
Anti-SARS-CoV-2 IgG against the S Protein: A Comparison of BNT162b2, mRNA-1273, ChAdOx1 nCoV-2019 and Ad26.COV2.S Vaccines

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
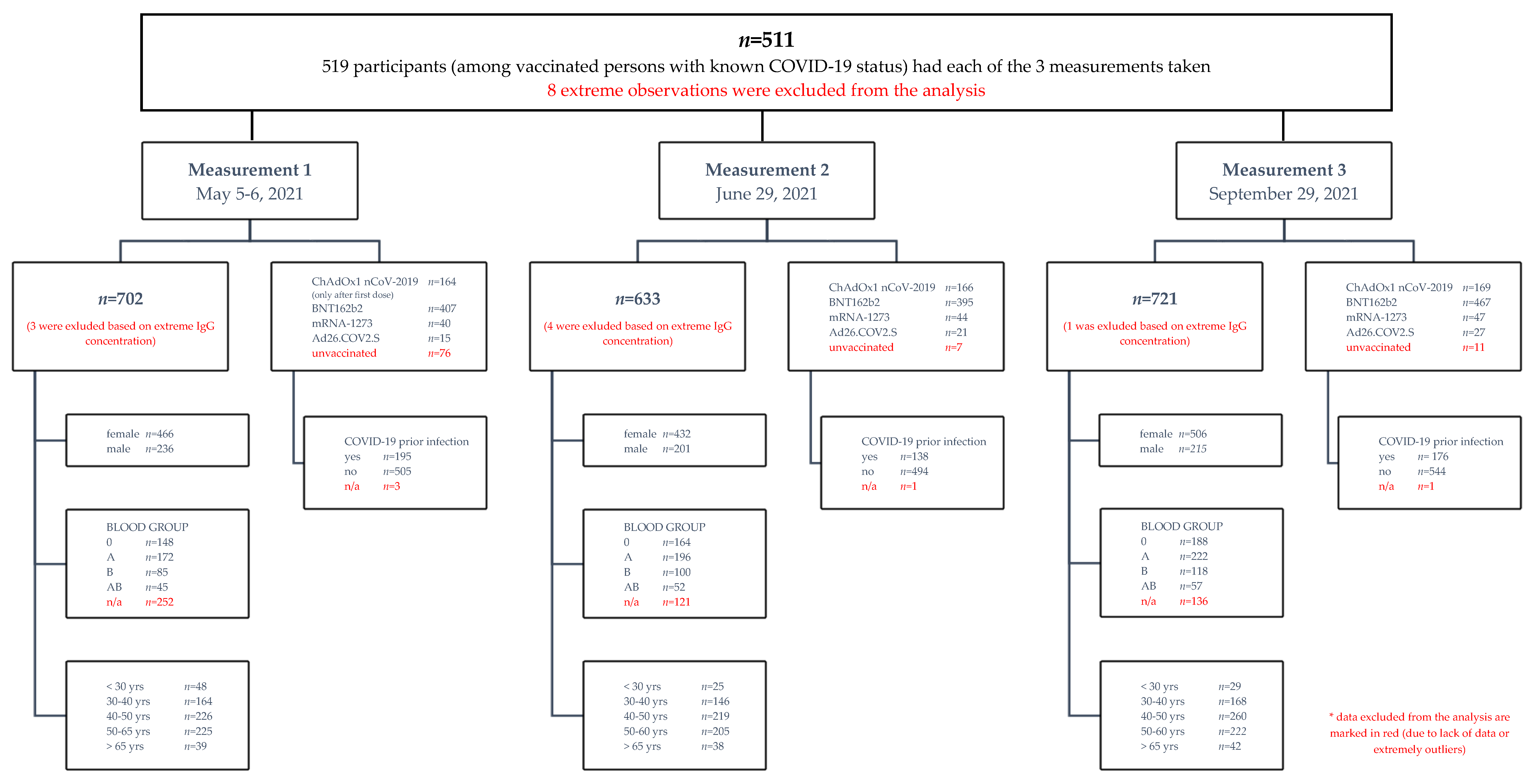
2.1. Cohort
2.2. Aim of Study
2.3. Enzyme-Linked Immunosorbent Assay for Detection of IgG Antibodies
2.4. Statistical Analysis
3. Results
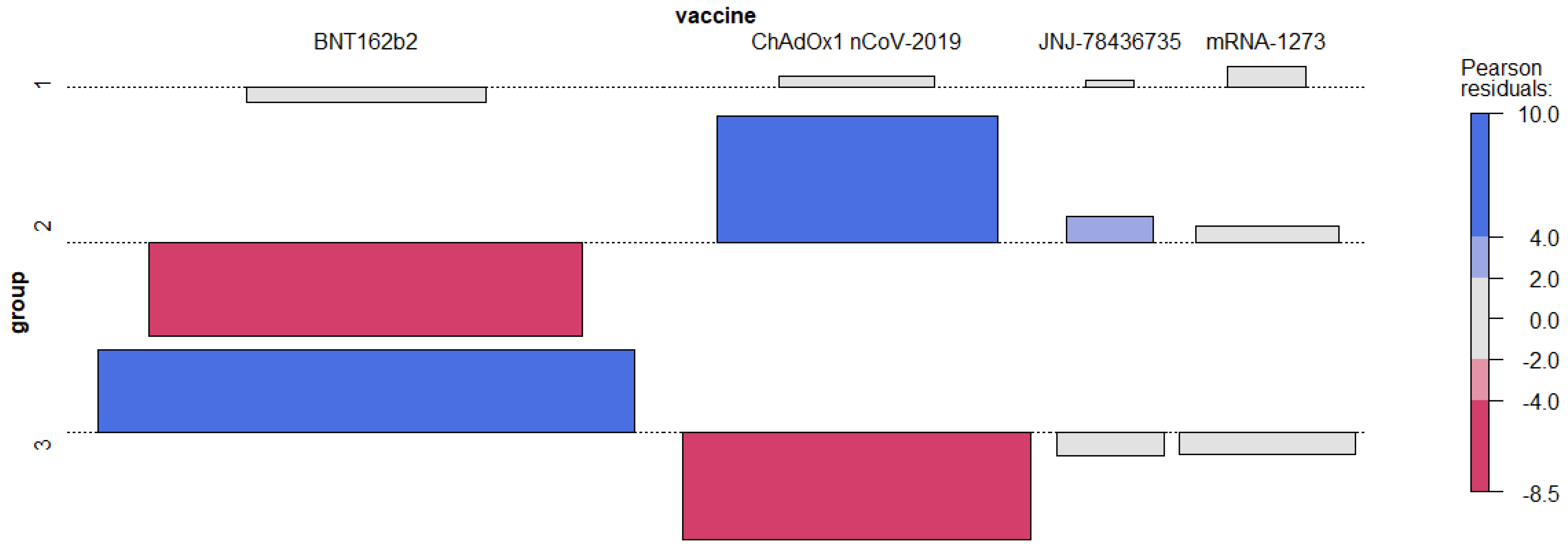
3.1. The Influence of Factors on the Level of Anti-SARS-CoV-2 IgG
- Group 1: up to 14 days between the planned date of the second measurement and the last dose of vaccine;
- Group 2: between 14 and 60 days; and
- Group 3: more than 60 days.
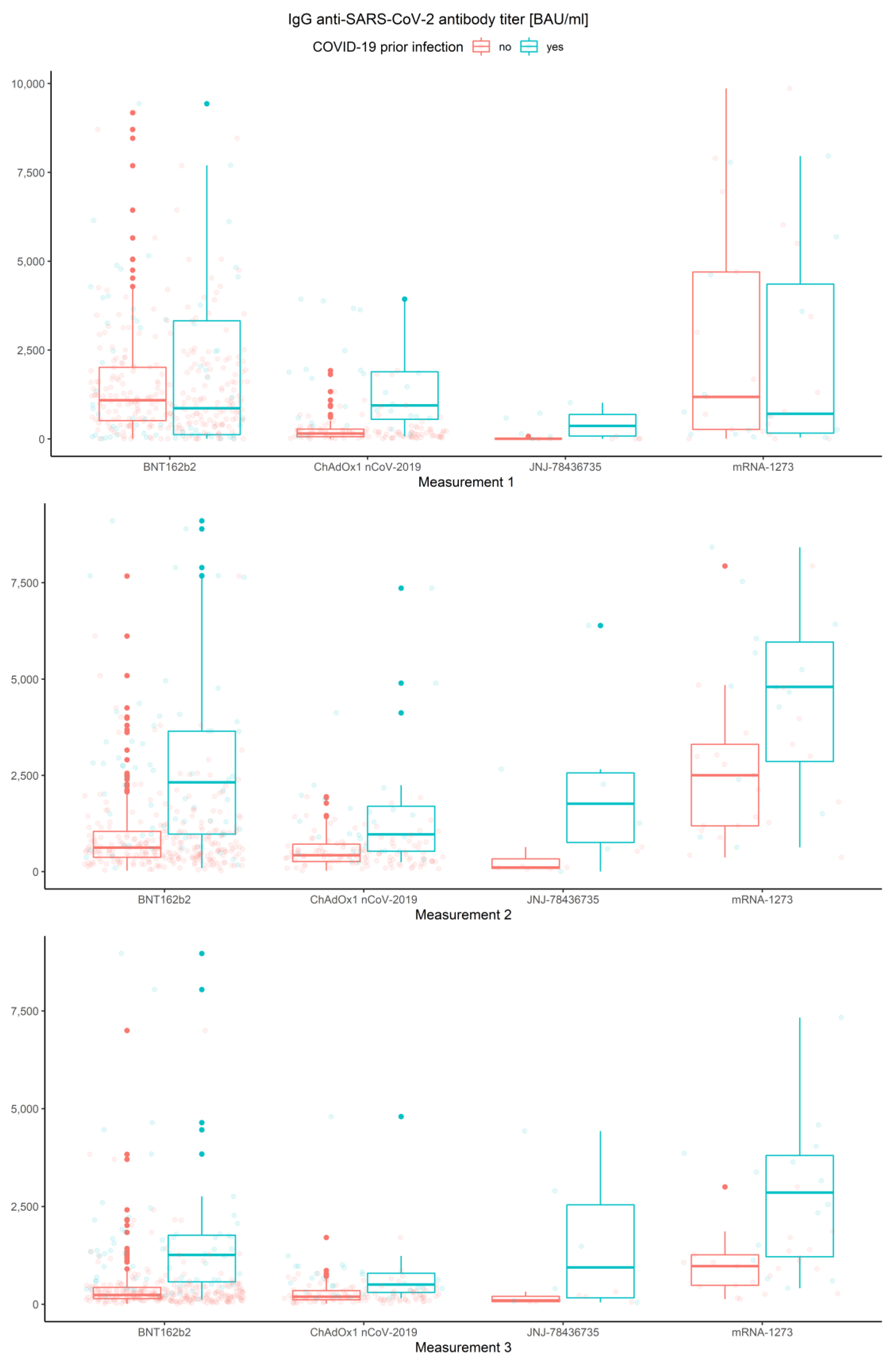
3.2. Measurements, Prior COVID-19 Infection and Vaccine Type
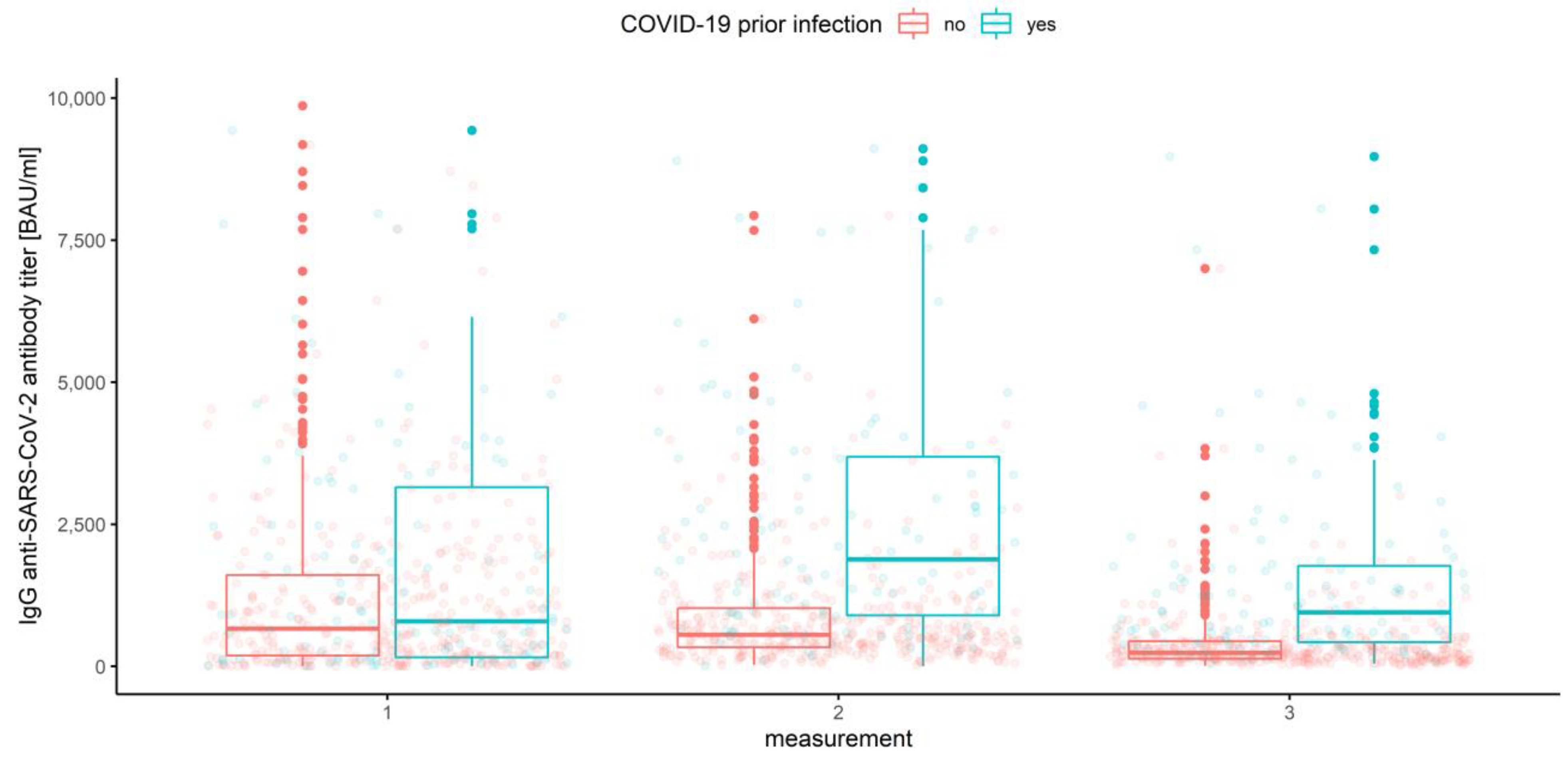
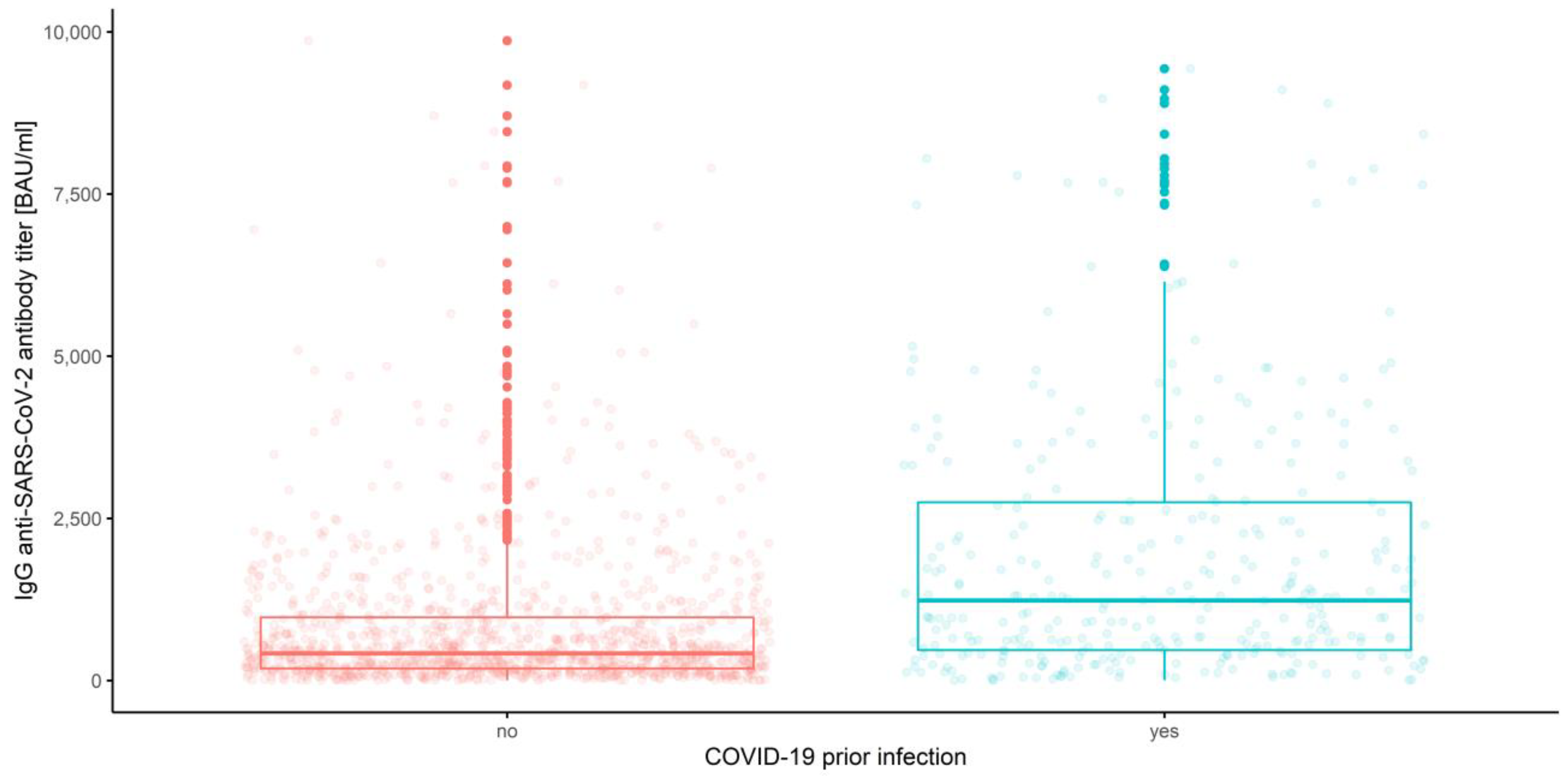
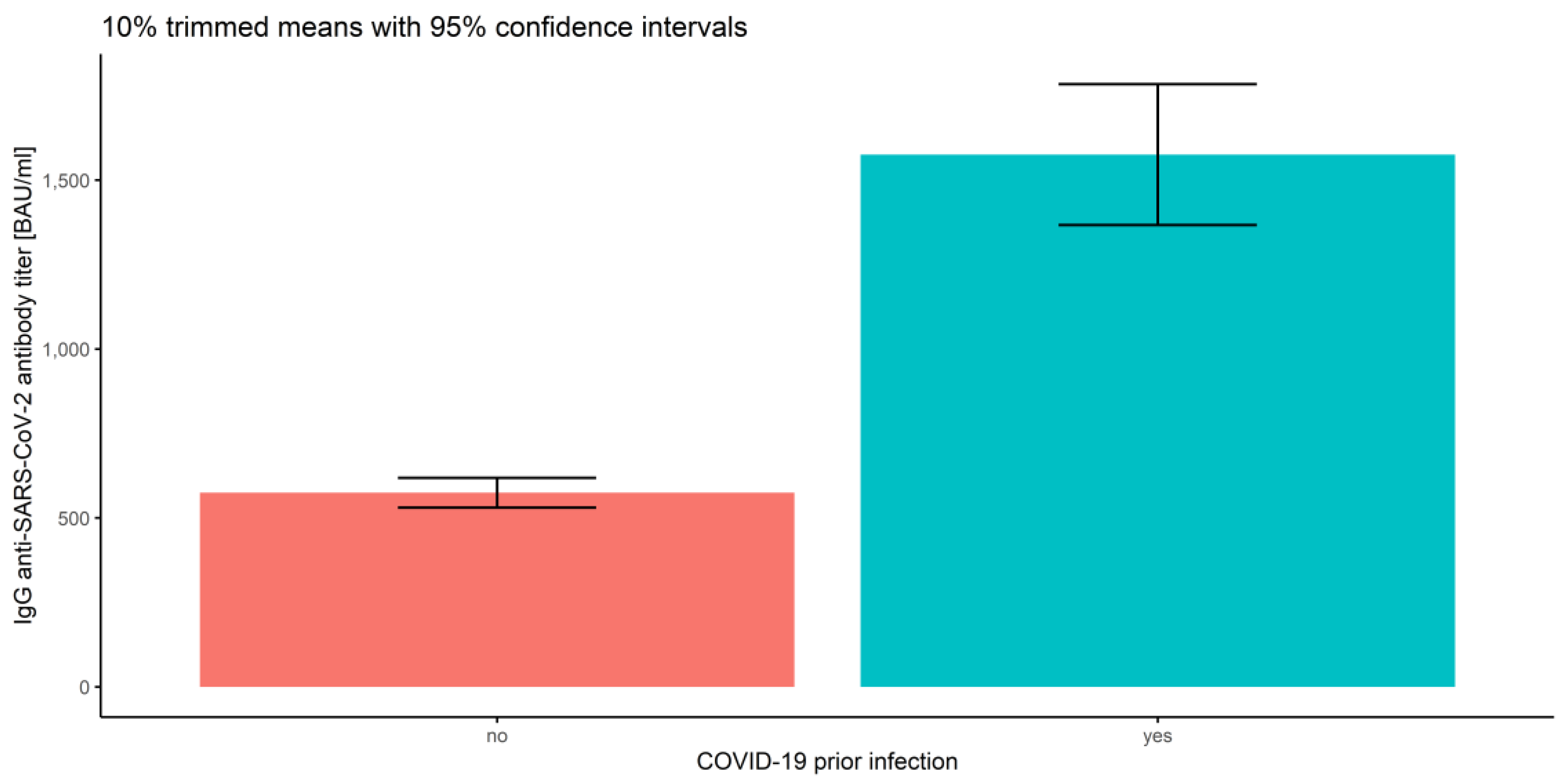
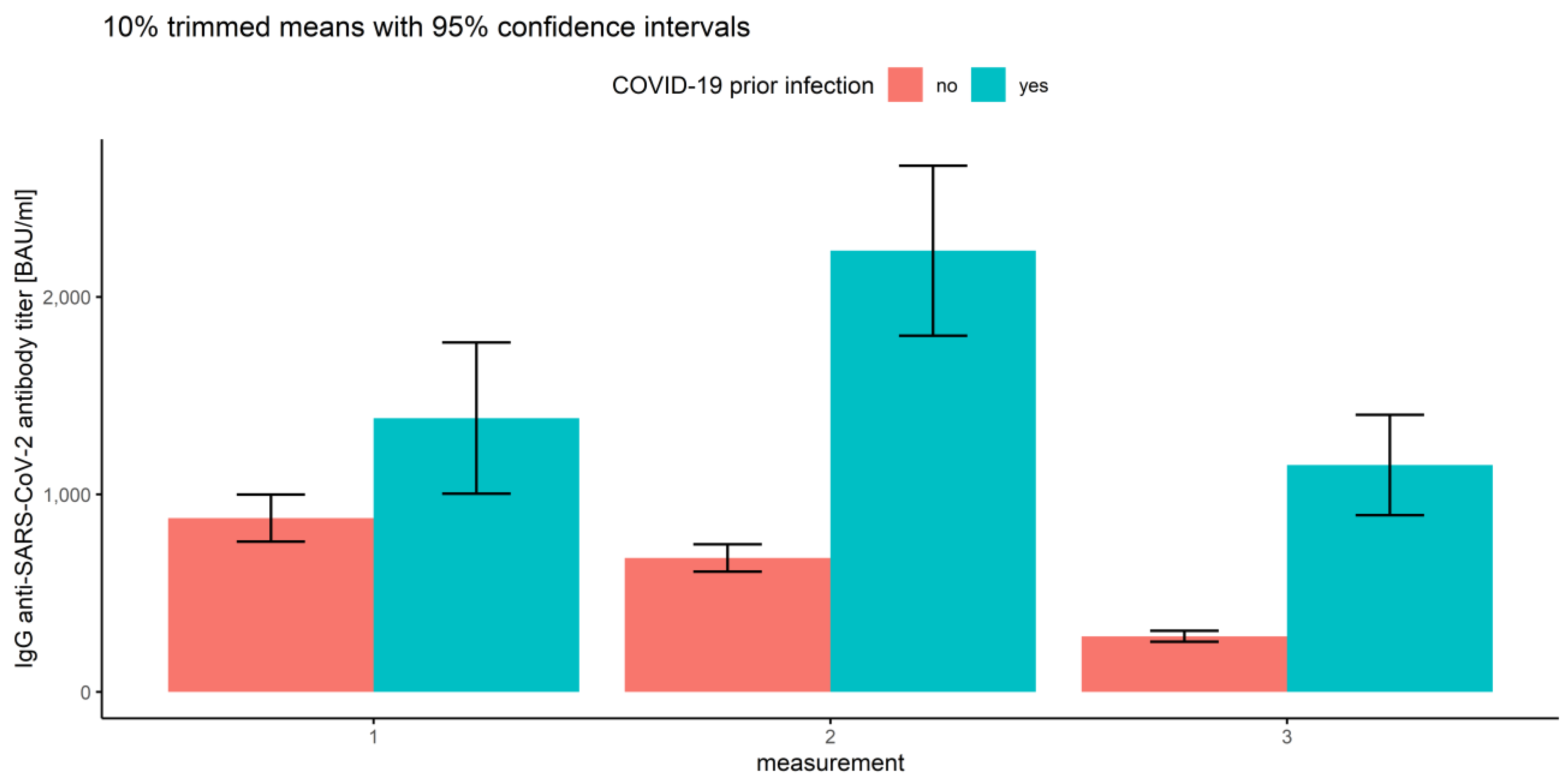
3.2.1. Measurements and Prior COVID-19 Infection
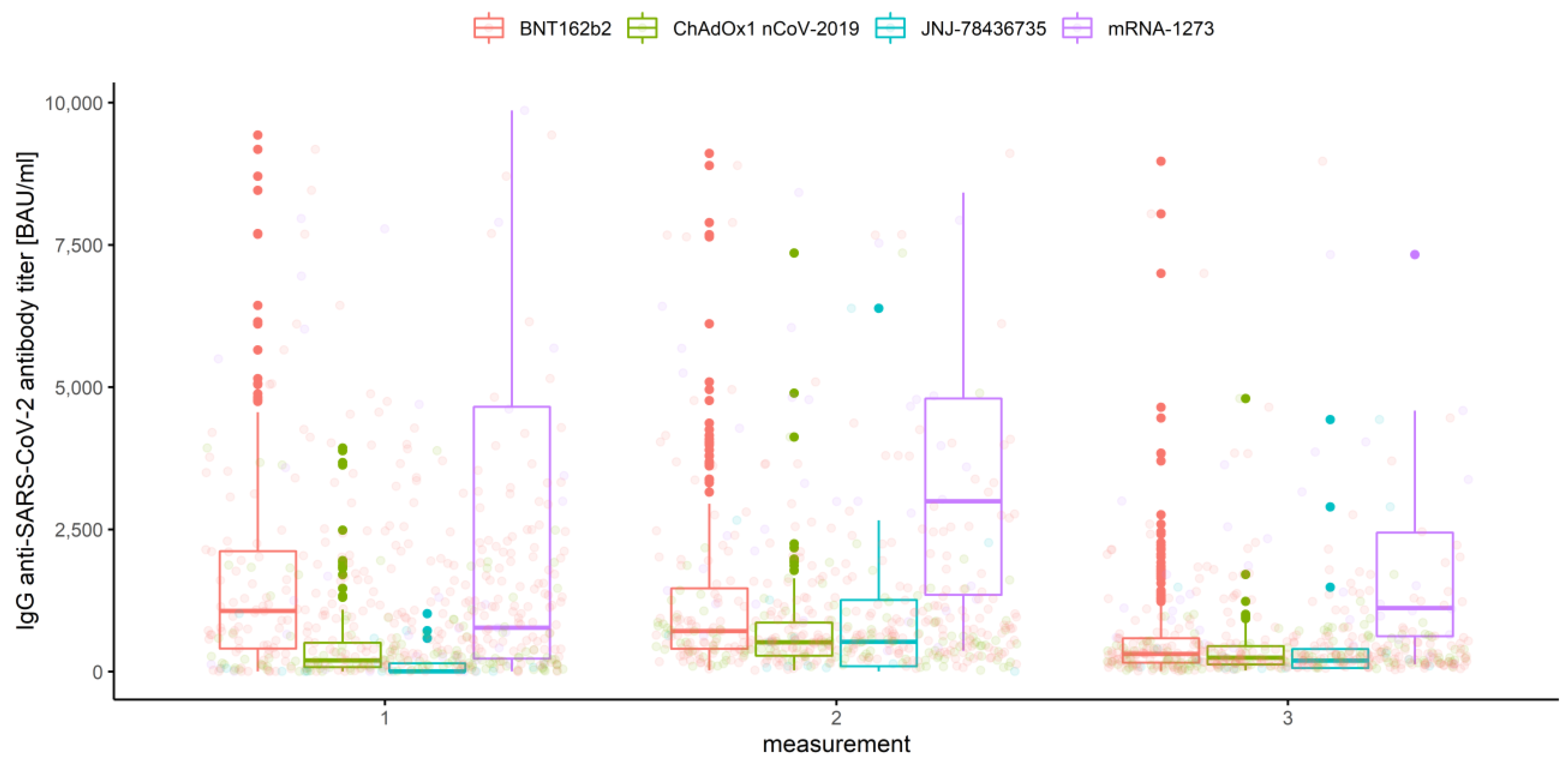
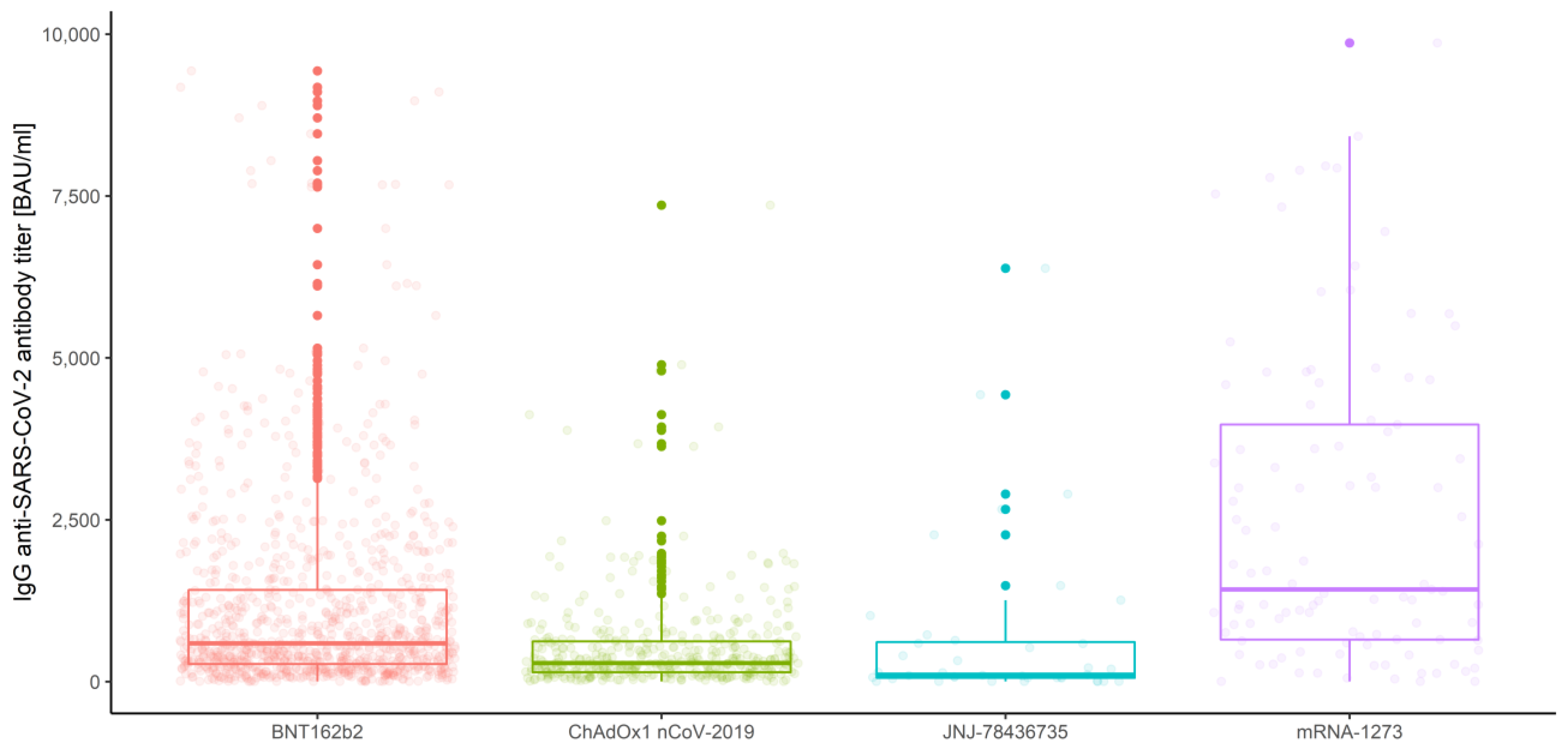
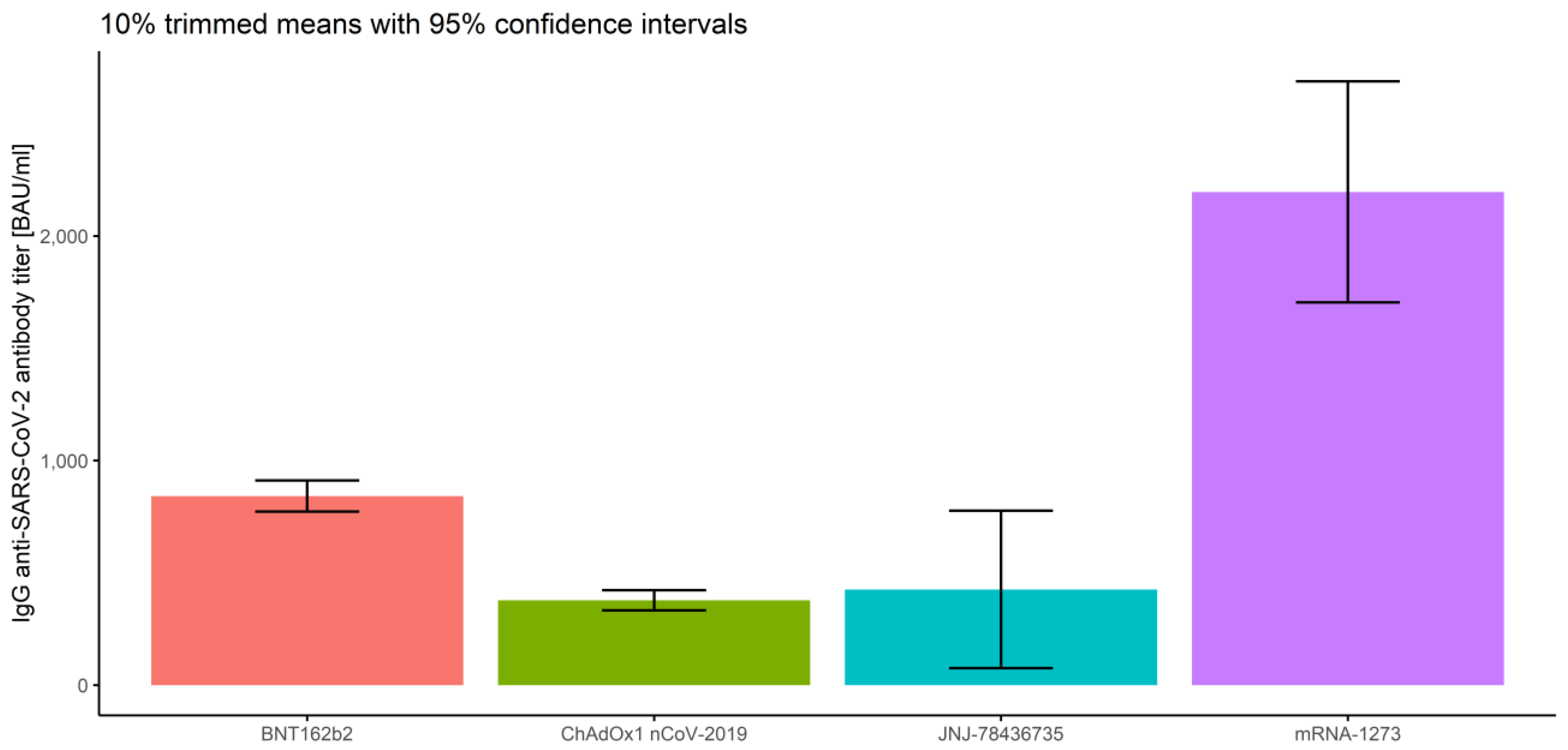
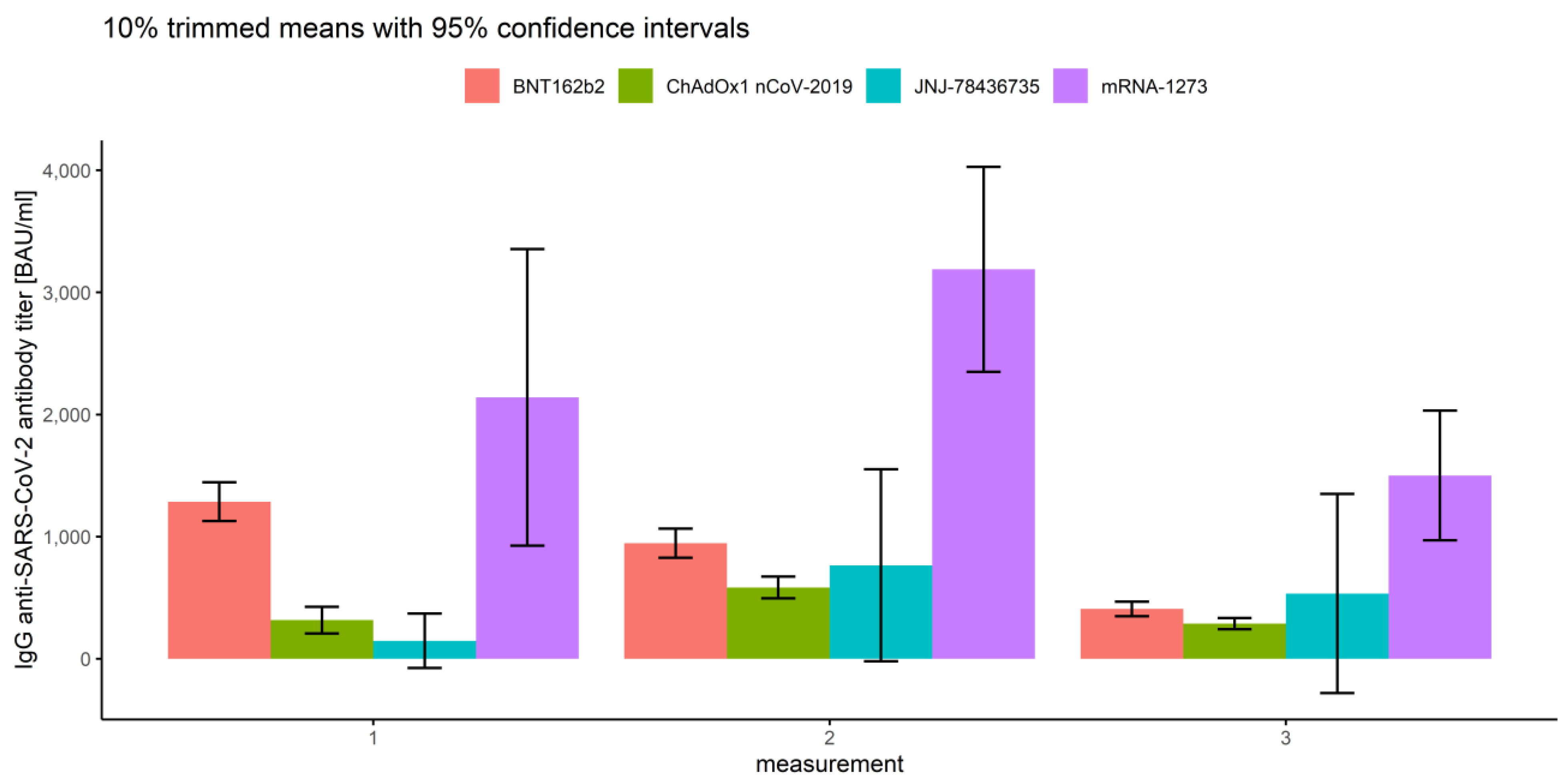
3.2.2. Association of Measurements and Vaccine Type
- In the first measurement, there are no significant differences between BNT162b2 and mRNA-1273 (p = 0.263), and between ChAdOx1 nCoV-2019 and JNJ-78436735 (p = 0.076).
- In the second measurement, there are no significant differences between BNT162b2 and JNJ-78436735 (p = 0.996), and between ChAdOx1 nCoV-2019 and JNJ-78436735 (p = 0.856).
- In the third measurement, there are no significant differences between BNT162b2 and JNJ-78436735 (p = 0.966), between ChAdOx1 nCoV-2019 and JNJ-78436735 (p = 0.665) and between JNJ-78436735 and mRNA-1273 (p = 0.220).
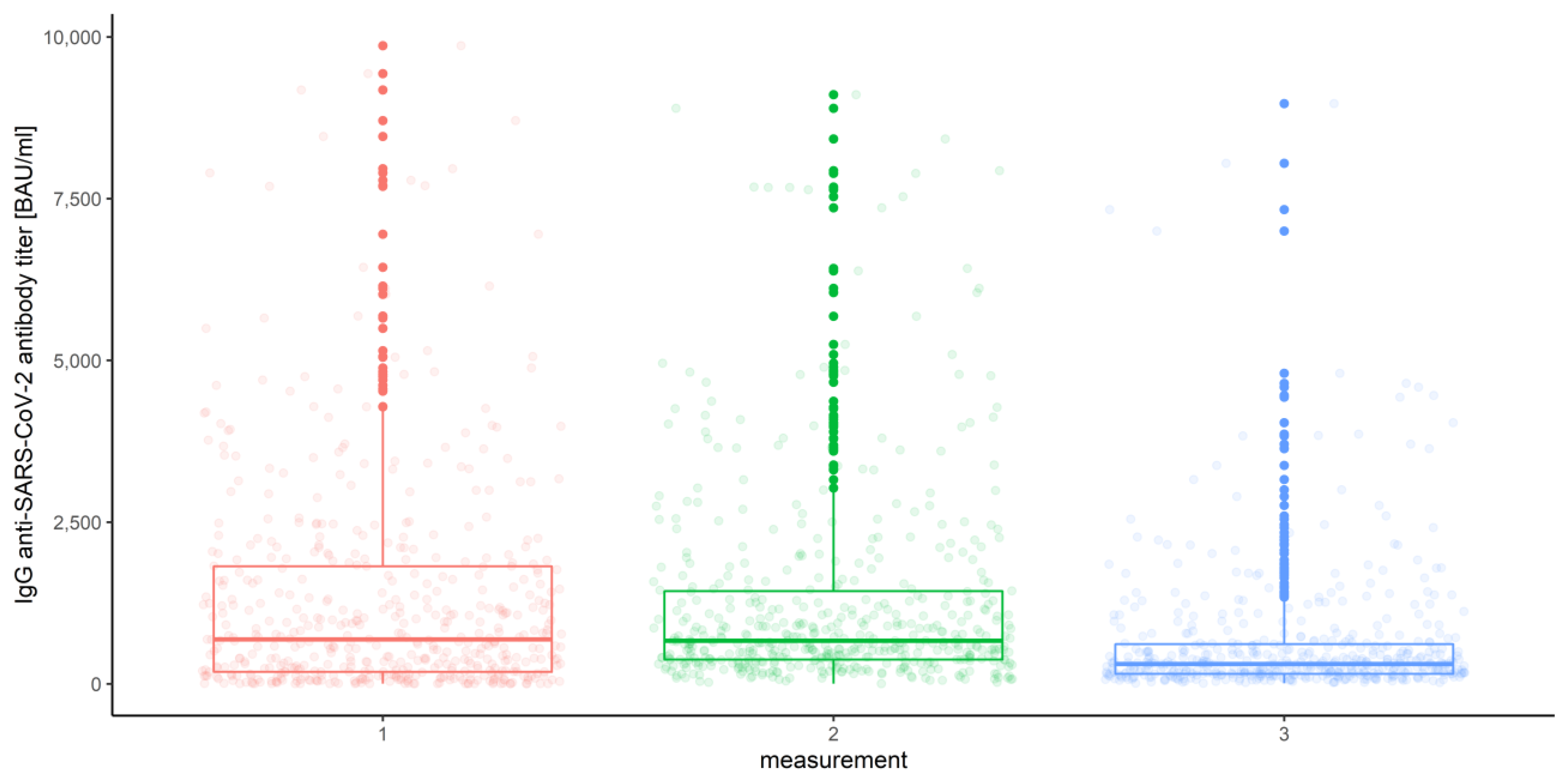
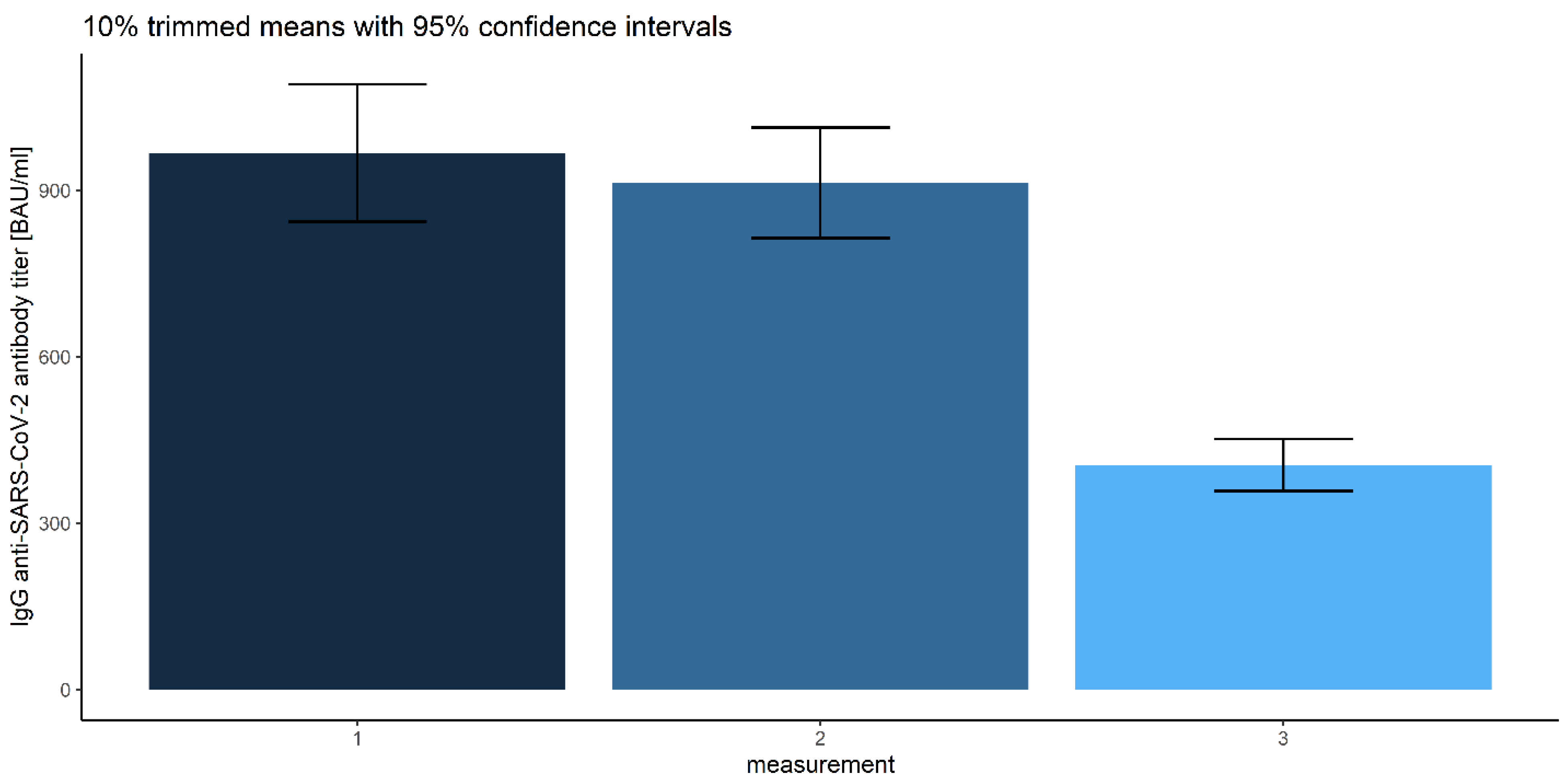
3.2.3. Measurements
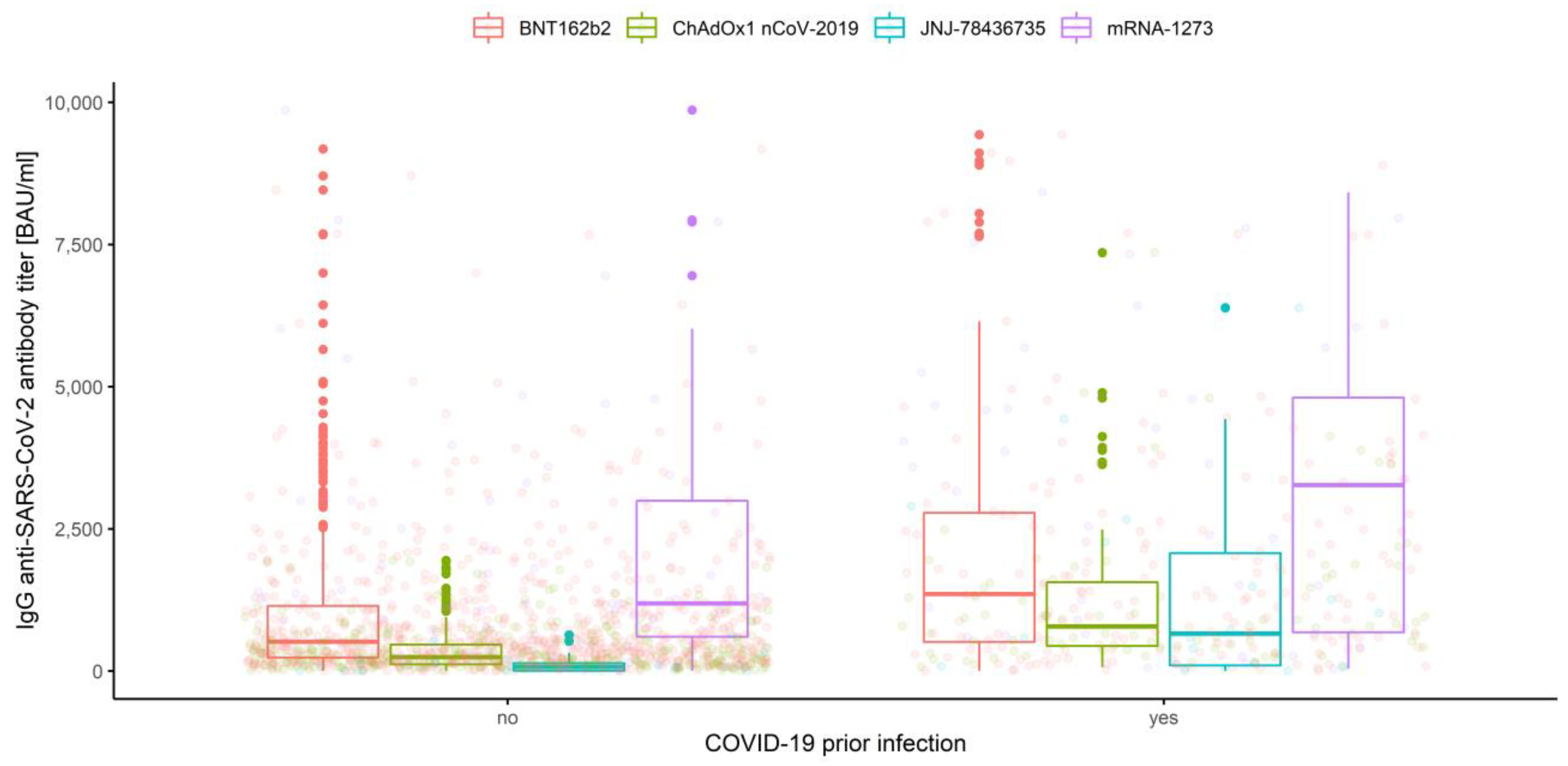
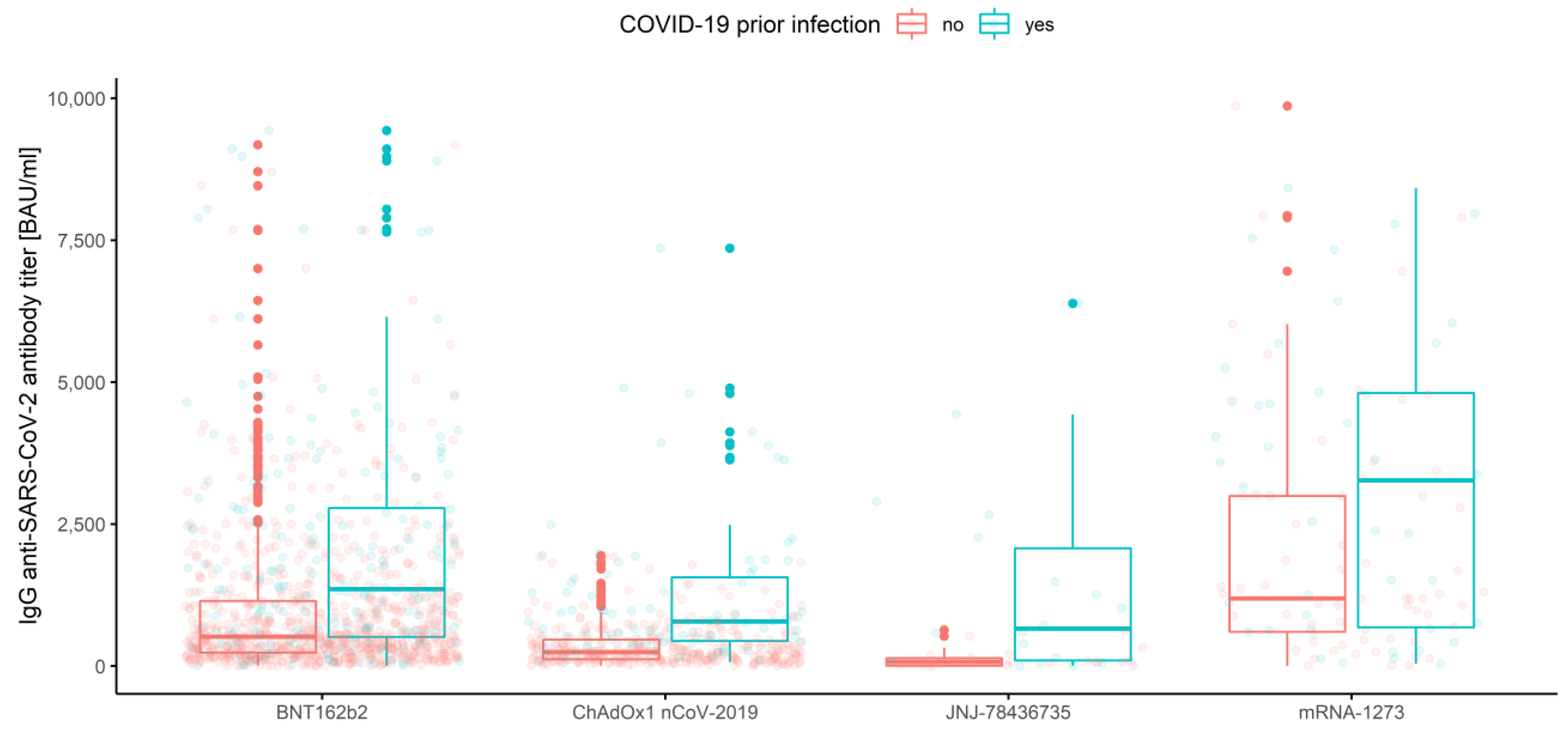
3.2.4. Association of Prior COVID-19 Infection and Vaccine Type
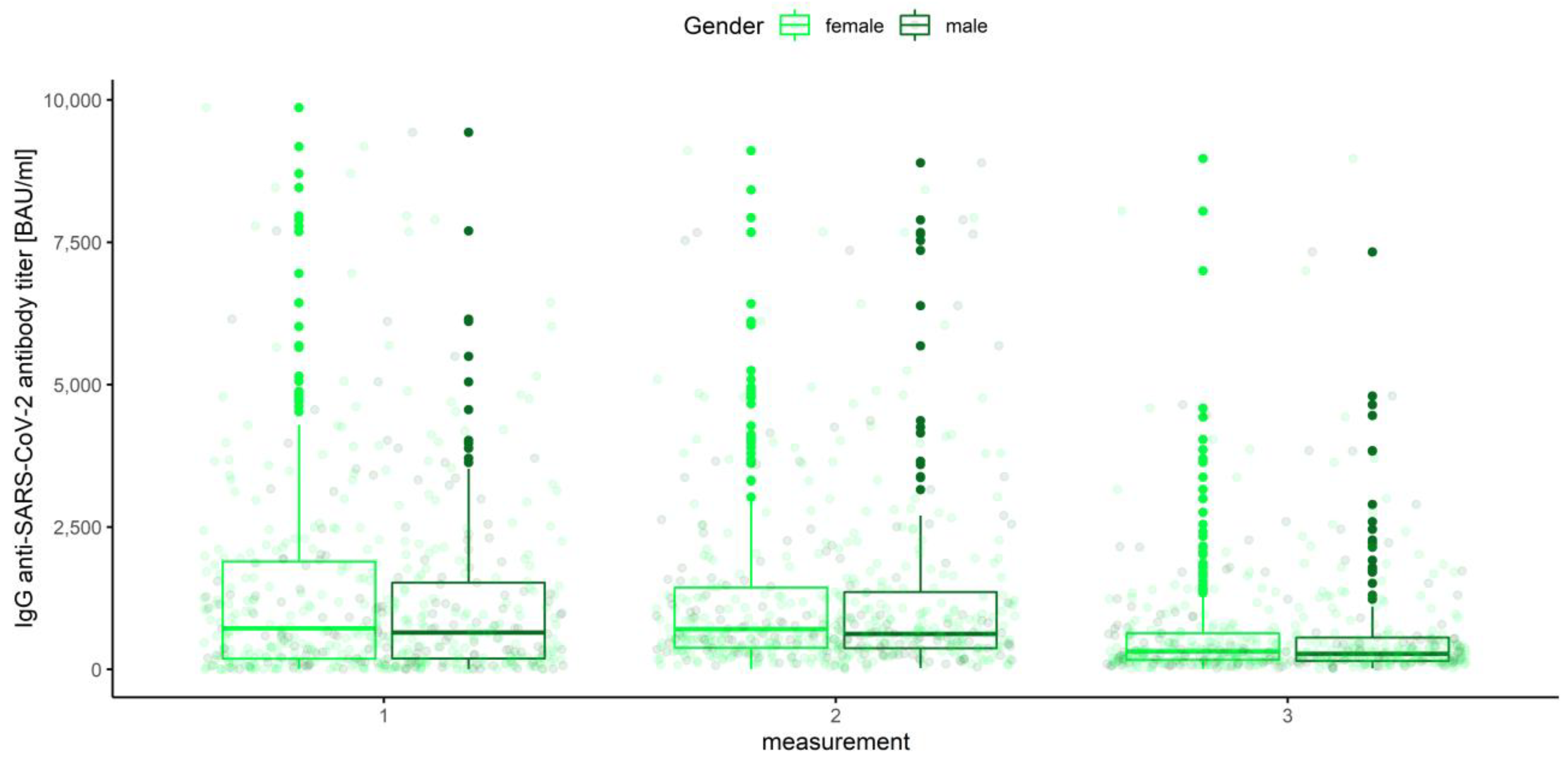
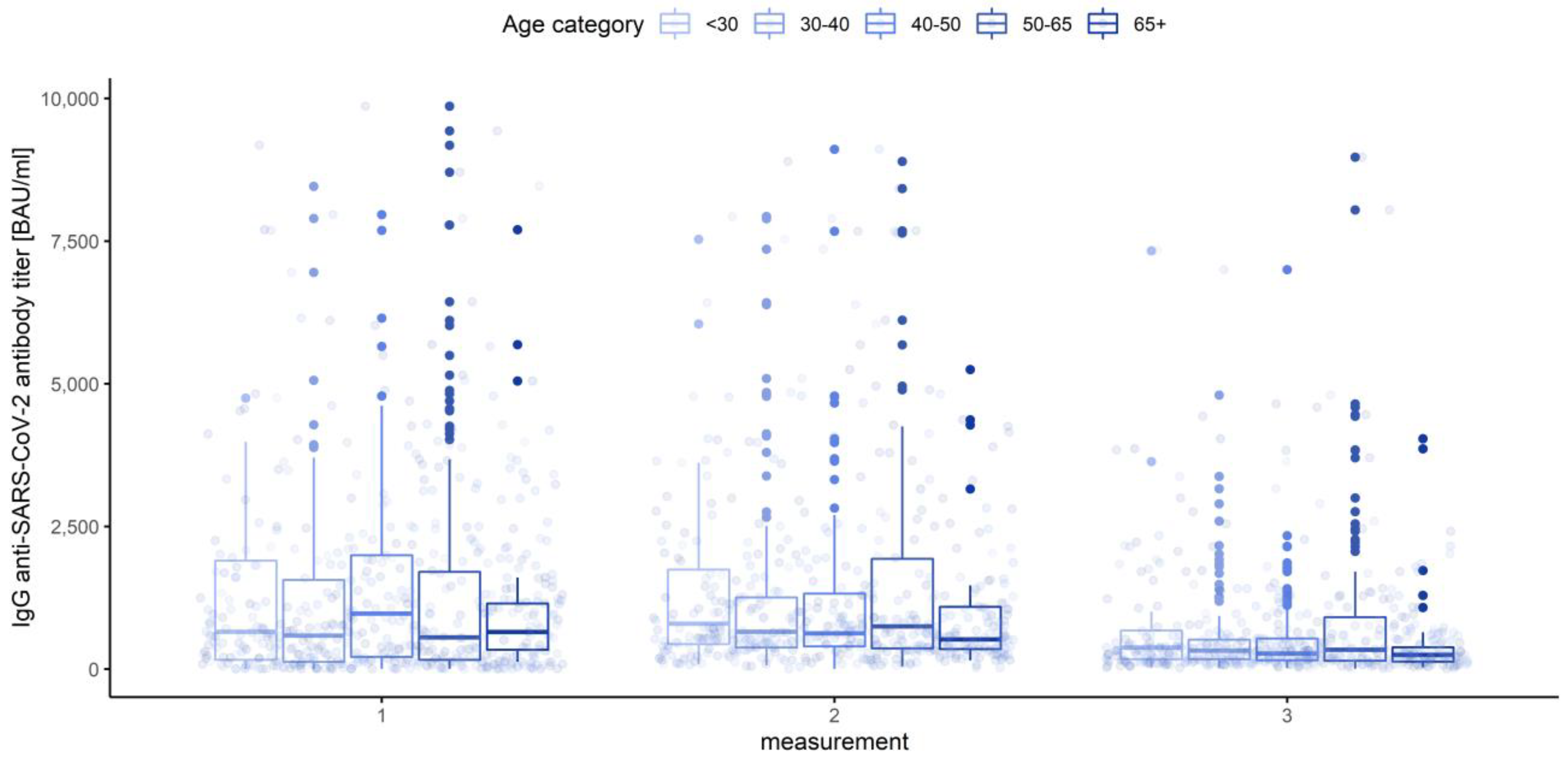
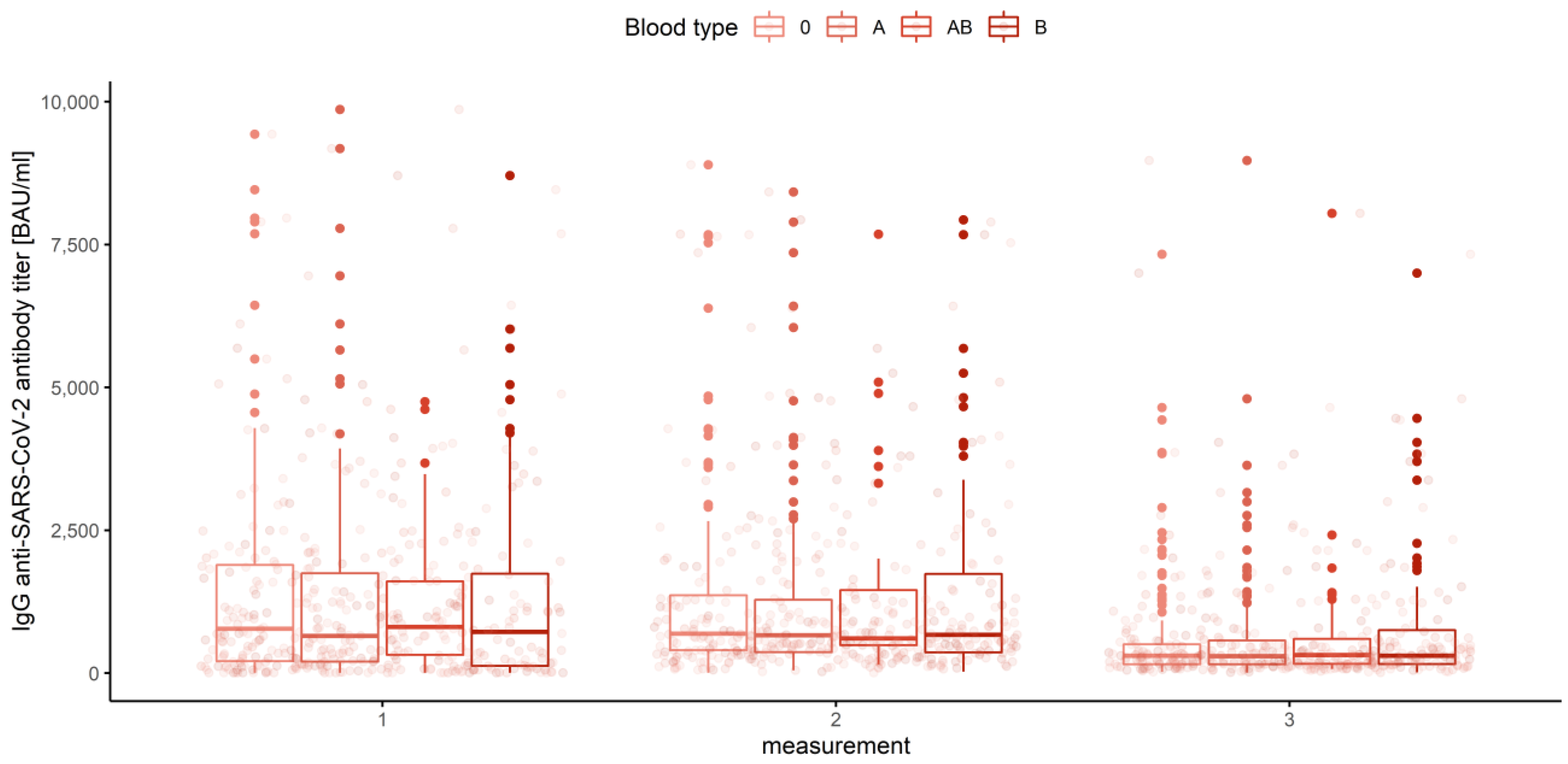
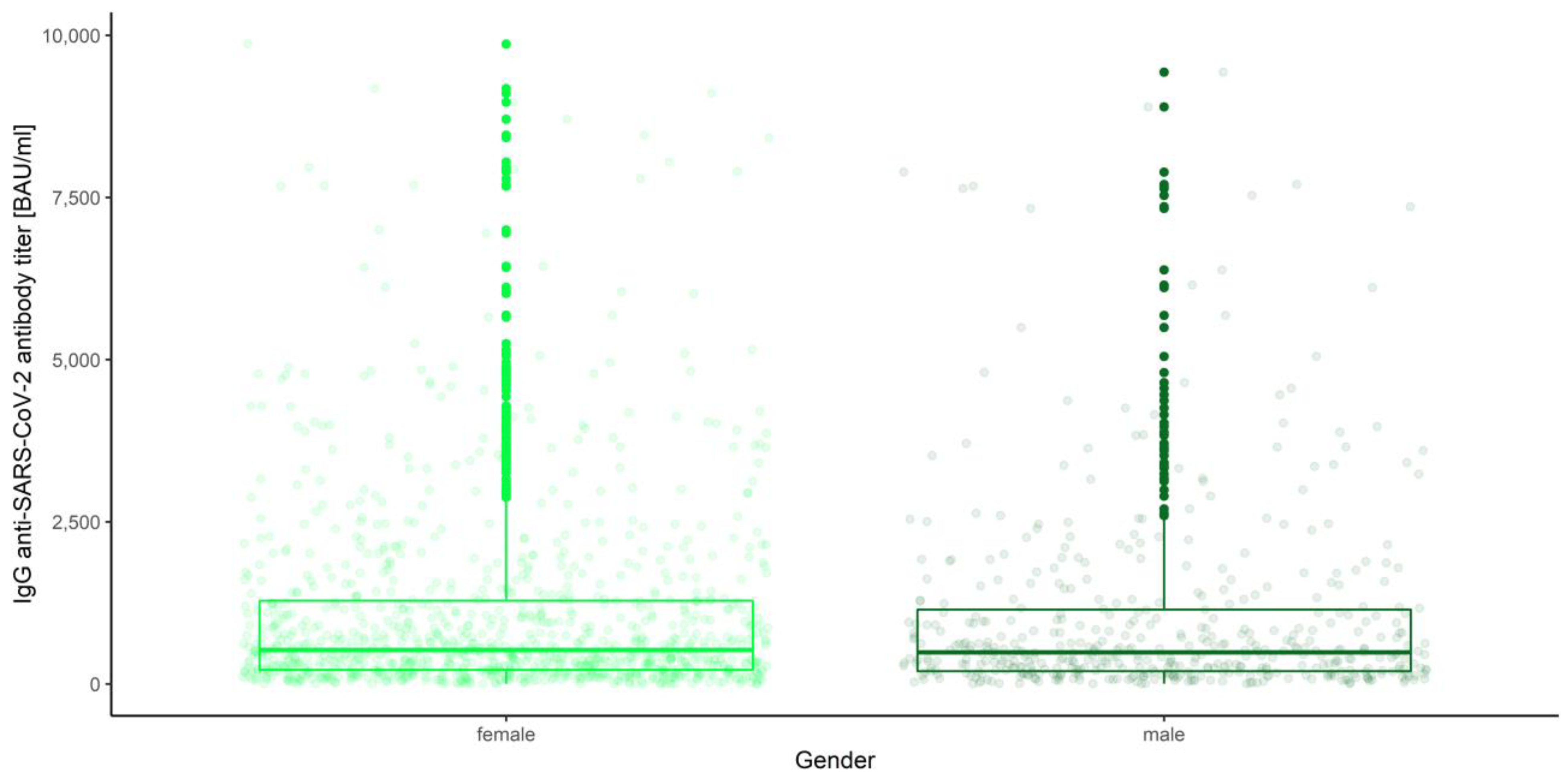
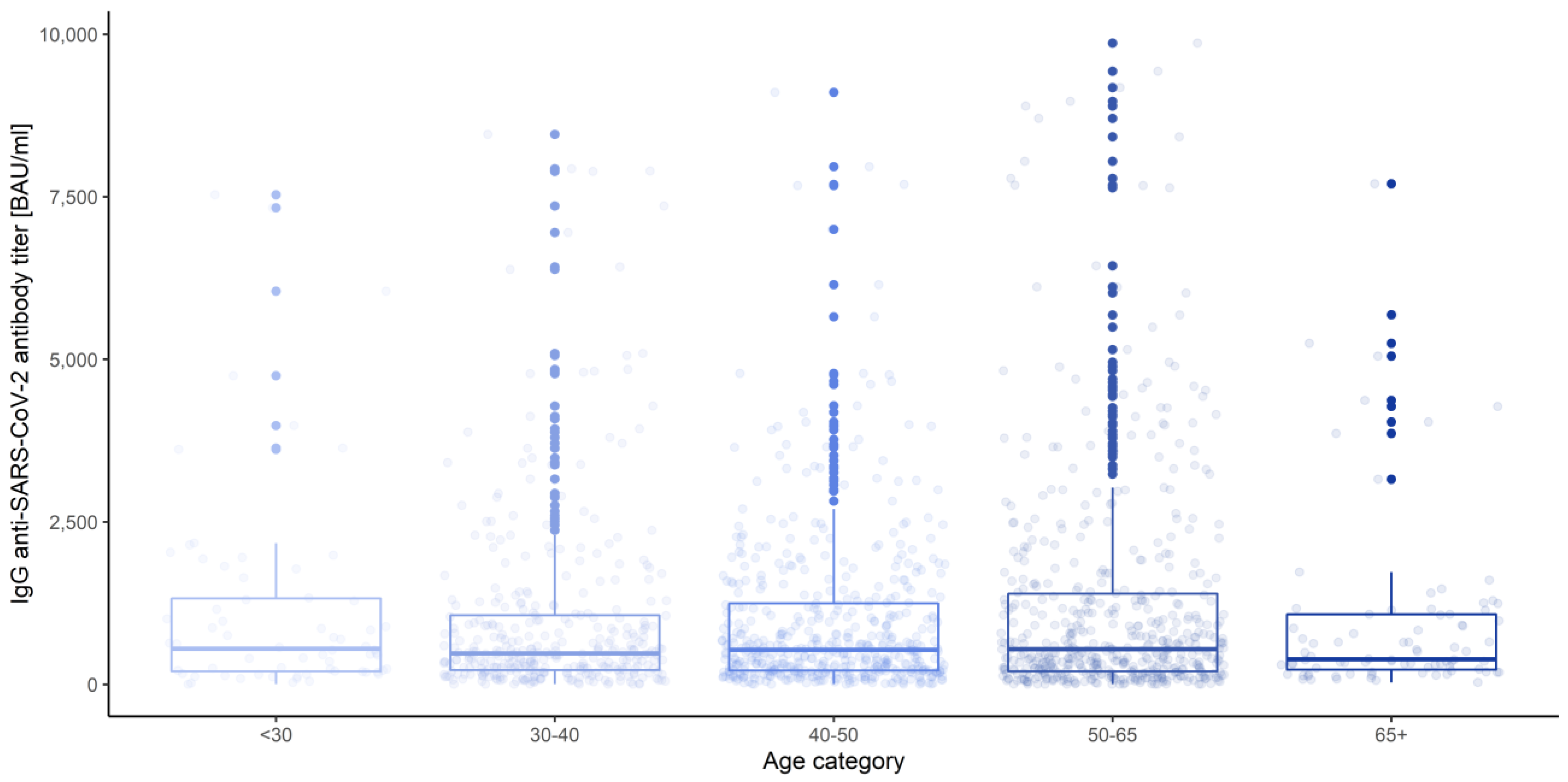
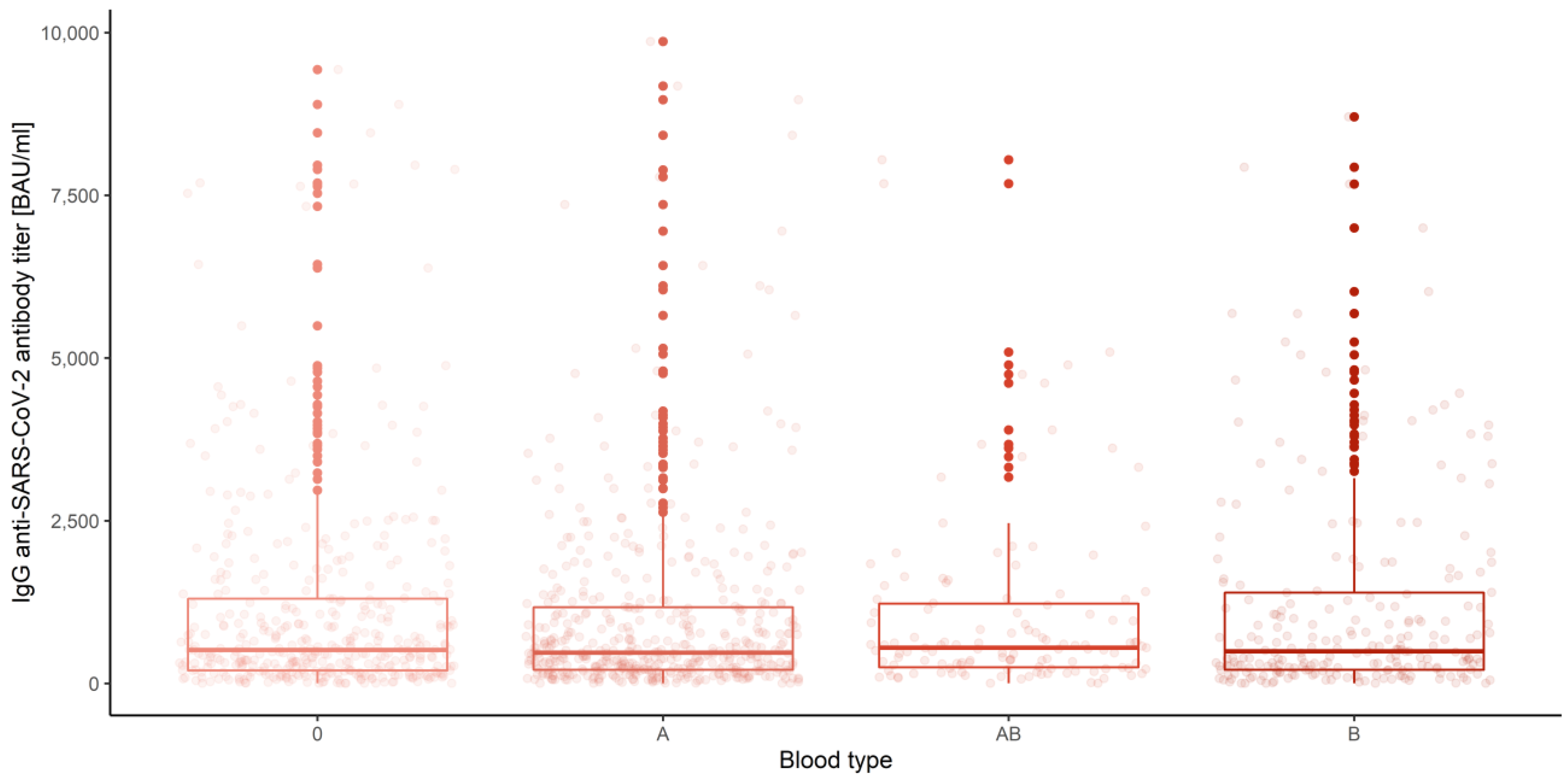
3.3. Gender, Age Category and Blood Group
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zollner, A.; Watschinger, C.; Rossler, A.; Farcet, M.R.; Penner, A.; Bohm, V.; Kiechl, S.J.; Stampfel, G.; Hintenberger, R.; Tilg, H.; et al. B and T cell response to SARS-CoV-2 vaccination in health care professionals with and without previous COVID-19. EBioMedicine 2021, 70, 103539. [Google Scholar] [CrossRef]
- COVID-19 Vaccination Report. Available online: https://www.gov.pl/web/szczepimysie/raport-szczepien-przeciwko-covid-19 (accessed on 11 December 2021).
- Turner, J.S.; O’Halloran, J.A.; Kalaidina, E.; Kim, W.; Schmitz, A.J.; Zhou, J.Q.; Lei, T.; Thapa, M.; Chen, R.E.; Case, J.B.; et al. SARS-CoV-2 mRNA vaccines induce persistent human germinal centre responses. Nature 2021, 596, 109–113. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Teijaro, J.R.; Farber, D.L. COVID-19 vaccines: Modes of immune activation and future challenges. Nat. Rev. Immunol. 2021, 21, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Lange, A.; Borowik, A.; Bochenska, J.; Rossowska, J.; Jaskula, E. Immune Response to COVID-19 mRNA Vaccine-A Pilot Study. Vaccines 2021, 9, 488. [Google Scholar] [CrossRef] [PubMed]
- Bongiovanni, M.; Liuzzi, G.; Schiavon, L.; Gianturco, L.; Giuliani, G. Evaluation of the immune response to COVID-19 vaccine mRNA BNT162b2 and correlation with previous COVID-19 infection. J. Clin. Virol. 2021, 143, 104962. [Google Scholar] [CrossRef] [PubMed]
- Lustig, Y.; Sapir, E.; Regev-Yochay, G.; Cohen, C.; Fluss, R.; Olmer, L.; Indenbaum, V.; Mandelboim, M.; Doolman, R.; Amit, S.; et al. BNT162b2 COVID-19 vaccine and correlates of humoral immune responses and dynamics: A prospective, single-centre, longitudinal cohort study in health-care workers. Lancet Respir. Med. 2021, 9, 999–1009. [Google Scholar] [CrossRef]
- Shroff, R.T.; Chalasani, P.; Wei, R.; Pennington, D.; Quirk, G.; Schoenle, M.V.; Peyton, K.L.; Uhrlaub, J.L.; Ripperger, T.J.; Jergovic, M.; et al. Immune responses to two and three doses of the BNT162b2 mRNA vaccine in adults with solid tumors. Nat. Med. 2021, 27, 2002–2011. [Google Scholar] [CrossRef]
- Available online: www.pfizerpro.com.pl (accessed on 18 October 2021).
- Oliver, S.E.; Gargano, J.W.; Marin, M.; Wallace, M.; Curran, K.G.; Chamberland, M.; McClung, N.; Campos-Outcalt, D.; Morgan, R.L.; Mbaeyi, S.; et al. The Advisory Committee on Immunization Practices’ Interim Recommendation for Use of Moderna COVID-19 Vaccine-United States, December 2020. MMWR Morb. Mortal. Wkly. Rep. 2021, 69, 1653–1656. [Google Scholar] [CrossRef]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef]
- Angeli, F.; Spanevello, A.; Reboldi, G.; Visca, D.; Verdecchia, P. SARS-CoV-2 vaccines: Lights and shadows. Eur. J. Intern. Med. 2021, 88, 1–8. [Google Scholar] [CrossRef]
- Summary of Product Characteristics Vaxzevria, COVID 19 Vaccine (ChAdOx1 S [Recombinant]). Available online: https://ec.europa.eu/health/documents/community-register/2021/20210408151651/anx_151651_pl.pdf (accessed on 19 October 2021).
- Abbasi, J. COVID-19 and mRNA Vaccines-First Large Test for a New Approach. JAMA 2020, 324, 1125–1127. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. The race for coronavirus vaccines: A graphical guide. Nature 2020, 580, 576–577. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency (EMA). Vaxzevria: COVID-19 Vaccine Safety Update. Available online: https://www.ema.europa.eu/ (accessed on 20 October 2021).
- Janssen Vaccines & Prevention. A Randomized, Double-Blind, Placebo-Controlled Phase 3 Study to Assess the Efficacy and Safety of Ad26.COV2.S for the Prevention of SARS-CoV-2-Mediated COVID-19 in Adults Aged 18 Years and Older ENSEMBLE Protocol VAC31518COV3001; Phase 3” (PDF). Available online: https://www.jnj.com/coronavirus/ensemble-1-study-protocol (accessed on 27 October 2021).
- U.S. Food and Drug Administration (FDA). FDA Briefing Document Janssen Ad26.COV2.S Vaccine for the Prevention of COVID-19 (Report). Available online: https://www.fda.gov/media/146217/download (accessed on 27 October 2021).
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against COVID-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Polinski, J.M.; Weckstein, A.R.; Batech, M.; Kabelac, C.; Kamath, T.; Harvey, R.; Jain, S.; Rassen, J.A.; Khan, N.; Schneeweiss, S. Effectiveness of the Single-Dose Ad26.COV2.S COVID Vaccine. MedRxiv 2021. [Google Scholar] [CrossRef]
- Hossain, M.K.; Hassanzadeganroudsari, M.; Apostolopoulos, V. The emergence of new strains of SARS-CoV-2. What does it mean for COVID-19 vaccines? Expert Rev. Vaccines 2021, 20, 635–638. [Google Scholar] [CrossRef]
- Harvala, H.; Robb, M.L.; Watkins, N.; Ijaz, S.; Dicks, S.; Patel, M.; Supasa, P.; Wanwisa, D.; Liu, C.; Mongkolsapaya, J.; et al. Convalescent plasma therapy for the treatment of patients with COVID-19: Assessment of methods available for antibody detection and their correlation with neutralising antibody levels. Transfus. Med. 2021, 31, 167–175. [Google Scholar] [CrossRef]
- Okba, N.M.A.; Muller, M.A.; Li, W.; Wang, C.; GeurtsvanKessel, C.H.; Corman, V.M.; Lamers, M.M.; Sikkema, R.S.; de Bruin, E.; Chandler, F.D.; et al. Severe Acute Respiratory Syndrome Coronavirus 2-Specific Antibody Responses in Coronavirus Disease Patients. Emerg. Infect. Dis. 2020, 26, 1478–1488. [Google Scholar] [CrossRef]
- Dolscheid-Pommerich, R.; Bartok, E.; Renn, M.; Kummerer, B.M.; Schulte, B.; Schmithausen, R.M.; Stoffel-Wagner, B.; Streeck, H.; Saschenbrecker, S.; Steinhagen, K.; et al. Correlation between a quantitative anti-SARS-CoV-2 IgG ELISA and neutralization activity. J. Med. Virol. 2022, 94, 388–392. [Google Scholar] [CrossRef]
- Tretyn, A.; Szczepanek, J.; Skorupa, M.; Jarkiewicz-Tretyn, J.; Sandomierz, D.; Dejewska, J.; Ciechanowska, K.; Jarkiewicz-Tretyn, A.; Koper, W.; Palgan, K. Differences in the Concentration of Anti-SARS-CoV-2 IgG Antibodies Post-COVID-19 Recovery or Post-Vaccination. Cells 2021, 10, 1952. [Google Scholar] [CrossRef]
- Wilcox, R. Introduction to Robust Estimation and Hypothesis Testing, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, S.S.; Prazeres, D.M.F.; Azevedo, A.M.; Marques, M.P.C. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021, 39, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Schlake, T.; Thess, A.; Thran, M.; Jordan, I. mRNA as novel technology for passive immunotherapy. Cell. Mol. Life Sci. 2019, 76, 301–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, B.; Lee, K. Stability of the osmoregulated promoter-derived proP mRNA is posttranscriptionally regulated by RNase III in Escherichia coli. J. Bacteriol. 2015, 197, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Weissman, D. Nucleoside Modified mRNA Vaccines for Infectious Diseases. Methods Mol. Biol. 2017, 1499, 109–121. [Google Scholar] [PubMed]
- Zarghampoor, F.; Azarpira, N.; Khatami, S.R.; Behzad-Behbahani, A.; Foroughmand, A.M. Improved translation efficiency of therapeutic mRNA. Gene 2019, 707, 231–238. [Google Scholar] [CrossRef]
- van Gils, M.J.; Lavell, A.H.A.; van der Straten, K.; Appelman, B.; Bontjer, I.; Poniman, M.; Burger, J.A.; Oomen, M.; Bouhuijs, J.H.; van Vught, L.A.; et al. Four SARS-CoV-2 vaccines induce quantitatively different antibody responses against SARS-CoV-2 variants. medRxiv 2021. [Google Scholar] [CrossRef]
- Steensels, D.; Pierlet, N.; Penders, J.; Mesotten, D.; Heylen, L. Comparison of SARS-CoV-2 Antibody Response Following Vaccination With BNT162b2 and mRNA-1273. JAMA 2021, 326, 1533–1535. [Google Scholar] [CrossRef]
- Cohn, B.A.; Cirillo, P.M.; Murphy, C.C.; Krigbaum, N.Y.; Wallace, A.W. SARS-CoV-2 vaccine protection and deaths among US veterans during 2021. Science 2021, eabm0620. [Google Scholar] [CrossRef]
- Naaber, P.; Tserel, L.; Kangro, K.; Sepp, E.; Jurjenson, V.; Adamson, A.; Haljasmagi, L.; Rumm, A.P.; Maruste, R.; Karner, J.; et al. Dynamics of antibody response to BNT162b2 vaccine after six months: A longitudinal prospective study. Lancet Reg. Health Eur. 2021, 10, 100208. [Google Scholar] [CrossRef] [PubMed]
- Israel, A.; Shenhar, Y.; Green, I.; Merzon, E.; Golan-Cohen, A.; Schaffer, A.A.; Ruppin, E.; Vinker, S.; Magen, E. Large-scale study of antibody titer decay following BNT162b2 mRNA vaccine or SARS-CoV-2 infection. medRxiv 2021. [Google Scholar] [CrossRef]
- Mangge, H.; Kneihsl, M.; Schnedl, W.; Sendlhofer, G.; Curcio, F.; Domenis, R. Immune Responses against SARS-CoV-2-Questions and Experiences. Biomedicines 2021, 9, 1342. [Google Scholar] [CrossRef] [PubMed]
- McClung, N.; Chamberland, M.; Kinlaw, K.; Matthew, D.B.; Wallace, M.; Bell, B.P.; Lee, G.M.; Talbot, H.K.; Romero, J.R.; Oliver, S.E.; et al. The Advisory Committee on Immunization Practices’ Ethical Principles for Allocating Initial Supplies of COVID-19 Vaccine-United States, 2020. Am. J. Transplant. 2021, 21, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Widge, A.T.; Rouphael, N.G.; Jackson, L.A.; Anderson, E.J.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Durability of Responses after SARS-CoV-2 mRNA-1273 Vaccination. N. Engl. J. Med. 2021, 384, 80–82. [Google Scholar] [CrossRef]
- Pegu, A.; O’Connell, S.E.; Schmidt, S.D.; O’Dell, S.; Talana, C.A.; Lai, L.; Albert, J.; Anderson, E.; Bennett, H.; Corbett, K.S.; et al. Durability of mRNA-1273 vaccine-induced antibodies against SARS-CoV-2 variants. Science 2021, 373, 1372–1377. [Google Scholar] [CrossRef] [PubMed]
- Jeewandara, C.; Kamaladasa, A.; Pushpakumara, P.D.; Jayathilaka, D.; Aberathna, I.S.; Danasekara, D.; Guruge, D.; Ranasinghe, T.; Dayarathna, S.; Pathmanathan, T.; et al. Immune responses to a single dose of the AZD1222/Covishield vaccine in health care workers. Nat. Commun. 2021, 12, 4617. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Pradhan, S.K.; Pati, S.; Sahu, S.; Nanda, R.K. Waning of Anti-spike Antibodies in AZD1222 (ChAdOx1) Vaccinated Healthcare Providers: A Prospective Longitudinal Study. Cureus 2021, 13, e19879. [Google Scholar] [CrossRef] [PubMed]
- Flaxman, A.; Marchevsky, N.G.; Jenkin, D.; Aboagye, J.; Aley, P.K.; Angus, B.; Belij-Rammerstorfer, S.; Bibi, S.; Bittaye, M.; Cappuccini, F.; et al. Reactogenicity and immunogenicity after a late second dose or a third dose of ChAdOx1 nCoV-19 in the UK: A substudy of two randomised controlled trials (COV001 and COV002). Lancet 2021, 398, 981–990. [Google Scholar] [CrossRef]
- Pouwels, K.B.; Pritchard, E.; Matthews, P.C.; Stoesser, N.; Eyre, D.W.; Vihta, K.D.; House, T.; Hay, J.; Bell, J.I.; Newton, J.N.; et al. Effect of Delta variant on viral burden and vaccine effectiveness against new SARS-CoV-2 infections in the UK. Nat. Med. 2021, 27, 2127–2135. [Google Scholar] [CrossRef]
- Sanderson, K. COVID vaccines protect against Delta, but their effectiveness wanes. Nature 2021. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, E.; Matthews, P.C.; Stoesser, N.; Eyre, D.W.; Gethings, O.; Vihta, K.D.; Jones, J.; House, T.; VanSteenHouse, H.; Bell, I.; et al. Impact of vaccination on new SARS-CoV-2 infections in the United Kingdom. Nat. Med. 2021, 27, 1370–1378. [Google Scholar] [CrossRef] [PubMed]
- Andrejko, K.L.; Pry, J.; Myers, J.F.; Jewell, N.P.; Openshaw, J.; Watt, J.; Jain, S.; Lewnard, J.A. California COVID-19 Case-Control Study Team. Prevention of COVID-19 by mRNA-based vaccines within the general population of California. Clin. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Collier, A.Y.; Yu, J.; McMahan, K.; Liu, J.; Chandrashekar, A.; Maron, J.S.; Atyeo, C.; Martinez, D.R.; Ansel, J.L.; Aguayo, R.; et al. Differential Kinetics of Immune Responses Elicited by COVID-19 Vaccines. N. Engl. J. Med. 2021, 385, 2010–2012. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.J.; Xue, M.; Zheng, P.; Lyu, J.; Zhan, Z.; Hu, H.; Zhang, Y.; Zhang, X.D.; Sun, B. Factors Affecting the Antibody Immunogenicity of Vaccines against SARS-CoV-2: A Focused Review. Vaccines 2021, 9, 869. [Google Scholar] [CrossRef] [PubMed]
- Pellini, R.; Venuti, A.; Pimpinelli, F.; Abril, E.; Blandino, G.; Campo, F.; Conti, L.; De Virgilio, A.; De Marco, F.; Di Domenico, E.G.; et al. Initial observations on age, gender, BMI and hypertension in antibody responses to SARS-CoV-2 BNT162b2 vaccine. EClinicalMedicine 2021, 36, 100928. [Google Scholar] [CrossRef] [PubMed]
- Collier, D.A.; Ferreira, I.A.T.M.; Kotagiri, P.; Datir, R.P.; Lim, E.Y.; Touizer, E.; Meng, B.; Abdullahi, A.; Baker, S.; Dougan, G.; et al. Age-related immune response heterogeneity to SARS-CoV-2 vaccine BNT162b2. Nature 2021, 596, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Notarte, K.I.; Ver, A.T.; Velasco, J.V.; Pastrana, A.; Catahay, J.A.; Salvagno, G.L.; Yap, E.P.H.; Martinez-Sobrido, L.; Torrelles, J.; Lippi, G.; et al. Effects of Age, Sex, Serostatus and Underlying Comorbidities on Humoral Response Post-SARS-CoV-2 Pfizer-BioNTech Vaccination—A Systematic Review. medRvix 2021. [Google Scholar] [CrossRef]
- Bajaj, V.; Gadi, N.; Spihlman, A.P.; Wu, S.C.; Choi, C.H.; Moulton, V.R. Aging, Immunity, and COVID-19: How Age Influences the Host Immune Response to Coronavirus Infections? Front. Physiol. 2020, 11, 571416. [Google Scholar] [CrossRef] [PubMed]
- Abu Jabal, K.; Ben-Amram, H.; Beiruti, K.; Batheesh, Y.; Sussan, C.; Zarka, S.; Edelstein, M. Impact of age, ethnicity, sex and prior infection status on immunogenicity following a single dose of the BNT162b2 mRNA COVID-19 vaccine: Real-world evidence from healthcare workers, Israel, December 2020 to January 2021. Eurosurveillance 2021, 26, 2100096. [Google Scholar] [CrossRef] [PubMed]
- Fischinger, S.; Boudreau, C.M.; Butler, A.L.; Streeck, H.; Alter, G. Sex differences in vaccine-induced humoral immunity. Semin. Immunopathol. 2019, 41, 239–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoiland, R.L.; Fergusson, N.A.; Mitra, A.R.; Griesdale, D.E.G.; Devine, D.V.; Stukas, S.; Cooper, J.; Thiara, S.; Foster, D.; Chen, L.Y.C.; et al. The association of ABO blood group with indices of disease severity and multiorgan dysfunction in COVID-19. Blood Adv. 2020, 4, 4981–4989. [Google Scholar] [CrossRef] [PubMed]
- Gerard, C.; Maggipinto, G.; Minon, J.M. COVID-19 and ABO blood group: Another viewpoint. Br. J. Haematol. 2020, 190, e93–e94. [Google Scholar] [CrossRef] [PubMed]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n (%) | Oxford/AstraZeneca | Pfizer/BioNTech | Moderna | Johnson & Johnson | Unvaccinated | |
|---|---|---|---|---|---|---|
| n | 221 (23.2%) | 562 (58.9%) | 56 (5.9%) | 30 (3.1%) | 85 (8.9%) | |
| Age (years) | ≤30 | 21 (9.50%) | 17 (3.02%) | 3 (5.36%) | 3 (10.00%) | 12 (14.12%) |
| 30–40 | 64 (28.96%) | 114 (20.28%) | 15 (26.79%) | 8 (26.67%) | 21 (24.71%) | |
| 40–50 | 75 (33.93%) | 210 (37.37%) | 15 (26.79%) | 7 (23.33%) | 25 (29.41%) | |
| 50–65 | 48 (21.71%) | 184 (32.74%) | 19 (33.93%) | 12 (40.00%) | 24 (28.24%) | |
| 65+ | 13 (5.88%) | 37 (6.58%) | 4 (7.14%) | 0 (0.00%) | 3 (3.53%) | |
| Gender | female | 136 (61.54%) | 388 (69.04%) | 42 (75.00%) | 22 (73.33%) | 55 (64.71%) |
| male | 85 (38.46%) | 174 (30.96%) | 14 (25.00%) | 8 (26.67%) | 30 (35.29%) | |
| COVID-19 history | no | 183 (82.81%) | 438 (77.94%) | 39 (69.64%) | 20 (66.67%) | 32 (38.10) |
| yes | 38 (17.19%) | 122 (22.06%) | 17 (30.36%) | 10 (33.33%) | 52 (61.90%) | |
| Blood group | 0 | 45 (20.36%) | 129 (22.95%) | 13 (23.21%) | 10 (33.33%) | 7 (8.23%) |
| A | 66 (29.86%) | 153 (27.22%) | 16 (28.57%) | 8 (26.67%) | 2 (2.36%) | |
| B | 28 (12.67%) | 87 (15.48%) | 10 (17.86%) | 5 (16.67%) | 0 (0.00%) | |
| AB | 16 (7.24%) | 43 (7.65%) | 3 (5.36%) | 2 (6.67%) | 1 (1.18%) | |
| n/a | 66 (29.86%) | 150 (26.69%) | 14 (25.00%) | 5 (16.67%) | 75 (88.23%) | |
| Measurement | Factor | Sample Mean | 10% Trimmed Mean | Lower 95% CI | Upper 95% CI |
|---|---|---|---|---|---|
| Prior COVID-19 infection | |||||
| 1 | No | 1184.74 | 880.03 | 761.05 | 999.00 |
| Yes | 1746.16 | 1386.48 | 1003.90 | 1769.06 | |
| 2 | No | 884.47 | 678.47 | 609.75 | 747.19 |
| Yes | 2582.02 | 2233.98 | 1803.26 | 2664.71 | |
| 3 | No | 392.70 | 281.67 | 254.05 | 309.29 |
| Yes | 1466.29 | 1148.91 | 895.19 | 1402.64 | |
| Overall | No | 820.63 | 574.73 | 530.44 | 619.01 |
| yes | 1931.49 | 1575.96 | 1367.56 | 1784.37 | |
| Vaccine type | |||||
| 1 | BNT162b2 | 1563.97 | 1286.18 | 1127.87 | 1444.49 |
| ChAdOx1nCoV-2019 | 496.19 | 316.36 | 206.78 | 425.95 | |
| JNJ-78436735 | 202.59 | 146.55 | −76.26 | 369.35 | |
| mRNA-1273 | 2508.98 | 2140.85 | 926.40 | 3355.30 | |
| 2 | BNT162b2 | 1252.67 | 945.77 | 826.15 | 1065.39 |
| ChAdOx1nCoV-2019 | 738.17 | 583.72 | 494.20 | 673.25 | |
| JNJ-78436735 | 1138.68 | 765.02 | −20.87 | 1550.90 | |
| mRNA-1273 | 3365.29 | 3189.02 | 2349.41 | 4028.62 | |
| 3 | BNT162b2 | 618.51 | 408.13 | 348.36 | 467.90 |
| ChAdOx1nCoV-2019 | 357.08 | 287.51 | 241.28 | 333.75 | |
| JNJ-78436735 | 796.12 | 534.04 | −281.35 | 1349.43 | |
| mRNA-1273 | 1714.50 | 1500.38 | 969.27 | 2031.50 | |
| Overall | BNT162b2 | 1145.10 | 842.10 | 772.61 | 911.59 |
| ChAdOx1nCoV-2019 | 530.48 | 377.82 | 333.23 | 422.41 | |
| JNJ-78436735 | 712.46 | 426.24 | 75.23 | 777.24 | |
| mRNA-1273 | 2529.59 | 2196.88 | 1704.35 | 2689.40 | |
| Measurement number | |||||
| 1 | 1307.79 | 967.35 | 843.27 | 1091.43 | |
| 2 | 1256.53 | 913.78 | 814.24 | 1013.32 | |
| 3 | 628.01 | 404.81 | 357.76 | 451.86 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczepanek, J.; Skorupa, M.; Goroncy, A.; Jarkiewicz-Tretyn, J.; Wypych, A.; Sandomierz, D.; Jarkiewicz-Tretyn, A.; Dejewska, J.; Ciechanowska, K.; Pałgan, K.; et al. Anti-SARS-CoV-2 IgG against the S Protein: A Comparison of BNT162b2, mRNA-1273, ChAdOx1 nCoV-2019 and Ad26.COV2.S Vaccines. Vaccines 2022, 10, 99. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10010099
Szczepanek J, Skorupa M, Goroncy A, Jarkiewicz-Tretyn J, Wypych A, Sandomierz D, Jarkiewicz-Tretyn A, Dejewska J, Ciechanowska K, Pałgan K, et al. Anti-SARS-CoV-2 IgG against the S Protein: A Comparison of BNT162b2, mRNA-1273, ChAdOx1 nCoV-2019 and Ad26.COV2.S Vaccines. Vaccines. 2022; 10(1):99. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10010099
Chicago/Turabian StyleSzczepanek, Joanna, Monika Skorupa, Agnieszka Goroncy, Joanna Jarkiewicz-Tretyn, Aleksandra Wypych, Dorota Sandomierz, Aleksander Jarkiewicz-Tretyn, Joanna Dejewska, Karolina Ciechanowska, Krzysztof Pałgan, and et al. 2022. "Anti-SARS-CoV-2 IgG against the S Protein: A Comparison of BNT162b2, mRNA-1273, ChAdOx1 nCoV-2019 and Ad26.COV2.S Vaccines" Vaccines 10, no. 1: 99. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10010099