16050677
Chicago/Turabian StyleKim, Hong-Gi, Prima F. Hillman, You-Jeung Lee, Ha-Eun Jeon, Byung-Kwan Lim, and Sang-Jip Nam. 2024. "Caboxamycin Inhibits Heart Inflammation in a Coxsackievirus B3-Induced Myocarditis Mouse Model" Viruses 16, no. 5: 677. https://0-doi-org.brum.beds.ac.uk/10.3390/v16050677