Design of an Epitope-Based Vaccine Ensemble for Animal Trypanosomiasis by Computational Methods


Abstract
:1. Introduction
2. Materials and Methods
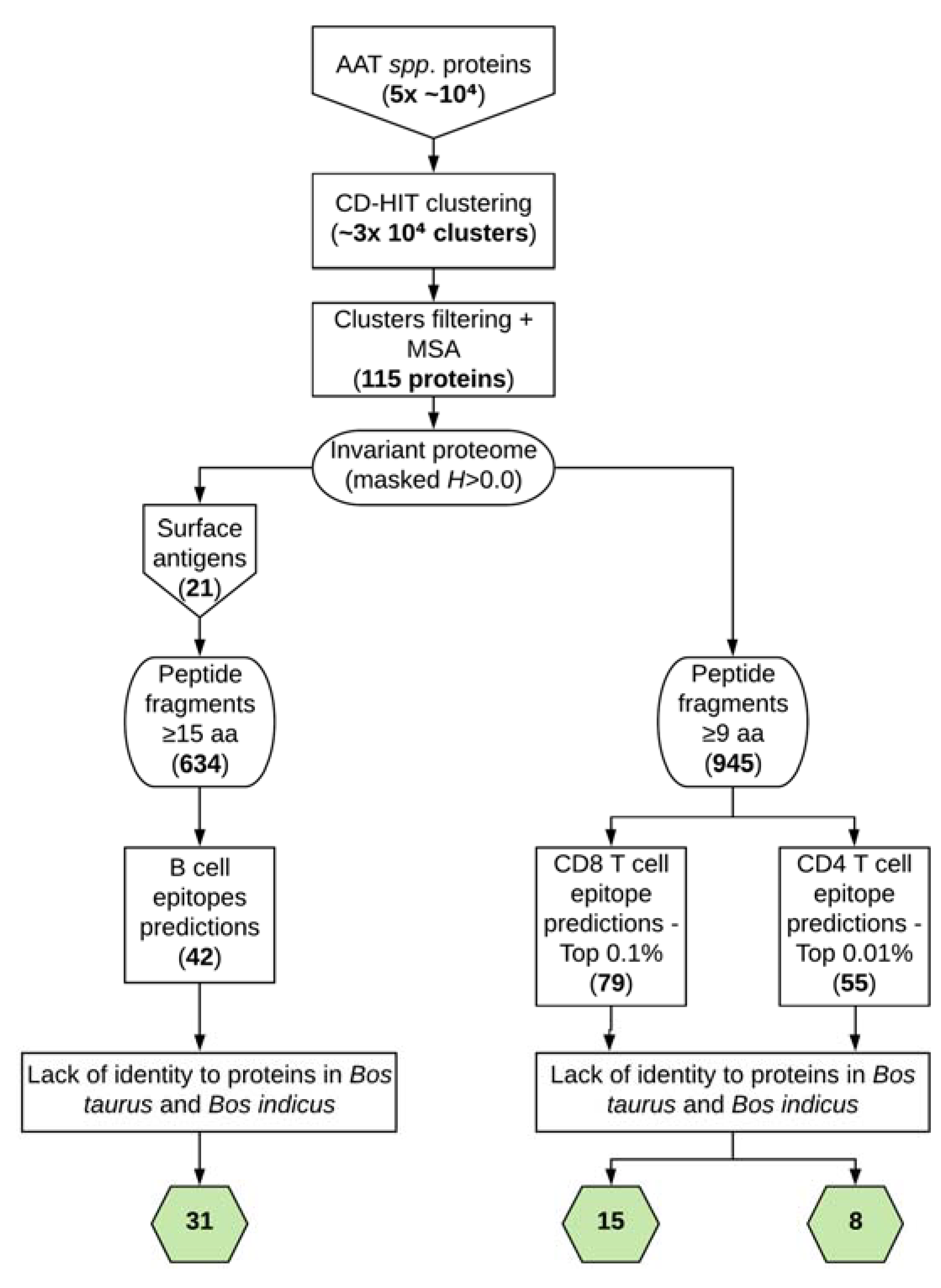
2.1. Collection of Protein Sequences from Trypanosome Spp. That Cause AAT
2.2. Generation of Clusters
2.3. Generation of the Invariable Proteome of Trypanosome Spp. Responsible for AAT
2.4. Prediction of B-Cell Epitopes
2.5. Prediction of T-cell Epitopes
2.6. Blast Searches for Epitopes Identity and Other Analysis Procedures
3. Results
3.1. Identification of the Invariant Proteome of AAT-Causing Trypanosomes
3.2. Prediction of B-Cell Epitopes
3.3. Prediction of T-Cell Epitopes
3.4. Design of a Genetic Vaccine Ensemble
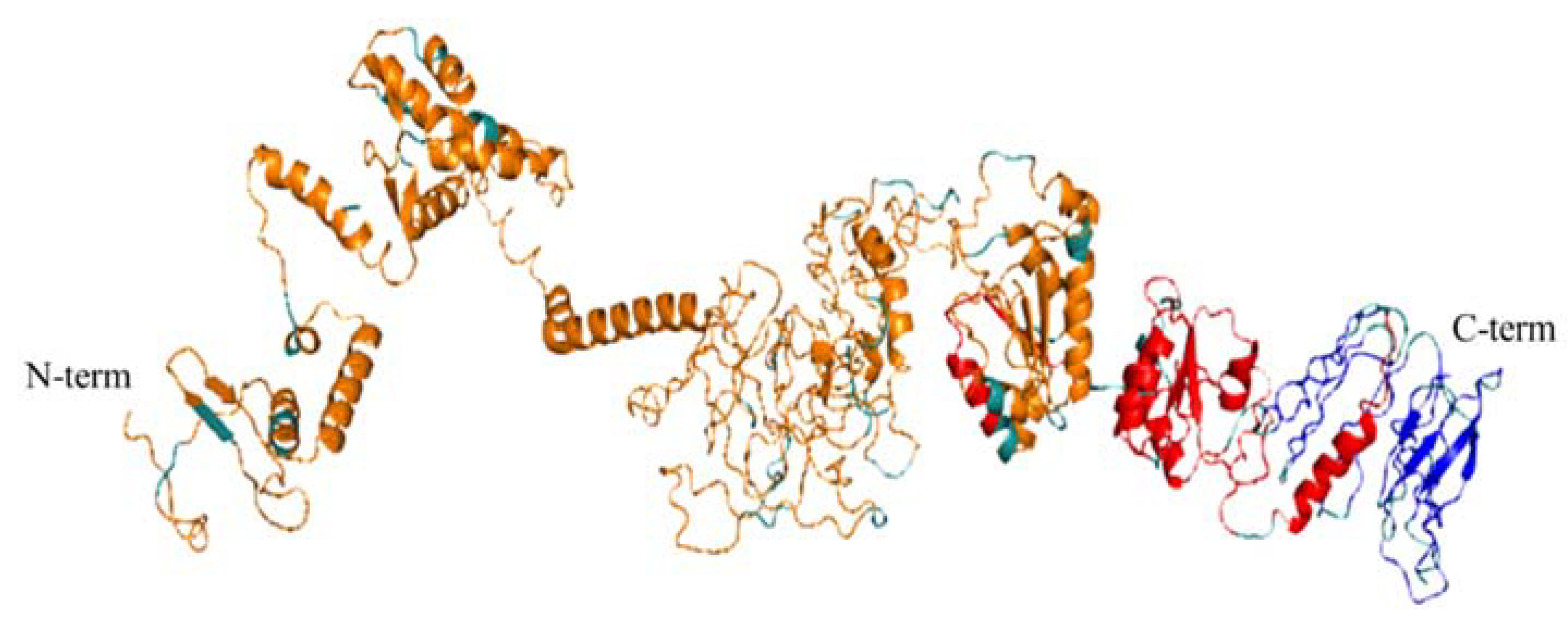
3.5. 3D Modelling of the Peptide Vaccine Ensemble
4. Discussion
5. Conclusions and Limitations
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
References
- Morrison, L.J.; Vezza, L.; Rowan, T.; Hope, J.C. Animal African trypanosomiasis: Time to increase focus on clinically relevant parasite and host species. Trends Parasitol. 2016, 32, 599–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, S.J.; Mansfield, J.M. Prospects for vaccination against pathogenic African trypanosomes. Parasite Immunol. 2016, 38, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Sileghem, M.; Naessens, J. Are CD8 T cells involved in control of African trypanosomiasis in a natural host environment? Eur. J. Immunol. 1995, 25, 1965–1971. [Google Scholar] [CrossRef] [PubMed]
- Yaro, M.; Munyard, K.A.; Stear, M.J.; Groth, D.M. Combatting African animal trypanosomiasis (AAT) in livestock: The potential role of trypanotolerance. Vet. Parasitol. 2016, 225, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekonnen, G.; Mohammed, E.F.; Kidane, W.; Nesibu, A.; Yohannes, H.; Van Reet, N.; Büscher, P.; Birhanu, H. Isometamidium chloride and homidium chloride fail to cure mice infected with Ethiopian Trypanosoma evansi type A and B. PLoS Negl. Trop. Dis. 2018, 12, e0006790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magez, S.; Caljon, G.; Tran, T.; Stijlemans, B.; Radwanska, M. Current status of vaccination against African trypanosomiasis. Parasitology 2010, 137, 2017–2027. [Google Scholar] [CrossRef] [Green Version]
- La Greca, F.; Magez, S. Vaccination against trypanosomiasis: Can it be done or is the trypanosome truly the ultimate immune destroyer and escape artist? Hum. Vaccines 2011, 7, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Stijlemans, B.; Radwanska, M.; De Trez, C.; Magez, S. African trypanosomes undermine humoral responses and vaccine development: Link with inflammatory responses? Front. Immunol. 2017, 8, 582. [Google Scholar] [CrossRef]
- Tewari, A.K.; Kurup, S.P.; Baidya, S.; Barta, J.R.; Sharma, B. Protective antibody and cytokine responses in mice following immunization with recombinant beta-tubulin and subsequent Trypanosoma evansi challenge. Parasites Vectors 2015, 8, 580. [Google Scholar] [CrossRef] [Green Version]
- Lança, A.S.; De Sousa, K.P.; Atouguia, J.; Prazeres, D.M.F.; Monteiro, G.A.; Silva, M.S. Trypanosoma brucei: Immunisation with plasmid DNA encoding invariant surface glycoprotein gene is able to induce partial protection in experimental African trypanosomiasis. Exp. Parasitol. 2011, 127, 18–24. [Google Scholar] [CrossRef]
- Taylor, K.A. Immune responses of cattle to African trypanosomes: Protective or pathogenic? Int. J. Parasitol. 1998, 28, 219–240. [Google Scholar] [CrossRef]
- Aslett, M.; Aurrecoechea, C.; Berriman, M.; Brestelli, J.; Brunk, B.P.; Carrington, M.; Depledge, D.P.; Fischer, S.; Gajria, B.; Gao, X.; et al. TriTrypDB: A functional genomic resource for the Trypanosomatidae. Nucleic Acids Res. 2010, 38, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C. The mathematical theory of commnication. Bell Syst. Technol. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.J.; Lee, C.Y.; Ibrahim, S.; Watts, P.; Shlomchik, M.; Weigert, M.; Litwin, S. A Shannon entropy analysis of immunoglobulin and T cell receptor. Mol. Immunol. 1997, 34, 1067–1082. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [Green Version]
- Dhanda, S.K.; Mahajan, S.; Paul, S.; Yan, Z.; Kim, H.; Jespersen, M.C.; Jurtz, V.; Adreatta, M.; Greenbaum, J.A.; Marcatili, P.; et al. IEDB-AR: Immune epitope database-analysis resource in 2019. Nucleic Acids Res. 2019, 47, W502–W506. [Google Scholar] [CrossRef] [Green Version]
- Farrell, D.; Jones, G.; Pirson, C.; Malone, K.; Rue-Albrecht, K.; Chubb, A.J.; Vordermeier, M.; Gordon, S.V. Integrated computational prediction and experimental validation identifies promiscuous T cell epitopes in the proteome of Mycobacterium bovis. Microb. Genom. 2016, 2, e000071. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.D.; Auffret, M.D.; Warr, A.; Wiser, A.H.; Press, M.O.; Langford, K.W.; Lichko, I.; Snelling, T.J.; Dewhurst, R.J.; Walker, A.W.; et al. Assembly of 913 microbial genomes from metagenomic sequencing of the cow rumen. Nat. Commun. 2018, 9, 870. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef]
- Pierleoni, A.; Martelli, P.; Casadio, R. PredGPI: A GPI-anchor predictor. BMC Bioinform. 2008, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Xu, J. RaptorX: Exploiting structure information for protein alignment by statistical inference. Proteins 2011, 79 (Suppl. S10), 161–171. [Google Scholar] [CrossRef] [Green Version]
- Suhrbier, A. Multi-epitope DNA vaccines. Immunol. Cell Biol. 1997, 75, 402–408. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Wang, G.; Pang, A.; Fu, B.; Yin, H.; Zhang, D. Increased survival time in mice vaccinated with a branched lysine multiple antigenic peptide containing B- and T-cell epitopes from T. gondii antigens. Vaccine 2011, 29, 8619–8623. [Google Scholar] [CrossRef]
- Choudhury, S.; Ganguly, A.; Chakrabarti, K.; Sharma, R.K.; Gupta, S.K. DNA vaccine encoding chimeric protein encompassing epitopes of human ZP3 and ZP4: Immunogenicity and characterization of antibodies. J. Reprod. Immunol. 2009, 79, 137–147. [Google Scholar] [CrossRef]
- Da’Dara, A.A.; Li, C.; Yu, X.; Zheng, M.; Zhou, J.; Shollenberger, L.M.; Li, Y.S.; Harn, D.A. Prime-boost vaccine regimen for SjTPI and SjC23schistosome vaccines, increase efficacy in water buffalo in a field trial in China. Front. Immunol. 2019, 10, 284. [Google Scholar] [CrossRef]
- Tollefsen, S.; Vordermeier, M.; Olsen, I.; Storset, A.K.; Reitan, L.J.; Clifford, D.; Lowrie, D.B.; Wiker, H.G.; Huygen, K.; Hewison, G.; et al. DNA injection in combination with electroporation: A novel method for vaccination of farmed ruminants. Scand. J. Immunol. 2003, 57, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Integrated DNA Technologies IDT—Codon Optimization Tool. Available online: https://eu.idtdna.com/CodonOpt (accessed on 20 November 2019).
- Stothard, P. Sequence Manipulation Suite (SMS): Reverse Translate. Available online: http://www.bioinformatics.org/sms2/rev_trans.html (accessed on 20 November 2019).
- Coustou, V.; Plazolles, N.; Guegan, F.; Baltz, T. Sialidases play a key role in infection and anaemia in Trypanosoma congolense animal trypanosomiasis. Cell. Microbiol. 2012, 14, 431–445. [Google Scholar] [CrossRef]
- Bragazzi, N.L.; Gianfredi, V.; Villarini, M.; Rosselli, R.; Nasr, A.; Hussein, A.; Martini, M.; Behzadifar, M. Vaccines meet big data: State-of-the-art and future prospects. From the classical 3Is (“isolate-inactivate-inject”) Vaccinology 1.0 to Vaccinology 3.0, vaccinomics, and beyond: A historical overview. Front. Public Health 2018, 6, 62. [Google Scholar] [CrossRef]
- Saadi, M.; Karkhah, A.; Nouri, H.R. Development of a multi-epitope peptide vaccine inducing robust T cell responses against brucellosis using immunoinformatics based approaches. Infect. Genet. Evol. 2017, 51, 227–234. [Google Scholar] [CrossRef]
- Maritz-Olivier, C.; Van Zyl, W.; Stutzer, C. A systematic, functional genomics, and reverse vaccinology approach to the identification of vaccine candidates in the cattle tick, Rhipicephalus microplus. Ticks Tick Borne Dis. 2012, 3, 179–187. [Google Scholar] [CrossRef]
- Kar, P.P.; Srivastava, A. Immuno-informatics analysis to identify novel vaccine candidates and design of a multi-epitope based vaccine candidate against Theileria parasites. Front. Immunol. 2018, 9, 2213. [Google Scholar] [CrossRef]
- Guedes, R.L.M.; Rodrigues, C.M.F.; Coatnoan, N.; Cosson, A.; Cadioli, F.A.; Garcia, H.A.; Gerber, A.L.; Machado, R.Z.; Minoprio, P.M.C.; Teixeira, M.M.G.; et al. A comparative in silico linear B-cell epitope prediction and characterization for South American and African Trypanosoma vivax strains. Genomics 2019, 111, 407–417. [Google Scholar] [CrossRef]
- Pandya, M.; Rasmussen, M.; Hansen, A.; Nielsen, M.; Buus, S.; Golde, W.; Barlow, J. A modern approach for epitope prediction: Identification of foot-and-mouth disease virus peptides binding bovine leukocyte antigen (BoLA) class I molecules. Immunogenetics 2015, 67, 691–703. [Google Scholar] [CrossRef]
- Nielsen, M.; Connelley, T.; Ternette, N. Improved prediction of bovine leucocyte antigens (BoLA) presented ligands by use of mass-spectrometry-determined ligand and in vitro binding data. J. Proteome Res. 2018, 17, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Zikova, A.; Schnaufer, A.; Dalley, R.A.; Panigrahi, A.K.; Stuart, K.D. The F(0)F(1)-ATP synthase complex contains novel subunits and is essential for procyclic Trypanosoma brucei. PLoS Pathog. 2009, 5, e1000436. [Google Scholar] [CrossRef] [Green Version]
- Simwango, M.; Ngonyoka, A.; Nnko, H.J.; Salekwa, L.P.; Ole-Neselle, M.; Kimera, S.I.; Gwakisa, P.S. Molecular prevalence of trypanosome infections in cattle and tsetse flies in the Maasai Steppe, northern Tanzania. Parasites Vectors 2017, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Dagenais, T.R.; Demick, K.P.; Bangs, J.D.; Forest, K.T.; Paulnock, D.M.; Mansfield, J.M. T-cell responses to the trypanosome variant surface glycoprotein are not limited to hypervariable subregions. Infect. Immun. 2009, 77, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Magez, S.; Radwanska, M.; Drennan, M.; Fick, L.; Baral, T.N.; Brombacher, F.; Baetselier, P.D. Interferon-γ and nitric oxide in combination with antibodies are key protective host immune factors during Trypanosoma congolense Tc13 infections. J. Infect. Dis. 2006, 193, 1575–1583. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, J.M.; Paulnock, D.M.; Hedberg, G.M. Bridging Innate and Adaptive Immunity in African Trypanosomiasis. In Trypanosomes and Trypanosomiasis; Springer-Nature: London, UK, 2013; pp. 89–114. [Google Scholar]
- Liu, G.; Sun, D.; Wu, H.; Zhang, M.; Huan, H.; Xu, J.; Zhang, X.; Zhou, H.; Shi, M. Distinct contributions of CD4+ and CD8+ T cells to pathogenesis of Trypanosoma brucei infection in the context of gamma interferon and interleukin-10. Infect. Immun. 2015, 83, 2785–2795. [Google Scholar] [CrossRef] [Green Version]
- Onah, D.N.; Hopkins, J.; Luckins, A.G. Effects of the depletion of CD8+ T cells and monocytes on the proliferative responses of peripheral blood leucocytes from Trypanosoma evansi-infected sheep. Vet. Parasitol. 2000, 92, 25–35. [Google Scholar] [CrossRef]
- Flynn, J.N.; Sileghem, M.; Williams, D.J. Parasite-specific T-cell responses of trypanotolerant and trypanosusceptible cattle during infection with Trypanosoma congolense. Immunology 1992, 75, 639–645. [Google Scholar]
- Radwanska, M.; Guirnalda, P.; De Trez, C.; Ryffel, B.; Black, S.; Magez, S. Trypanosomiasis-induced B cell apoptosis results in loss of protective anti-parasite antibody responses and abolishment of vaccine-induced memory responses. PLoS Pathog. 2008, 4, e1000078. [Google Scholar] [CrossRef] [Green Version]
- Horn, D. Antigenic variation in African trypanosomes. Mol. Biochem. Parasitol. 2017, 195, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Villarreal, D.O.; Talbott, K.T.; Choo, D.K.; Shedlock, D.J.; Weiner, D.B. Synthetic DNA vaccine strategies against persistent viral infections. Expert Rev. Vaccines 2013, 12, 537–554. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Xiang, P.; Li, Q. Investigations of the effect of DNA size in transient transfection assay using dual luciferase system. Anal. Biochem. 2005, 346, 289–294. [Google Scholar] [CrossRef]
- Jazayeri, S.D.; Poh, C.L. Recent advances in delivery of veterinary DNA vaccines against avian pathogens. Vet. Res. 2019, 50, 78. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Liu, H.; Zhang, K.; Xue, Q.; Chen, K.; Xue, Y. Minimum dose, antigen content, and immunization duration of a trivalent vaccine of inactivated Haemophilus parasuis serovars 4, 5, and 12 against Glässer’s disease in pigs. Can. J. Vet. Res. 2016, 80, 287–293. [Google Scholar]
- Tabel, H.; Wei, G.; Bull, H.J. Immunosuppression: Cause for failures of vaccines against African trypanosomiases. PLoS Negl. Trop. Dis. 2013, 7, e2090. [Google Scholar] [CrossRef] [Green Version]
- Young, J.L.; Dean, D.A. Electroporation-mediated gene delivery. Adv. Genet. 2015, 89, 49–88. [Google Scholar]




{kind=link}
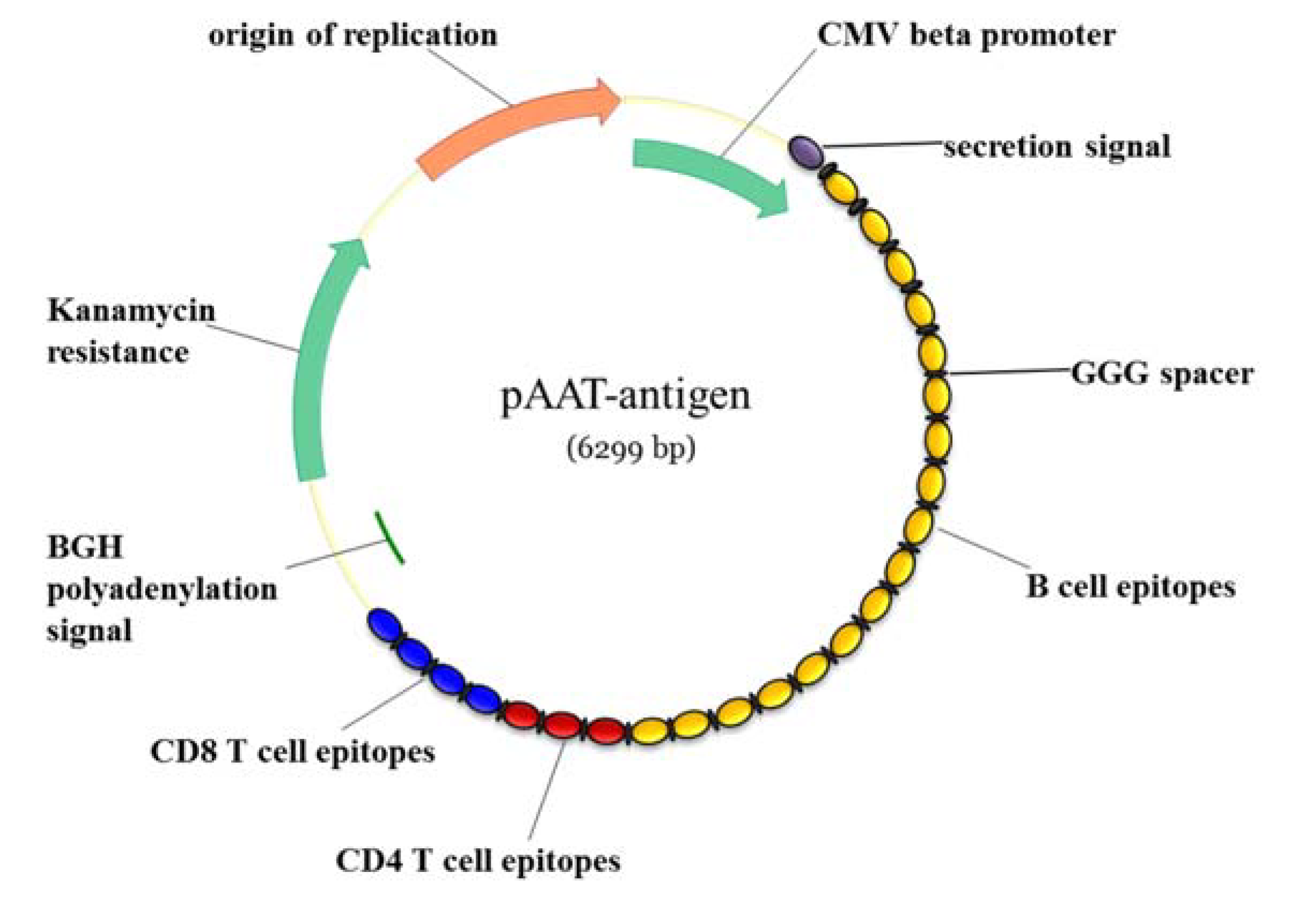{kind=link}
{kind=link}
| Epitope | Length | Antigen 1 | Annotation 1 | TargetP 2 | Bos taurus Hit 3 | Id (%) 4 | Bos indicus Hit 3 | Id (%) 4 |
|---|---|---|---|---|---|---|---|---|
| MQYGSTPKDIRYGIE | 15 | TcIL3000.11.15250.1 | chaperonin HSP60, mitochondrial precursor | Other (5) | NP_001029859.1 | 28.6 | XP_019842974.1 | 31.6 |
| GFTSPYFVTNTKSQKC | 16 | TcIL3000.11.15250.1 | chaperonin HSP60, mitochondrial precursor | Other (5) | AAI02078.1 | 56.3 | XP_019829938.1 | 56.3 |
| VFTGAQMISEDLGLSLDQS | 19 | TcIL3000.11.15250.1 | chaperonin HSP60, mitochondrial precursor | Other (5) | XP_024847167.1 | 23.3 | XP_019820980.1 | 47.6 |
| TISRDECILMEGGGSAIAVE ERVQMIRDMIAAEDHEYNRERL | 42 | TcIL3000.11.15250.1 | chaperonin HSP60, mitochondrial precursor | Other (5) | XP_024847167.1 | 23.3 | XP_019820980.1 | 47.6 |
| RIEDRGLEDKEKREGLN | 17 | TcIL3000.11.15250.1 | chaperonin HSP60, mitochondrial precursor | Other (5) | XP_024847167.1 | 23.3 | XP_019811187.1 | 47.1 |
| MRKRVNESQAPLPAL | 15 | TcIL3000.11.15250.1 | chaperonin HSP60, mitochondrial precursor | Other (5) | NP_001029859.1 | 28.6 | XP_019842974.1 | 31.6 |
| KCIYYVTGDSKKKLETSPFIEQAKRRG | 27 | TcIL3000_0_26140.1 | Heat shock protein 83, putative | Other (1) | NP_001073105.1 | 48.1 | CCA61548.1 | 48.1 |
| EYIPRAFPVKSTTGL | 15 | TcIL3000_10_3450.1 | hypothetical protein, conserved | M (4) | XP_024851069.1 | 65.4 | XP_019820927.1 | 65.4 |
| ECQDDWCTLEKRYFW | 15 | TcIL3000_10_3450.1 | hypothetical protein, conserved | M (4) | XP_024851069.1 | 65.4 | XP_019820927.1 | 65.4 |
| LETAQKWNEWRFWELDPEQVKKVAREDQNIGR EGLGYNSPWEQVVREDFDKRKKALTQEEMAE LKRMDTERMARETAAYKERKDRIRDDLEKARG | 95 | TcIL3000_10_3450.1 | hypothetical protein, conserved | M (4) | NP_001029859.1 | 28.6 | XP_019842974.1 | 31.6 |
| KFLNKRFAVDKDLQRMQPGKRYSGKTAD | 28 | TcIL3000_10_3450.1 | hypothetical protein, conserved | M (4) | NP_001029859.1 | 28.6 | XP_019834515.1 | 28.6 |
| PFGRFAHTPTVLPDSSIDLSYEVPWW | 26 | TcIL3000_10_510.1 | hypothetical protein, conserved | Other (3) | NP_001029859.1 | 28.6 | XP_019842974.1 | 31.6 |
| DSLCTLNMVPGVSVYGEKRVEVGATQ | 26 | TcIL3000_10_6460.1 | fibrillarin, putative | Other (3) | XP_024851069.1 | 65.4 | XP_019820927.1 | 65.4 |
| TGQFEKAGEQKEREGKH | 17 | TcIL3000_10_970.1 | intraflagellar transport protein 172, putative | M (5) | XP_024847167.1 | 23.3 | XP_019820980.1 | 47.6 |
| DFDCTDFPKKYPMPKSSS | 18 | TcIL3000_10_970.1 | intraflagellar transport protein 172, putative | M (5) | - | - | - | - |
| ADDGYVGYDSVPFHRYNR | 18 | TcIL3000_7_2400.1 | hypothetical protein, conserved | M (4) | - | - | - | - |
| LGCATCKMPNDINEA | 15 | TcIL3000_7_510.1 | hypothetical protein, conserved | Other (3) | NP_001029859.1 | 28.6 | XP_019842974.1 | 31.6 |
| IIKGCNVFELDGSMSDVHQSI | 21 | TcIL3000_7_510.1 | hypothetical protein, conserved | Other (3) | AAI02078.1 | 56.3 | XP_019829938.1 | 56.3 |
| TSAAGLSPDWLEAFFTNVAYNT | 22 | TcIL3000_7_510.1 | hypothetical protein, conserved | Other (3) | XP_024847167.1 | 23.3 | XP_019820980.1 | 47.6 |
| KSLRMNFFTVCERCVLKEM | 19 | TcIL3000_7_510.1 | hypothetical protein, conserved | Other (3) | NP_001029859.1 | 28.6 | XP_019842974.1 | 31.6 |
| EKGPSTLPPLEHLFVASVYLAAQRQFTNLFFF | 32 | TcIL3000_7_510.1 | hypothetical protein, conserved | Other (3) | XP_024851069.1 | 65.4 | XP_019820927.1 | 65.4 |
| DLKEWCKQYEQLTNDMLRLR | 20 | TcIL3000_7_510.1 | hypothetical protein, conserved | Other (3) | - | - | - | - |
| ELPGAITHKSILELR | 15 | TcIL3000_8_4670.1 | hypothetical protein, conserved | Other (2) | XP_024851069.1 | 65.4 | XP_019820927.1 | 65.4 |
| QSQQQLLASTRGGMPAR | 17 | TcIL3000_8_4670.1 | hypothetical protein, conserved | Other (2) | XP_024847167.1 | 23.3 | XP_019816904.1 | 23.3 |
| SGVRHLRMAGDGTVGQN | 17 | TcIL3000_9_6470.1 | hypothetical protein, conserved | M (5) | XP_024847167.1 | 23.3 | XP_019820980.1 | 47.6 |
| DWLERQFIDNCATPERDP | 18 | TcIL3000_9_6470.1 | hypothetical protein, conserved | M (5) | XP_024851069.1 | 65.4 | XP_019820927.1 | 65.4 |
| LLDWEDFGIPREDLYR | 16 | TcIL3000_9_6470.1 | hypothetical protein, conserved | M (5) | NP_001029859.1 | 28.6 | XP_019842974.1 | 31.6 |
| QQTRFDNTTEEKLRSL TYTQTDKTVDEYYG | 30 | TevSTIB805.5.5170-t26_1 | hypothetical protein, conserved | S (3) | XP_024847167.1 | 23.3 | XP_019816904.1 | 23.3 |
| FPDPPFNFSAVVPER | 15 | TevSTIB805.5.5170-t26_1 | hypothetical protein, conserved | S (3) | XP_024851069.1 | 65.4 | XP_019820927.1 | 65.4 |
| SGVKANDTGVGPNTTNTAGGA | 21 | TevSTIB805.5.5170-t26_1 | hypothetical protein, conserved | S (3) | XP_024847167.1 | 23.3 | XP_019820980.1 | 47.6 |
| CVDIIDWRDLDEMLNNRTDEVVEKSL | 26 | TevSTIB805.5.5170-t26_1 | hypothetical protein, conserved | S (3) | - | - | - | - |
| Epitope | Antigen 1 | Annotation 1 | HLAs (human) 2 | Bos taurus Hit 3 | Id (%) 4 | Bos indicus Hit3 | Id (%) 4 |
|---|---|---|---|---|---|---|---|
| AQIHDIYRMIPPLQVVLVSAT | TcIL3000.11.9080.1 | ATP-dependent RNA helicase FAL1, putative | HLA-DRB3*02:02 | NP_001039653 | 66.7 | XP_019837752 | 66.7 |
| FKAQIHDIYRMIPPLQVVLVS | TcIL3000.11.9080.1 | ATP-dependent RNA helicase FAL1, putative | HLA-DRB1*09:01 | NP_001039653 | 66.7 | XP_019837752 | 66.7 |
| GLVIERRLSDKHFVF | TcIL3000_0_22360.1 | NADH-dependent fumarate reductase | HLA-DRB3*01:01 | 3FE5_A | 46.7 | XP_019825021 | 46.7 |
| RINLVVQFDMASDADSYLHRV | TcIL3000_10_400.1 | ATP-dependent RNA helicase SUB2, putative | HLA-DRB3*01:01 | NP_001029924 | 57.1 | XP_019819542 | 57.1 |
| DGRIGIILMDNITEVQSGQK | TcIL3000_7_6050.1 | ATP synthase F1, alpha subunit, putative | HLA-DRB3*01:01 | XP_005203166 | 40.0 | XP_019836600 | 40.0 |
| AKNKFFYMYVQELNYLIRF | TcIL3000_7_6050.1 | ATP synthase F1, alpha subunit, putative | HLA-DPA1*02:01 _DPB1*14:01 | XP_005205520 | 36.8 | XP_019814319 | 36.8 |
| ASYANVWEMDDPYRFLQTEQD | TcIL3000_9_4570.1 | Chromosome passenger complex (CPC) protein INCENP N terminal, putative | HLA-DRB3*01:01 | DAA31134 | 38.1 | XP_019813076 | 38.1 |
| DYTNRIIRQMLHNVAALSCNK | TevSTIB805.10.4720-t26_1 | Pumillo RNA binding protein PUF1 | HLA-DRB4*01:01 | XP_002697522 | 38.1 | XP_019840994 | 38.1 |
| Epitope | Antigen 1 | Annotation 1 | BoLA 2 | Bos taurus Hit 3 | Id (%) 4 | Bos indicus Hit 3 | Id (%) 4 |
|---|---|---|---|---|---|---|---|
| MMHAYASRY | TcIL3000.11.1250.1 | NADH dehydrogenase [ubiquinone] iron -sulfur protein 7, mitochondrial | BoLA-N:05501, BoLA-N:02401 | DAA15150.1 | 66.7 | XP_019842744.1 | 66.7 |
| YPAVRVYPV | TcIL3000.11.9220.1 | NADH dehydrogenase subunit NB6M, putative | BoLA-N:00501 | DAA32829.1 | 66.7 | XP_019825300.1 | 66.7 |
| FGPWFVEPV | TcIL3000_0_05230.1 | proteasome beta 3 subunit, putative | BoLA-N:05301, BoLA-N:00402, BoLA-N:00401, BoLA-N:00102, BoLA-N:00103, BoLA-N:00101 | NP_789861.1 | 55.6 | XP_019823249.1 | 55.6 |
| VTPSIHYTM | TcIL3000_0_22360.1 | NADH-dependent fumarate reductase | BoLA-N:05301, BoLA-N:03801, BoLA-N:00402, BoLA-N:00401 | XP_024843166.1 | 66.7 | XP_019834535.1 | 66.7 |
| ATFEVFHTI | TcIL3000_10_13200.1 | methyltransferase domain containing protein, putative | BoLA-N:05401 | XP_005225549.2 | 66.7 | XP_019808611.1 | 66.7 |
| LQYHKYGCL | TcIL3000_10_3580.1 | ubiquinol-cytochrome c reductase complex 14kD subunit, putative | BoLA-HD6 | NP_001179015.1 | 66.7 | XP_019841558.1 | 66.7 |
| YGPSWHCVM | TcIL3000_10_3900.1 | dynein light chain, putative | BoLA-N:05301, BoLA-N:04001, BoLA-N:00402, BoLA-N:00401, BoLA-N:00102, BoLA-N:00103, BoLA-N:00101 | XP_015328853.2 | 55.6 | XP_019837337.1 | 55.6 |
| DMYLHQHEF | TcIL3000_10_970.1 | intraflagellar transport protein 172, putative | BoLA-NC1:00101, BoLA-NC1:00201, BoLA-NC1:00301 | XP_002693758.2 | 55.6 | XP_019830009.1 | 55.6 |
| EEMKYVAGL | TcIL3000_10_970.1 | intraflagellar transport protein 172, putative | BoLA-N:04101 | NP_001192842.1 | 66.7 | XP_019822633.1 | 66.7 |
| RKYETTWEM | TcIL3000_6_4030.1 | hypothetical protein, conserved | BoLA-D18.4 | AAI42017.1 | 55.6 | XP_019813499.1 | 55.6 |
| EHNWMFAEI | TcIL3000_9_4570.1 | Chromosome passenger complex (CPC) protein INCENP N terminal, putative | BoLA-N:05001 | XP_024855981.1 | 55.6 | XP_019826963.1 | 55.6 |
| SEMRAWYWK | TcIL3000_9_4570.1 | Chromosome passenger complex (CPC) protein INCENP N terminal, putative | BoLA-N:04201 | NP_001095539.1 | 55.6 | XP_019817650.1 | 66.7 |
| AMWSQDSPY | TcIL3000_9_6470.1 | hypothetical protein, conserved | BoLA-N:00901 | NP_001193092.1 | 66.7 | XP_019825232.1 | 66.7 |
| ARYEYFLAY | TevSTIB805.10.7310-t26_1 | Sterol methyltransferase, putative | BoLA-N:02201 | XP_005212827.1 | 55.6 | XP_019825865.1 | 55.6 |
| THETHSFLY | TvY486_1100540:mRNA | dynein light chain, putative | BoLA-N:04801 | XP_024852897.1 | 66.7 | XP_019822516.1 | 66.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michel-Todó, L.; Bigey, P.; Reche, P.A.; Pinazo, M.-J.; Gascón, J.; Alonso-Padilla, J. Design of an Epitope-Based Vaccine Ensemble for Animal Trypanosomiasis by Computational Methods. Vaccines 2020, 8, 130. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010130
Michel-Todó L, Bigey P, Reche PA, Pinazo M-J, Gascón J, Alonso-Padilla J. Design of an Epitope-Based Vaccine Ensemble for Animal Trypanosomiasis by Computational Methods. Vaccines. 2020; 8(1):130. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010130
Chicago/Turabian StyleMichel-Todó, Lucas, Pascal Bigey, Pedro A Reche, María-Jesus Pinazo, Joaquim Gascón, and Julio Alonso-Padilla. 2020. "Design of an Epitope-Based Vaccine Ensemble for Animal Trypanosomiasis by Computational Methods" Vaccines 8, no. 1: 130. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010130