Yersinia pestis Antigen F1 but Not LcrV Induced Humoral and Cellular Immune Responses in Humans Immunized with Live Plague Vaccine—Comparison of Immunoinformatic and Immunological Approaches

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Study Subjects
2.3. Y. pestis Recombinant Antigens
2.4. Isolation of PBMCs, Proliferation Assay, and Cytokine Profiling
2.5. Serum Antibody Detection
2.6. Antibody Isotyping
2.7. In Silico Prediction of Linear and Conformational B-Cell Epitopes for F1 and LcrV
2.8. Allergenicity and Toxicity Prediction
2.9. Identification of F1 Immunoreactive Linear B-Cell Epitopes Using Epitope Mapping
2.10. Statistical Analysis
3. Results
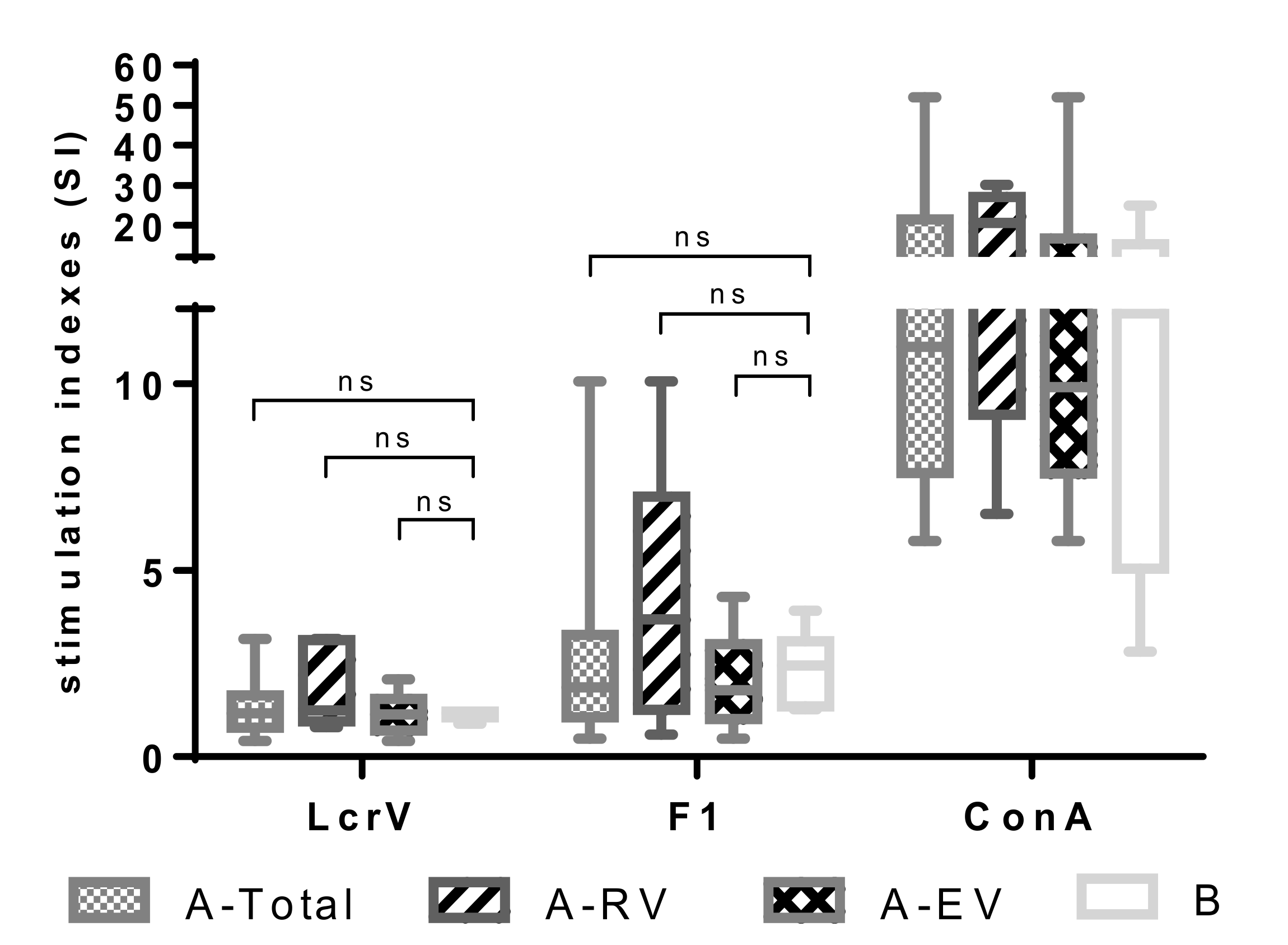
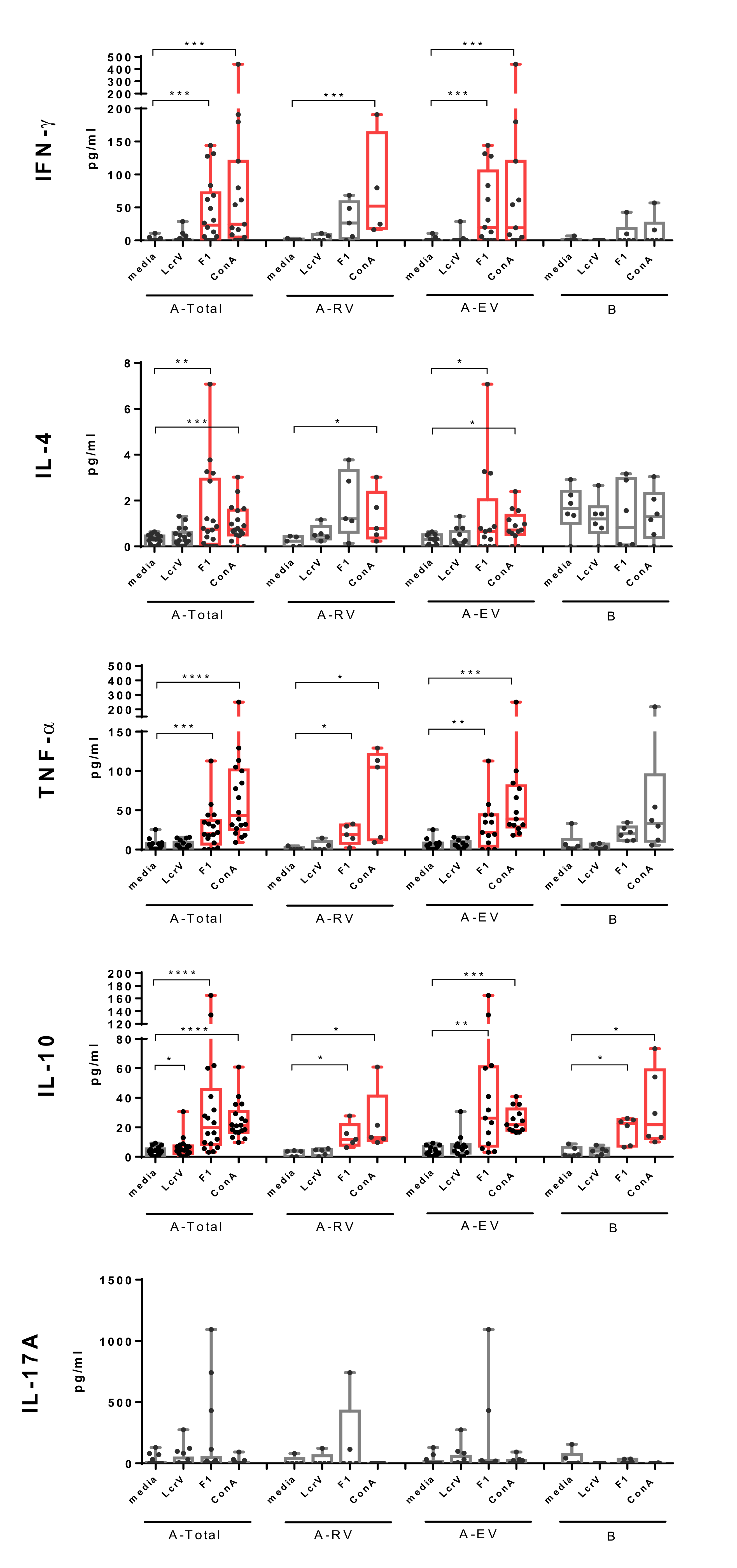
3.1. Proliferative and Cytokine Response to In Vitro Antigen Stimuli
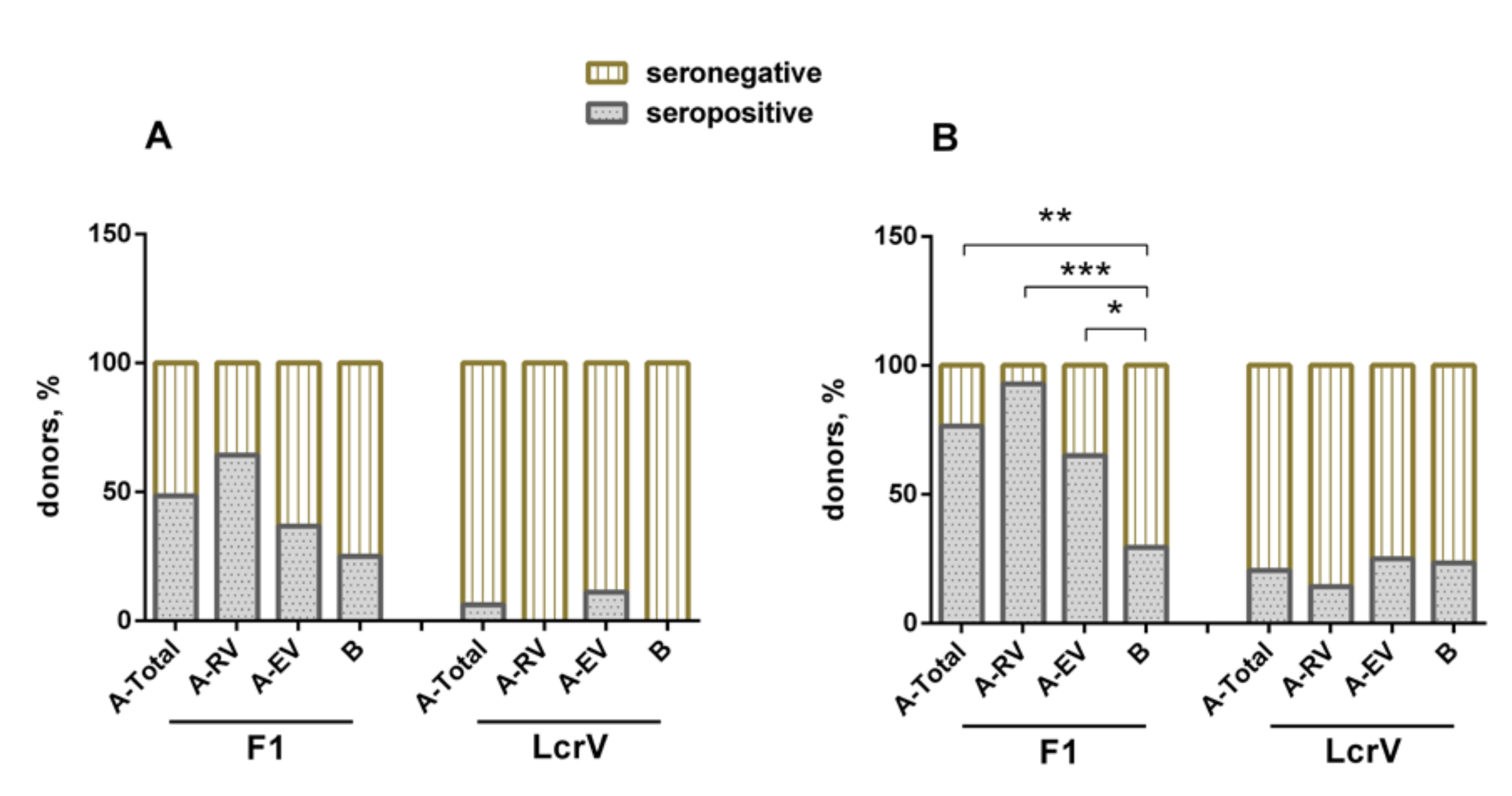
3.2. Humoral Immune Response to F1 and LcrV in Vaccinees
3.3. In Silico Prediction of B-Cell Linear Epitopes of F1 and LcrV
3.4. Epitope Mapping of F1 Antigen with the Use of Overlapping Peptide Library
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delany, I.; Rappuoli, R.; De Gregorio, E. Vaccines for the 21st century. EMBO Mol. Med. 2014, 6, 708–720. [Google Scholar] [CrossRef]
- De Groot, A.S.; Moise, L.; Terry, F.; Gutierrez, A.H.; Hindocha, P.; Richard, G.; Hoft, D.F.; Ross, T.M.; Noe, A.R.; Takahashi, Y.; et al. Better Epitope Discovery, Precision Immune Engineering, and Accelerated Vaccine Design Using Immunoinformatics Tools. Front. Immunol. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed]
- Korber, B.T.; Labute, M.; Yusim, K. Immunoinformatics Comes of Age. PLoS Comput. Biol. 2006, 2, e71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groot, A.S.; Bosma, A.; Chinai, N.; Frost, J.; Jesdale, B.M.; Gonzalez, M.A.; Martin, W.; Saint-Aubin, C. From genome to vaccine: In silico predictions, ex vivo verification. Vaccine 2001, 19, 4385–4395. [Google Scholar] [CrossRef]
- Islam, S.T.; Zaman, S.; Khan, K.; Uddin, M.I.; Chakraborty, S.; Nishat, N.S.; Ibnat, N.; Alam, M.M.; Bhuiyan, T.R.; Qadri, F.; et al. Multi-Epitope Cluster Ep85B within the Mycobacterial Protein Ag85B Elicits Cell-Mediated and Humoral Responses in Mice. Turk. J. Immunol. 2018, 6, 108–117. [Google Scholar] [CrossRef]
- Feodorova, V.A.; Sayapina, L.V.; Corbel, M.J.; Motin, V.L. Russian vaccines against especially dangerous bacterial pathogens. Emerg. Microbes Infect. 2014, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Feodorova, V.A.; Motin, V.L. Plague vaccines: Current developments and future perspectives. Emerg. Microbes Infect. 2012, 1, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, T.B.; Mellado-Sánchez, G.; Jorgensen, A.L.; Moore, S.; Nataro, J.P.; Pasetti, M.F.; Baillie, L. Development of a multiple-antigen protein fusion vaccine candidate that confers protection against Bacillus anthracis and Yersinia pestis. PLoS Negl. Trop. Dis. 2019, 13, e0007644. [Google Scholar] [CrossRef]
- Singh, A.K.; Curtiss, R.; Sun, W. A Recombinant Attenuated Yersinia pseudotuberculosis Vaccine Delivering a Y. pestis YopENt138-LcrV Fusion Elicits Broad Protection against Plague and Yersiniosis in Mice. Infect. Immun. 2019, 87, 87. [Google Scholar] [CrossRef]
- Tao, P.; Mahalingam, M.; Rao, V. Highly Effective Soluble and Bacteriophage T4 Nanoparticle Plague Vaccines Against Yersinia pestis. Breast Cancer 2016, 1403, 499–518. [Google Scholar] [CrossRef] [Green Version]
- Batra, L.; Verma, S.K.; Nagar, D.P.; Saxena, N.; Pathak, P.; Pant, S.C.; Tuteja, U. HSP70 Domain II of Mycobacterium tuberculosis Modulates Immune Response and Protective Potential of F1 and LcrV Antigens of Yersinia pestis in a Mouse Model. PLoS Negl. Trop. Dis. 2014, 8, e3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, R.; Motin, V.L.; Brubaker, R.R. Suppression of cytokines in mice by protein A-V antigen fusion peptide and restoration of synthesis by active immunization. Infect. Immun. 1995, 63, 3021–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, D.G.; Anderson, G.W.; Mauro, J.; Welkos, S.L.; Andrews, G.P.; Adamovicz, J.; Friedlander, A.M. Protection against experimental bubonic and pneumonic plague by a recombinant capsular F1-V antigen fusion protein vaccine. Vaccine 1998, 16, 1131–1137. [Google Scholar] [CrossRef]
- Moore, B.D.; New, R.; Butcher, W.; Mahood, R.; Steward, J.; Bayliss, M.; MacLeod, C.; Bogus, M.; Williamson, E. Dual route vaccination for plague with emergency use applications. Vaccine 2018, 36, 5210–5217. [Google Scholar] [CrossRef]
- Sha, J.; Kirtley, M.L.; Klages, C.; Erova, T.E.; Telepnev, M.; Ponnusamy, D.; Fitts, E.C.; Baze, W.B.; Sivasubramani, S.K.; Lawrence, W.S.; et al. A Replication-Defective Human Type 5 Adenovirus-Based Trivalent Vaccine Confers Complete Protection against Plague in Mice and Nonhuman Primates. Clin. Vaccine Immunol. 2016, 23, 586–600. [Google Scholar] [CrossRef] [Green Version]
- Mizel, S.B.; Graff, A.H.; Sriranganathan, N.; Ervin, S.; Lees, C.J.; Lively, M.O.; Hantgan, R.R.; Thomas, M.J.; Wood, J.; Bell, B. Flagellin-F1-V Fusion Protein Is an Effective Plague Vaccine in Mice and Two Species of Nonhuman Primates. Clin. Vaccine Immunol. 2008, 16, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Zvi, A.; Rotem, S.; Zauberman, A.; Elia, U.; Aftalion, M.; Bar-Haim, E.; Mamroud, E.; Cohen, O. Novel CTL epitopes identified through a Y. pestis proteome-wide analysis in the search for vaccine candidates against plague. Vaccine 2017, 35, 5995–6006. [Google Scholar] [CrossRef]
- Sabhnani, L.; Rao, D.N. Identification of immunodominant epitope of F1 antigen of Yersinia pestis. FEMS Immunol. Med Microbiol. 2000, 27, 155–162. [Google Scholar] [CrossRef]
- Musson, J.A.; Ingram, R.; Durand, G.; Ascough, S.; Waters, E.L.; Hartley, M.G.; Robson, T.; Maillere, B.; Williamson, E.D.; Sriskandan, S.; et al. Repertoire of HLA-DR1-Restricted CD4 T-Cell Responses to Capsular Caf1 Antigen of Yersinia pestis in Human Leukocyte Antigen Transgenic Mice. Infect. Immun. 2010, 78, 4356–4362. [Google Scholar] [CrossRef] [Green Version]
- Sabhnani, L.; Manocha, M.; Sridevi, K.; Shashikiran, D.; Rayanade, R.; Rao, D. Developing subunit immunogens using B and T cell epitopes and their constructs derived from the F1 antigen of Yersinia pestis using novel delivery vehicles. FEMS Immunol. Med. Microbiol. 2003, 38, 215–229. [Google Scholar] [CrossRef] [Green Version]
- Braciale, V.L.; Nash, M.; Sinha, N.; Zudina, I.V.; Motin, V.L. Correlates of Immunity Elicited by Live Yersinia pestis Vaccine. In National Institute of Allergy and Infectious Diseases, NIH; Humana Press: Totowa, NJ, USA, 2008; pp. 473–480. [Google Scholar] [CrossRef]
- Feodorova, V.A.; Lyapina, A.M.; Khizhnyakova, M.A.; Zaitsev, S.S.; Sayapina, L.V.; Arseneva, T.E.; Trukhachev, A.L.; Lebedeva, S.A.; Telepnev, M.V.; Ulianova, O.V.; et al. Humoral and cellular immune responses to Yersinia pestis Pla antigen in humans immunized with live plague vaccine. PLoS Negl. Trop. Dis. 2018, 12, e0006511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponomarenko, J.V.; Bui, H.-H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitrov, I.; Bangov, I.; Flower, D.R.; Doytchinova, I. AllerTOP v.2—a server for in silico prediction of allergens. J. Mol. Model. 2014, 20, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Raghava, G.P.S. Open Source Drug Discovery Consortium In Silico Approach for Predicting Toxicity of Peptides and Proteins. PLoS ONE 2013, 8, e73957. [Google Scholar] [CrossRef] [Green Version]
- Doytchinova, I.; Flower, D.R. VAXIJN: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Randall, A.Z.; Sweredoski, M.J.; Baldi, P. SCRATCH: A protein structure and structural feature prediction server. Nucleic Acids Res. 2005, 33, W72–W76. [Google Scholar] [CrossRef] [Green Version]
- A Feodorova, V.; Corbel, M.J. Prospects for new plague vaccines. Expert Rev. Vaccines 2009, 8, 1721–1738. [Google Scholar] [CrossRef]
- Benner, G.E.; Andrews, G.P.; Byrne, W.R.; Strachan, S.D.; Sample, A.K.; Heath, D.G.; Friedlander, A.M. Immune response to Yersinia outer proteins and other Yersinia pestis antigens after experimental plague infection in mice. Infect Immun 1999, 67, 1922–1928. [Google Scholar] [CrossRef]
- Li, B.; Du, C.; Zhou, L.; Bi, Y.; Wang, X.; Wen, L.; Guo, Z.; Song, Z.; Yang, R. Humoral and Cellular Immune Responses to Yersinia pestis Infection in Long-Term Recovered Plague Patients. Clin. Vaccine Immunol. 2012, 19, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Butler, T.; Hudson, B.W. The serological response to Yersinia pestis infection. Bull. World Heal. Organ. 1977, 55, 39–42. [Google Scholar]
- Li, B.; Jiang, L.; Song, Q.; Yang, J.; Chen, Z.; Guo, Z.; Zhou, D.; Du, Z.; Song, Y.; Wang, J.; et al. Protein Microarray for Profiling Antibody Responses to Yersinia pestis Live Vaccine. Infect. Immun. 2005, 73, 3734–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, V.; Chitralekha, K.; Bakshi, A.; Tomar, D.; Deshmukh, R.; Baig, M.; Rao, D. Inducing systemic and mucosal immune responses to B-T construct of F1 antigen of Yersinia pestis in microsphere delivery. Vaccine 2006, 24, 3279–3289. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhu, Z.; Dankmeyer, J.L.; Wormald, M.M.; Fast, R.L.; Worsham, P.L.; Cote, C.K.; Amemiya, K.; Dimitrov, D.S. Human Anti-Plague Monoclonal Antibodies Protect Mice from Yersinia pestis in a Bubonic Plague Model. PLoS ONE 2010, 5, e13047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, R.; Naqvi, R.A.; Kumar, S.; Bhat, A.A.; Rao, D. Multiple Antigen Peptide Containing B and T Cell Epitopes of F1 Antigen of Yersinia pestis Showed Enhanced Th1 Immune Response in Murine Model. Scand. J. Immunol. 2013, 77, 361–371. [Google Scholar] [CrossRef]
- Liu, W.; Ren, J.; Zhang, J.; Song, X.; Liu, S.; Chi, X.; Chen, Y.; Wen, Z.; Li, J.; Chen, W. Identification and characterization of a neutralizing monoclonal antibody that provides complete protection against Yersinia pestis. PLoS ONE 2017, 12, e0177012. [Google Scholar] [CrossRef]
- Devdariani, Z.L.; A Fedorova, V.; Gromova, O.V.; Taranenko, T.M. Comparative incidence of detection of specific antibodies to Yersinia pestis capsular antigen and lipopolysaccharide in humans immunized with pest vaccine. Klin. Lab. Diagn. 1997, 39–41. [Google Scholar] [CrossRef]
- Andrews, G.P.; Strachan, S.T.; Benner, G.E.; Sample, A.K.; Anderson, G.W.; Adamovicz, J.J.; Welkos, S.L.; Pullen, J.K.; Friedlander, A.M. Protective Efficacy of Recombinant Yersinia Outer Proteins against Bubonic Plague Caused by Encapsulated and Nonencapsulated Yersinia pestis. Infect. Immun. 1999, 67, 1533–1537. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhou, L.; Guo, J.; Wang, X.; Ni, B.; Ke, Y.; Zhu, Z.; Guo, Z.; Yang, R. High-Throughput Identification of New Protective Antigens from a Yersinia pestis Live Vaccine by Enzyme-Linked Immunospot Assay. Infect. Immun. 2009, 77, 4356–4361. [Google Scholar] [CrossRef] [Green Version]
- Frey, S.E.; Lottenbach, K.; Graham, I.; Anderson, E.; Bajwa, K.; May, R.C.; Mizel, S.B.; Graff, A.; Belshe, R.B. A phase I safety and immunogenicity dose escalation trial of plague vaccine, Flagellin/F1/V, in healthy adult volunteers (DMID 08-0066). Vaccine 2017, 35, 6759–6765. [Google Scholar] [CrossRef]
- Hamzabegovic, F.; Goll, J.B.; Hooper, W.F.; Frey, S.; Gelber, C.E.; Abate, G. Flagellin adjuvanted F1/V subunit plague vaccine induces T cell and functional antibody responses with unique gene signatures. NPJ Vaccines 2020, 5, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Quenee, L.E.; Cornelius, C.A.; Ciletti, N.A.; Elli, D.; Schneewind, O. Yersinia pestis caf1 Variants and the Limits of Plague Vaccine Protection. Infect. Immun. 2008, 76, 2025–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeure, C.E.; Derbise, A.; Guillas, C.; Gerke, C.; Cauchemez, S.; Carniel, E.; Pizarro-Cerdá, J. Humoral and cellular immune correlates of protection against bubonic plague by a live Yersinia pseudotuberculosis vaccine. Vaccine 2019, 37, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhou, L.; Qi, Z.; Zhang, Q.; Dai, R.; Yang, Y.; Cui, B.; Wang, H.; Yang, R.; Wang, X. Long-Term Observation of Subunit Vaccine F1-rV270 against Yersinia pestis in Mice. Clin. Vaccine Immunol. 2009, 17, 199–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, C.; Titecat, M.; Poiret, S.; Cayet, D.; Boutillier, D.; Simonet, M.; Sirard, J.-C.; Lemaître, N.; Sebbane, F. Characterization of the protective immune response to Yersinia pseudotuberculosis infection in mice vaccinated with an LcrV-secreting strain of Lactococcus lactis. Vaccine 2016, 34, 5762–5767. [Google Scholar] [CrossRef]
- Sanapala, S.; Rahav, H.; Patel, H.; Sun, W.; Curtiss, R. Multiple antigens of Yersinia pestis delivered by live recombinant attenuated Salmonella vaccine strains elicit protective immunity against plague. Vaccine 2016, 34, 2410–2416. [Google Scholar] [CrossRef] [Green Version]
- Arnaboldi, P.M.; Sambir, M.; Arco, C.D.; Peters, L.A.; Seegers, J.F.; Mayer, L.; McCormick, A.A.; Dattwyler, R.J. Intranasal delivery of a protein subunit vaccine using a Tobacco Mosaic Virus platform protects against pneumonic plague. Vaccine 2016, 34, 5768–5776. [Google Scholar] [CrossRef] [Green Version]
- Leary, S.E.; Griffin, K.F.; Garmory, H.S.; Williamson, E.D.; Titball, R.W. Expression of an F1/V fusion protein in attenuated Salmonella typhimurium and protection of mice against plague. Microb. Pathog. 1997, 23, 167–179. [Google Scholar] [CrossRef]
- Uppada, S.B.; Bhat, A.A.; Sah, A.; Donthamshetty, R.N. Enhanced humoral and mucosal immune responses after intranasal immunization with chimeric multiple antigen peptide of LcrV antigen epitopes of Yersinia pestis coupled to palmitate in mice. Vaccine 2011, 29, 9352–9360. [Google Scholar] [CrossRef]
- Khan, A.A.; Babu, J.P.; Gupta, G.; Rao, D. Identifying B and T cell epitopes and studying humoral, mucosal and cellular immune responses of peptides derived from V antigen of Yersinia pestis. Vaccine 2008, 26, 316–332. [Google Scholar] [CrossRef]
- Pullen, J.K.; Anderson, G.W.; Welkos, S.L.; Friedlander, A.M. Analysis of the Yersinia pestis V Protein for the Presence of Linear Antibody Epitopes. Infect. Immun. 1998, 66, 521–527. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
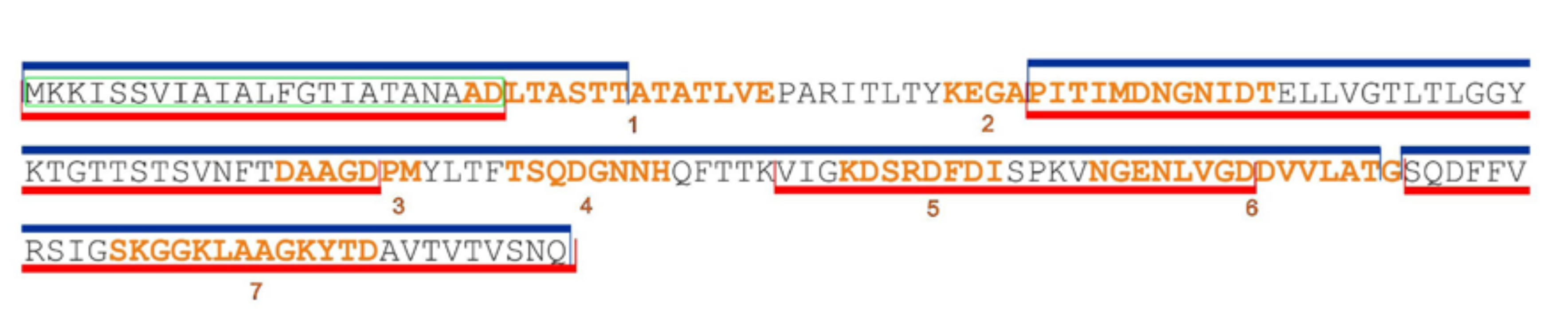{kind=link}
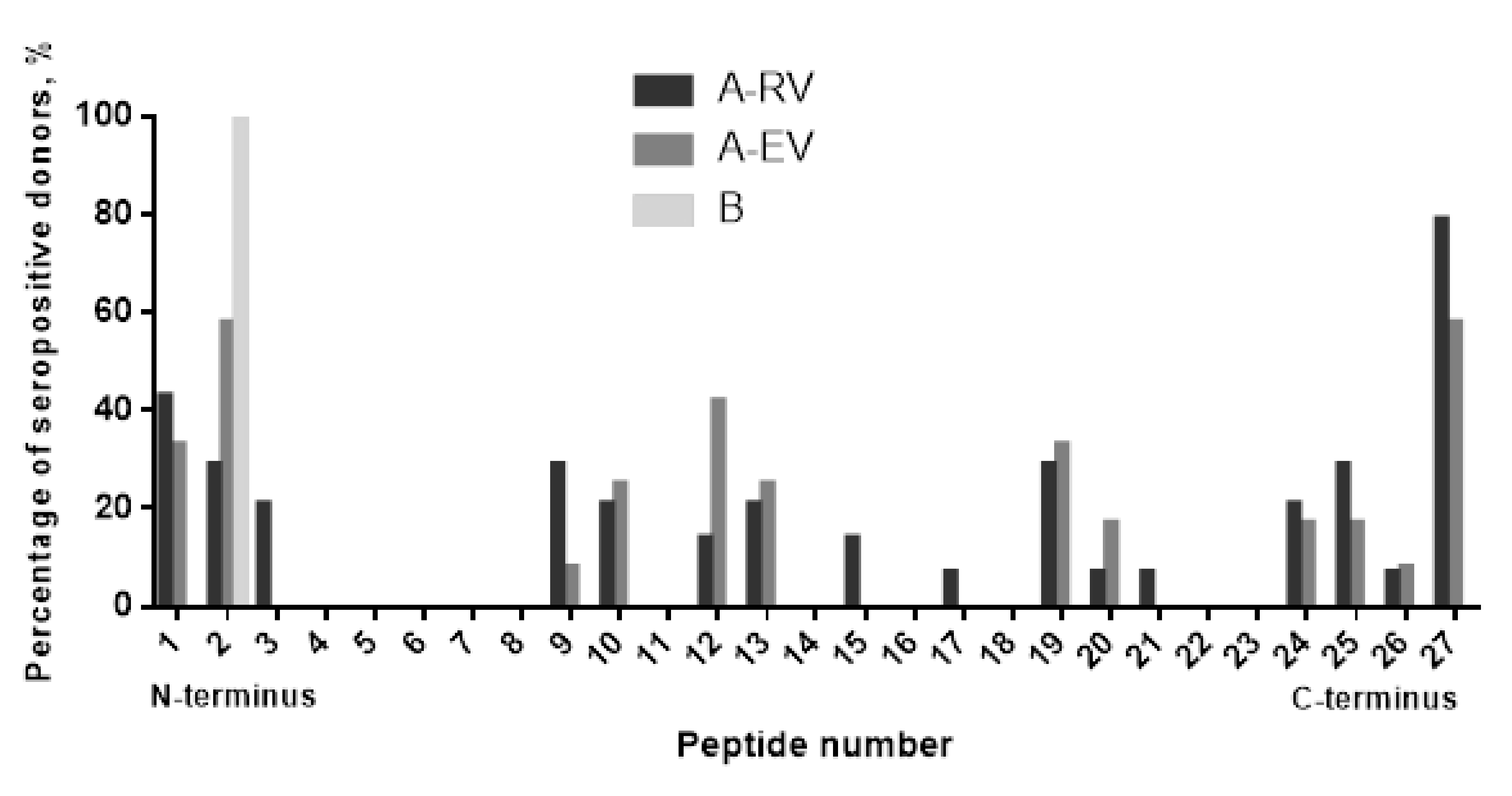{kind=link}
| Antigen | Epitope Number | B-cell Epitope 1 | Position | Number of Residues | Antigenicity 2 | Allergenicity 2 | Toxicity 3 | 3D Structure | |
|---|---|---|---|---|---|---|---|---|---|
| Antigenicity Scores | Status | ||||||||
| F1 | 1 | ADLTASTTATATLVE | 22–36 | 15 | 0.7645 | Antigen | Probable allergen | No | Figure S10A, Supplementary Materials |
| 2 | KEGAPITIMDNGNIDT | 45–60 | 16 | 0.5891 | Antigen | Nonallergen | No | ||
| 3 | DAAGDPM | 85–91 | 7 | 0.6144 | Antigen | Nonallergen | No | ||
| 4 | TSQDGNNH | 96–103 | 8 | 1.7902 | Antigen | Probable allergen | No | ||
| 5 | KDSRDFDI | 112–119 | 8 | 1.7530 | Antigen | Probable allergen | No | ||
| 6 | NGENLVGDDVVLATG | 124–138 | 15 | 0.6398 | Antigen | Allergen | No | ||
| 7 | SKGGKLAAGKYTD | 149–161 | 13 | 1.1808 | Antigen | Nonallergen | No | ||
| LcrV | 1 | GSSVLEELVQLVKDKNID | 28–45 | 18 | −0.0587 | Nonantigen | Nonallergen | No | Figure S10A, Supplementary Materials |
| 2 | KYDPRKDSEVFANRVITDDIELLKK | 49–73 | 25 | 0.5326 | Antigen | Allergen | No | ||
| 3 | AYFLPEDAILKGGHYDNQLQN | 76–96 | 21 | 0.3679 | Nonantigen | Nonallergen | No | ||
| 4 | ESSPNTQ | 106–112 | 7 | 1.6615 | Antigen | Allergen | No | ||
| 5 | NKHLSSSGT | 175–183 | 9 | 1.0585 | Antigen | Allergen | No | ||
| 6 | IHDKS | 186–190 | 5 | N/A | N/A | Allergen | No | ||
| 7 | MDKNLYGYTDEEIFKAS | 194–210 | 17 | 0.3717 | Nonantigen | Nonallergen | No | ||
| 8 | PQTTIQVDGSEKKIV | 220–234 | 15 | 1.0058 | Antigen | Nonallergen | No | ||
| 9 | GNLKNSYSYNKDNNELSHFATTCSDKSRP | 251–279 | 29 | 0.6307 | Antigen | Nonallergen | No | ||
| Antigen | Epitope # | B-cell Epitope Residues and Position 1 | Number of Residues | Score | 3D Structure |
|---|---|---|---|---|---|
| F1 | 1 | B:A22, B:D23, B:L24, B:T25, B:A26, B:S27, B:T28, B:T29, B:A30, B:T31, B:A32, B:T33, B:L34, B:V35, B:E36, B:P37 | 16 | 0.849 | Figure S10B, Supplementary Materials |
| 2 | B:K45, B:E46, B:G47, B:A48, B:P49, B:I50, B:T51, B:I52, B:M53, B:D54, B:N55, B:G56, B:N57, B:I58, B:D59, B:T60, B:L62, B:T96, B:S97, B:Q98, B:D99, B:G100, B:N101, B:N102, B:H103, B:S149, B:K150, B:G151, B:G152, B:K153, B:L154, B:A155, B:A156, B:G157, B:K158, B:Y159, B:T160, B:D161 | 38 | 0.672 | ||
| 3 | B:G75, B:T77, B:S78, B:T79, B:D85, B:A86, B:A87, B:G88, B:D89, B:P90, B:M91, B:Y92, B:K112, B:D113, B:S114, B:R115, B:D116, B:F117, B:D118, B:I119, B:S120, B:P121, B:K122, B:N124, B:G125, B:E126, B:N127, B:L128, B:V129, B:G130, B:D131, B:D132, B:V133, B:V134, B:L135 | 35 | 0.567 | ||
| LcrV | 1 | A:N175, A:K176, A:H177, A:L178, A:S179, A:S180, A:S181, A:G182, A:T183, A:I186, A:H187, A:D188, A:K189, A:G251, A:N252, A:L253, A:K254, A:N255, A:S256, A:S258, A:Y259, A:N260, A:K261, A:D262, A:N263, A:N264, A:E265, A:L266, A:S267, A:H268, A:F269, A:A270, A:T271, A:T272, A:C273, A:S274, A:D275, A:K276, A:S277, A:R278, A:P279 | 41 | 0.766 | Figure S10B, Supplementary Materials |
| 2 | A:K49, A:Y50, A:D51, A:P52, A:R53, A:K54, A:D55, A:S56, A:E57, A:V58, A:F59, A:A60, A:N61, A:R62, A:D148 | 15 | 0.741 | ||
| 3 | A:N192, A:D195, A:K196, A:N197, A:L198, A:G200, A:Y201, A:T202, A:D203, A:E204, A:E205, A:I206, A:F207, A:K208, A:A209, A:S210, A:P220, A:Q221, A:T222, A:T223, A:I224, A:Q225, A:V226, A:D227, A:G228, A:S229, A:E230, A:K231, A:K232, A:I233, A:V234, A:S235 | 32 | 0.729 | ||
| 4 | A:G28, A:S29, A:S30, A:V31, A:L32, A:E33, A:E34, A:V36, A:Q37, A:L38, A:V39, A:K40, A:D41, A:K42, A:N43, A:I44, A:D45, A:V63, A:I64, A:T65, A:D66, A:D67, A:I68, A:E69, A:L70, A:K72, A:K73, A:A76, A:F78, A:L79, A:P80, A:E81, A:D82, A:A83, A:I84, A:L85, A:K86, A:G87, A:G88, A:H89, A:Y90, A:D91, A:N92, A:Q93, A:L94, A:Q95, A:N96, A:E106, A:S108, A:P109, A:N110, A:T111, A:Q112 | 53 | 0.689 | ||
| 5 | A:D313, A:S314, A:R318 | 3 | 0.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feodorova, V.A.; Lyapina, A.M.; Khizhnyakova, M.A.; Zaitsev, S.S.; Saltykov, Y.V.; Motin, V.L. Yersinia pestis Antigen F1 but Not LcrV Induced Humoral and Cellular Immune Responses in Humans Immunized with Live Plague Vaccine—Comparison of Immunoinformatic and Immunological Approaches. Vaccines 2020, 8, 698. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8040698
Feodorova VA, Lyapina AM, Khizhnyakova MA, Zaitsev SS, Saltykov YV, Motin VL. Yersinia pestis Antigen F1 but Not LcrV Induced Humoral and Cellular Immune Responses in Humans Immunized with Live Plague Vaccine—Comparison of Immunoinformatic and Immunological Approaches. Vaccines. 2020; 8(4):698. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8040698
Chicago/Turabian StyleFeodorova, Valentina A., Anna M. Lyapina, Maria A. Khizhnyakova, Sergey S. Zaitsev, Yury V. Saltykov, and Vladimir L. Motin. 2020. "Yersinia pestis Antigen F1 but Not LcrV Induced Humoral and Cellular Immune Responses in Humans Immunized with Live Plague Vaccine—Comparison of Immunoinformatic and Immunological Approaches" Vaccines 8, no. 4: 698. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8040698