
Chimeric RHDV Virus-Like Particles Displaying Foot-and-Mouth Disease Virus Epitopes Elicit Neutralizing Antibodies and Confer Partial Protection in Pigs

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus, Cells and Peptides
2.2. Generation of Recombinant Baculovirus Transfer Vectors
2.3. Generation of Recombinant Baculoviruses
2.4. Expression and Purification of Recombinant Chimeric RHDV VLPs
2.5. Transmission Electron Microscopy
2.6. Mice Immunization
2.7. Immunization and Infections of Pigs
2.8. Detection of Specific Anti-RHDV-VLP Antibodies by ELISA
2.9. Detection of Specific Anti-FMDV Antibodies by ELISA
2.10. Virus Neutralization Test (VNT)
2.11. IFN-γ Detection by ELISPOT
2.12. Lymphoproliferation Assay
2.13. Data and Statistical Analysis
2.14. Ethics Statement
3. Results
3.1. Design and Characterization of RHDV-Based Chimeric VLPs Displaying FMDV B- and T-Cell Epitopes
3.2. Immune Responses Elicited by Chimeric VLPs Displaying FMDV B- and T-Cell Epitopes in Mice
3.2.1. Chimeric VLPs Displaying FMDV Epitopes Elicit Specific Humoral Responses including Neutralizing Antibodies
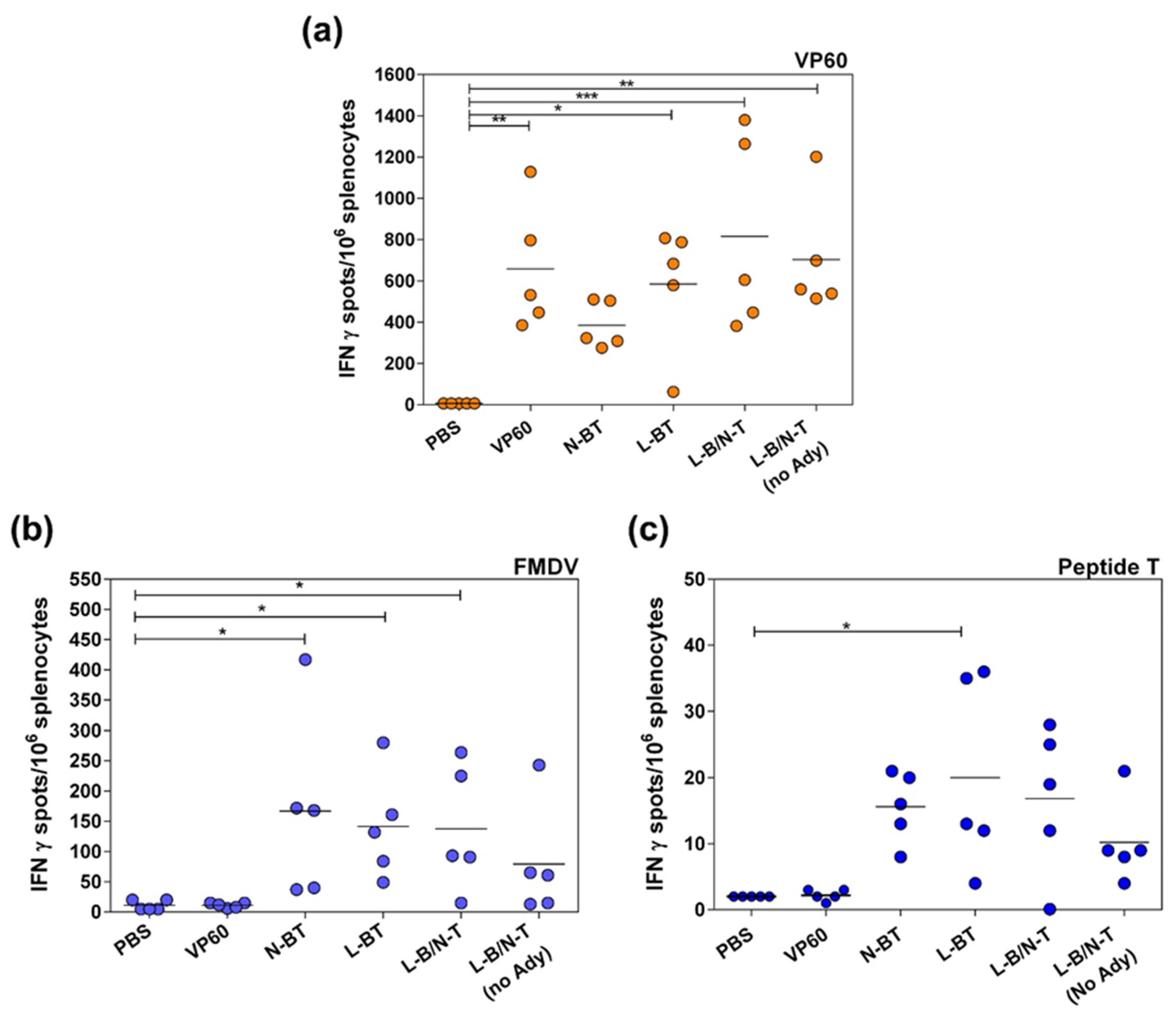
3.2.2. T-Cell Responses Elicited by Chimeric VLPs Displaying FMDV Epitopes in Mice
3.3. Immune Response and Protection Conferred by Chimeric VLP L-B/N-T in Pigs
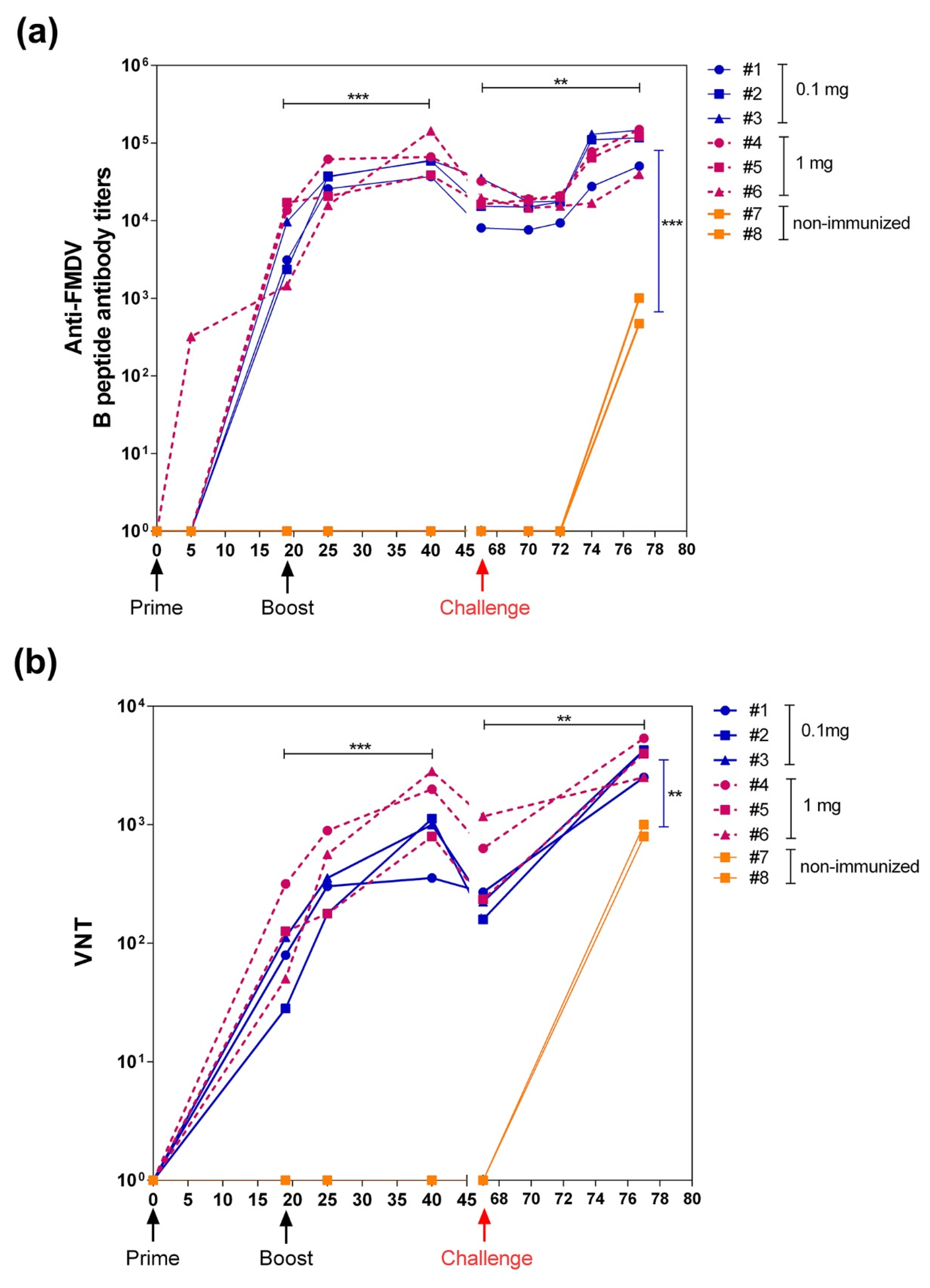
3.3.1. VLP L-B/N-T Elicits Robust Humoral FMDV-Specific Response and VNT Titers in Pigs
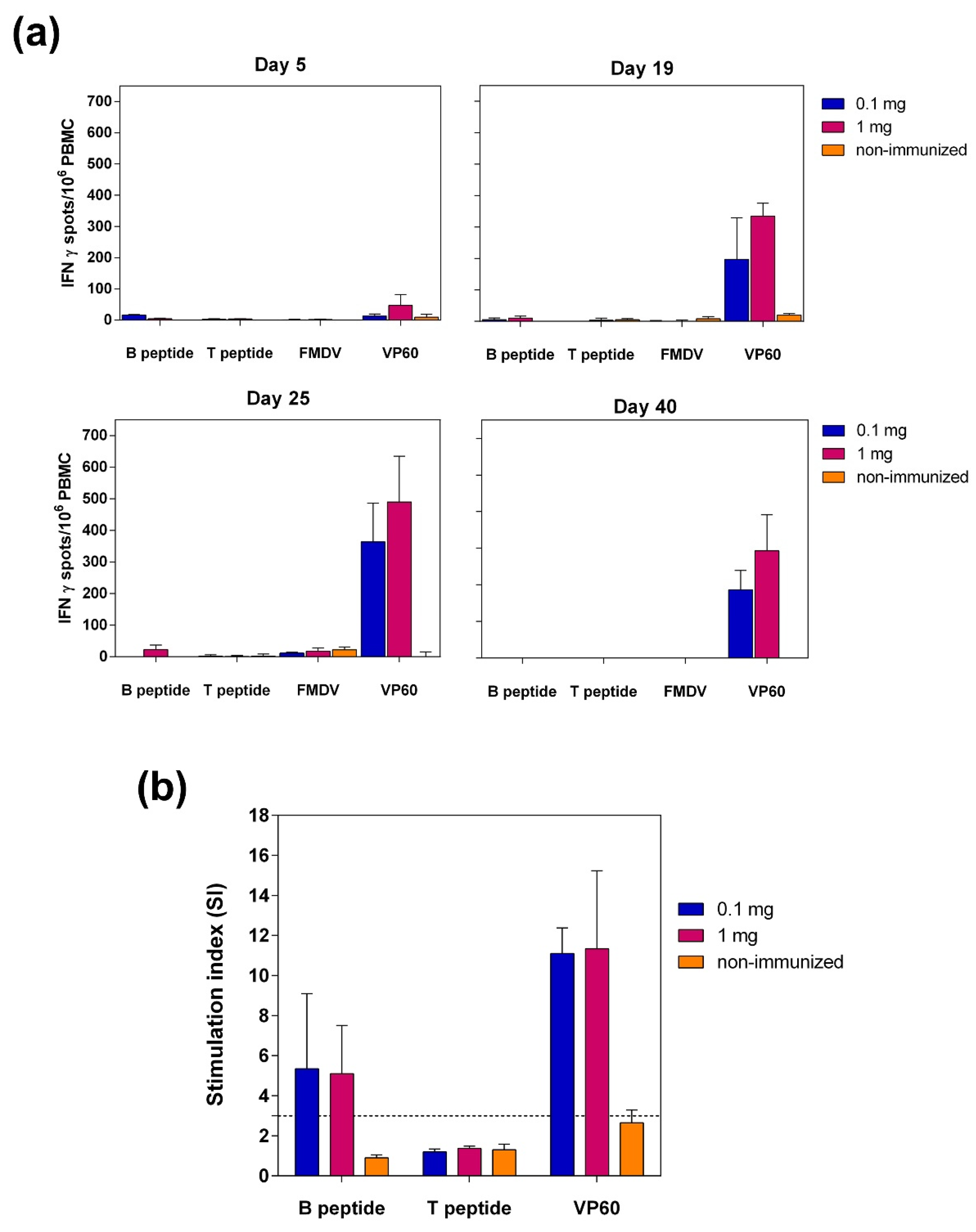
3.3.2. Specific Cellular Immune Responses Elicited by VLP L-B/N-T in Pigs
3.3.3. Immunization with VLP L-B/N-T Provides Partial Clinical Protection against an FMDV Challenge in Pigs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, J.E.; Chiu, W. Structures of virus and virus-like particles. Curr. Opin. Struct. Biol. 2000, 10, 229–235. [Google Scholar] [CrossRef]
- Mateu, M.G. Assembly, Engineering and Applications of Virus-Based Protein Nanoparticles. Adv. Exp. Med. Biol. 2016, 940, 83–120. [Google Scholar] [CrossRef]
- Lua, L.H.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P. Bioengineering virus-like particles as vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Zeltins, A. Construction and characterization of virus-like particles: A review. Mol. Biotechnol. 2013, 53, 92–107. [Google Scholar] [CrossRef]
- Pushko, P.; Pumpens, P.; Grens, E. Development of virus-like particle technology from small highly symmetric to large complex virus-like particle structures. Intervirology 2013, 56, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, M.O.; Augusto, G.; Bachmann, M.F. The 3Ds in virus-like particle based-vaccines: “Design, Delivery and Dynamics”. Immunol. Rev. 2020, 296, 155–168. [Google Scholar] [CrossRef]
- Mohsen, M.O.; Gomes, A.C.; Vogel, M.; Bachmann, M.F. Interaction of Viral Capsid-Derived Virus-Like Particles (VLPs) with the Innate Immune System. Vaccines 2018, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Kelly, H.G.; Kent, S.J.; Wheatley, A.K. Immunological basis for enhanced immunity of nanoparticle vaccines. Expert Rev. Vaccines 2019, 18, 269–280. [Google Scholar] [CrossRef]
- Crisci, E.; Barcena, J.; Montoya, M. Virus-like particles: The new frontier of vaccines for animal viral infections. Vet. Immunol. Immunopathol. 2012, 148, 211–225. [Google Scholar] [CrossRef]
- Charlton Hume, H.K.; Vidigal, J.; Carrondo, M.J.T.; Middelberg, A.P.J.; Roldao, A.; Lua, L.H.L. Synthetic biology for bioengineering virus-like particle vaccines. Biotechnol. Bioeng. 2019, 116, 919–935. [Google Scholar] [CrossRef] [Green Version]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major findings and recent advances in virus-like particle (VLP)-based vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Moreno, N.; Mena, I.; Angulo, I.; Gomez, Y.; Crisci, E.; Montoya, M.; Caston, J.R.; Blanco, E.; Barcena, J. Rabbit hemorrhagic disease virus capsid, a versatile platform for foreign B-cell epitope display inducing protective humoral immune responses. Sci. Rep. 2016, 6, 31844. [Google Scholar] [CrossRef]
- Mateu, M.G. Virus engineering: Functionalization and stabilization. Protein Eng. Des. Sel. 2011, 24, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Brune, K.D.; Leneghan, D.B.; Brian, I.J.; Ishizuka, A.S.; Bachmann, M.F.; Draper, S.J.; Biswas, S.; Howarth, M. Plug-and-Display: Decoration of Virus-Like Particles via isopeptide bonds for modular immunization. Sci. Rep. 2016, 6, 19234. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, M.F.; Jennings, G.T. Therapeutic vaccines for chronic diseases: Successes and technical challenges. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2815–2822. [Google Scholar] [CrossRef] [Green Version]
- Barcena, J.; Blanco, E. Design of novel vaccines based on virus-like particles or chimeric virions. Subcell Biochem. 2013, 68, 631–665. [Google Scholar] [CrossRef]
- Lee, K.L.; Twyman, R.M.; Fiering, S.; Steinmetz, N.F. Virus-based nanoparticles as platform technologies for modern vaccines. Wiley Interdiscip Rev. Nanomed. Nanobiotechnol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Recule, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Celio, A.P.; et al. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Valicek, L.; Smid, B.; Rodak, L.; Kudrna, J. Electron and immunoelectron microscopy of rabbit haemorrhagic disease virus (RHDV). Arch. Virol. 1990, 112, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Vautherot, J.F.; Madelaine, M.F.; Le Gall, G.; Rasschaert, D. Recombinant rabbit hemorrhagic disease virus capsid protein expressed in baculovirus self-assembles into viruslike particles and induces protection. J. Virol. 1994, 68, 6794–6798. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xu, F.; Liu, J.; Gao, B.; Liu, Y.; Zhai, Y.; Ma, J.; Zhang, K.; Baker, T.S.; Schulten, K.; et al. Atomic model of rabbit hemorrhagic disease virus by cryo-electron microscopy and crystallography. PLoS Pathog. 2013, 9, e1003132. [Google Scholar] [CrossRef] [Green Version]
- Leuthold, M.M.; Dalton, K.P.; Hansman, G.S. Structural analysis of a rabbit hemorrhagic disease virus binding to histo-blood group antigens. J. Virol. 2015, 89, 2378–2387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcena, J.; Guerra, B.; Angulo, I.; Gonzalez, J.; Valcarcel, F.; Mata, C.P.; Caston, J.R.; Blanco, E.; Alejo, A. Comparative analysis of rabbit hemorrhagic disease virus (RHDV) and new RHDV2 virus antigenicity, using specific virus-like particles. Vet. Res. 2015, 46, 106. [Google Scholar] [CrossRef] [Green Version]
- Barcena, J.; Verdaguer, N.; Roca, R.; Morales, M.; Angulo, I.; Risco, C.; Carrascosa, J.L.; Torres, J.M.; Caston, J.R. The coat protein of Rabbit hemorrhagic disease virus contains a molecular switch at the N-terminal region facing the inner surface of the capsid. Virology 2004, 322, 118–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisci, E.; Almanza, H.; Mena, I.; Cordoba, L.; Gomez-Casado, E.; Caston, J.R.; Fraile, L.; Barcena, J.; Montoya, M. Chimeric calicivirus-like particles elicit protective anti-viral cytotoxic responses without adjuvant. Virology 2009, 387, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Luque, D.; Gonzalez, J.M.; Gomez-Blanco, J.; Marabini, R.; Chichon, J.; Mena, I.; Angulo, I.; Carrascosa, J.L.; Verdaguer, N.; Trus, B.L.; et al. Epitope insertion at the N-terminal molecular switch of the rabbit hemorrhagic disease virus T = 3 capsid protein leads to larger T = 4 capsids. J. Virol. 2012, 86, 6470–6480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight-Jones, T.J.; Robinson, L.; Charleston, B.; Rodriguez, L.L.; Gay, C.G.; Sumption, K.J.; Vosloo, W. Global Foot-and-Mouth Disease Research Update and Gap Analysis: 2—Epidemiology, Wildlife and Economics. Transbound. Emerg. Dis. 2016, 63 (Suppl. 1), 14–29. [Google Scholar] [CrossRef]
- Robinson, L.; Knight-Jones, T.J.; Charleston, B.; Rodriguez, L.L.; Gay, C.G.; Sumption, K.J.; Vosloo, W. Global Foot-and-Mouth Disease Research Update and Gap Analysis: 3—Vaccines. Transbound Emerg. Dis. 2016, 63 (Suppl. 1), 30–41. [Google Scholar] [CrossRef]
- Pfaff, E.; Mussgay, M.; Bohm, H.O.; Schulz, G.E.; Schaller, H. Antibodies against a preselected peptide recognize and neutralize foot and mouth disease virus. EMBO J. 1982, 1, 869–874. [Google Scholar] [CrossRef]
- Acharya, R.; Fry, E.; Stuart, D.; Fox, G.; Rowlands, D.; Brown, F. The three-dimensional structure of foot-and-mouth disease virus at 2.9 A resolution. Nature 1989, 337, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Blanco, E.; Garcia-Briones, M.; Sanz-Parra, A.; Gomes, P.; De Oliveira, E.; Valero, M.L.; Andreu, D.; Ley, V.; Sobrino, F. Identification of T-cell epitopes in nonstructural proteins of foot-and-mouth disease virus. J. Virol. 2001, 75, 3164–3174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubillos, C.; de la Torre, B.G.; Jakab, A.; Clementi, G.; Borras, E.; Barcena, J.; Andreu, D.; Sobrino, F.; Blanco, E. Enhanced mucosal immunoglobulin A response and solid protection against foot-and-mouth disease virus challenge induced by a novel dendrimeric peptide. J. Virol. 2008, 82, 7223–7230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, E.; Guerra, B.; de la Torre, B.G.; Defaus, S.; Dekker, A.; Andreu, D.; Sobrino, F. Full protection of swine against foot-and-mouth disease by a bivalent B-cell epitope dendrimer peptide. Antiviral. Res. 2016, 129, 74–80. [Google Scholar] [CrossRef]
- Canas-Arranz, R.; Forner, M.; Defaus, S.; Rodriguez-Pulido, M.; de Leon, P.; Torres, E.; Bustos, M.J.; Borrego, B.; Saiz, M.; Blanco, E.; et al. A bivalent B-cell epitope dendrimer peptide can confer long-lasting immunity in swine against foot-and-mouth disease. Transbound Emerg. Dis. 2020, 67, 1614–1622. [Google Scholar] [CrossRef] [PubMed]
- Cubillos, C.; de la Torre, B.G.; Barcena, J.; Andreu, D.; Sobrino, F.; Blanco, E. Inclusion of a specific T cell epitope increases the protection conferred against foot-and-mouth disease virus in pigs by a linear peptide containing an immunodominant B cell site. Virol. J. 2012, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Blanco, E.; Cubillos, C.; Moreno, N.; Barcena, J.; de la Torre, B.G.; Andreu, D.; Sobrino, F. B epitope multiplicity and B/T epitope orientation influence immunogenicity of foot-and-mouth disease peptide vaccines. Clin. Dev. Immunol. 2013, 2013, 475960. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.J.; Black, L. Response of young pigs to foot-and-mouth disease oil emulsion vaccination in the presence and absence of maternally derived neutralising antibodies. Res. Vet. Sci. 1986, 41, 33–39. [Google Scholar] [CrossRef]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Chapter 3.1.8. Foot and Mouth Disease (Infection with Foot and Mouth Disease Virus). Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.08_FMD.pdf (accessed on 6 May 2021).
- Golding, S.M.; Hedger, R.S.; Talbot, P. Radial immuno-diffusion and serum-neutralisation techniques for the assay of antibodies to swine vesicular disease. Res. Vet. Sci. 1976, 20, 142–147. [Google Scholar] [CrossRef]
- Perez-Filgueira, D.M.; Resino-Talavan, P.; Cubillos, C.; Angulo, I.; Barderas, M.G.; Barcena, J.; Escribano, J.M. Development of a low-cost, insect larvae-derived recombinant subunit vaccine against RHDV. Virology 2007, 364, 422–430. [Google Scholar] [CrossRef]
- Almanza, H.; Cubillos, C.; Angulo, I.; Mateos, F.; Caston, J.R.; van der Poel, W.H.; Vinje, J.; Barcena, J.; Mena, I. Self-assembly of the recombinant capsid protein of a swine norovirus into virus-like particles and evaluation of monoclonal antibodies cross-reactive with a human strain from genogroup II. J. Clin. Microbiol. 2008, 46, 3971–3979. [Google Scholar] [CrossRef] [Green Version]
- Cubillos-Zapata, C.; Angulo, I.; Almanza, H.; Borrego, B.; Zamora-Ceballos, M.; Caston, J.R.; Mena, I.; Blanco, E.; Barcena, J. Precise location of linear epitopes on the capsid surface of feline calicivirus recognized by neutralizing and non-neutralizing monoclonal antibodies. Vet. Res. 2020, 51, 59. [Google Scholar] [CrossRef] [PubMed]
- Rouco, C.; Abrantes, J.; Serronha, A.; Lopes, A.M.; Maio, E.; Magalhaes, M.J.; Blanco, E.; Barcena, J.; Esteves, P.J.; Santos, N.; et al. Epidemiology of RHDV2 (Lagovirus europaeus/GI.2) in free-living wild European rabbits in Portugal. Transbound Emerg. Dis. 2018, 65, e373–e382. [Google Scholar] [CrossRef] [PubMed]
- Crisci, E.; Fraile, L.; Novellas, R.; Espada, Y.; Cabezon, R.; Martinez, J.; Cordoba, L.; Barcena, J.; Benitez-Ribas, D.; Montoya, M. In vivo tracking and immunological properties of pulsed porcine monocyte-derived dendritic cells. Mol. Immunol. 2015, 63, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Peers-Adams, A.; Win, S.J.; Scullion, S.; Wilson, M.; Young, V.L.; Jennings, P.; Ward, V.K.; Baird, M.A.; Young, S.L. Antigen incorporated in virus-like particles is delivered to specific dendritic cell subsets that induce an effective antitumor immune response in vivo. J. Immunother. 2013, 36, 11–19. [Google Scholar] [CrossRef]
- Win, S.J.; Ward, V.K.; Dunbar, P.R.; Young, S.L.; Baird, M.A. Cross-presentation of epitopes on virus-like particles via the MHC I receptor recycling pathway. Immunol. Cell Biol. 2011, 89, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.; Al-Barwani, F.; Baird, M.A.; Young, V.L.; Larsen, D.S.; Ward, V.K.; Young, S.L. Functionalisation of Virus-Like Particles Enhances Antitumour Immune Responses. J. Immunol. Res. 2019, 2019, 5364632. [Google Scholar] [CrossRef]
- Donaldson, B.; Al-Barwani, F.; Pelham, S.J.; Young, K.; Ward, V.K.; Young, S.L. Multi-target chimaeric VLP as a therapeutic vaccine in a model of colorectal cancer. J. Immunother. Cancer 2017, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Peacey, M.; Wilson, S.; Perret, R.; Ronchese, F.; Ward, V.K.; Young, V.; Young, S.L.; Baird, M.A. Virus-like particles from rabbit hemorrhagic disease virus can induce an anti-tumor response. Vaccine 2008, 26, 5334–5337. [Google Scholar] [CrossRef]
- Canas-Arranz, R.; de Leon, P.; Forner, M.; Defaus, S.; Bustos, M.J.; Torres, E.; Andreu, D.; Blanco, E.; Sobrino, F. Immunogenicity of a Dendrimer B2T Peptide Harboring a T-Cell Epitope From FMDV Non-structural Protein 3D. Front. Vet. Sci. 2020, 7, 498. [Google Scholar] [CrossRef]
- Chang, J.; Zhang, Y.; Yang, D.; Jiang, Z.; Wang, F.; Yu, L. Potent neutralization activity against type O foot-and-mouth disease virus elicited by a conserved type O neutralizing epitope displayed on bovine parvovirus virus-like particles. J. Gen. Virol. 2019, 100, 187–198. [Google Scholar] [CrossRef]
- Li, X.; Meng, X.; Wang, S.; Li, Z.; Yang, L.; Tu, L.; Diao, W.; Yu, C.; Yu, Y.; Yan, C.; et al. Virus-like particles of recombinant PCV2b carrying FMDV-VP1 epitopes induce both anti-PCV and anti-FMDV antibody responses. Appl. Microbiol. Biotechnol. 2018, 102, 10541–10550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Guo, Y.J.; Wang, K.Y.; Lu, K.; Li, K.; Zhu, Y.; Sun, S.H. Enhanced immunogenicity of modified hepatitis B virus core particle fused with multiepitopes of foot-and-mouth disease virus. Scand J. Immunol. 2007, 65, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, F.C.; Turco, C.S.; Miraglia, M.C.; Bessone, F.A.; Franco, R.; Perez-Filgueira, M.; Sala, J.M.; Capozzo, A.V. The role of viral particle integrity in the serological assessment of foot-and-mouth disease virus vaccine-induced immunity in swine. PLoS ONE 2020, 15, e0232782. [Google Scholar] [CrossRef] [PubMed]
- Singanallur, N.B.; Dekker, A.; Eble, P.L.; van Hemert-Kluitenberg, F.; Weerdmeester, K.; Horsington, J.; Vosloo, W.W. Emergency Foot-and-Mouth Disease Vaccines A Malaysia 97 and A22 Iraq 64 Offer Good Protection against Heterologous Challenge with A Variant Serotype A ASIA/G-IX/SEA-97 Lineage Virus. Vaccines 2020, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueda, P.; Moron, G.; Sarraseca, J.; Leclerc, C.; Casal, J.I. Influence of flanking sequences on presentation efficiency of a CD8+ cytotoxic T-cell epitope delivered by parvovirus-like particles. J. Gen. Virol. 2004, 85, 563–572. [Google Scholar] [CrossRef]
- Alves, M.P.; Guzylack-Piriou, L.; Juillard, V.; Audonnet, J.C.; Doel, T.; Dawson, H.; Golde, W.T.; Gerber, H.; Peduto, N.; McCullough, K.C.; et al. Innate immune defenses induced by CpG do not promote vaccine-induced protection against foot-and-mouth disease virus in pigs. Clin. Vaccine Immunol. 2009, 16, 1151–1157. [Google Scholar] [CrossRef] [Green Version]
- McCullough, K.C.; De Simone, F.; Brocchi, E.; Capucci, L.; Crowther, J.R.; Kihm, U. Protective immune response against foot-and-mouth disease. J. Virol. 1992, 66, 1835–1840. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inoculum | Vaccine Dose (mg) | Pig # | First Day with Vesicles 1 | Number of Vesicles 2 | Fever 3 | SN 4 Prime/Boost/Challenge | Protection 5 |
|---|---|---|---|---|---|---|---|
| RHDV VLP L-B/N-T | 0.1 | #1 | 7 | 3 | 39.8 (4) | 1.9/2.4/3.4 | Partial protection |
| #2 | 7 | 1 | No fever | 1.5/2.2/3.6 | Full protection | ||
| #3 | 7 | 3 | 40.9 (4) | 2.0/2.3/3.6 | Partial protection | ||
| 1 | #4 | 4 | 4 | 40.1 (6) | 2.5/2.8/3.7 | Partial protection | |
| #5 | 7 | 3 | 39.9 (5) | 2.1/2.4/3.6 | Partial protection | ||
| #6 | 7 | 3 | No fever | 1.7/3.6/3.4 | Partial protection | ||
| Non-immunized | - | #7 | 4 | 4 | 40.7 (4) | 0/0/3.0 | Non-protected |
| #8 | 3 | 4 | 40.0 (4) | 0/0/2.9 | Non-protected |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangel, G.; Bárcena, J.; Moreno, N.; Mata, C.P.; Castón, J.R.; Alejo, A.; Blanco, E. Chimeric RHDV Virus-Like Particles Displaying Foot-and-Mouth Disease Virus Epitopes Elicit Neutralizing Antibodies and Confer Partial Protection in Pigs. Vaccines 2021, 9, 470. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050470
Rangel G, Bárcena J, Moreno N, Mata CP, Castón JR, Alejo A, Blanco E. Chimeric RHDV Virus-Like Particles Displaying Foot-and-Mouth Disease Virus Epitopes Elicit Neutralizing Antibodies and Confer Partial Protection in Pigs. Vaccines. 2021; 9(5):470. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050470
Chicago/Turabian StyleRangel, Giselle, Juan Bárcena, Noelia Moreno, Carlos P. Mata, José R. Castón, Alí Alejo, and Esther Blanco. 2021. "Chimeric RHDV Virus-Like Particles Displaying Foot-and-Mouth Disease Virus Epitopes Elicit Neutralizing Antibodies and Confer Partial Protection in Pigs" Vaccines 9, no. 5: 470. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050470