Seasonality of Nosema ceranae Infections and Their Relationship with Honey Bee Populations, Food Stores, and Survivorship in a North American Region


 ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Screening and Selection of Experimental Colonies
2.2. Infection Intensity
2.3. Spore Viability
2.4. Nosema ceranae Prevalence in Bees (Percentage of Infected Bees)
2.5. Colony Evaluations and Relationship with Nosema ceranae Infections
2.6. Effect of Nosema ceranae Infections on Bee Survivorship in Colonies
2.7. Statistical Analyses
3. Results
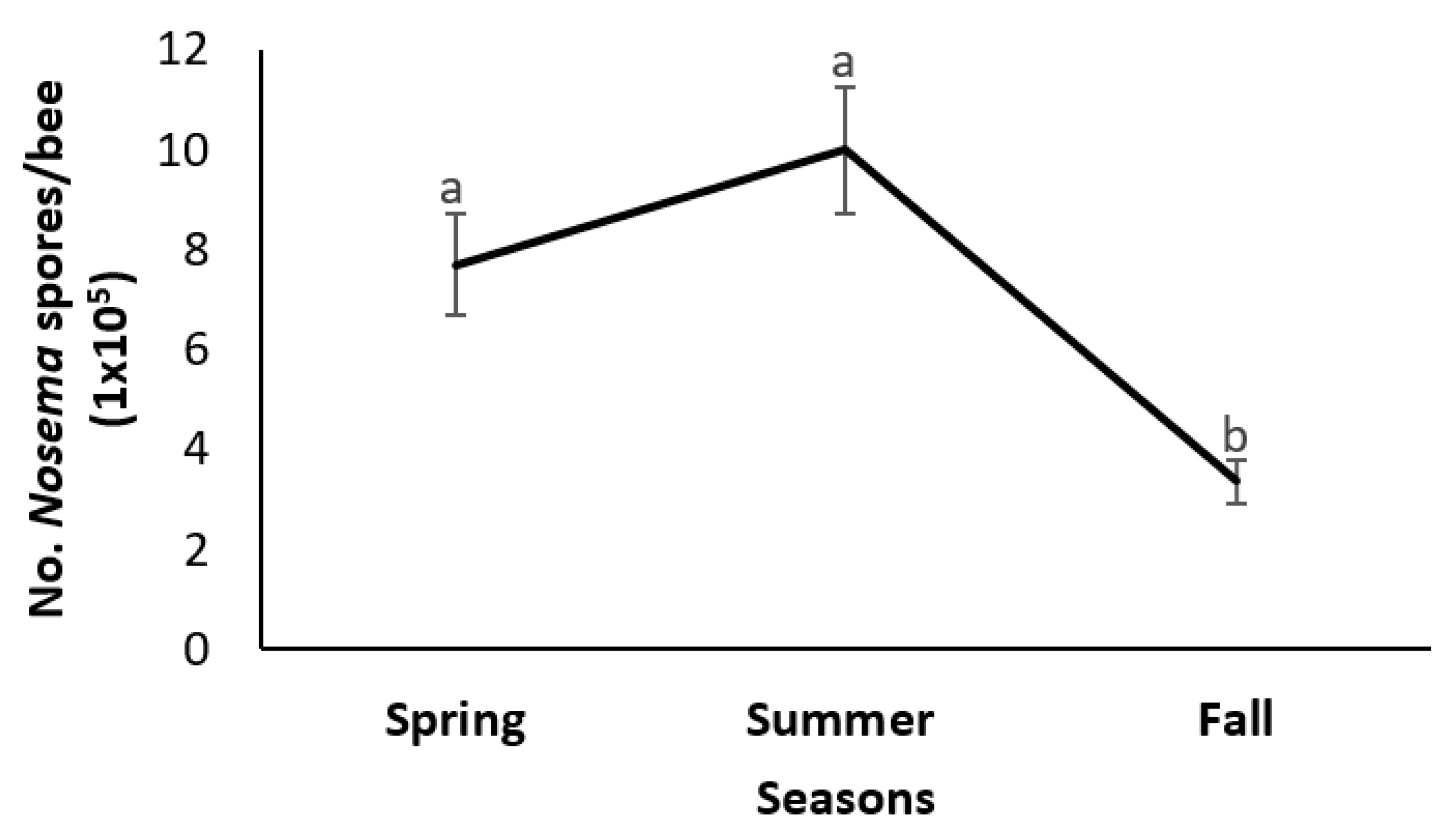
3.1. Intensity, Prevalence, and Viability of Nosema ceranae Infections
3.2. Nosema ceranae Infections and Colony Conditions
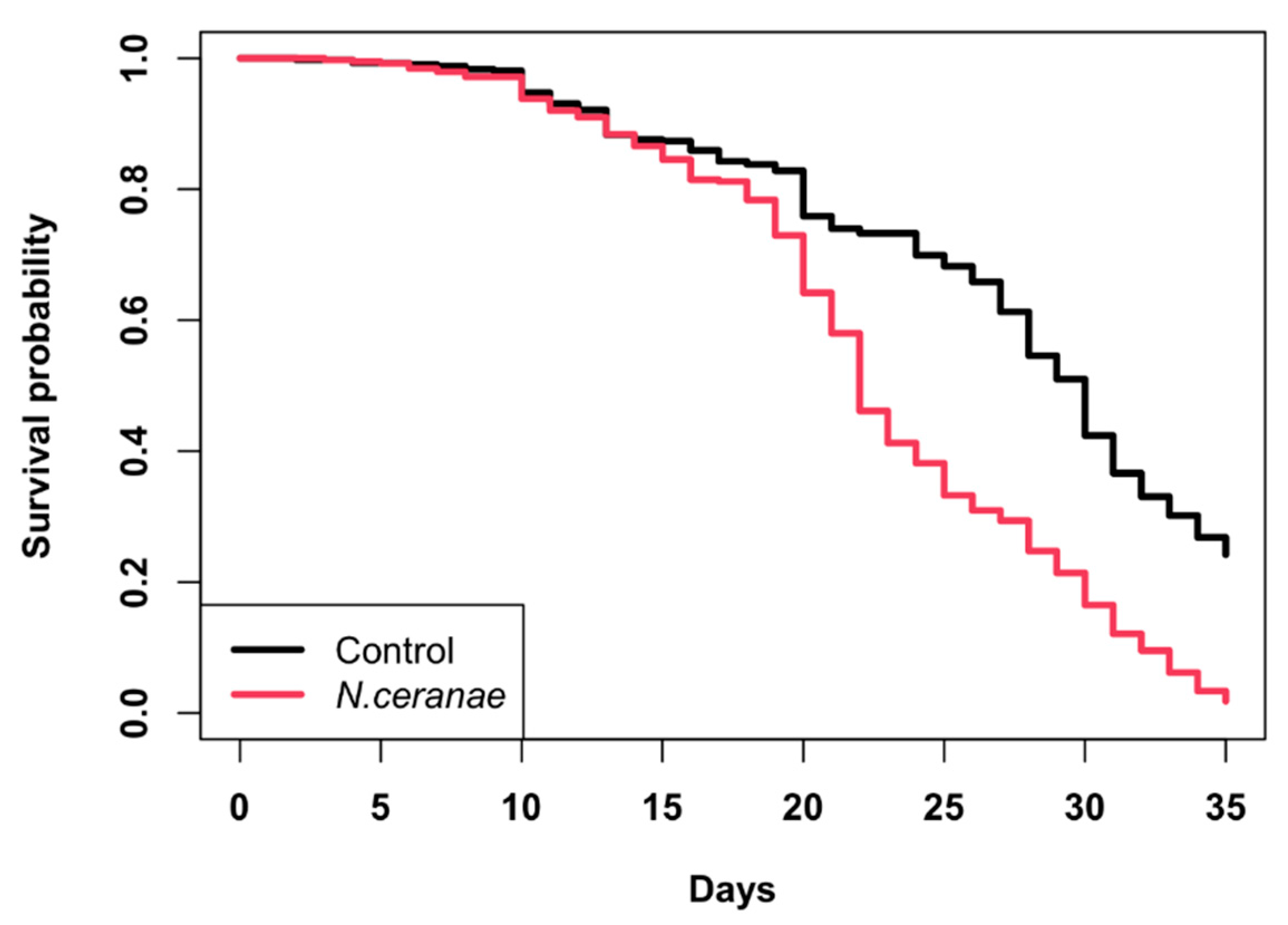
3.3. Survivorship of Bees in Colonies
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fries, I.; Feng, F.; da Silva, A.; Slemenda, S.B.; Pieniazek, N.J. Nosema ceranae n. sp. (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur. J. Parasitol. 1996, 32, 356–365. [Google Scholar] [CrossRef]
- Higes, M.; Martín, R.; Meana, A. Nosema ceranae, a new microsporidian parasite in honeybees in Europe. J. Invertebr. Pathol. 2006, 92, 93–95. [Google Scholar] [CrossRef]
- Huang, W.F.; Jiang, J.H.; Chen, Y.W.; Wang, C.-H. A Nosema ceranae isolate from the honeybee Apis mellifera. Apidologie 2007, 38, 30–37. [Google Scholar] [CrossRef]
- Fries, I. Protozoa. In Honey Bee Pests, Predators and Diseases, 3rd ed.; Morse, R.A., Flottum, K., Eds.; A I Root Company: Medina, OH, USA, 1997; pp. 57–76. [Google Scholar]
- Fries, I. Infectivity and multiplication of Nosema apis Z. in the ventriculus of the honey bee. Apidologie 1988, 19, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Goblirsch, M. Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie 2018, 49, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Cilia, G.; Cabbri, R.; Maiorana, G.; Cardaio, I.; Dall’Olio, R.; Nanetti, A. A novel TaqMan® assay for Nosema ceranae quantification in honey bee, based on the protein coding gene Hsp70. Eur. J. Protistol. 2018, 63, 44–50. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Bartolomé, C.; Chejanovsky, N.; Le Conte, Y.; Dalmon, A.; Dussaubat, C.; García-Palencia, P.; Meana, A.; Pinto, M.A.; Soroker, V.; et al. Nosema ceranae in Apis mellifera: A 12 years postdetection perspective. Environ. Microbiol. 2018, 20, 1302–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cilia, G.; Sagona, S.; Giusti, M.; Jarmela dos Santos, P.E.; Nanetti, A.; Felicioli, A. Nosema ceranae infection in honeybee samples from Tuscanian archipelago (Central Italy) investigated by two qPCR methods. Saudi J. Biol. Sci. 2019, 26, 1553–1556. [Google Scholar] [CrossRef]
- Meixner, M.D.; Francis, R.M.; Gajda, A.; Kryger, P.; Andonov, S.; Uzunov, A.; Topolska, G.; Costa, C.; Amiri, E.; Berg, S.; et al. Occurrence of parasites and pathogens in honey bee colonies used in a European genotype-environment interactions experiment. J. Apic. Res. 2014, 53, 215–229. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Botías, C.; Barrios, L.; Martínez-Salvador, A.; Meana, A.; Mayack, C.; Higes, M. Comparison of the energetic stress associated with experimental Nosema ceranae and Nosema apis infection of honeybees (Apis mellifera). Parasitol. Res. 2011, 109, 605–612. [Google Scholar] [CrossRef]
- Higes, M.; Martín-Hernández, R.; Meana, A. Nosema ceranae in Europe: An emergent type C nosemosis. Apidologie 2010, 41, 375–392. [Google Scholar] [CrossRef] [Green Version]
- Traver, B.E.; Williams, M.R.; Fell, R.D. Comparison of within hive sampling and seasonal activity of Nosema ceranae in honey bee colonies. J. Invertebr. Pathol. 2012, 109, 187–193. [Google Scholar] [CrossRef]
- Ibrahim, A.; Melathopoulos, A.P.; Pernal, S.F. Integrated management of Nosema and detection of antibiotic residues. Hivelights 2012, 25, 11–17. [Google Scholar]
- Sánchez Collado, J.G.; Higes, M.; Barrio, L.; Martín-Hernández, R. Flow cytometry analysis of Nosema species to assess spore viability and longevity. Parasitol. Res. 2014, 113, 1695–1701. [Google Scholar] [CrossRef]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef]
- Paxton, R.J.; Klee, J.; Korpela, S.; Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 2007, 38, 558–565. [Google Scholar] [CrossRef]
- Williams, G.R.; Shutler, D.; Burgher-MacLellan, K.L.; Rogers, R.E.L. Infra-Population and community dynamics of the parasites Nosema apis and Nosema ceranae, and consequences for honey bee (Apis mellifera) hosts. PLoS ONE 2014, 9, e99465. [Google Scholar] [CrossRef] [Green Version]
- Goblirsch, M.; Huang, Z.Y.; Spivak, M. Physiological and behavioral changes in honey bees (Apis mellifera) induced by Nosema ceranae infection. PLoS ONE 2013, 8, e58165. [Google Scholar] [CrossRef]
- Milbrath, M.O.; van Tran, T.; Huang, W.-F.; Solter, L.F.; Tarpy, D.R.; Lawrence, F.; Huang, Z.Y. Comparative virulence and competition between Nosema apis and Nosema ceranae in honey bees (Apis mellifera). J. Invertebr. Pathol. 2015, 125, 9–15. [Google Scholar] [CrossRef]
- Huang, W.-F.; Solter, L.; Aronstein, K.; Huang, Z. Infectivity and virulence of Nosema ceranae and Nosema apis in commercially available North American honey bees. J. Invertebr. Pathol. 2015, 124, 107–113. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Meana, A.; Prieto, L.; Salvador, A.M.; Garrido-Bailón, E.; Higes, M. Outcome of colonization of Apis mellifera by Nosema ceranae. Appl. Environ. Microbiol. 2007, 73, 6331–6338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G.; et al. How natural infection by Nosema ceranae causes honeybee Colony Collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; Martín-Hernández, R.; Garrido-Bailón, E.; González-Porto, A.V.; García-Palencia, P.; Meana, A.; Nozal, M.J.D.; Mayo, R.; Bernal, J.L. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 2009, 1, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumann, P. Predictive markers of honey bee colony collapse. PLoS ONE 2012, 7, e32151. [Google Scholar] [CrossRef] [PubMed]
- Siede, R.; Berg, S.; Meixner, M. Effects of symptomless infections with Nosema sp. on honey bee colonies. In Proceedings of the OIE-Symposium Diagnosis and Control of Bee Diseases, Freiburg, Germany, 26–28 August 2008. [Google Scholar]
- Gisder, S.; Hedtke, K.; Möckel, N.; Frielitz, M.-C.; Linde, A.; Genersch, E. Five-year cohort study of Nosema spp. in Germany: Does climate shape virulence and assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 2010, 76, 3032–3038. [Google Scholar] [CrossRef] [Green Version]
- Stevanovic, J.; Simeunovic, P.; Gajic, B.; Lakic, N.; Radovic, D.; Fries, I.; Stanimirovic, Z. Characteristics of Nosema ceranae infection in Serbian honey bee colonies. Apidologie 2013, 44, 522–536. [Google Scholar] [CrossRef] [Green Version]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic survey of microbes in honey bee Colony Collapse Disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Currie, R.W.; Pernal, S.F.; Guzman-Novoa, E. Honey bee colony losses in Canada. J. Apic. Res. 2010, 49, 104–106. [Google Scholar] [CrossRef]
- Guzman-Novoa, E.; Eccles, L.; Calvete, Y.; McGowan, J.; Kelly, P.G.; Correa-Benítez, A. Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada. Apidologie 2010, 41, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Hardy, J.; Villalobos, E.; Martín-Hernández, R.; Nikaido, S.; Higes, M. Do the honeybee pathogens Nosema ceranae and deformed wing virus act synergistically? Environ. Microbiol. Rep. 2013, 5, 506–510. [Google Scholar] [CrossRef] [Green Version]
- Emsen, B.; Guzman-Novoa, E.; Hamiduzzaman, M.M.; Eccles, L.; Lacey, B.; Ruiz-Pérez, R.A.; Nasr, M. Higher prevalence and levels of Nosema ceranae than Nosema apis infections in Canadian honey bee colonies. Parasitol. Res. 2016, 115, 175–181. [Google Scholar] [CrossRef]
- Guerrero-Molina, C.; Correa-Benítez, A.; Hamiduzzaman, M.M.; Guzman-Novoa, E. Nosema ceranae is an old resident of honey bee (Apis mellifera) colonies in Mexico, causing infection levels of one million spores per bee or higher during summer and fall. J. Invertebr. Pathol. 2016, 141, 38–40. [Google Scholar] [CrossRef] [PubMed]
- Invernizzi, C.; Abud, C.; Tomasco, I.H.; Harriet, J.; Ramallo, G.; Campá, J.; Katz, H.; Gardiol, G.; Mendoza, Y. Presence of Nosema ceranae in honeybees (Apis mellifera) in Uruguay. J. Invertebr. Pathol. 2009, 101, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Novoa, E.; Hamiduzzaman, M.M.; Arechavaleta-Velasco, M.E.; Koleoglu, G.; Valizadeh, P.; Correa-Benítez, A. Nosema ceranae has parasitized Africanized honey bees in Mexico since at least 2004. J. Apic. Res. 2011, 50, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Holt, H.L.; Grozinger, C.M. Approaches and challenges to managing Nosema (Microspora: Nosematidae) parasites in honey bee (Hymenoptera: Apidae) Colonies. J. Econ. Entomol. 2016, 109, 1487–1503. [Google Scholar] [CrossRef] [PubMed]
- Gary, N.E.; Lorenzen, K. Vacuum devices for capturing and partitioning commingled subpopulations of honey bees (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 1990, 83, 1152–1154. [Google Scholar] [CrossRef]
- Fries, I.; Chauzat, M.; Chen, Y.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standard methods for Nosema research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Hamiduzzaman, M.M.; Guzman-Novoa, E.; Goodwin, P.H. A multiplex PCR assay to diagnose and quantify Nosema infections in honey Bees (Apis mellifera). J. Invertebr. Pathol. 2010, 105, 151–155. [Google Scholar] [CrossRef]
- World Organization for Animal Health (OIE) Terrestrial Manual 2018 Section 3.2.4. Nosemosis of Honey Bees. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.02.04_NOSEMOSIS_FINAL.pdf (accessed on 6 August 2020).
- Copley, T.R.; Chen, H.; Giovenazzo, P.; Houle, E.; Jabaji, S.H. Prevalence and seasonality of Nosema species in Québec honey bees. Can. Entomol. 2012, 144, 577–588. [Google Scholar] [CrossRef]
- McGowan, J.; De la Mora, A.; Goodwin, P.H.; Habash, M.; Hamiduzzaman, M.M.; Kelly, P.G.; Guzman-Novoa, E. Viability and infectivity of fresh and cryopreserved Nosema ceranae spores. J. Microbiol. Meth. 2016, 131, 16–22. [Google Scholar] [CrossRef]
- Fenoy, S.; Rueda, C.; Higes, M.; Martín-Hernández, R.; del Aguila, C. High-level resistance of Nosema ceranae, a parasite of the honeybee, to temperature and desiccation. Appl. Environ. Microbiol. 2009, 75, 6886–6889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, A.T.; Robertson, L.J.; Smith, H.V. Viability of Cryptosporidium parvum Oocysts: Correlation of in vitro excystation with inclusion or exclusion of fluorogenic vital dyes. Appl. Environ. Microbiol. 1992, 58, 3488–3493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaplane, K.S.; van der Steen, J.; Guzman-Novoa, E. Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Jaycox, E.R. Laboratory Diagnosis of Bee Diseases; Publication H-688; University of Illinois: Urbana, IL, USA, 1977. [Google Scholar]
- Ontario Treatment Recommendations for Honey Bee Disease and Mite Control. Available online: http://www.omafra.gov.on.ca/english/food/inspection/bees/2017-treatment.htm (accessed on 10 August 2020).
- Mortensen, A.N.; Jack, C.J.; McConnell, M.; Teigen, L.; Ellis, J. How to Quantify Nosema Spores Infection Rate in a Honey Bee Colony; Bulletin ENY-167; University of Florida Extension. Available online: https://edis.ifas.ufl.edu/in1123 (accessed on 10 August 2020).
- Woyke, J. Correlations and interactions between population, length of worker life and honey production by honeybees in a temperate region. J. Apic. Res. 1984, 23, 148–156. [Google Scholar] [CrossRef]
- Szabo, T.I.; Lefkovitch, L.P. Effect of brood production and population size on honey production of honeybee colonies in Alberta, Canada. Apidologie 1989, 20, 157–163. [Google Scholar] [CrossRef]
- Guzman-Novoa, E.; Gary, N.E. Genotypic variability of components of foraging behavior in honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1993, 86, 715–721. [Google Scholar] [CrossRef]
- Unger, P.; Guzman-Novoa, E. Honey bee (Hymenoptera: Apidae) guarding behaviour may not be a rare task. Can. Entomol. 2012, 144, 538–541. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org/ (accessed on 10 August 2020).
- Jack, C.J.; Lucas, H.M.; Webster, T.C.; Sagili, R.R. Colony level prevalence and intensity of Nosema ceranae in Honey bees (Apis mellifera L.). PLoS ONE 2016, 11, e0163522. [Google Scholar] [CrossRef] [Green Version]
- Pickard, R.S.; El-Shemy, A.A.M. Seasonal variation in the infection of honeybee colonies with Nosema apis Zander. J. Apic. Res. 1989, 28, 93–100. [Google Scholar] [CrossRef]
- Smith, M.L. The Honey bee parasite Nosema ceranae: Transmissible via food exchange? PLoS ONE 2012, 7, e43319. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.-F.; Solter, L.F. Comparative development and tissue tropism of Nosema apis and Nosema ceranae. J. Invertebr. Pathol. 2013, 113, 35–41. [Google Scholar] [CrossRef]
- Williams, G.R.; Shutler, D.; Little, C.M.; Burgher-Maclellan, K.L.; Rogers, R.E.L. The microsporidian Nosema ceranae, the antibiotic fumagilin-B®, and western honey bee (Apis mellifera) colony strength. Apidologie 2011, 42, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Botías, C.; Martín-Hernández, R.; Barrios, L.; Meana, A.; Higes, M. Nosema spp. infection and its negative effects on honey bees (Apis mellifera Iberiensis) at the colony level. Vet. Res. 2013, 44, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villa, J.D.; Bourgeois, A.L.; Danka, R.G. Negative evidence for effects of genetic origin of bees on Nosema ceranae, positive evidence for effects of Nosema ceranae on bees. Apidologie 2013, 44, 511–518. [Google Scholar] [CrossRef]
- Becerra-Guzmán, F.; Guzman-Novoa, E.; Correa-Benítez, A.; Zozaya-Rubio, A. Length of life, age at first foraging and foraging life of Africanized and European honey bee (Apis mellifera) workers, during conditions of resource abundance. J. Apic. Res. 2005, 44, 151–156. [Google Scholar] [CrossRef]
- Eiri, D.M.; Suwannapong, G.; Endler, M.; Nieh, J.C. Nosema ceranae can infect honey bee larvae and reduces subsequent adult longevity. PLoS ONE 2015, 10, e0126330. [Google Scholar] [CrossRef] [Green Version]
- Mattila, H.R.; Otis, G.W. Effects of pollen availability and Nosema infection during the spring on division of labor and survival of worker honey bees (Hymenoptera: Apidae). Environ. Entomol. 2006, 35, 708–717. [Google Scholar] [CrossRef]
- Dussaubat, C.; Maisonnasse, A.; Crauser, D.; Beslay, D.; Costagliola, G.; Soubeyrand, S.; Kretzchmar, A.; Le Conte, Y. Flight behavior and pheromone changes associated to Nosema ceranae infection of honey bee workers (Apis mellifera) in field conditions. J. Invertebr. Pathol. 2013, 113, 42–51. [Google Scholar] [CrossRef]
- Zheng, H.-Q.; Lin, Z.-G.; Huang, S.-K.; Sohr, A.; Wu, L.; Chen, Y.P. Spore loads may not be used alone as a direct indicator of the severity of Nosema ceranae infection in honey bees Apis mellifera (Hymenoptera:Apidae). J. Econ. Entomol. 2014, 107, 2037–2044. [Google Scholar] [CrossRef]
- Dussaubat, C.; Brunet, J.L.; Higes, M.; Colbourne, J.K.; Lopez, J.; Choi, J.H.; Martín-Hernández, R.; Botías, C.; Cousin, M.; McDonnell, C.; et al. Gut pathology and responses to the microsporidium Nosema ceranae in the honey bee Apis mellifera. PLoS ONE 2012, 7, e37017. [Google Scholar] [CrossRef] [Green Version]
- Kevan, P.G.; Guzman-Novoa, E.; Skinner, A.; Englesdorp, D.V. Colony Collapse Disorder (CCD) in Canada: Do we have a problem? Hivelights 2007, 20, 14–16. [Google Scholar]
- Williams, G.R.; Tarpy, D.R.; vanEngelsdorp, D.; Chauzat, M.P.; Cox-Foster, D.L.; Delaplane, K.S.; Neumann, P.; Pettis, J.S.; Rogers, R.E.L.; Shutler, D. Colony Collapse Disorder in context. BioEssays 2010, 32, 845–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Novoa, E. Colony collapse disorder and other threats to honey bees. In One Health Case Studies: Addressing Complex. Problems in a Changing World; Cork, S., Hall, D.C., Liljebjelke, K., Eds.; 5M Publishing Ltd.: Sheffielf, UK, 2016; pp. 204–216. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Season | % Infected Bees ± SE | % Spore Viability ± SE |
|---|---|---|
| Spring | 73.9 ± 1.8 a | 59.0 ± 6.0 a |
| Summer | 68.0 ± 1.8 a | 13.7 ± 1.6 b |
| Fall | 48.0 ± 5.6 b | 16.3 ± 1.8 b |
| Infection Category | No. Spores Per Bee 1 | No. Frames with Bees | No. Frames with Brood | No. Frames with Pollen | No. Frames with Honey |
|---|---|---|---|---|---|
| Low | <1.0 | 19.7 ± 1.0 a | 8.4 ± 0.3 a | 2.1 ± 0.1 a | 14.1 ± 1.2 a |
| Threshold | 1.0–2.0 | 17.9 ± 1.3 a,b | 6.9 ± 0.6 b | 1.7 ± 0.2 a,b | 9.0 ± 1.3 b |
| High | >2.0 | 14.3 ± 2.6 b | 6.7 ± 1.4 b | 1.4 ± 0.3 b | 4.6 ± 1.6 c |
| Variables Correlated | Correlation | p |
|---|---|---|
| No. spores–Frames w/bees | −0.33 | <0.0001 |
| No. spores–Frames w/brood | −0.31 | <0.0001 |
| No. spores–Frames w/pollen | −0.24 | 0.0024 |
| No. spores–Frames w/honey | −0.52 | <0.0001 |
| No. spores–Colony growth | −0.18 | <0.05 |
| Frames w/bees–Frames w/brood | 0.56 | <0.0001 |
| Frames w/bees–Frames w/pollen | 0.30 | 0.0002 |
| Frames w/bees–Frames w/honey | 0.74 | <0.0001 |
| Frames w/bees–Colony growth | 0.96 | <0.0001 |
| Frames w/brood–Frames w/pollen | 0.02 | 0.7678 |
| Frames w/brood–Frames w/honey | 0.47 | <0.0001 |
| Frames w/brood–Colony growth | 0.38 | <0.0001 |
| Frames w/pollen–Frames w/honey | 0.44 | <0.0001 |
| Frames w/pollen–Colony growth | 0.23 | <0.01 |
| Frames w/honey–Colony growth | 0.79 | <0.01 |
| Variables | Low | Threshold | High |
|---|---|---|---|
| No. spores/bee 1 | 1.88 ± 0.25 | 1.75 ± 0.61 | 3.21 ± 1.58 |
| No. Frames w/bees | 6.3 ± 0.5 | 6.3 ± 0.9 | 5.5 ± 1.3 |
| No. Frames w/brood | 3.4 ± 0.3 | 4.1 ± 0.6 | 3.0 ± 0.5 |
| No. Frames w/pollen | 1.9 ± 0.2 | 2.1 ± 0.2 | 1.9 ± 0.9 |
| No. Frames w/honey | 8.3 ± 0.4 | 8.2 ± 0.7 | 9.7 ± 0.7 |
| % Colony survival | 64.2 | 55.6 | 43.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emsen, B.; De la Mora, A.; Lacey, B.; Eccles, L.; Kelly, P.G.; Medina-Flores, C.A.; Petukhova, T.; Morfin, N.; Guzman-Novoa, E. Seasonality of Nosema ceranae Infections and Their Relationship with Honey Bee Populations, Food Stores, and Survivorship in a North American Region. Vet. Sci. 2020, 7, 131. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci7030131
Emsen B, De la Mora A, Lacey B, Eccles L, Kelly PG, Medina-Flores CA, Petukhova T, Morfin N, Guzman-Novoa E. Seasonality of Nosema ceranae Infections and Their Relationship with Honey Bee Populations, Food Stores, and Survivorship in a North American Region. Veterinary Sciences. 2020; 7(3):131. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci7030131
Chicago/Turabian StyleEmsen, Berna, Alvaro De la Mora, Brian Lacey, Les Eccles, Paul G. Kelly, Carlos A. Medina-Flores, Tatiana Petukhova, Nuria Morfin, and Ernesto Guzman-Novoa. 2020. "Seasonality of Nosema ceranae Infections and Their Relationship with Honey Bee Populations, Food Stores, and Survivorship in a North American Region" Veterinary Sciences 7, no. 3: 131. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci7030131