
Impacts of Quarterly Sow Mass Vaccination with a Porcine Reproductive and Respiratory Syndrome Virus Type 1 (PRRSV-1) Modified Live Vaccine in Two Herds


Abstract
:1. Introduction
2. Materials and Methods
2.1. Farms
2.2. Experimental Design
2.3. Laboratory Analysis
2.4. Statistical Analysis
3. Results
3.1. RT-qPCR
3.2. Sequence Analysis
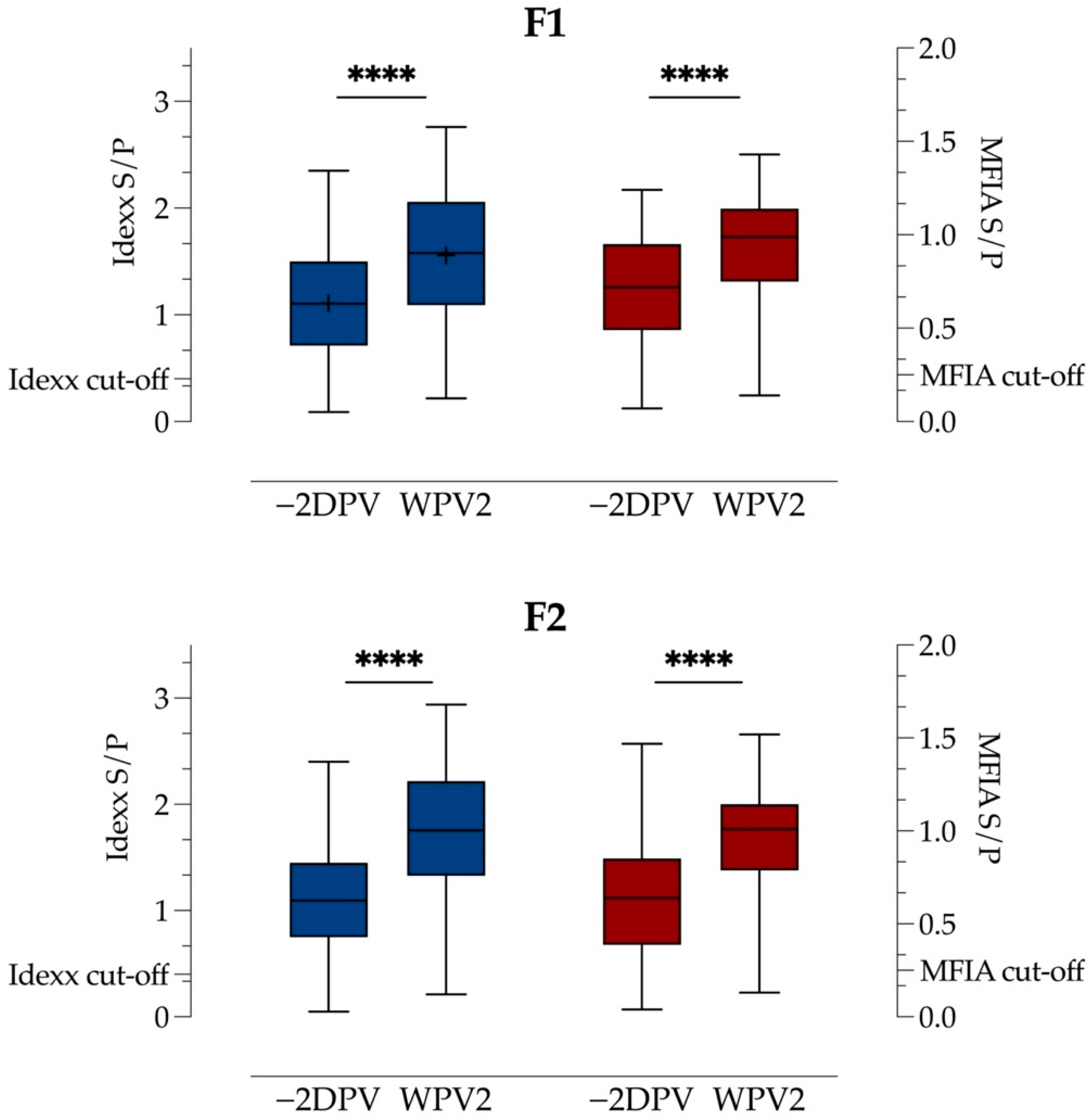
3.3. Test for Antibodies
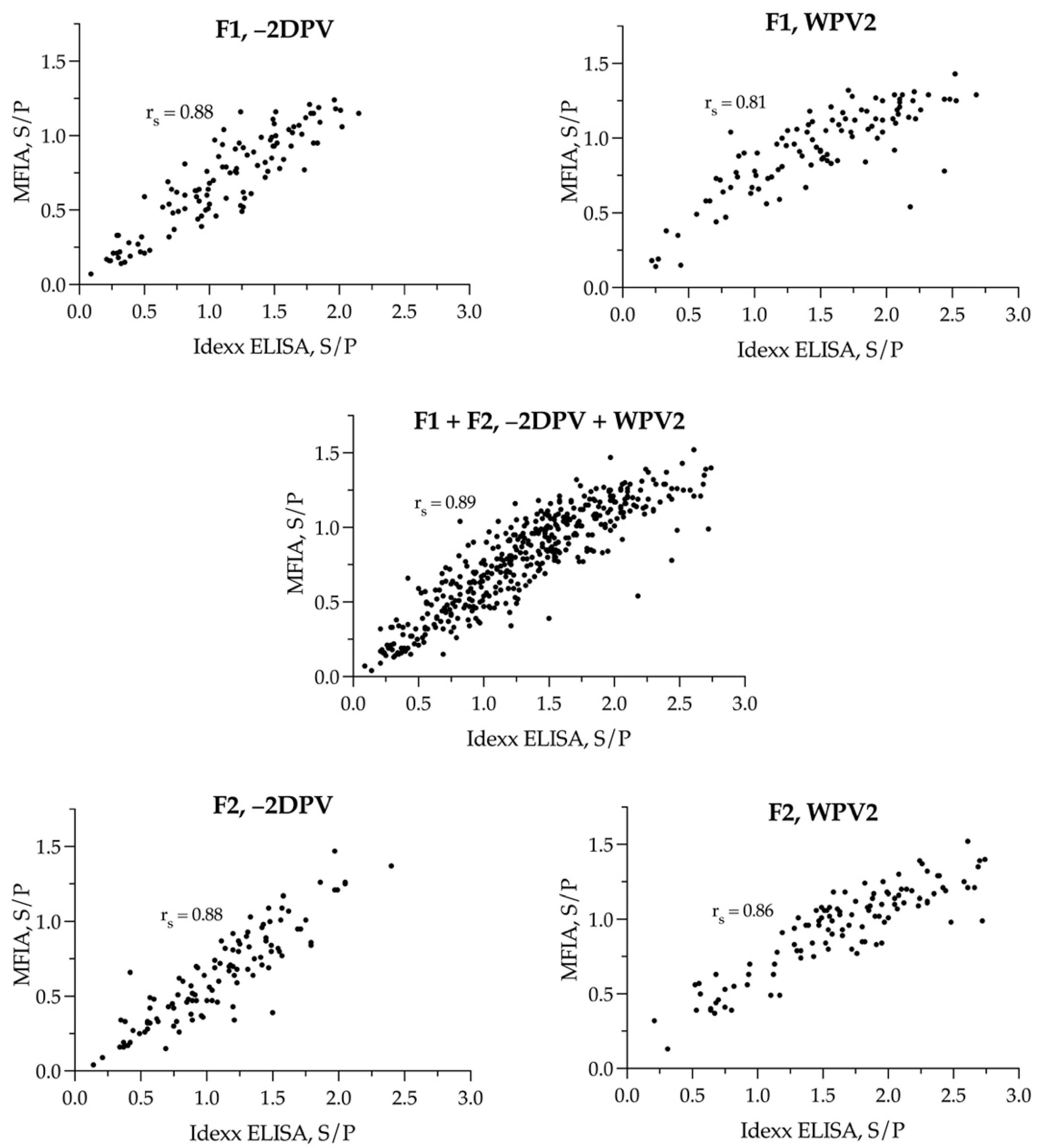
3.4. Agreement in Results between the Idexx ELISA and the MFIA Serological Tests
4. Discussion
4.1. Virus
4.2. Effect of SMV
4.3. Higher Response with Lower Value −2DPV
4.4. Agreement between Idexx ELISA and MFIA
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kvisgaard, L.K.; Kristensen, C.S.; Ryt-Hansen, P.; Pedersen, K.; Stadejek, T.; Trebbien, R.; Andresen, L.O.; Larsen, L.E. A recombination between two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) vaccine strains has caused severe outbreaks in Danish pigs. Transbound. Emerg. Dis. 2020, 67, 1786–1796. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, D.J.; Polson, D.D.; Torremorell, M.; Morrison, B.; Rowland, R.R.; Snelson, H.; Straw, B.; Yeske, P.; Zimmerman, J. Terminology for Classifying Swine Herds by Porcine Reproductive and Respiratory Syndrome Virus Status. J. Swine Health Prod. 2011, 19, 13. [Google Scholar]
- Kristensen, C.S.; Christiansen, M.G.; Pedersen, K.; Larsen, L.E. Production losses five months after outbreak with a recombinant of two PRRSV vaccine strains in 13 Danish sow herds. Porc. Health Manag. 2020, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Zuckermann, F.A.; Garcia, E.A.; Luque, I.D.; Christopher-Hennings, J.; Doster, A.; Brito, M.; Osorio, F. Assessment of the efficacy of commercial porcine reproductive and respiratory syndrome virus (PRRSV) vaccines based on measurement of serologic response, frequency of gamma-IFN-producing cells and virological parameters of protection upon challenge. Vet. Microbiol. 2007, 123, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Aly, S.S.; Cano, J.P.; Polson, D.; Kass, P.H.; Perez, A.M. Stochastic model of porcine reproductive and respiratory syndrome virus control strategies on a swine farm in the United States. Am. J. Vet. Res. 2014, 75, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Linhares, D.C.L.; Johnson, C.; Morrison, R.B. Economic Analysis of Vaccination Strategies for PRRS Control. PLoS ONE 2015, 10, e0144265. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Park, C.; Choi, K.; Jeong, J.; Kang, I.; Park, S.-J.; Chae, C. Comparison of Two Commercial Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Modified Live Vaccines against Heterologous Type 1 and Type 2 PRRSV Challenge in Growing Pigs. Clin. Vaccine Immunol. 2015, 22, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Pejsak, Z.; Markowska-Daniel, I. Randomised, placebo-controlled trial of a live vaccine against porcine reproductive and respiratory syndrome virus in sows on infected farms. Vet. Rec. 2006, 158, 475–478. [Google Scholar] [CrossRef]
- Alexopoulos, C.; Kritas, S.; Kyriakis, C.S.; Tzika, E. Sow performance in an endemically porcine reproductive and respiratory syndrome (PRRS)-infected farm after sow vaccination with an attenuated PRRS vaccine. Vet. Microbiol. 2005, 111, 151–157. [Google Scholar] [CrossRef]
- Madapong, A.; Saeng-Chuto, K.; Boonsoongnern, A.; Tantituvanont, A.; Nilubol, D. Cell-mediated immune response and protective efficacy of porcine reproductive and respiratory syndrome virus modified-live vaccines against co-challenge with PRRSV-1 and PRRSV-2. Sci. Rep. 2020, 10, 1649. [Google Scholar] [CrossRef]
- Díaz, I.; Gimeno, M.; Callén, A.; Pujols, J.; López, S.; Charreyre, C.; Joisel, F.; Mateu, E. Comparison of different vaccination schedules for sustaining the immune response against porcine reproductive and respiratory syndrome virus. Vet. J. 2013, 197, 438–444. [Google Scholar] [CrossRef]
- Bassaganya-Riera, J.; Thacker, B.J.; Yu, S.; Strait, E.; Wannemuehler, M.J.; Thacker, E.L. Impact of Immunizations with Porcine Reproductive and Respiratory Syndrome Virus on Lymphoproliferative Recall Responses of CD8+ T Cells. Viral Immunol. 2004, 17, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Díaz, I.; Genís-Jorquera, B.; Martin-Valls, G.; Mateu, E. Using commercial ELISAs to assess humoral response in sows repeatedly vaccinated with modified live porcine reproductive and respiratory syndrome virus. Vet. Rec. 2020, 186, 123. [Google Scholar] [CrossRef]
- Wagstrom, E.A.; Chang, C.-C.; Yoon, K.-J.; Zimmerman, J.J. Shedding of porcine reproductive and respiratory syndrome virus in mammary gland secretions of sows. Am. J. Vet. Res. 2001, 62, 1876–1880. [Google Scholar] [CrossRef]
- Baker, B.; Thacker, E.; Thacker, B.; Vincent, A. A Preliminary Investigation into Possible PRRSV Anergy Induction from Repeated Immunization with a Modified Live Vaccine. In Proceedings of the 26th Allen D. Leman Swine Conference, St. Paul, MN, USA, 18–21 September 1999; p. 31. [Google Scholar]
- Cano, J.P.; Dee, S.A.; Murtaugh, M.P.; Rovira, A.; Morrison, R.B. Infection dynamics and clinical manifestations following experimental inoculation of gilts at 90 days of gestation with a low dose of porcine reproductive and respiratory syndrome virus. Can. J. Vet. Res. 2009, 73, 303–307. [Google Scholar]
- Rowland, R.R. The interaction between PRRSV and the late gestation pig fetus. Virus Res. 2010, 154, 114–122. [Google Scholar] [CrossRef]
- Khurana, S.; Hahn, M.; Coyle, E.M.; King, L.R.; Lin, T.-L.; Treanor, J.; Sant, A.; Golding, H. Repeat vaccination reduces antibody affinity maturation across different influenza vaccine platforms in humans. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilalta, C.; Sanhueza, J.; Garrido, J.; Murray, D.; Morrison, R.; Corzo, C.A.; Torremorell, M. Indirect assessment of porcine reproductive and respiratory syndrome virus status in pigs prior to weaning by sampling sows and the environment. Vet. Microbiol. 2019, 237, 108406. [Google Scholar] [CrossRef] [PubMed]
- Kittawornrat, A.; Panyasing, Y.; Goodell, C.; Wang, C.; Gauger, P.; Harmon, K.; Rauh, R.; Desfresne, L.; Levis, I.; Zimmerman, J. Porcine reproductive and respiratory syndrome virus (PRRSV) surveillance using pre-weaning oral fluid samples detects circulation of wild-type PRRSV. Vet. Microbiol. 2014, 168, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Lebret, A.; Boulbria, G.; Berton, P.; Moalic, P.-Y.; Le Guennec, J.; Bouchet, F.; Auvigne, V.; Normand, V. Monitoring PRRSV-1 in suckling piglets in an endemic herd using reverse transcriptase quantitative real time polymerase chain reaction: Comparison of the rate of detection in serum and oral fluid samples and evaluation of pooling. Porc. Health Manag. 2019, 5, 8. [Google Scholar] [CrossRef]
- Kvisgaard, L.; Larsen, L.; Kristensen, C.; Paboeuf, F.; Renson, P.; Bourry, O. Challenge of Naïve and Vaccinated Pigs with a Vaccine-Derived Recombinant Porcine Reproductive and Respiratory Syndrome Virus 1 Strain (Horsens Strain). Vaccines 2021, 9, 417. [Google Scholar] [CrossRef]
- Wernike, K.; Bonilauri, P.; Dauber, M.; Errington, J.; Leblanc, N.; Revilla-Fernández, S.; Hjulsager, C.K.; Isaksson, M.; Stadejek, T.; Beer, M.; et al. Porcine reproductive and respiratory syndrome virus. J. Vet. Diagn. Investig. 2012, 24, 855–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvisgaard, L.K.; Hjulsager, C.K.; Brar, M.S.; Leung, F.C.; Larsen, L.E. Genetic dissection of complete genomes of Type 2 PRRS viruses isolated in Denmark over a period of 15 years. Vet. Microbiol. 2013, 167, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Kvisgaard, L.K.; Hjulsager, C.K.; Kristensen, C.S.; Lauritsen, K.T.; Larsen, L.E. Genetic and antigenic characterization of complete genomes of Type 1 Porcine Reproductive and Respiratory Syndrome viruses (PRRSV) isolated in Denmark over a period of 10 years. Virus Res. 2013, 178, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrowski, M.; Galeota, J.A.; Jar, A.M.; Platt, K.B.; Osorio, F.A.; Lopez, O.J. Identification of Neutralizing and Nonneutralizing Epitopes in the Porcine Reproductive and Respiratory Syndrome Virus GP5 Ectodomain. J. Virol. 2002, 76, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebret, A.; Berton, P.; Normand, V.; Messager, I.; Robert, N.; Bouchet, F.; Brissonnier, M.; Boulbria, G. PRRSV detection by qPCR in processing fluids and serum samples collected in a positive stable breeding herd following mass vaccination of sows with a modified live vaccine. Porc. Health Manag. 2021, 7, 1–5. [Google Scholar] [CrossRef]
- Torrents, D.; Miranda, J.; Pedrazuela, R.; Gauger, P.C.; Ramirez, A.; Linhares, D.C.L. Implementation of PRRSV status classification system in swine breeding herds from a large integrated group in Spain. Porc. Health Manag. 2019, 5, 1–8. [Google Scholar] [CrossRef]
- Wiedermann, U.; Garner-Spitzer, E.; Wagner, A. Primary vaccine failure to routine vaccines: Why and what to do? Hum. Vaccines Immunother. 2016, 12, 239–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronski, D.M.; Chambers, C.; Sabaiduc, S.; De Serres, G.; Winter, A.-L.; Dickinson, J.; Krajden, M.; Gubbay, J.B.; Drews, S.J.; Martineau, C.; et al. A Perfect Storm: Impact of Genomic Variation and Serial Vaccination on Low Influenza Vaccine Effectiveness During the 2014–2015 Season. Clin. Infect. Dis. 2016, 63, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, N.; Takeishi, K. Change in the efficacy of influenza vaccination after repeated inoculation under antigenic mismatch: A systematic review and meta-analysis. Vaccine 2018, 36, 949–957. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Time | Herd | n of Stables/Approx. Pigs Tested | n of Stables with Positive Samples | |
|---|---|---|---|---|
| PRRSV-1 | PRRSV-2 | |||
| −2DPV | N1 | 10/2000 | 1 | 2 |
| N2 | 10/2000 | 0 | 1 | |
| WPV2 | N1 | 13/2600 | 0 | 0 |
| N2 | 8/1600 | 0 | 1 | |
| WPV12 | N1 | 13/2600 | 0 | 4 |
| N2 | 5/1000 | 2 | 0 | |
| F1 | F2 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parity | Sows (n) | Idexx ELISA | MFIA | Both | Sows (n) | Idexx ELISA | MFIA | Both | ||||||
| −2DPV | WPV2 | −2DPV | WPV2 | −2DPV | WPV2 | −2DPV | WPV2 | −2DPV | WPV2 | −2DPV | WPV2 | |||
| 1 | 21 | 3 | 0 | 5 | 1 | 3 | 0 | 30 | 0 | 1 | 2 | 1 | 0 | 1 |
| 2 | 15 | 6 | 3 | 6 | 3 | 6 | 2 | 21 | 1 | 0 | 1 | 0 | 1 | 0 |
| 3 | 32 | 3 | 0 | 3 | 0 | 2 | 0 | 34 | 6 | 1 | 5 | 0 | 4 | 0 |
| 4 | 27 | 5 | 1 | 4 | 1 | 3 | 1 | 21 | 1 | 0 | 2 | 0 | 1 | 0 |
| 5 | 15 | 1 | 0 | 1 | 0 | 1 | 0 | 11 | 1 | 0 | 1 | 0 | 1 | 0 |
| 6 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 |
| 7 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 120 | 18 | 4 | 19 | 5 | 15 | 3 | 120 | 9 | 2 | 11 | 1 | 7 | 1 |
| 0 | - | - | - | 7 | 10 | - | - | - | - | - | 4 | 11 | - | - |
| +PRRSV-2 | - | - | - | 1 | 3 | - | - | - | - | - | 2 | 0 | - | - |
| Neg → Pos | - | 14 | 13 | 9 | - | 8 | 9 | 6 | ||||||
| Neg → Neg | - | 4 | 4 | 3 | - | 1 | 1 | 0 | ||||||
| Pos → Neg | - | 0 | 0 | 0 | - | 1 | 0 | 0 | ||||||
| IC → Pos | - | - | 5 | - | - | - | 3 | - | ||||||
| IC → Neg | - | - | 0 | - | - | - | 0 | - | ||||||
| Pos → IC | - | - | 9 | - | - | - | 9 | - | ||||||
| Neg → IC | - | - | 1 | - | - | - | 1 | - | ||||||
| Idexx ELISA Positive | Idexx ELISA Negative | Sum | |
|---|---|---|---|
| MFIA-positive | 394 | 7 | 401 |
| MFIA-negative | 10 | 26 | 36 |
| Sum | 404 | 33 | 437 |
| κ | SE | 95% CI | Observed Agreements | Agreements by Chance | |
|---|---|---|---|---|---|
| −2DPV | 0.74 | 0.07 | 0.60–0.87 | 213 (94.25%) | 176.2 (77.95%) |
| WPV2 | 0.66 | 0.16 | 0.34–0.97 | 207 (98.10%) | 199.3 (94.47%) |
| Total | 0.73 | 0.06 | 0.61–0.85 | 420 (96.11%) | 373.4 (85.45%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedersen, K.; Kristensen, C.S.; Kvisgaard, L.K.; Larsen, L.E. Impacts of Quarterly Sow Mass Vaccination with a Porcine Reproductive and Respiratory Syndrome Virus Type 1 (PRRSV-1) Modified Live Vaccine in Two Herds. Vaccines 2021, 9, 1057. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9101057
Pedersen K, Kristensen CS, Kvisgaard LK, Larsen LE. Impacts of Quarterly Sow Mass Vaccination with a Porcine Reproductive and Respiratory Syndrome Virus Type 1 (PRRSV-1) Modified Live Vaccine in Two Herds. Vaccines. 2021; 9(10):1057. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9101057
Chicago/Turabian StylePedersen, Kasper, Charlotte Sonne Kristensen, Lise Kirstine Kvisgaard, and Lars Erik Larsen. 2021. "Impacts of Quarterly Sow Mass Vaccination with a Porcine Reproductive and Respiratory Syndrome Virus Type 1 (PRRSV-1) Modified Live Vaccine in Two Herds" Vaccines 9, no. 10: 1057. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9101057