Additional Evidence on Serological Correlates of Protection against Measles: An Observational Cohort Study among Once Vaccinated Children Exposed to Measles

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants
2.3. Data Collection
2.4. Laboratory Tests
2.5. Case Classification
2.6. Statistical Methods
2.7. Ethics Statement
3. Results
3.1. Descriptive Results
3.2. Correlates of Protection
3.3. Sensitivity Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Laboratory Methods
References
- Rota, P.A.; Moss, W.J.; Takeda, M.; de Swart, R.L.; Thompson, K.M.; Goodson, J.L. Measles. Nat. Rev. Dis. Prim. 2016, 2, 16049. [Google Scholar] [CrossRef] [PubMed]
- Laksono, B.M.; de Vries, R.D.; Verburgh, R.J.; Visser, E.G.; de Jong, A.; Fraaij, P.L.; Ruijs, W.L.; Nieuwenhuijse, D.F.; van den Ham, H.J.; Koopmans, M.P. Studies into the mechanism of measles-associated immune suppression during a measles outbreak in the Netherlands. Nat. Commun. 2018, 9, 4944. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.J.; Metcalf, C.J.; de Swart, R.L.; Osterhaus, A.D.; Grenfell, B.T. Long-term measles-induced immunomodulation increases overall childhood infectious disease mortality. Science 2015, 348, 694–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabbagh, A.; Patel, M.K.; Dumolard, L.; Gacic-Dobo, M.; Mulders, M.N.; Okwo-Bele, J.M.; Kretsinger, K.; Papania, M.J.; Rota, P.A.; Goodson, J.L. Progress Toward Regional Measles Elimination—Worldwide, 2000-2016. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 1148–1153. [Google Scholar] [CrossRef]
- Mollema, L.; Smits, G.P.; Berbers, G.A.; Van Der Klis, F.R.; Van Binnendijk, R.S.; De Melker, H.E.; Hahne, S.J. High risk of a large measles outbreak despite 30 years of measles vaccination in The Netherlands. Epidemiol. Infect. 2014, 142, 1100–1108. [Google Scholar] [CrossRef]
- Krugman, S. Further-attenuated measles vaccine: Characteristics and use. Rev. Infect. Dis. 1983, 5, 477–481. [Google Scholar] [CrossRef]
- Christenson, B.; Bottiger, M. Measles antibody: Comparison of long-term vaccination titres, early vaccination titres and naturally acquired immunity to and booster effects on the measles virus. Vaccine 1994, 12, 129–133. [Google Scholar] [CrossRef]
- Van den Hof, S.; Berbers, G.A.; de Melker, H.E.; Conyn-van Spaendonck, M.A. Sero-epidemiology of measles antibodies in the Netherlands, a cross-sectional study in a national sample and in communities with low vaccine coverage. Vaccine 1999, 18, 931–940. [Google Scholar] [CrossRef]
- Panum, P.L. Observations made during the epidemic of measles on the Faroe Islands in the year 1846. Bibiliothek Laeg. 1847, 1, 270–344. [Google Scholar]
- LeBaron, C.W.; Beeler, J.; Sullivan, B.J.; Forghani, B.; Bi, D.; Beck, C.; Audet, S.; Gargiullo, P. Persistence of measles antibodies after 2 doses of measles vaccine in a postelimination environment. Arch. Pediatr. Adolesc. Med. 2007, 161, 294–301. [Google Scholar] [CrossRef]
- Davidkin, I.; Jokinen, S.; Broman, M.; Leinikki, P.; Peltola, H. Persistence of measles, mumps, and rubella antibodies in an MMR-vaccinated cohort: A 20-year follow-up. J. Infect. Dis. 2008, 197, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Haralambieva, I.H.; Ovsyannikova, I.G.; O’Byrne, M.; Pankratz, V.S.; Jacobson, R.M.; Poland, G.A. A large observational study to concurrently assess persistence of measles specific B-cell and T-cell immunity in individuals following two doses of MMR vaccine. Vaccine 2011, 29, 4485–4491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, J.B.; Rota, J.S.; Hickman, C.J.; Sowers, S.B.; Mercader, S.; Rota, P.A.; Bellini, W.J.; Huang, A.J.; Doll, M.K.; Zucker, J.R.; et al. Outbreak of measles among persons with prior evidence of immunity, New York City, 2011. Clin. Infect. Dis. 2014, 58, 1205–1210. [Google Scholar] [PubMed]
- Hahne, S.J.; Nic Lochlainn, L.M.; van Burgel, N.D.; Kerkhof, J.; Sane, J.; Yap, K.B.; van Binnendijk, R.S. Measles Outbreak Among Previously Immunized Healthcare Workers, the Netherlands, 2014. J. Infect. Dis. 2016, 214, 1980–1986. [Google Scholar] [CrossRef] [PubMed]
- Mathias, R.G.; Meekison, W.G.; Arcand, T.A.; Schechter, M.T. The role of secondary vaccine failures in measles outbreaks. Am. J. Public Health 1989, 79, 475–478. [Google Scholar] [CrossRef]
- Cherry, J.D.; Zahn, M. Clinical Characteristics of Measles in Previously Vaccinated and Unvaccinated Patients in California. Clin. Infect. Dis. 2018, 67, 1315–1319. [Google Scholar] [CrossRef] [Green Version]
- Strebel, P.M.; Papania, M.J.; Gastañaduy, P.A.; Goodson, J.L. 37—Measles Vaccines. In Plotkin’s Vaccines, 7th ed.; Plotkin, S.A., Orenstein, W.A., Offit, P.A., Edwards, K.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 579–618. [Google Scholar]
- Cohen, B.J.; Audet, S.; Andrews, N.; Beeler, J. Plaque reduction neutralization test for measles antibodies: Description of a standardised laboratory method for use in immunogenicity studies of aerosol vaccination. Vaccine 2007, 26, 59–66. [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef]
- Chen, R.T.; Markowitz, L.E.; Albrecht, P.; Stewart, J.A.; Mofenson, L.M.; Preblud, S.R.; Orenstein, W.A. Measles antibody: Reevaluation of protective titers. J. Infect. Dis. 1990, 162, 1036–1042. [Google Scholar] [CrossRef]
- Samb, B.; Aaby, P.; Whittle, H.C.; Seck, A.M.; Rahman, S.; Bennett, J.; Markowitz, L.; Simondon, F. Serologic status and measles attack rates among vaccinated and unvaccinated children in rural Senegal. Pediatr. Infect. Dis. J. 1995, 14, 203–209. [Google Scholar] [CrossRef]
- Muller, C.P.; Huiss, S.; Schneider, F. Secondary immune responses in parents of children with recent measles. Lancet 1996, 348, 1379–1380. [Google Scholar] [CrossRef]
- Lee, M.S.; Nokes, D.J.; Hsu, H.M.; Lu, C.F. Protective titres of measles neutralising antibody. J. Med. Virol. 2000, 62, 511–517. [Google Scholar] [CrossRef]
- Van Lier, E.A.; Oomen, P.J.; Giesbers, H.; van Vliet, J.A.; Drijfhout, I.H.; Zonnenberg-Hoff, I.F.; De Melker, H.E. Vaccinatiegraad en Jaarverslag Rijksvaccinatieprogramma Nederland 2018; RIVM: Bilthoven, The Netherlands, 2019. [Google Scholar]
- Van den Hof, S.; Conyn-van Spaendonck, M.A.; van Steenbergen, J.E. Measles epidemic in the Netherlands, 1999–2000. J. Infect. Dis. 2002, 186, 1483–1486. [Google Scholar] [CrossRef] [PubMed]
- Bijkerk, H.; Bilkert-Mooiman, M.A.; Houtters, H.J. The immunization status of patients registered with measles during the 1987-88 epidemic. Ned. Tijdschr. Voor Geneeskd. 1989, 133, 29–32. [Google Scholar]
- Ruijs, W.L.; Hautvast, J.L.; van Ansem, W.J.; Akkermans, R.P.; van′t Spijker, K.; Hulscher, M.E.; van der Velden, K. Measuring vaccination coverage in a hard to reach minority. Eur. J. Public Health 2012, 22, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Wallinga, J.; Heijne, J.C.; Kretzschmar, M. A measles epidemic threshold in a highly vaccinated population. PLoS Med. 2005, 2, e316. [Google Scholar] [CrossRef]
- Woudenberg, T.; van Binnendijk, R.S.; Sanders, E.A.; Wallinga, J.; de Melker, H.E.; Ruijs, W.L.; Hahne, S.J. Large measles epidemic in the Netherlands, May 2013 to March 2014: Changing epidemiology. Euro Surveill. 2017, 22, 30443. [Google Scholar] [CrossRef]
- Smits, G.P.; van Gageldonk, P.G.; Schouls, L.M.; van der Klis, F.R.; Berbers, G.A. Development of a bead-based multiplex immunoassay for simultaneous quantitative detection of IgG serum antibodies against measles, mumps, rubella, and varicella-zoster virus. Clin. Vaccine Immunol. 2012, 19, 396–400. [Google Scholar] [CrossRef]
- De Swart, R.L.; Vos, H.W.; UytdeHaag, F.G.; Osterhaus, A.D.; van Binnendijk, R.S. Measles virus fusion protein- and hemagglutinin-transfected cell lines are a sensitive tool for the detection of specific antibodies by a FACS-measured immunofluorescence assay. J. Virol. Methods 1998, 71, 35–44. [Google Scholar] [CrossRef]
- El Mubarak, H.S.; Ibrahim, S.A.; Vos, H.W.; Mukhtar, M.M.; Mustafa, O.A.; Wild, T.F.; Osterhaus, A.D.; de Swart, R.L. Measles virus protein-specific IgM, IgA, and IgG subclass responses during the acute and convalescent phase of infection. J. Med. Virol. 2004, 72, 290–298. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Commission implementing decision (EU) 2018/945 of 22 June 2018 on the communicable diseases and related special health issues to be covered by epidemiological surveillance as well as relevant case definitions. Off. J. Eur. Union 2018, 30–31. [Google Scholar]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- DeLong, E.R.; DeLong, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Muller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Aaby, P.; Bukh, J.; Leerhoy, J.; Lisse, I.M.; Mordhorst, C.H.; Pedersen, I.R. Vaccinated children get milder measles infection: A community study from Guinea-Bissau. J. Infect. Dis. 1986, 154, 858–863. [Google Scholar] [CrossRef]
- Paunio, M.; Peltola, H.; Valle, M.; Davidkin, I.; Virtanen, M.; Heinonen, O.P. Explosive school-based measles outbreak: Intense exposure may have resulted in high risk, even among revaccinees. Am. J. Epidemiol. 1998, 148, 1103–1110. [Google Scholar] [CrossRef]
- Cohen, B.J.; Doblas, D.; Andrews, N. Comparison of plaque reduction neutralisation test (PRNT) and measles virus-specific IgG ELISA for assessing immunogenicity of measles vaccination. Vaccine 2008, 26, 6392–6397. [Google Scholar] [CrossRef]
- Whittle, H.C.; Aaby, P.; Samb, B.; Jensen, H.; Bennett, J.; Simondon, F. Effect of subclinical infection on maintaining immunity against measles in vaccinated children in West Africa. Lancet 1999, 353, 98–102. [Google Scholar] [CrossRef]
- Ruckdeschel, J.C.; Graziano, K.D.; Mardiney, M.R., Jr. Additional evidence that the cell-associated immune system is the primary host defense against measles (rubeola). Cell. Immunol. 1975, 17, 11–18. [Google Scholar] [CrossRef]
- Griffin, D.E. The Immune Response in Measles: Virus Control, Clearance and Protective Immunity. Viruses 2016, 8, 282. [Google Scholar] [CrossRef]
- Kremer, J.R.; Schneider, F.; Muller, C.P. Waning antibodies in measles and rubella vaccinees—A longitudinal study. Vaccine 2006, 24, 2594–2601. [Google Scholar] [CrossRef]
- Sowers, S.B.; Rota, J.S.; Hickman, C.J.; Mercader, S.; Redd, S.; McNall, R.J.; Williams, N.; McGrew, M.; Walls, M.L.; Rota, P.A.; et al. High Concentrations of Measles Neutralizing Antibodies and High-Avidity Measles IgG Accurately Identify Measles Reinfection Cases. Clin. Vaccine Immunol. 2016, 23, 707–716. [Google Scholar] [PubMed] [Green Version]
- Woudenberg, T. The tip of the iceberg: Incompleteness of measles reporting during a large outbreak in The Netherlands in 2013–2014. Epidemiol. Infect. 2018, 147, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.J.; Parry, R.P.; Doblas, D.; Samuel, D.; Warrener, L.; Andrews, N.; Brown, D. Measles immunity testing: Comparison of two measles IgG ELISAs with plaque reduction neutralisation assay. J. Virol. Methods 2006, 131, 209–212. [Google Scholar] [CrossRef] [PubMed]




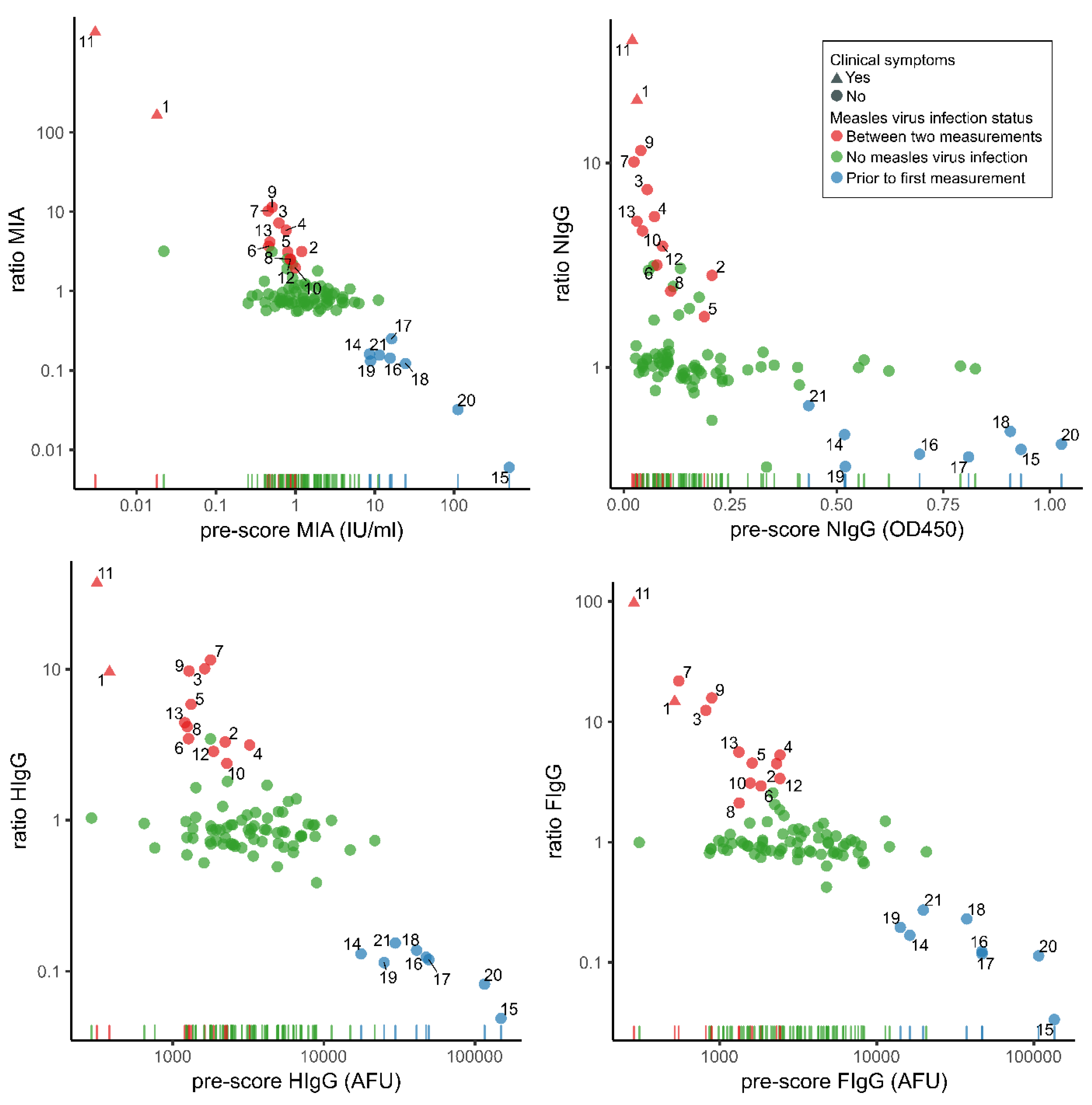{kind=link}
{kind=link}
{kind=link}
| Characteristic | N (%) |
|---|---|
| Sex | |
| Boy | 41 (45) |
| Girl | 50 (55) |
| Enrollment | |
| School | 70 (77) |
| National vaccination register | 21 (23) |
| Age at first MMR in months (IQR) | 14.5 (14.4–15.1) |
| Median follow-up time in months (IQR) | 8.4 (6.6–8.4) |
| Self-reported symptoms along the follow–up | |
| Fever | 38 (42) |
| Rash | 3 (3) |
| Cough | 46 (51) |
| Runny nose | 42 (46) |
| Conjunctivitis | 8 (9) |
| Exposure to MV * | |
| High | 82 (90) |
| Medium | 9 (10) |
| Age at first sampling in years (IQR) | 6.5 (5.5–7.5) |
| Age at second sampling in years (IQR) | 7.3 (6.2–8.2) |
| Antibody Concentrations Prior to Exposure (IU/mL) | Attack Rate (Cases/Exposed) (%) |
|---|---|
| ≤0.06 | 67 (2/3) |
| 0.35–1.21 | 22 (6/27) |
| 1.21–2.54 | 19 (5/27) |
| >2.54 | 0 (0/26) |
| Total | 16 (13/83) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woudenberg, T.; van Binnendijk, R.; Veldhuijzen, I.; Woonink, F.; Ruijs, H.; van der Klis, F.; Kerkhof, J.; de Melker, H.; de Swart, R.; Hahné, S. Additional Evidence on Serological Correlates of Protection against Measles: An Observational Cohort Study among Once Vaccinated Children Exposed to Measles. Vaccines 2019, 7, 158. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines7040158
Woudenberg T, van Binnendijk R, Veldhuijzen I, Woonink F, Ruijs H, van der Klis F, Kerkhof J, de Melker H, de Swart R, Hahné S. Additional Evidence on Serological Correlates of Protection against Measles: An Observational Cohort Study among Once Vaccinated Children Exposed to Measles. Vaccines. 2019; 7(4):158. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines7040158
Chicago/Turabian StyleWoudenberg, Tom, Rob van Binnendijk, Irene Veldhuijzen, Frits Woonink, Helma Ruijs, Fiona van der Klis, Jeroen Kerkhof, Hester de Melker, Rik de Swart, and Susan Hahné. 2019. "Additional Evidence on Serological Correlates of Protection against Measles: An Observational Cohort Study among Once Vaccinated Children Exposed to Measles" Vaccines 7, no. 4: 158. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines7040158