1. Introduction
West Nile virus (WNV) is an arthropod borne flavivirus that mainly circulates between bird populations and its natural mosquito host,
Culex spp [
1,
2,
3]. Occasionally, a virus can spill over and cause infections in humans, an inadvertent host. Although mostly asymptomatic, WNV infections can cause a range of symptoms in humans, from mild febrile illness to more severe diseases such as paralysis and meningitis [
4]. In 1999, WNV caused a major outbreak of fever and encephalitis in New York City. This particularly virulent strain of WNV, named WNV
NY99, caused an unusually high rate of neurological symptoms with 63% of the patients developing encephalitis and a 12% mortality rate [
5,
6]. Apart from the occasional human outbreaks, horses are known to incur serious WNV infections, representing 96.9% of all mammalian cases caused by WNV [
7,
8,
9]. Like humans, horses are dead-end hosts, as the viremia is not sufficient to sustain transmission to mosquitoes [
10]. Several vaccines have been developed and licensed for equine use, but so far there are still none licensed for use in humans [
11]. It is crucial for a vaccine to be both safe and highly effective. One of the major concerns about sub-unit and inactivated vaccines is low immunogenicity, which usually has to be complemented with a strong adjuvant to induce the required antibody response and usually requires frequent re-vaccinations. On multiple occasions, this has been linked to unwanted allergic reactions [
12]. Live-attenuated vaccines are highly effective and, in most cases, eliminate the need for an adjuvant. However, these bring higher risk of the virus reverting to virulence, thus making them inappropriate for use in humans who are immunocompromised [
13,
14,
15].
Previously, we reported the generation of BinJ/WNV
KUN-prME, a chimeric flavivirus that encodes the structural prME genes of WNV
KUN on the genetic backbone of the insect-specific flavivirus (ISF) Binjari virus (BinJV) nonstructural protein genes [
16]. During vertebrate infection, the flavivirus envelope (E) proteins engage with cellular receptors leading to virus internalization and replication. To prevent this, virus neutralization by antibodies directed to the EDIII receptor-binding domain of the virus is one of the requirements for the host to be protected [
17,
18,
19]. We previously demonstrated that BinJ/WNV
KUN-prME authentically presents all E protein epitopes, including those in EDIII, when compared to the wildtype WNV
KUN. BinJ/WNV
KUN-prME chimera can be produced to high titers in insect cells but exhibits an insect-specific phenotype and is unable to replicate in vertebrate cells. This provides a critical element of safety in the context of its assessment as a vaccine. Unlike previously reported chimeric flavivirus vaccines based on YFV or DENV backbones, the inability of the BinJ/WNV-prME chimeric virus to replicate in vaccinated individuals, eliminates any risk of reversion to virulence and thus would be more suitable for use in immunocompromised people and pregnant women.
Here, we report the assessment of immunogenicity and efficacy of BinJ/WNVKUN-prME as a novel WNV vaccine candidate and demonstrate protection of mice against lethal challenge with the virulent WNVNY99 strain. In addition, we show that further inactivation treatment of this vaccine does not adversely influence epitope presentation or protection in vivo.
2. Materials and Methods
2.1. Animal Ethics Statement
All animal work was carried out in accordance with the “Australian Code for the Care and Use of Animals for Scientific Purposes” as defined by the National Health and Medical Research Council of Australia. All experiments had received approval by the University of Queensland Animal Ethics Committee (permits SCMB/008/18 and SCMB/361/17). Three- to six-week-old CD1 mice were purchased from the Animal Resources Centre, Murdoch, Western Australia.
2.2. Cell Culture
C6/36 (Aedes albopictus) mosquito cells were grown in RPMI 1640 medium supplemented with 5% fetal bovine serum (FBS) at 28 °C. Vero African green monkey (Cercopithecus aethiops) kidney cells were maintained in Dulbecco’s modified Eagle medium (DMEM) with 5% FBS and were incubated at 37 °C with 5% CO2. All media was supplemented with penicillin (50 U/mL), streptomycin (50 μg/mL) and 2 mM L-glutamine (PSG).
2.3. Generation of BinJ/WNVKUN-prME
RNA extraction, Circular Polymerase Extension Reaction and transfection to obtain the BinJ/WNV
KUN-prME chimera was previously described [
16].
2.4. Viruses
BinJV was isolated in our laboratory from Ae. normanensis mosquitoes as previously reported in Hobsen-Peters et al. [
16]. The NY99-4132 strain of WNV was obtained from the Division of Vector-Borne Infectious Diseases, Centers for Diseases Control, Fort Collins, CO, USA [
20]. The KUNV-NSW2011 strain of WNV was isolated in Australia from horse brains at the Elizabeth Macarthur Agriculture Institute, Menangle, New South Wales, Australia [
21].
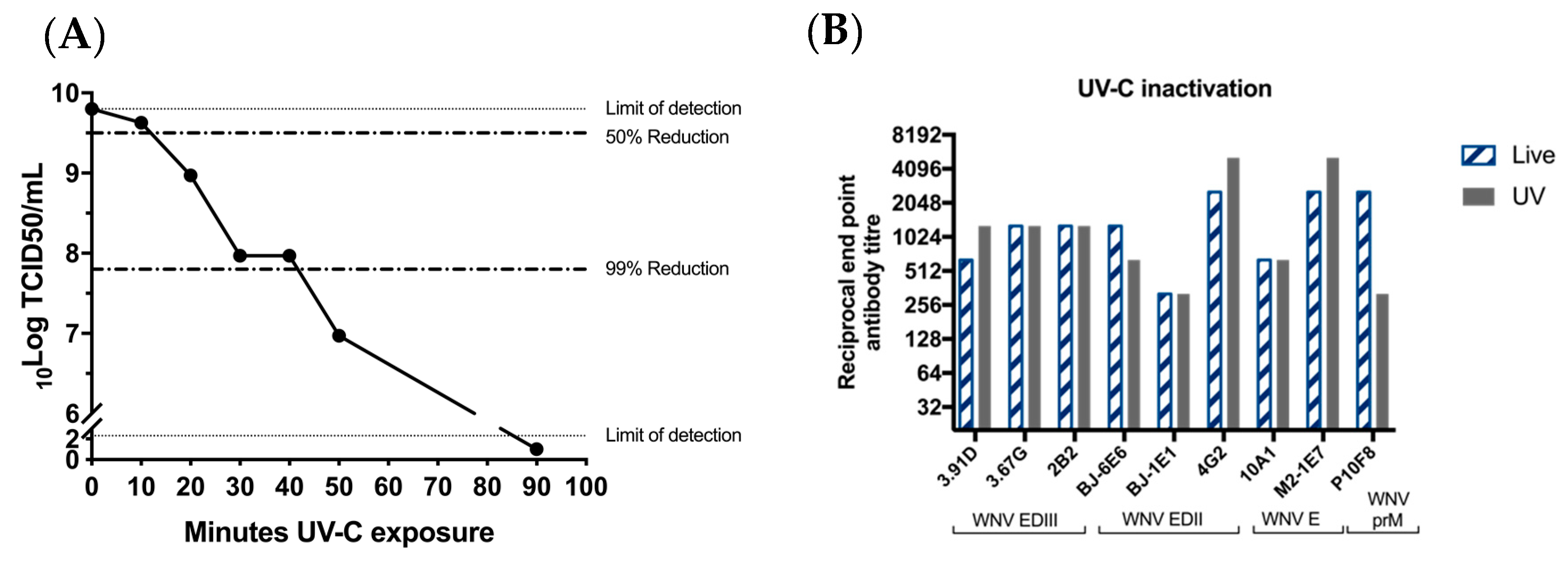
Monoclonal Antibodies
All monoclonal antibodies and their general characteristics that were used in this study have been reported previously [
16]. WNV-specific monoclonal antibodies used were 3.91D, 3.67G, 2B2, 10A1 (anti-WNV E) and P10F8 (anti-WNV-prM). Antibodies BJ-6E6, BJ-1E1, 4G4 were cross-reactive between BinJV and vertebrate-infecting flaviviruses (VIFs). VIF-specific antibodies used in this study were 4G2 and M2-1E7, which were not able to detect BinJV.
2.5. Enzyme-linked Immunosorbent Assay (ELISA)
Plates (detailed in 2.5.1 and 2.5.2) were blocked in 150 μL ELISA blocking buffer [0.05M tris-HCl (pH 8.0), 1 mM EDTA, 0.15 M NaCl, 0.05% (v/v) Tween 20 and 0.2% (w/v) casein] for 1 h at room temperature. Heat-inactivated mouse serum (56 °C/30 min; 1/50 dilution in blocking buffer) was added to first column, titrated out across the plate in doubling dilutions (for total IgG fixed-cell ELISA) or primary monoclonal antibody hybridoma supernatant (coating ELISA;
Section 2.5.2) and incubated at 37 °C with 5% CO
2 for 1 h, with a total volume of 50 μL per well [
16]. After 3 washes with PBS-T, 50 μL/well horseradish peroxidase-labelled goat anti-mouse Ig was added and incubated at 37 °C with CO
2 for 1 h. Five PBS-T washes were performed before adding 100 μL substrate buffer [1 mM 2,2-azino- bis(3-ethylbenzthiazoline-6-sulfonic acid) and 3 mM H
2O
2 in substrate buffer [0.1 M citric acid with 0.2 M Na
2HPO
4 (pH 4.2)]]. Substrate was incubated at room temperature for 1 h in the dark. ODs were measured at 405 nm in spectrophotometer.
2.5.1. Fixed-Cell ELISA Plate Preparation
C6/36 cells were seeded at 104 cells per well in a 96-well plate (Corning Incorporated Costar). The following day, cells were infected with WNVKUN at a multiplicity of infection (MOI) of 0.1 and fixed 5 days post-infection in fixative buffer (20% acetone with 0.02% bovine serum albumin (BSA) in PBS).
2.5.2. Antigen-Coated ELISA on Purified Chimeric Virus
Live or inactivated purified BinJ/WNVKUN-prME virions were diluted in PBS to a final concentration of 80 ng per well. One-hundred microliters per well was added to high-binding 96-well plates (Greiner) and left to bind overnight at 4 °C. The plates were washed 3 times in PBS-T prior to blocking.
2.6. Vaccine Production and Purification
C6/36 cells were pre-seeded in T175 culture flasks and infected with BinJ/WNVKUN-prME a MOI 0.1, 3 days post-seeding. Virus inoculum was prepared in RPMI with 2% FBS, and PSG, and 4 mL were added per flask. Cells were incubated for 1 h at room temperature while rocking. Virus inoculum was then discarded and 20 mL of fresh RPMI media with 2% FBS and PSG were added. Cells were incubated at 28 °C. Virus supernatant was collected at 3 dpi and clarified at 10,000× g for 30 min in a tabletop centrifuge to remove any cell debris. Fresh media was added back onto infected cells and harvesting was repeated every 2 days, for a maximum of 5 harvests. Clarified supernatant was stored at 4 °C until virus purification.
2.6.1. Vaccine Purification
Polyethylene glycol 6000 (40% PEG6000 in NTE) was added to the virus supernatant in a 1:4 ratio. The solution was stirred slowly overnight at 4 °C. Virus-PEG solution was then centrifuged at 12,000× g for 2 h at 4 °C to pellet the virus. The pellet was resuspended in 5 mL cold NTE buffer [120 mM NaCl, 10 mM Tris, 1 mM EDTA (pH 8.0)]. A 40% sucrose cushion was layered under the virus suspension and centrifuged at 28,000 ×g for 2 h at 4 °C. The virus pellet was resuspended in 250 μL NTE buffer and buffer exchanged into PBS using 100 kDa centricon tubes. Purified virus was stored at 4 °C.
2.6.2. Quantification of Vaccine
Purified BinJ/WNV
KUN-prME was run on an unreduced SDS-PAGE along with a pre-made BSA standard curve (0.1, 1, 2, 2.5, 3, 3.5 and 10 mg/mL). Band intensity was determined using Fiji-ImageJ software and yield of E monomer was resolved by comparison to BSA standard [
16].
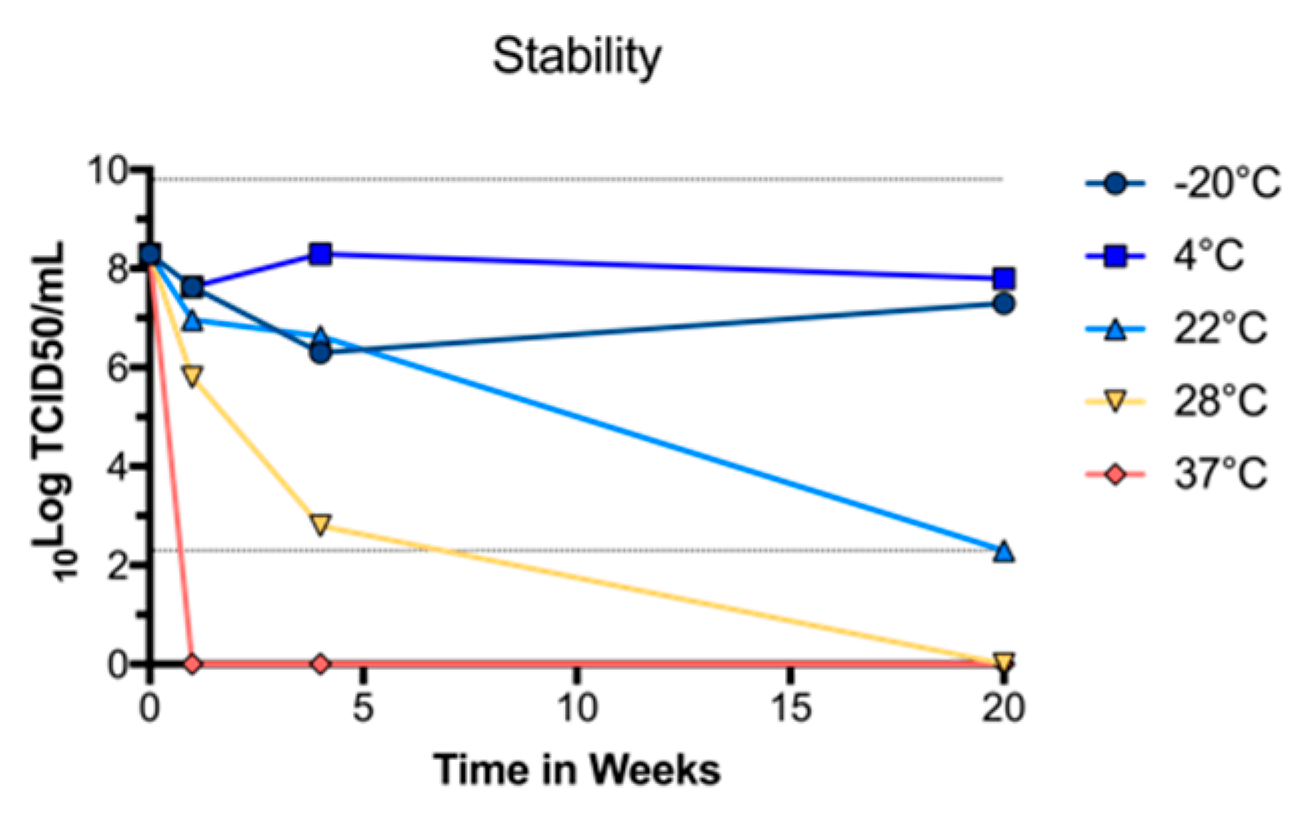
2.7. UV Inactivation of BinJ/WNVKUN-prME Vaccine
To determine optimal UV-C exposure time, purified BinJ/WNV
KUN-prME dose was prepared in PBS in a 6-well plate and placed on ice in the biosafety cabinet. The lid was removed from the plate and the vaccine was exposed to UV-C for different lengths of time. The optimal exposure time was determined by performing a TCID
50 assay to ensure that no infectious virus remained [
20]. Data was extrapolated based on the TCID
50 results to determine the time required for complete inactivation. For vaccine preparation, purified BinJ/WNV
KUN-prME vaccine was exposed to UV-C for 90 min and then stored at 4 °C until time of vaccination. A TCID
50 assay was performed before vaccination to ensure inactivation.
2.8. Micro-Neutralization Assay of Mouse Serum Samples
Micro-neutralization assays were performed using Vero cells. Mouse sera were heated to 56 °C for 30 min to inactivate complement. Serum samples were added in 96-well plates, starting at a 1 in 20 dilution, then titrated out in doubling dilutions. Hundred infectious units of wildtype virus (WNV
NY99 or WNV
KUN) was added per well and the virus-serum mixture incubated for 1 h at 37 °C + 5% CO
2 before adding 10
4 Vero cells per well. The culture plates were incubated for 5 days and then fixed in 20% acetone with 0.02% bovine serum albumin (BSA) in PBS. Neutralizing antibody titers were determined in a fixed-cell ELISA (
Section 2.5 and
Section 2.5.1) using 4G4 monoclonal antibody.
2.9. Vaccination of CD1 Mice with BinJ/WNVKUN-prME
Vaccine doses were prepared in PBS no more than 1 h before vaccination and kept on ice. All vaccines were administered subcutaneously at the base of the tail using a 27-gauge insulin needle, with a maximum volume of 50 μL. Booster doses were given 21 days apart. Animals were monitored after vaccination for potential adverse effects.
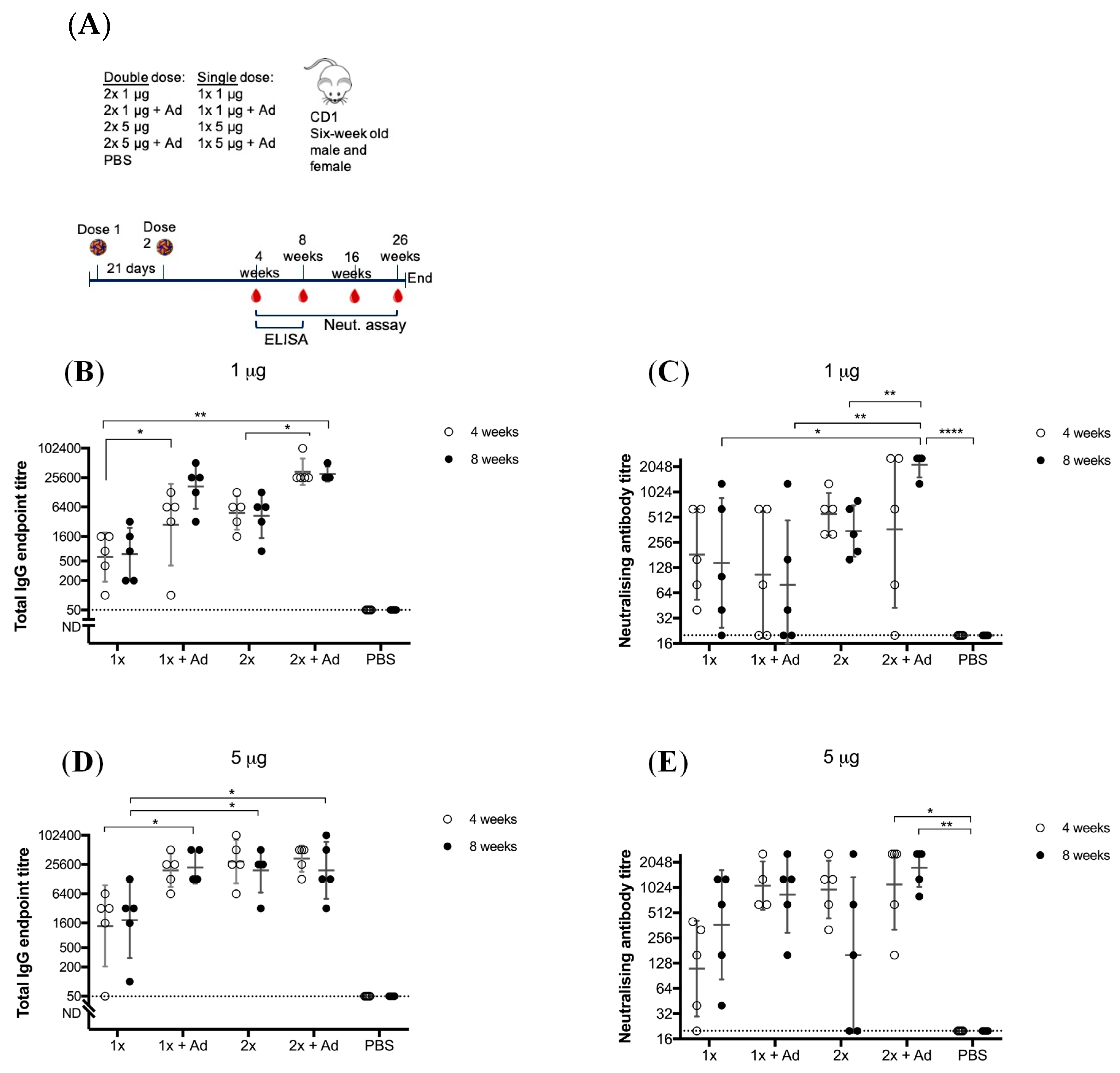
2.9.1. Assessment of Immunogenicity of BinJ/WNVKUN-prME with and without AdvaxTM
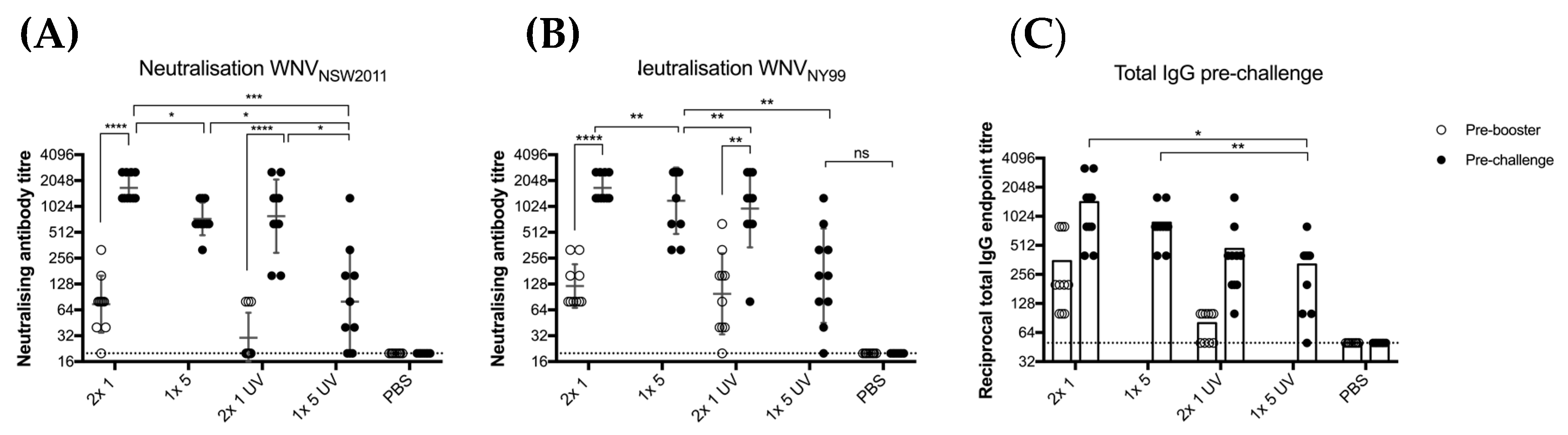
Six-week old male and female CD1 mice were immunized with either 1 or 5 μg BinJ/WNVKUN-prME with half of the mice receiving a booster dose. Vaccination groups included: 1 × 1 μg, 2 × 1 μg, 1 × 5 μg, 2 × 5 μg, each group either with or without AdvaxTM (Vaxine Pty Ltd., Adelaide, Australia) and a non-vaccinated PBS control (n = 5 per group, either 2 males and 3 females or 3 males and 2 females). Adjuvanted doses of BinJ/WNVKUN-prME with AdvaxTM were prepared within 2 h of vaccination by combining the vaccine with AdvaxTM (1mg/mouse) in PBS. Mice were tail bled at 4, 8, 16 and 26 weeks post-last vaccination by making a minor incision in the tail. Blood was collected in BD Microtainer® (Becton, Dickinson & Co., Franklin Lakes, NJ, USA) gel tubes and was allowed to settle for 30 min before serum was separated by centrifugation for 10 min at 11,000 x g. Sera were stored at −20 °C. The majority of male mice were culled prior to week 26 due aggressive behavior. Therefore, only complete data sets are presented in this study for four- and eight-weeks post-vaccination. Antibody titers were determined in fixed cell ELISA (with WNVKUN) and micro-neutralization assays.
2.9.2. Vaccination with Live and UV Inactivated BinJ/WNVKUN-prME
Three-week old male and female CD1 mice were immunized with 1 or 5 μg purified BinJ/WNV
KUN-prME in PBS, either live virus or UV-inactivated. Mice vaccinated with 1 μg received a booster dose after 21 days. Vaccination groups included: 2 × 1 μg dose, 1 × 5 μg dose, 2 × 1 μg dose UV, 1 × 5 μg dose UV and a PBS control group (
n = 10 per group, 5 males and 5 females). Tail bleeds were collected 2 days before booster and virus challenge (
Section 2.9.1).
2.10. Virus Challenge with WNVNY99
WNV
NY99 was propagated in C6/36 cells by infecting a confluent monolayer with MOI 0.01. Virus was incubated for 1 h at 28 °C before removing inoculum and adding fresh RPMI with 2% FBS and PSG. After 5 days, the supernatant was replaced with fresh media to minimize the occurrence of defective interfering particles in the virus stock [
22]. New supernatant was harvested the following day and clarified by centrifugation to remove cell debris. Virus was stored in RPMI with 10% FBS at −80 °C. Virus was titrated using the TCID
50 method on Vero cells and fixed 5 dpi [
23].
2.10.1. WNVNY99 Challenge in CD1 Mice
Mice were infected intraperitoneally with 10
3 infectious units of WNV
NY99 21 days post-vaccination. Mice were monitored twice daily for any clinical signs and weighed once daily. To determine viremia levels, tail bleeds were taken daily, with each individual mouse bled no more than once every three days. Animals were humanely euthanized when weight loss was ≥20% of initial body weight and/or if animal had severe clinical signs, including but not limited to a depressed appearance, abdominal swelling or neurological signs. After virus challenge, animals were culled when reaching a humane endpoint as stipulated by the AEC permit or at the end of the trial on day 21 post-infection. A final cardiac bleed was performed on animals under deep anesthesia (50 mg/kg of ketamine and 10 mg/kg of xylazine). Following cervical dislocation, the head and spines were collected from all animals and fixed in 10% neutral-buffered formaldehyde for 24–36 h, after which the tissues were transferred to 70% ethanol until further processing. Additional tissues (intestines, liver, spleen, kidneys, lung, heart) were also collected and processed from animals experiencing clinical signs of disease. Heads and spines were decalcified in 8% formic acid in water (
v/v) for 4–5 days and then trimmed and subjected to routine processing for paraffin-embedding. Five µm sections were stained with hematoxylin and eosin (H and E) and examined on a Nikon Eclipse 51 E microscope. Digital microphotographs were taken using a Nikon DS-Fi1 camera with a DS-U2 unit and NIS elements F 4.60 software. Images are reproduced without manipulations other than cropping and adjustment of light intensity. Immunohistochemistry for detection of flavivirus NS1 protein in tissues was performed on 4–5 µm FFPE sections as previously described in detail [
24].
2.10.2. Viremia Titrations in Immune-Plaque Assay (IPA)
Blood obtained via tail bleeds were centrifuged to separate serum and frozen at −80 °C. C6/36 cells were pre-seeded at a concentration of 10
5 cells per well in a 96-well plate. Sera were titrated out in RPMI with 2% FBS and PSG in tenfold serial dilutions and transferred onto pre-seeded cells (25 μL per well). Cells were incubated to infect for 2 h at 28 °C, 5% CO
2, before removing the inoculum and adding 200 μL of overlay media (2X M199 medium supplemented with 5% FBS, PSG and 2% carboxymethyl cellulose. Media was removed from the cells 48 h after infection and cells were fixed in 100 μL 80% ice-cold acetone in PBS at −20 °C for 30 min, then air-dried. For probing, plates were blocked with 100 μL KPL milk diluent/blocking solution in PBS-T at 37 °C for 1 h. Fifty microliters of purified hE16 [
25,
26] antibody (1μg/mL) was added and incubated for 1 h at 37 °C to label for WNV E protein. Plates were washed 3 times in PBS-T before adding 50 μL of secondary antibody (IRDye 800CW goat anti-human IgG, LI-COR at 12.5 ng/well) and incubated at 37 °C for 1 h. After five PBS-T washes, plates were air-dried and scanned on Odyssey Clx reader with the following specifications: channel = 800, intensity = auto, resolution = 42 μm and focal length = 3mm. Resulting viral titers are shown as foci forming units per mL (FFU/mL) [
27].
2.11. Statistical Analysis
Statistical analysis was performed using Prism 8 for MacOS (version 8.4.2 (464) 2020). Multiple t-tests were used for analysis of IgG and neutralizing antibody titers. For weight loss comparison, repeated-measures ANOVA analysis of variance was used. Survival curve was analyzed using a Mantel–Cox (log-rank) test.
4. Discussion
Here, we demonstrate that a BinJ/WNV
KUN-prME chimeric vaccine induces high and long-lasting levels of virus neutralizing antibody responses that protect mice against lethal challenge with a virulent WNV strain. Despite being non-infectious for vertebrates, the vaccine showed good efficacy after a single dose in the absence of an adjuvant. This supports our previous studies that showed similar protective efficacy of a BinJ/ZIKV-prME vaccine in a mouse model of ZIKV disease [
16].
We also previously reported that BinJ/WNV
KUN-prME and BinJ/ZIKV-prME particles exhibit authentic epitope presentation compared to the wild type pathogen [
16]. Thus, it is likely that the structural and antigenic integrity of the BinJ/WNV
KUN-prME vaccine is at least partially responsible for the potent protective immunity we have demonstrated here. However, as reported previously, we also hypothesize that these BinJV-chimeric vaccines may have an inherent self-adjuvating capability, possibly caused by initial synthesis of viral replicative dsRNA intermediates upon entry of the vertebrate cell before replication is aborted by innate responses and other mechanisms [
16,
30]. To investigate this further, we showed that after inactivation of the purified BinJ/WNV
KUN-prME vaccine with UV-C, despite significantly lower neutralizing antibody titers compared to untreated vaccine (
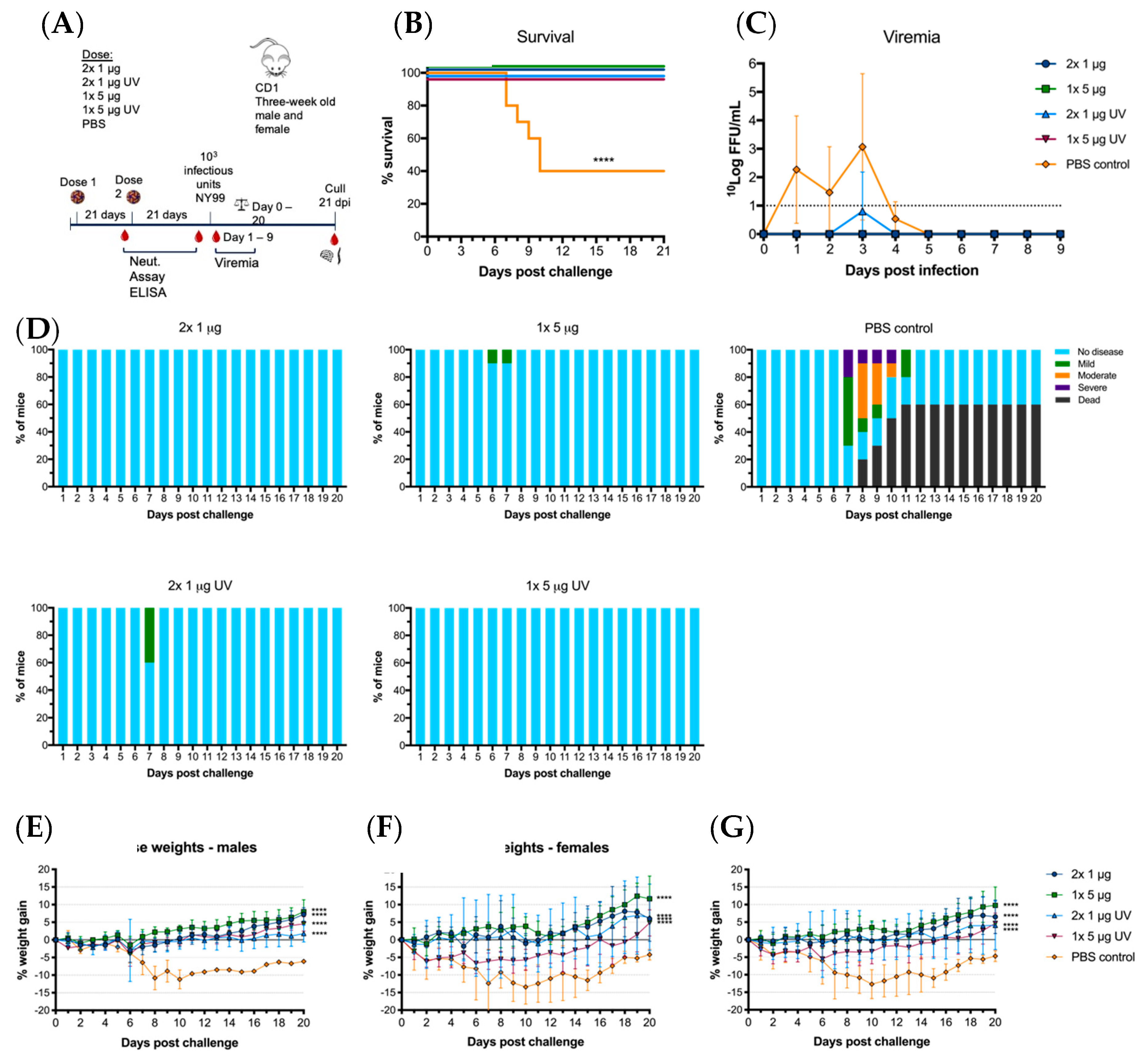
p < 0.0021), immunized mice were still protected from lethal challenge and significant clinical signs of disease. This is consistent with our demonstration that UV inactivation did not significantly alter the epitope presentation. Hence, even though this vaccine is non-infectious to vertebrates, if issues arise in regard to licensing of a GMO-vaccine, there is a viable option for inactivating the chimeric vaccine with little compromise to efficacy.
Despite the robust protective responses induced in the absence of an adjuvant, we did observe a significantly enhanced response in some vaccine groups when the vaccine was formulated with the polysaccharide Advax
TM adjuvant. Advax
TM is a GMP-grade delta-inulin polysaccharide-based adjuvant that has been shown to induce a balanced Th1/Th2 immune response to human vaccines in a number of clinical trials with minimal adverse reactions [
28,
29]. Indeed, we also show that, when formulated with an inactivated Japanese encephalitis virus (JEV) vaccine preparation, Advax
TM induces a robust neutralizing antibody response against JEV in horses and is well tolerated in foals and pregnant mares [
32]. These results suggest that, should further dose sparing be required for a BinJ/WNV
KUN-prME vaccine, Advax
TM provides a suitable adjuvant formulation for further studies.
Our findings also warrant further comparisons of safety and efficacy of the BinJ/WNV
KUN-prME vaccine with other WNV vaccines that have been approved for veterinary use or have been assessed as candidates for a human WNV vaccine in terms of safety and efficacy. Two inactivated vaccines are currently approved for use in horses; Innovator (Pfizer) and Vetera
® WNV (Boehringer Ingelheim). Both require two doses four to six weeks apart with recommended booster doses every 6–12 months. A recombinant canary pox-vectored vaccine (Recomibitek
®, Merial) is also given as two doses four to six weeks apart. To date, no WNV vaccines have been approved for use in humans. However, three candidate vaccines have entered clinical trials. The live-attenuated ChimeriVax-WNV vaccine, which uses the YFV-17D backbone to present the structural WNV prM-E genes, and a similar chimeric vaccine based on the DENV-4 genome backbone (WN-DEN4). Both vaccines were well tolerated and induced significant neutralization titers after a single dose [
11]. A DNA-based, non-infectious vaccine expressing the WNV prM-E proteins has also been assessed in phase I trials. This vaccine was given as three doses and was well tolerated [
33,
34]. It is clear that our results with the BinJV-based chimeric vaccine platform eliminates a number of safety concerns associated with replicating vaccines, thereby providing a distinct advantage over live-attenuated chimeric vaccines. Furthermore, the induction of immunity after a single, unadjuvanted dose provides a favorable initial comparison with existing inactivated and recombinant vaccines.
The ability of this chimeric virus to replicate to high titers in insect cells will be a distinct advantage for large scale manufacturing and efficient vaccine production. Furthermore, the inability of the BinJ/WNVKUN-prME vaccine to replicate in vertebrate cells provides a significant safety aspect to allow propagation of the vaccine in low biocontainment facilities. However, further research is required to generate cell banks of mosquito cell lines for GMP approval. Further studies are also required to investigate whether prior vaccination with a BinJV-based chimera has any effect on efficacy of other BinJV-based vaccines that are subsequently given.
Because BinJ/WNVKUN-prME replicates exceptionally well in mosquito cell culture, we also acknowledge the remote possibility of wild mosquito populations acquiring and subsequently transmitting the vaccine virus. However, preliminary studies have shown that Culex annulirostris, a major mosquito vector of arboviruses in Australasia, cannot transmit BinJ/WNVKUN-prME, even when fed very high doses of the virus in a blood meal. This suggests that there is negligible risk in this regard. We are currently undertaking further studies with other important mosquito species for publication elsewhere.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




