
Telomere Length, Proviral Load and Neurologic Impairment in HTLV-1 and HTLV-2-Infected Subjects

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Subjects
2.2. Laboratory Measures
2.3. Statistical Analysis
3. Results
3.1. Study Population
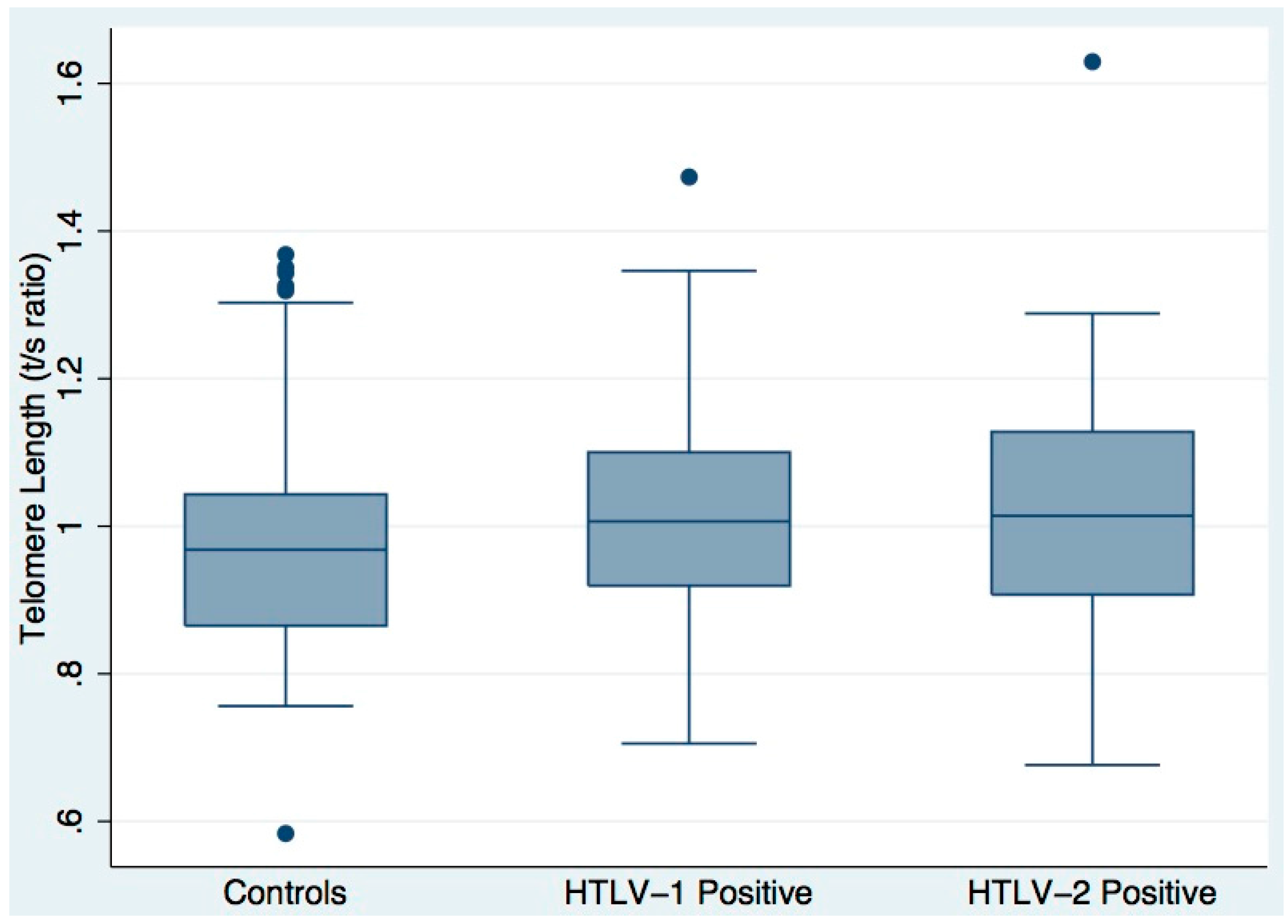
3.2. Telomere Length
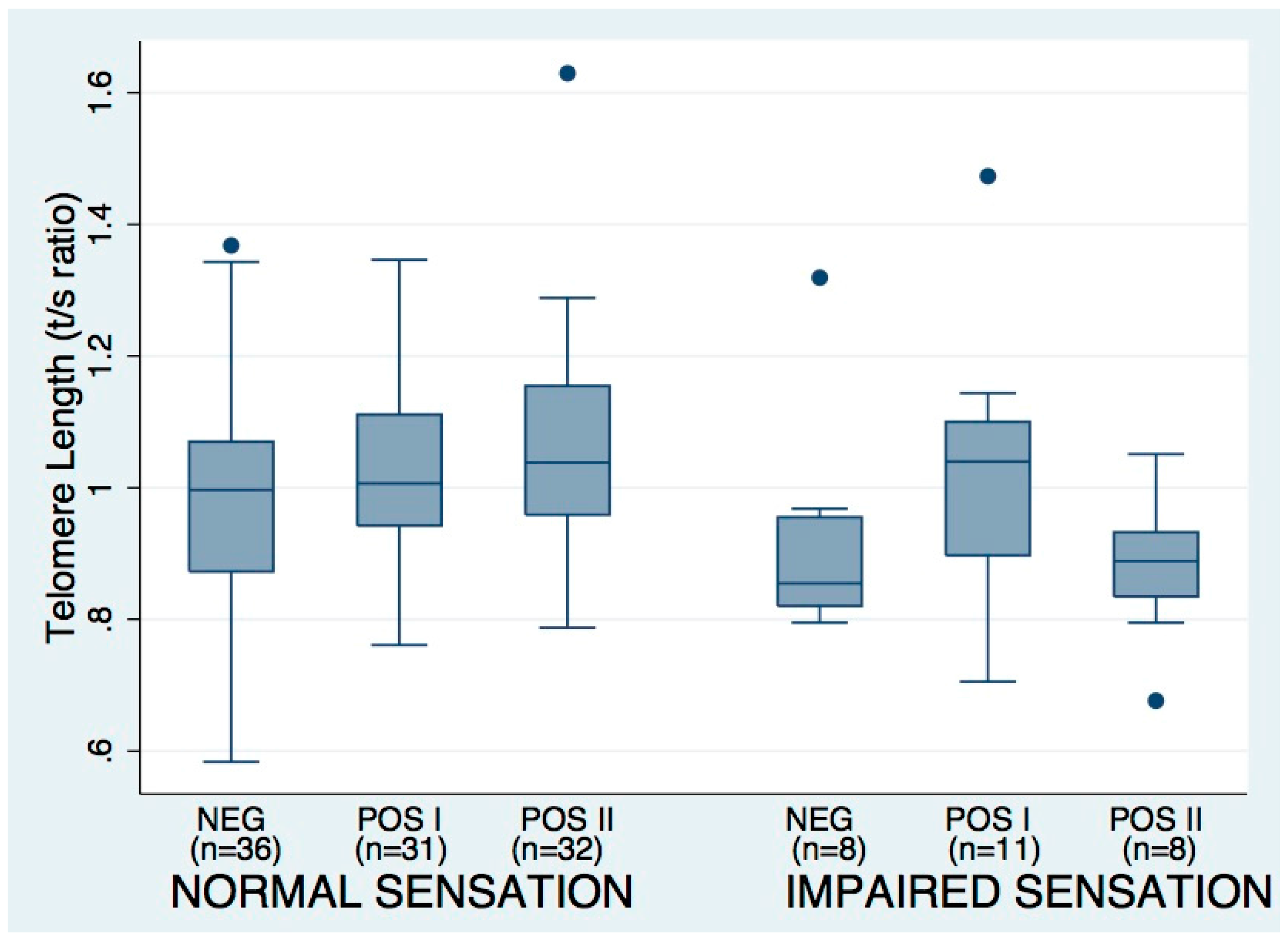
3.3. Tuning Fork Sensation Impairment Associations
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Orland, J.R.; Wang, B.; Wright, D.J.; Nass, C.C.; Garratty, G.; Smith, J.W.; Newman, B.; Smith, D.M.; Murphy, E.L. Increased mortality associated with htlv-ii infection in blood donors: A prospective cohort study. Retrovirology 2004, 1, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arisawa, K.; Sobue, T.; Yoshimi, I.; Soda, M.; Shirahama, S.; Doi, H.; Katamine, S.; Saito, H.; Urata, M. Human t-lymphotropic virus type-i infection, survival and cancer risk in southwestern japan: A prospective cohort study. Cancer Causes Control 2003, 14, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Biswas, H.H.; Kaidarova, Z.M.A.; Garratty, G.; Gibble, J.W.; Newman, B.H.; Smith, J.W.; Ziman, A.; Fridey, J.L.; Sacher, R.A.; Murphy, E.L. Increased all-cause and cancer mortality in htlv-ii infection. J. Acquir. Immune Defic. Syndr. 2010, 54, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Van Tienen, C.; van der Schim Loeff, M.; Peterson, I.; Cotten, M.; Andersson, S.; Holmgren, B.; Vincent, T.; de Silva, T.; Rowland-Jones, S.; Aaby, P.; et al. HTLV-1 and HIV-2 infection are associated with increased mortality in a rural west african community. PLoS ONE 2011, 6, e29026. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.L.; Glynn, S.A.; Fridey, J.; Smith, J.W.; Sacher, R.A.; Nass, C.C.; Ownby, H.E.; Wright, D.J.; Nemo, G.J. Increased incidence of infectious diseases during prospective follow-up of human t-lymphotropic virus type II- and I-infected blood donors. Retrovirus epidemiology donor study. Arch. Intern. Med. 1999, 159, 1485–1491. [Google Scholar] [CrossRef] [PubMed]
- Biswas, H.H.; Engstrom, J.W.; Kaidarova, Z.; Garratty, G.; Gibble, J.W.; Newman, B.H.; Smith, J.W.; Ziman, A.; Fridey, J.L.; Sacher, R.A.; et al. Neurologic abnormalities in HTLV-I- and HTLV-II-infected individuals without overt myelopathy. Neurology 2009, 73, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Ziskoven, C.; Jager, M.; Kircher, J.; Patzer, T.; Bloch, W.; Brixius, K.; Krauspe, R. Physiology and pathophysiology of nitrosative and oxidative stress in osteoarthritic joint destruction. Can. J. Physiol. Pharmacol. 2011, 89, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Yudoh, K.; Nguyen, V.T.; Nakamura, H.; Hongo-Masuko, K.; Kato, T.; Nishioka, K. Potential involvement of oxidative stress in cartilage senescence and development of osteoarthritis: Oxidative stress induces chondrocyte telomere instability and downregulation of chondrocyte function. Arthritis Res. Ther. 2005, 7, R380–R391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Moreno, M.; Tamayo, M.; Soto-Hermida, A.; Mosquera, A.; Oreiro, N.; Fernandez-Lopez, C.; Fernandez, J.L.; Rego-Perez, I.; Blanco, F.J. Mtdna haplogroup j modulates telomere length and nitric oxide production. BMC Musculoskelet. Disord. 2011, 12, 283. [Google Scholar] [CrossRef] [PubMed]
- Tattermusch, S.; Skinner, J.A.; Chaussabel, D.; Banchereau, J.; Berry, M.P.; McNab, F.W.; O’Garra, A.; Taylor, G.P.; Bangham, C.R. Systems biology approaches reveal a specific interferon-inducible signature in HTLV-1 associated myelopathy. PLoS Pathog. 2012, 8, e1002480. [Google Scholar] [CrossRef] [PubMed]
- Calado, R.T.; Young, N.S. Telomere diseases. N. Engl. J. Med. 2009, 361, 2353–2365. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Shao, L.; Colmegna, I.; Goronzy, J.J.; Weyand, C.M. Telomerase insufficiency in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2009, 106, 4360–4365. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, A.L.; Kronmal, R.A.; Kimura, M.; Gardner, J.P.; Psaty, B.M.; Jenny, N.S.; Tracy, R.P.; Hardikar, S.; Aviv, A. Leukocyte telomere length and mortality in the cardiovascular health study. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2011, 66, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Codd, V.; Nelson, C.P.; Albrecht, E.; Mangino, M.; Deelen, J.; Buxton, J.L.; Hottenga, J.J.; Fischer, K.; Esko, T.; Surakka, I.; et al. Identification of seven loci affecting mean telomere length and their association with disease. Nat. Genet. 2013, 45, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Haycock, P.C.; Heydon, E.E.; Kaptoge, S.; Butterworth, A.S.; Thompson, A.; Willeit, P. Leucocyte telomere length and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2014, 349, g4227. [Google Scholar] [CrossRef] [PubMed]
- Shammas, M.A. Telomeres, lifestyle, cancer, and aging. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Kubuki, Y.; Suzuki, M.; Sasaki, H.; Toyama, T.; Yamashita, K.; Maeda, K.; Ido, A.; Matsuoka, H.; Okayama, A.; Nakanishi, T.; et al. Telomerase activity and telomere length as prognostic factors of adult T-cell leukemia. Leuk. Lymphoma 2005, 46, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Gabet, A.S.; Mortreux, F.; Charneau, P.; Riou, P.; Duc-Dodon, M.; Wu, Y.; Jeang, K.T.; Wattel, E. Inactivation of htert transcription by tax. Oncogene 2003, 22, 3734–3741. [Google Scholar] [CrossRef] [PubMed]
- Bellon, M.; Nicot, C. Telomerase: A crucial player in htlv-i-induced human t-cell leukemia. Cancer Genom. Proteom. 2007, 4, 21–25. [Google Scholar]
- Zane, L.; Sibon, D.; Capraro, V.; Galia, P.; Karam, M.; Delfau-Larue, M.H.; Gilson, E.; Gessain, A.; Gout, O.; Hermine, O.; et al. HTLV-1 positive and negative T cells cloned from infected individuals display telomerase and telomere genes deregulation that predominate in activated but untransformed CD4+ T cells. Int. J. Cancer 2012, 131, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Sinha-Datta, U.; Horikawa, I.; Michishita, E.; Datta, A.; Sigler-Nicot, J.C.; Brown, M.; Kazanji, M.; Barrett, J.C.; Nicot, C. Transcriptional activation of hTERT through the NF-kappab pathway in HTLV-I-transformed cells. Blood 2004, 104, 2523–2531. [Google Scholar] [CrossRef] [PubMed]
- Song, L.L.; Ponomareva, L.; Shen, H.; Duan, X.; Alimirah, F.; Choubey, D. Interferon-inducible IFI16, a negative regulator of cell growth, down-regulates expression of human telomerase reverse transcriptase (hTERT) gene. PLoS ONE 2010, 5, e8569. [Google Scholar] [CrossRef] [PubMed]
- Perkins, B.A.; Olaleye, D.; Zinman, B.; Bril, V. Simple screening tests for peripheral neuropathy in the diabetes clinic. Diabetes Care 2001, 24, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Martina, I.S.; van Koningsveld, R.; Schmitz, P.I.; van der Meche, F.G.; van Doorn, P.A. Measuring vibration threshold with a graduated tuning fork in normal aging and in patients with polyneuropathy. European inflammatory neuropathy cause and treatment (incat) group. J. Neurol. Neurosurg. Psychiatry 1998, 65, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Epel, E.; Cheon, J.; Kroenke, C.; Sinclair, E.; Bigos, M.; Wolkowitz, O.; Mellon, S.; Blackburn, E. Analyses and comparisons of telomerase activity and telomere length in human T and B cells: Insights for epidemiology of telomere maintenance. J. Immunol. Methods 2010, 352, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Chafets, D.M.; Busch, M.P.; Murphy, E.L. Quantitation of htlv-i and ii proviral load using real-time quantitative pcr with sybr green chemistry. J. Clin. Virol. 2004, 31, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Aubert, G.; Lansdorp, P.M. Telomeres and aging. Physiol. Rev. 2008, 88, 557–579. [Google Scholar] [CrossRef] [PubMed]
- Iwama, H.; Ohyashiki, K.; Ohyashiki, J.H.; Hayashi, S.; Yahata, N.; Ando, K.; Toyama, K.; Hoshika, A.; Takasaki, M.; Mori, M.; et al. Telomeric length and telomerase activity vary with age in peripheral blood cells obtained from normal individuals. Hum. Genet. 1998, 102, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.; Usuku, K.; Matsumoto, W.; Kodama, D.; Takenouchi, N.; Moritoyo, T.; Hashiguchi, S.; Ichinose, M.; Bangham, C.R.; Izumo, S.; et al. Analysis of HTLV-I proviral load in 202 HAM/TSP patients and 243 asymptomatic HTLV-I carriers: High proviral load strongly predisposes to HAM/TSP. J. Neurovirol. 1998, 4, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Dowd, J.B.; Bosch, J.A.; Steptoe, A.; Blackburn, E.H.; Lin, J.; Rees-Clayton, E.; Aiello, A.E. Cytomegalovirus is associated with reduced telomerase activity in the whitehall II cohort. Exp. Gerontol. 2013, 48, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Mirabello, L.; Huang, W.Y.; Wong, J.Y.; Chatterjee, N.; Reding, D.; Crawford, E.D.; De Vivo, I.; Hayes, R.B.; Savage, S.A. The association between leukocyte telomere length and cigarette smoking, dietary and physical variables, and risk of prostate cancer. Aging Cell 2009, 8, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, Y. Pathology of chronic myelopathy associated with HTLV-I infection (HAM/TSP). J. Neurol. Sci. 1990, 96, 103–123. [Google Scholar] [CrossRef]
- Sato, K.; Niessner, A.; Kopecky, S.L.; Frye, R.L.; Goronzy, J.J.; Weyand, C.M. Trail-expressing T cells induce apoptosis of vascular smooth muscle cells in the atherosclerotic plaque. J. Exp. Med. 2006, 203, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Fidanboylu, M.; Griffiths, L.A.; Flatters, S.J. Global inhibition of reactive oxygen species (ROS) inhibits paclitaxel-induced painful peripheral neuropathy. PLoS ONE 2011, 6, e25212. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, M.; Sasannejad, P.; Foroughipour, M.; Shahami, S.; Shoeibi, A. Prevalence of peripheral neuropathy in patients with HTLV-1 associated myelopathy/tropical spastic paraparesis (HAM/TSP). Acta Neurol. Belg. 2011, 111, 41–44. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| HTLV-NEG | HTLV-1 POS | HTLV-2 POS | |
|---|---|---|---|
| n = 45 | n = 45 | n = 45 | |
| Age | |||
| 35–45 | 3 | 3 | 3 |
| 45–55 | 12 | 12 | 12 |
| >56 | 30 | 30 | 30 |
| Mean, Median | 58.5, 58.7 | 58.4, 59.1 | 60.1, 59.3 |
| Sex | |||
| Male | 8 | 8 | 8 |
| Female | 37 | 37 | 37 |
| Race/Ethnicity | |||
| White | 15 | 15 | 15 |
| Black | 22 | 22 | 22 |
| Hispanic * | 6 | 3 | 8 |
| Japanese * | 1 | 4 | 0 |
| Native American * | 1 | 1 | 0 |
| Country of Birth | |||
| USA | 43 | 37 | 43 |
| Caribbean | 1 | 3 | 0 |
| Central & South America | 0 | 3 | 1 |
| Mediterranean | 1 | 1 | 0 |
| Japan & Taiwan | 0 | 2 | 1 |
| Geographic Region of Enrollment | |||
| Eastern US | 10 | 11 | 10 |
| Central US | 16 | 16 | 8 |
| Western US | 19 | 18 | 27 |
| Smoking, Alcohol & Drug Use | |||
| % Current Smoker | 6.7 | 22.2 | 17.8 |
| % History of Smoking | 46.7 | 43.2 | 68.9 |
| Pack-years (mean, median) | 5.3, 0 | 9.6, 0 | 9.1, 2 |
| Drinks/week (mean, median) | 2.0, 0.1 | 1.6, 0 | 1.3, 0.1 |
| BMI (mean, median) | 30.7, 29.7 | 30.2, 28.3 | 32.3, 31.6 |
| History of Injection Drug Use | 0 | 0 | 8 |
| Log10 Proviral Load | |||
| Median (Range) | NA | −3.79 (−6.00, −1.80) | −4.96 (−6.00, −2.33) |
| Vibration Sensation | |||
| Impaired | 8 | 11 | 8 |
| Not Impaired | 36 | 31 | 32 |
| HAM/TSP | NA | 0 | 2 |
| Group | Variable | Beta (Unadjusted) | p-Value | Beta (Adjusted) | p-Value * |
|---|---|---|---|---|---|
| All subjects | Age | −0.005 | 0.003 | −0.004 | 0.007 |
| Smoking | −0.002 | 0.015 | −0.001 | 0.076 | |
| Sex (male) | 0.061 | 0.097 | 0.050 | 0.165 | |
| Controls | Age | −0.008 | 0.007 | −0.009 | 0.006 |
| HTLV-1 | Log10 proviral load | −0.032 | 0.069 | −0.023 | 0.167 |
| Smoking | −0.003 | 0.009 | −0.002 | 0.052 | |
| HTLV-2 | No significant associations | ||||
| Group | Variable | Odds Ratio | 95% CI | Odds Ratio * | 95% CI * |
|---|---|---|---|---|---|
| All subjects | Age | 1.062 | (1.008, 1.119) | 1.046 | (0.989, 1.106) |
| Smoking | 1.033 | (1.004, 1.063) | 1.026 | (0.997, 1.057) | |
| Loge T/S ratio | 0.714 | (0.534, 0.956) | 0.826 | (0.610, 1.118) | |
| Controls | Age | 1.130 | (1.009, 1.258) | 1.129 | (0.988, 1.292) |
| HTLV-1 | No significant associations | ||||
| HTLV-2 | Loge T/S ratio | 0.380 | (0.176, 0.820) | 0.400 | (0.178, 0.897) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usadi, B.; Bruhn, R.; Lin, J.; Lee, T.-H.; Blackburn, E.; Murphy, E.L. Telomere Length, Proviral Load and Neurologic Impairment in HTLV-1 and HTLV-2-Infected Subjects. Viruses 2016, 8, 221. https://0-doi-org.brum.beds.ac.uk/10.3390/v8080221
Usadi B, Bruhn R, Lin J, Lee T-H, Blackburn E, Murphy EL. Telomere Length, Proviral Load and Neurologic Impairment in HTLV-1 and HTLV-2-Infected Subjects. Viruses. 2016; 8(8):221. https://0-doi-org.brum.beds.ac.uk/10.3390/v8080221
Chicago/Turabian StyleUsadi, Benjamin, Roberta Bruhn, Jue Lin, Tzong-Hae Lee, Elizabeth Blackburn, and Edward L. Murphy. 2016. "Telomere Length, Proviral Load and Neurologic Impairment in HTLV-1 and HTLV-2-Infected Subjects" Viruses 8, no. 8: 221. https://0-doi-org.brum.beds.ac.uk/10.3390/v8080221