
The Standard Scrapie Cell Assay: Development, Utility and Prospects

Abstract
:1. Introduction
PrPRes Amplification Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Designation | Species | Tissue or Cell of Origin | Prion Strain | References |
|---|---|---|---|---|
| Neuronal | ||||
| N2a | Mouse | Neuroblastoma | Chandler, RML, 139A, 22L, C506, Fukuoka-1, FU CJD | [11,12,30,31] |
| GT1 | Mouse | Hypothalamic | Chandler, RML, 139A, 22L, FU CJD, M1000 | [30,32] |
| SMB | Mouse | Scrapie-infected mesodermal cells | Chandler, 139A, 22F, 79A | [30,33,34] |
| SN56 | Mouse | Cholinergic septal cells | Chandler, ME7, 22L | [35] |
| CAD | Mouse | Catecholaminergic | RML, 22L, ME7, 301C | [14] |
| PC12 | Rat | Pheochromocytoma | 139A, ME7 | [36,37] |
| Non-Neuronal Cell Lines | ||||
| C2C12 | Mouse | Myotubes | RML, ME7, 22L | [38,39] |
| NIH/3T3 | Mouse | Fibroblast | 22L | [40] |
| moRK13 | Rabbit | Epithelial cell line expressing mouse PrPC | 22L, Chandler, M1000, mo sCJD, Fukuoka-1 | [41,42] |
| voRK13 | Rabbit | Epithelial cell line expressing vole PrPC | vole-adapted BSE | [41] |
| ovRK13/ RoV9 | Rabbit | Epithelial cell line expressing ovine PrPC | PG127, LA404, SSBP/1, scrapie field isolates | [43,44] |
| SSCA Cell Lines | ||||
| PK1 | Mouse | N2a | RML, 22L | [45,46,47] |
| R33 | Mouse | N2a | RML, 22L | [14,46] |
| CAD5 | Mouse | CAD | RML, 22L, ME7, 301C | [14,46] |
| LD9 | Mouse | L929 | RML, 22L, ME7 | [14,46] |
| L929 | Mouse | Fibroblast | RML, 22L, ME7 | [40] |
| Elk21- | Rabbit | RK13: epithelial cell line expressing elk PrPC | CWD | [48] |
2. The Standard Scrapie Cell Assay
2.1. Establishing the Standard Scrapie Cell Assay


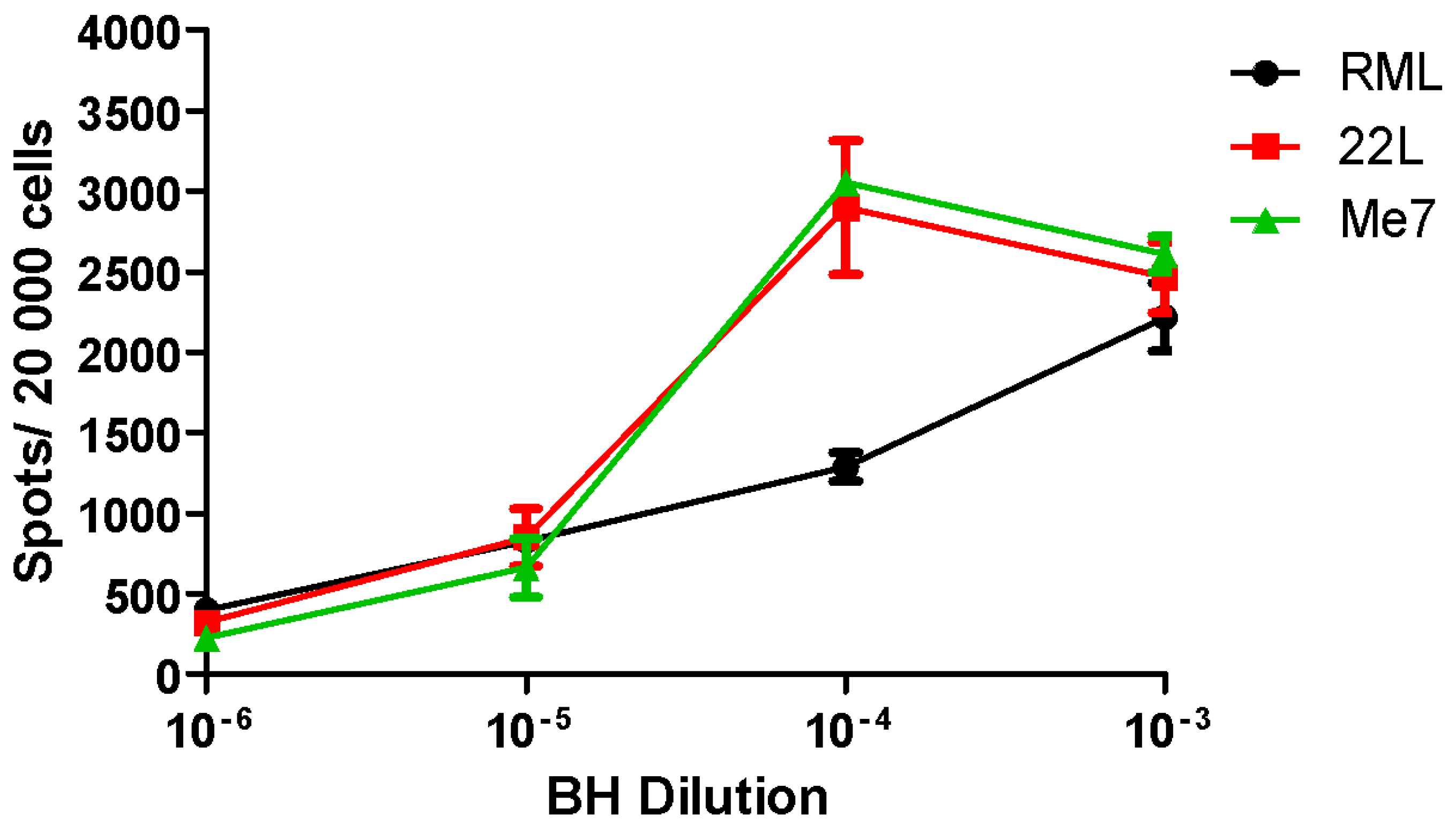
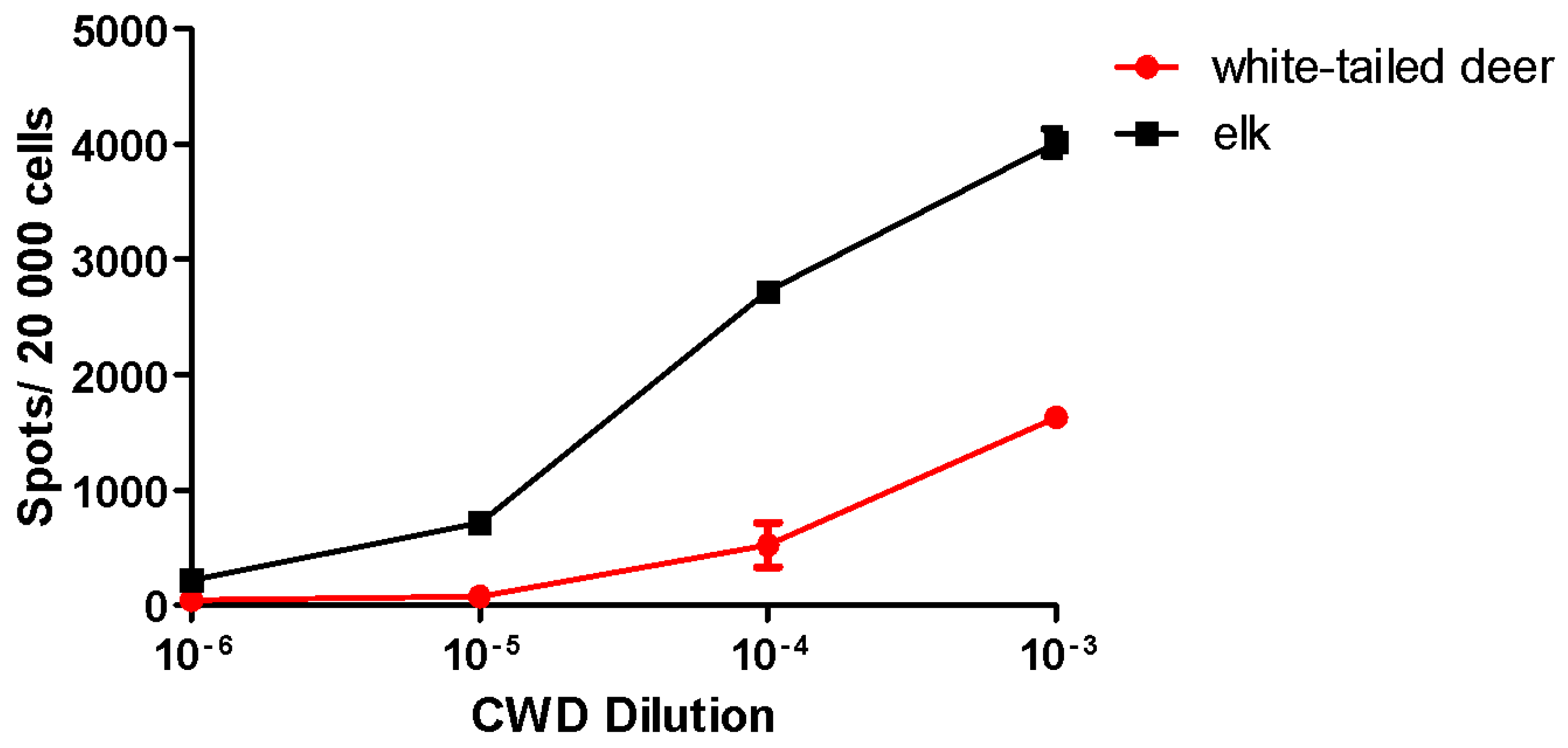
2.2. Utility and Sensitivity
Prion Strain Titration, Discrimination and Analysis
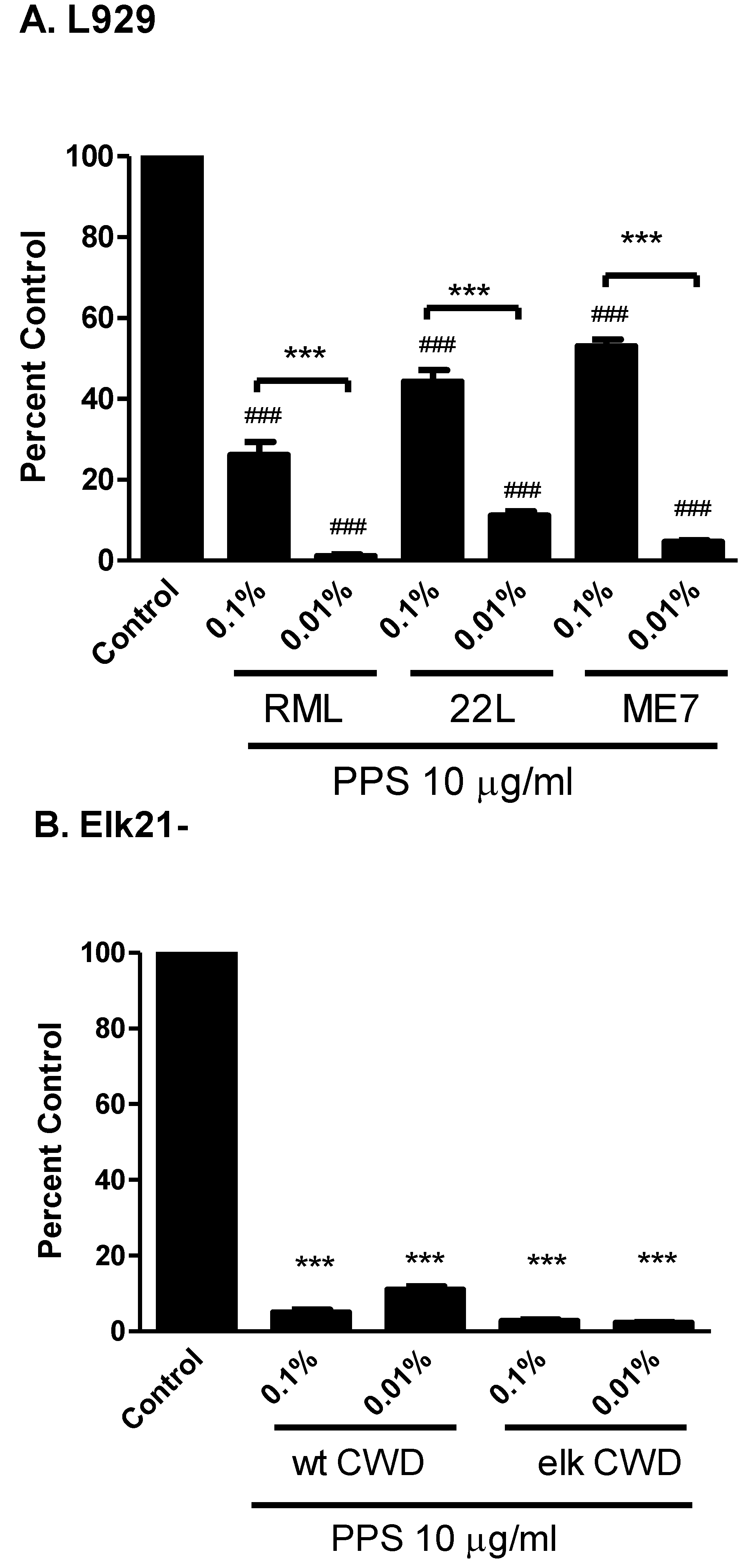
3. Anti-Prion Compounds and Drugs

4. Prion Infectivity and Decontamination Studies
5. Detection of Infectivity from Environmental Samples
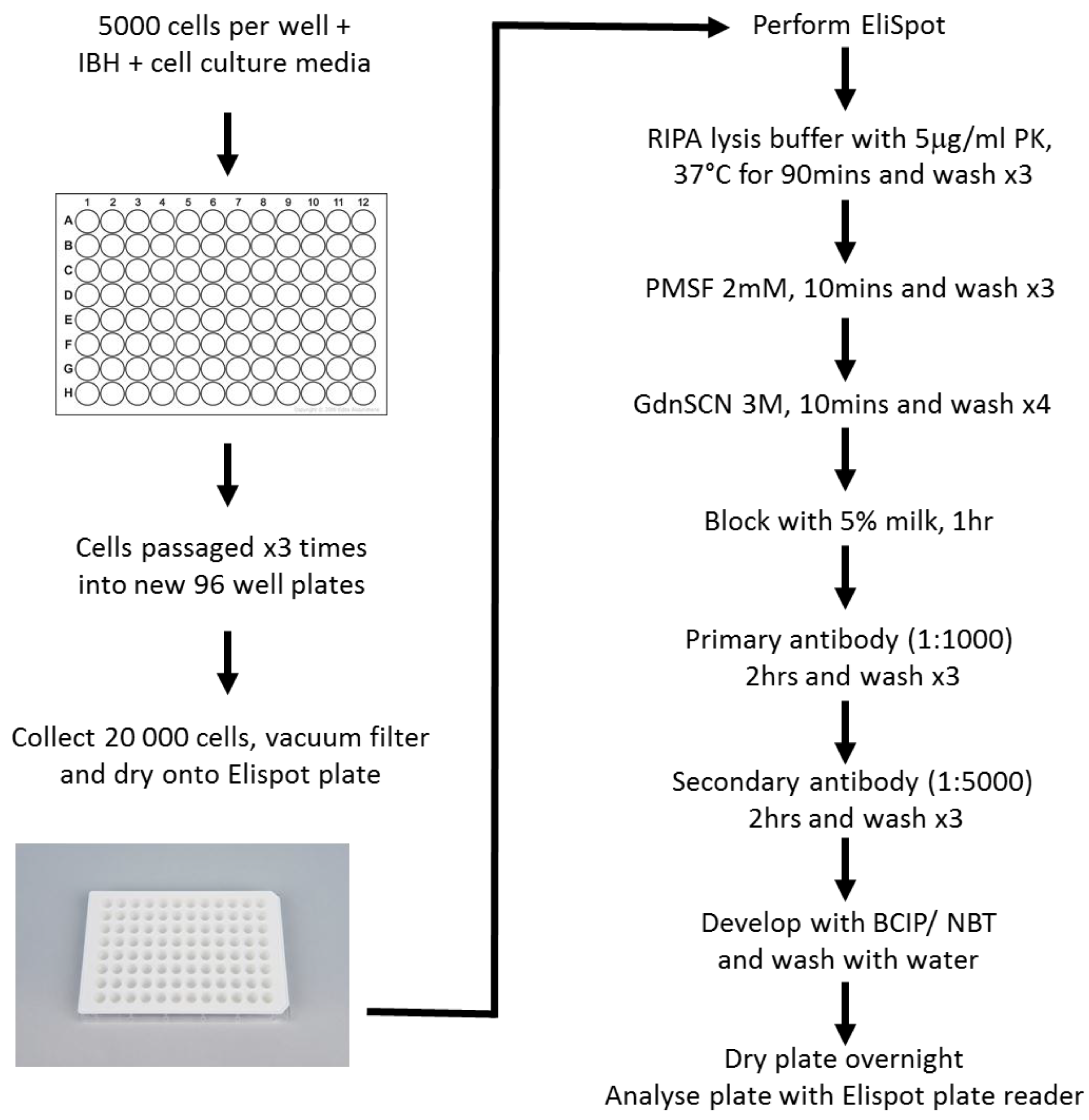
6. SSCA Materials and Methods

7. Limitations of the SSCA
8. Summary and Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Collinge, J. Prion diseases of humans and animals: Their causes and molecular basis. Annu. Rev. Neurosci. 2001, 24, 519–550. [Google Scholar] [CrossRef] [PubMed]
- Prusiner, S.B. Molecular biology of prion diseases. Science 1991, 252, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.S. Self-replication and scrapie. Nature 1967, 215, 1043–1044. [Google Scholar] [CrossRef] [PubMed]
- Bueler, H.; Aguzzi, A.; Sailer, A.; Greiner, R.A.; Autenried, P.; Aguet, M.; Weissmann, C. Mice devoid of prp are resistant to scrapie. Cell 1993, 73, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Knaus, K.J.; Morillas, M.; Swietnicki, W.; Malone, M.; Surewicz, W.K.; Yee, V.C. Crystal structure of the human prion protein reveals a mechanism for oligomerization. Nat. Struct. Biol. 2001, 8, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Riek, R.; Wider, G.; Billeter, M.; Hornemann, S.; Glockshuber, R.; Wuthrich, K. Prion protein nmr structure and familial human spongiform encephalopathies. Proc. Natl. Acad. Sci. USA 1998, 95, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Prusiner, S.B.; Cochran, S.P.; Groth, D.F.; Downey, D.E.; Bowman, K.A.; Martinez, H.M. Measurement of the scrapie agent using an incubation time interval assay. Ann. Neurol. 1982, 11, 353–358. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, I. Prion protein and developments in its detection. Transfus. Med. 2001, 11, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Gyllberg, H.; Löfgren, K. Immunodetection of prpsc using western and slot blotting techniques. In Prion Protein Protocols; Hill, A., Ed.; Humana Press: New York, NY, USA, 2008; Volume 459, pp. 35–48. [Google Scholar]
- Lee, D.C.; Stenland, C.J.; Hartwell, R.C.; Ford, E.K.; Cai, K.; Miller, J.L.; Gilligan, K.J.; Rubenstein, R.; Fournel, M.; Petteway, S.R., Jr. Monitoring plasma processing steps with a sensitive western blot assay for the detection of the prion protein. J. Virol. Methods 2000, 84, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Butler, D.A.; Scott, M.R.; Bockman, J.M.; Borchelt, D.R.; Taraboulos, A.; Hsiao, K.K.; Kingsbury, D.T.; Prusiner, S.B. Scrapie-infected murine neuroblastoma cells produce protease-resistant prion proteins. J. Virol. 1988, 62, 1558–1564. [Google Scholar] [PubMed]
- Nishida, N.; Harris, D.A.; Vilette, D.; Laude, H.; Frobert, Y.; Grassi, J.; Casanova, D.; Milhavet, O.; Lehmann, S. Successful transmission of three mouse-adapted scrapie strains to murine neuroblastoma cell lines overexpressing wild-type mouse prion protein. J. Virol. 2000, 74, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Race, R.E.; Caughey, B.; Graham, K.; Ernst, D.; Chesebro, B. Analyses of frequency of infection, specific infectivity, and prion protein biosynthesis in scrapie-infected neuroblastoma cell clones. J. Virol. 1988, 62, 2845–2849. [Google Scholar] [PubMed]
- Mahal, S.P.; Baker, C.A.; Demczyk, C.A.; Smith, E.W.; Julius, C.; Weissmann, C. Prion strain discrimination in cell culture: The cell panel assay. Proc. Natl. Acad. Sci. USA 2007, 104, 20908–20913. [Google Scholar] [CrossRef] [PubMed]
- Saborio, G.P.; Permanne, B.; Soto, C. Sensitive detection of pathological prion protein by cyclic amplification of protein misfolding. Nature 2001, 411, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, R.; Wilham, J.M.; Christensen, L.; Hughson, A.G.; Moore, R.A.; Johnson, L.M.; Onwubiko, H.A.; Priola, S.A.; Caughey, B. Simplified ultrasensitive prion detection by recombinant prp conversion with shaking. Nat. Methods 2008, 5, 211–212. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Saborio, G.P.; Anderes, L. Cyclic amplification of protein misfolding: Application to prion-related disorders and beyond. Trends Neurosci. 2002, 25, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Castilla, J.; Saa, P.; Morales, R.; Abid, K.; Maundrell, K.; Soto, C. Protein misfolding cyclic amplification for diagnosis and prion propagation studies. Methods Enzymol. 2006, 412, 3–21. [Google Scholar] [PubMed]
- Fujihara, A.; Atarashi, R.; Fuse, T.; Ubagai, K.; Nakagaki, T.; Yamaguchi, N.; Ishibashi, D.; Katamine, S.; Nishida, N. Hyperefficient prp sc amplification of mouse-adapted bse and scrapie strain by protein misfolding cyclic amplification technique. FEBS J. 2009, 276, 2841–2848. [Google Scholar] [CrossRef] [PubMed]
- Barria, M.A.; Gonzalez-Romero, D.; Soto, C. Cyclic amplification of prion protein misfolding. Methods Mol. Biol. 2012, 849, 199–212. [Google Scholar] [PubMed]
- Castilla, J.; Gonzalez-Romero, D.; Saa, P.; Morales, R.; de Castro, J.; Soto, C. Crossing the species barrier by prp(sc) replication in vitro generates unique infectious prions. Cell 2008, 134, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Weber, P.; Giese, A.; Piening, N.; Mitteregger, G.; Thomzig, A.; Beekes, M.; Kretzschmar, H.A. Generation of genuine prion infectivity by serial pmca. Vet. Microbiol. 2007, 123, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Barria, M.A.; Mukherjee, A.; Gonzalez-Romero, D.; Morales, R.; Soto, C. De novo generation of infectious prions in vitro produces a new disease phenotype. PLoS Pathog. 2009, 5, e1000421. [Google Scholar] [CrossRef] [PubMed]
- Shaked, G.M.; Meiner, Z.; Avraham, I.; Taraboulos, A.; Gabizon, R. Reconstitution of prion infectivity from solubilized protease-resistant prp and nonprotein components of prion rods. J. Biol. Chem. 2001, 276, 14324–14328. [Google Scholar] [CrossRef] [PubMed]
- Deleault, N.R.; Lucassen, R.W.; Supattapone, S. Rna molecules stimulate prion protein conversion. Nature 2003, 425, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, X.; Yuan, C.G.; Ma, J. Generating a prion with bacterially expressed recombinant prion protein. Science 2010, 327, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Saunders, S.E.; Bartz, J.C.; Shikiya, R.A. Protein misfolding cyclic amplification of prions. J. Vis. Exp. 2012. [Google Scholar] [CrossRef]
- Saa, P.; Sferrazza, G.F.; Ottenberg, G.; Oelschlegel, A.M.; Dorsey, K.; Lasmezas, C.I. Strain-specific role of rnas in prion replication. J. Virol. 2012, 86, 10494–10504. [Google Scholar] [CrossRef] [PubMed]
- Cosseddu, G.M.; Nonno, R.; Vaccari, G.; Bucalossi, C.; Fernandez-Borges, N.; di Bari, M.A.; Castilla, J.; Agrimi, U. Ultra-efficient prp(sc) amplification highlights potentialities and pitfalls of pmca technology. PLoS Pathog. 2011, 7, e1002370. [Google Scholar] [CrossRef] [PubMed]
- Grassmann, A.; Wolf, H.; Hofmann, J.; Graham, J.; Vorberg, I. Cellular aspects of prion replication in vitro. Viruses 2013, 5, 374–405. [Google Scholar] [CrossRef] [PubMed]
- Race, R. The scrapie agent in vitro. Curr. Top. Microbiol. Immunol. 1991, 172, 181–193. [Google Scholar] [PubMed]
- Schatzl, H.M.; Laszlo, L.; Holtzman, D.M.; Tatzelt, J.; DeArmond, S.J.; Weiner, R.I.; Mobley, W.C.; Prusiner, S.B. A hypothalamic neuronal cell line persistently infected with scrapie prions exhibits apoptosis. J. Virol. 1997, 71, 8821–8831. [Google Scholar] [PubMed]
- Birkett, C.R.; Hennion, R.M.; Bembridge, D.A.; Clarke, M.C.; Chree, A.; Bruce, M.E.; Bostock, C.J. Scrapie strains maintain biological phenotypes on propagation in a cell line in culture. EMBO J. 2001, 20, 3351–3358. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.C.; Haig, D.A. Evidence for the multiplication of scrapie agent in cell culture. Nature 1970, 225, 100–101. [Google Scholar] [CrossRef] [PubMed]
- Baron, G.S.; Magalhaes, A.C.; Prado, M.A.; Caughey, B. Mouse-adapted scrapie infection of sn56 cells: Greater efficiency with microsome-associated versus purified prp-res. J. Virol. 2006, 80, 2106–2117. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, R.; Carp, R.I.; Callahan, S.M. In vitro replication of scrapie agent in a neuronal model: Infection of pc12 cells. J. Gen. Virol. 1984, 65, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, R.; Deng, H.; Race, R.E.; Ju, W.; Scalici, C.L.; Papini, M.C.; Kascsak, R.J.; Carp, R.I. Demonstration of scrapie strain diversity in infected pc12 cells. J. Gen. Virol. 1992, 73, 3027–3031. [Google Scholar] [CrossRef] [PubMed]
- Dlakic, W.M.; Grigg, E.; Bessen, R.A. Prion infection of muscle cells in vitro. J. Virol. 2007, 81, 4615–4624. [Google Scholar] [CrossRef] [PubMed]
- Herbst, A.; Banser, P.; Velasquez, C.D.; Mays, C.E.; Sim, V.L.; Westaway, D.; Aiken, J.M.; McKenzie, D. Infectious prions accumulate to high levels in non proliferative c2c12 myotubes. PLoS Pathog. 2013, 9, e1003755. [Google Scholar] [CrossRef] [PubMed]
- Vorberg, I.; Raines, A.; Story, B.; Priola, S.A. Susceptibility of common fibroblast cell lines to transmissible spongiform encephalopathy agents. J. Infect. Dis. 2004, 189, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Courageot, M.P.; Daude, N.; Nonno, R.; Paquet, S.; di Bari, M.A.; le Dur, A.; Chapuis, J.; Hill, A.F.; Agrimi, U.; Laude, H.; et al. A cell line infectible by prion strains from different species. J. Gen. Virol. 2008, 89, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Lawson, V.A.; Vella, L.J.; Stewart, J.D.; Sharples, R.A.; Klemm, H.; Machalek, D.M.; Masters, C.L.; Cappai, R.; Collins, S.J.; Hill, A.F. Mouse-adapted sporadic human creutzfeldt-jakob disease prions propagate in cell culture. Int. J. Biochem. Cell Biol. 2008, 40, 2793–2801. [Google Scholar] [CrossRef] [PubMed]
- Vilette, D.; Andreoletti, O.; Archer, F.; Madelaine, M.F.; Vilotte, J.L.; Lehmann, S.; Laude, H. Ex vivo propagation of infectious sheep scrapie agent in heterologous epithelial cells expressing ovine prion protein. Proc. Natl. Acad. Sci. USA 2001, 98, 4055–4059. [Google Scholar] [CrossRef] [PubMed]
- Sabuncu, E.; Petit, S.; le Dur, A.; Lan Lai, T.; Vilotte, J.L.; Laude, H.; Vilette, D. Prp polymorphisms tightly control sheep prion replication in cultured cells. J. Virol. 2003, 77, 2696–2700. [Google Scholar] [CrossRef] [PubMed]
- Klohn, P.C.; Stoltze, L.; Flechsig, E.; Enari, M.; Weissmann, C. A quantitative, highly sensitive cell-based infectivity assay for mouse scrapie prions. Proc. Natl. Acad. Sci. USA 2003, 100, 11666–11671. [Google Scholar] [CrossRef] [PubMed]
- Mahal, S.P.; Demczyk, C.A.; Smith, E.W., Jr.; Klohn, P.C.; Weissmann, C. Assaying prions in cell culture: The standard scrapie cell assay (ssca) and the scrapie cell assay in end point format (scepa). Methods Mol. Biol. 2008, 459, 49–68. [Google Scholar] [PubMed]
- Enari, M.; Flechsig, E.; Weissmann, C. Scrapie prion protein accumulation by scrapie-infected neuroblastoma cells abrogated by exposure to a prion protein antibody. Proc. Natl. Acad. Sci. USA 2001, 98, 9295–9299. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Napier, D.; Khaychuck, V.; Angers, R.; Graham, C.; Telling, G. Cell-based quantification of chronic wasting disease prions. J. Virol. 2010, 84, 8322–8326. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, R.; Satoh, K.; Sano, K.; Fuse, T.; Yamaguchi, N.; Ishibashi, D.; Matsubara, T.; Nakagaki, T.; Yamanaka, H.; Shirabe, S.; et al. Ultrasensitive human prion detection in cerebrospinal fluid by real-time quaking-induced conversion. Nat. Med. 2011, 17, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Orru, C.D.; Wilham, J.M.; Vascellari, S.; Hughson, A.G.; Caughey, B. New generation quic assays for prion seeding activity. Prion 2012, 6, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Wilham, J.M.; Orru, C.D.; Bessen, R.A.; Atarashi, R.; Sano, K.; Race, B.; Meade-White, K.D.; Taubner, L.M.; Timmes, A.; Caughey, B. Rapid end-point quantitation of prion seeding activity with sensitivity comparable to bioassays. PLoS Pathog. 2010, 6, e1001217. [Google Scholar] [CrossRef] [PubMed]
- Colby, D.W.; Zhang, Q.; Wang, S.; Groth, D.; Legname, G.; Riesner, D.; Prusiner, S.B. Prion detection by an amyloid seeding assay. Proc. Natl. Acad. Sci. USA 2007, 104, 20914–20919. [Google Scholar] [CrossRef] [PubMed]
- McGuire, L.I.; Peden, A.H.; Orru, C.D.; Wilham, J.M.; Appleford, N.E.; Mallinson, G.; Andrews, M.; Head, M.W.; Caughey, B.; Will, R.G.; et al. Real time quaking-induced conversion analysis of cerebrospinal fluid in sporadic creutzfeldt-jakob disease. Ann. Neurol. 2012, 72, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Bosque, P.J.; Prusiner, S.B. Cultured cell sublines highly susceptible to prion infection. J. Virol. 2000, 74, 4377–4386. [Google Scholar] [CrossRef] [PubMed]
- Kocisko, D.A.; Caughey, B. Searching for anti-prion compounds: Cell-based high-throughput in vitro assays and animal testing strategies. Methods Enzymol. 2006, 412, 223–234. [Google Scholar] [PubMed]
- Leblanc, P.; Alais, S.; Porto-Carreiro, I.; Lehmann, S.; Grassi, J.; Raposo, G.; Darlix, J.L. Retrovirus infection strongly enhances scrapie infectivity release in cell culture. EMBO J. 2006, 25, 2674–2685. [Google Scholar] [CrossRef] [PubMed]
- Collinge, J.; Sidle, K.C.; Meads, J.; Ironside, J.; Hill, A.F. Molecular analysis of prion strain variation and the aetiology of “new variant” cjd. Nature 1996, 383, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Beringue, V.; Bencsik, A.; le Dur, A.; Reine, F.; Lai, T.L.; Chenais, N.; Tilly, G.; Biacabe, A.G.; Baron, T.; Vilotte, J.L.; et al. Isolation from cattle of a prion strain distinct from that causing bovine spongiform encephalopathy. PLoS Pathog. 2006, 2, e112. [Google Scholar] [CrossRef] [PubMed]
- Fraser, H.; Bruce, M. Argyrophilic plaques in mice inoculated with scrapie from particular sources. Lancet 1973, 1, 617–618. [Google Scholar] [CrossRef] [PubMed]
- Bruce, M.E.; McBride, P.A.; Farquhar, C.F. Precise targeting of the pathology of the sialoglycoprotein, prp, and vacuolar degeneration in mouse scrapie. Neurosci. Lett. 1989, 102, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fraser, H.; Dickinson, A.G. The sequential development of the brain lesion of scrapie in three strains of mice. J. Comp. Pathol. 1968, 78, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Jenny, A.L.; Taylor, W.D.; Marsh, R.F.; Rubenstein, R.; Race, R.E. Immunohistochemical detection of prion protein in sheep with scrapie. J. Vet. Diagn. Investig. 1993, 5, 309–316. [Google Scholar] [CrossRef]
- DeArmond, S.J.; Yang, S.L.; Lee, A.; Bowler, R.; Taraboulos, A.; Groth, D.; Prusiner, S.B. Three scrapie prion isolates exhibit different accumulation patterns of the prion protein scrapie isoform. Proc. Natl. Acad. Sci. USA 1993, 90, 6449–6453. [Google Scholar] [CrossRef] [PubMed]
- Oelschlegel, A.M.; Fallahi, M.; Ortiz-Umpierre, S.; Weissmann, C. The extended cell panel assay characterizes the relationship of prion strains rml, 79a, and 139a and reveals conversion of 139a to 79a-like prions in cell culture. J. Virol. 2012, 86, 5297–5303. [Google Scholar] [CrossRef] [PubMed]
- Mahal, S.P.; Jablonski, J.; Suponitsky-Kroyter, I.; Oelschlegel, A.M.; Herva, M.E.; Oldstone, M.; Weissmann, C. Propagation of rml prions in mice expressing prp devoid of gpi anchor leads to formation of a novel, stable prion strain. PLoS Pathog. 2012, 8, e1002746. [Google Scholar] [CrossRef] [PubMed]
- Vorberg, I.; Priola, S.A. Molecular basis of scrapie strain glycoform variation. J. Biol. Chem. 2002, 277, 36775–36781. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Johnson, J.; Vanderloo, J.P.; Keane, D.; Aiken, J.M.; McKenzie, D. Prion protein polymorphisms in white-tailed deer influence susceptibility to chronic wasting disease. J. Gen. Virol. 2006, 87, 2109–2114. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, K.I.; Spraker, T.R.; Hamburg, L.K.; Besser, T.E.; Brayton, K.A.; Knowles, D.P. Polymorphisms in the prion precursor functional gene but not the pseudogene are associated with susceptibility to chronic wasting disease in white-tailed deer. J. Gen. Virol. 2004, 85, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, K.I.; Besser, T.E.; Miller, M.W.; Cline, T.F.; Spraker, T.R.; Jenny, A.L.; Wild, M.A.; Zebarth, G.L.; Williams, E.S. Prp genotypes of captive and free-ranging rocky mountain elk (cervus elaphus nelsoni) with chronic wasting disease. J. Gen. Virol. 1999, 80, 2765–2769. [Google Scholar] [PubMed]
- Trevitt, C.R.; Collinge, J. A systematic review of prion therapeutics in experimental models. Brain: J. Neurol. 2006, 129, 2241–2265. [Google Scholar] [CrossRef]
- McCarthy, J.M.; Franke, M.; Resenberger, U.K.; Waldron, S.; Simpson, J.C.; Tatzelt, J.; Appelhans, D.; Rogers, M.S. Anti-prion drug mppig5 inhibits prp(c) conversion to prp(sc). PLoS One 2013, 8, e55282. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Kang, H.E.; Telling, G.C. Quinacrine promotes replication and conformational mutation of chronic wasting disease prions. Proc. Natl. Acad. Sci. USA 2014, 111, 6028–6033. [Google Scholar] [CrossRef] [PubMed]
- Edgeworth, J.A.; Jackson, G.S.; Clarke, A.R.; Weissmann, C.; Collinge, J. Highly sensitive, quantitative cell-based assay for prions adsorbed to solid surfaces. Proc. Natl. Acad. Sci. USA 2009, 106, 3479–3483. [Google Scholar] [CrossRef] [PubMed]
- Zobeley, E.; Flechsig, E.; Cozzio, A.; Enari, M.; Weissmann, C. Infectivity of scrapie prions bound to a stainless steel surface. Mol. Med. 1999, 5, 240–243. [Google Scholar] [PubMed]
- Flechsig, E.; Hegyi, I.; Enari, M.; Schwarz, P.; Collinge, J.; Weissmann, C. Transmission of scrapie by steel-surface-bound prions. Mol. Med. 2001, 7, 679–684. [Google Scholar] [PubMed]
- Weissmann, C.; Enari, M.; Klohn, P.C.; Rossi, D.; Flechsig, E. Transmission of prions. J. Infect. Dis. 2002, 186 (Suppl. 2), S157–S165. [Google Scholar] [CrossRef]
- Gregori, L. A prototype assay to detect vcjd-infected blood. Lancet 2011, 377, 444–446. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W.; Williams, E.S.; Hobbs, N.T.; Wolfe, L.L. Environmental sources of prion transmission in mule deer. Emerg. Infect. Dis. 2004, 10, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Saunders, S.E.; Bartelt-Hunt, S.L.; Bartz, J.C. Prions in the environment: Occurrence, fate and mitigation. Prion 2008, 2, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W.; Williams, E.S. Prion disease: Horizontal prion transmission in mule deer. Nature 2003, 425, 35–36. [Google Scholar] [CrossRef] [PubMed]
- Pulford, B.; Spraker, T.R.; Wyckoff, A.C.; Meyerett, C.; Bender, H.; Ferguson, A.; Wyatt, B.; Lockwood, K.; Powers, J.; Telling, G.C.; et al. Detection of prpcwd in feces from naturally exposed rocky mountain elk (cervus elaphus nelsoni) using protein misfolding cyclic amplification. J. Wildl. Dis. 2012, 48, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Seeger, H.; Heikenwalder, M.; Zeller, N.; Kranich, J.; Schwarz, P.; Gaspert, A.; Seifert, B.; Miele, G.; Aguzzi, A. Coincident scrapie infection and nephritis lead to urinary prion excretion. Science 2005, 310, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Mathiason, C.K.; Powers, J.G.; Dahmes, S.J.; Osborn, D.A.; Miller, K.V.; Warren, R.J.; Mason, G.L.; Hays, S.A.; Hayes-Klug, J.; Seelig, D.M.; et al. Infectious prions in the saliva and blood of deer with chronic wasting disease. Science 2006, 314, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Genovesi, S.; Leita, L.; Sequi, P.; Andrighetto, I.; Sorgato, M.C.; Bertoli, A. Direct detection of soil-bound prions. PLoS One 2007, 2, e1069. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.J.; Phillips, K.E.; Schramm, P.T.; McKenzie, D.; Aiken, J.M.; Pedersen, J.A. Prions adhere to soil minerals and remain infectious. PLoS Pathog. 2006, 2, e32. [Google Scholar] [CrossRef] [PubMed]
- Chasseigneaux, S.; Pastore, M.; Britton-Davidian, J.; Manie, E.; Stern, M.H.; Callebert, J.; Catalan, J.; Casanova, D.; Belondrade, M.; Provansal, M.; et al. Genetic heterogeneity versus molecular analysis of prion susceptibility in neuroblasma n2a sublines. Arch. Virol. 2008, 153, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Herva, M.E.; Weissmann, C. Cell-specific susceptibility to prion strains is a property of the intact cell. Prion 2012, 6, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Neale, M.H.; Mountjoy, S.J.; Edwards, J.C.; Vilette, D.; Laude, H.; Windl, O.; Saunders, G.C. Infection of cell lines with experimental and natural ovine scrapie agents. J. Virol. 2010, 84, 2444–2452. [Google Scholar] [CrossRef] [PubMed]
- Edgeworth, J.A.; Gros, N.; Alden, J.; Joiner, S.; Wadsworth, J.D.; Linehan, J.; Brandner, S.; Jackson, G.S.; Weissmann, C.; Collinge, J. Spontaneous generation of mammalian prions. Proc. Natl. Acad. Sci. USA 2010, 107, 14402–14406. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van der Merwe, J.; Aiken, J.; Westaway, D.; McKenzie, D. The Standard Scrapie Cell Assay: Development, Utility and Prospects. Viruses 2015, 7, 180-198. https://0-doi-org.brum.beds.ac.uk/10.3390/v7010180
Van der Merwe J, Aiken J, Westaway D, McKenzie D. The Standard Scrapie Cell Assay: Development, Utility and Prospects. Viruses. 2015; 7(1):180-198. https://0-doi-org.brum.beds.ac.uk/10.3390/v7010180
Chicago/Turabian StyleVan der Merwe, Jacques, Judd Aiken, David Westaway, and Debbie McKenzie. 2015. "The Standard Scrapie Cell Assay: Development, Utility and Prospects" Viruses 7, no. 1: 180-198. https://0-doi-org.brum.beds.ac.uk/10.3390/v7010180