
Addition of Reduced Glutathione (GSH) to Freezing Medium Reduces Intracellular ROS Levels in Donkey Sperm

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Donkeys and Sperm Samples
2.2. Experimental Design
2.3. Sperm Cryopreservation
2.4. Sperm Thawing
2.5. Sperm Motility Analysis
2.6. Sperm Viability Analysis
2.7. Evaluation of Sperm through Flow Cytometry
2.7.1. Plasma Membrane Integrity Analysis (SYBR-14/PI)
2.7.2. Acrosome Membrane Integrity Analysis (PNA-FITC/PI)
2.7.3. Mitochondrial Membrane Potential (MMP) Analysis (JC-1)
2.7.4. Intracellular Reactive Oxygen Species (ROS) Levels Analysis: H2O2 (H2DCFDA/PI) and O2− (HE/YO-PRO-1)
2.8. Statistical Analyses
3. Results
3.1. Sperm Motility
3.2. Motile Sperm Subpopulations
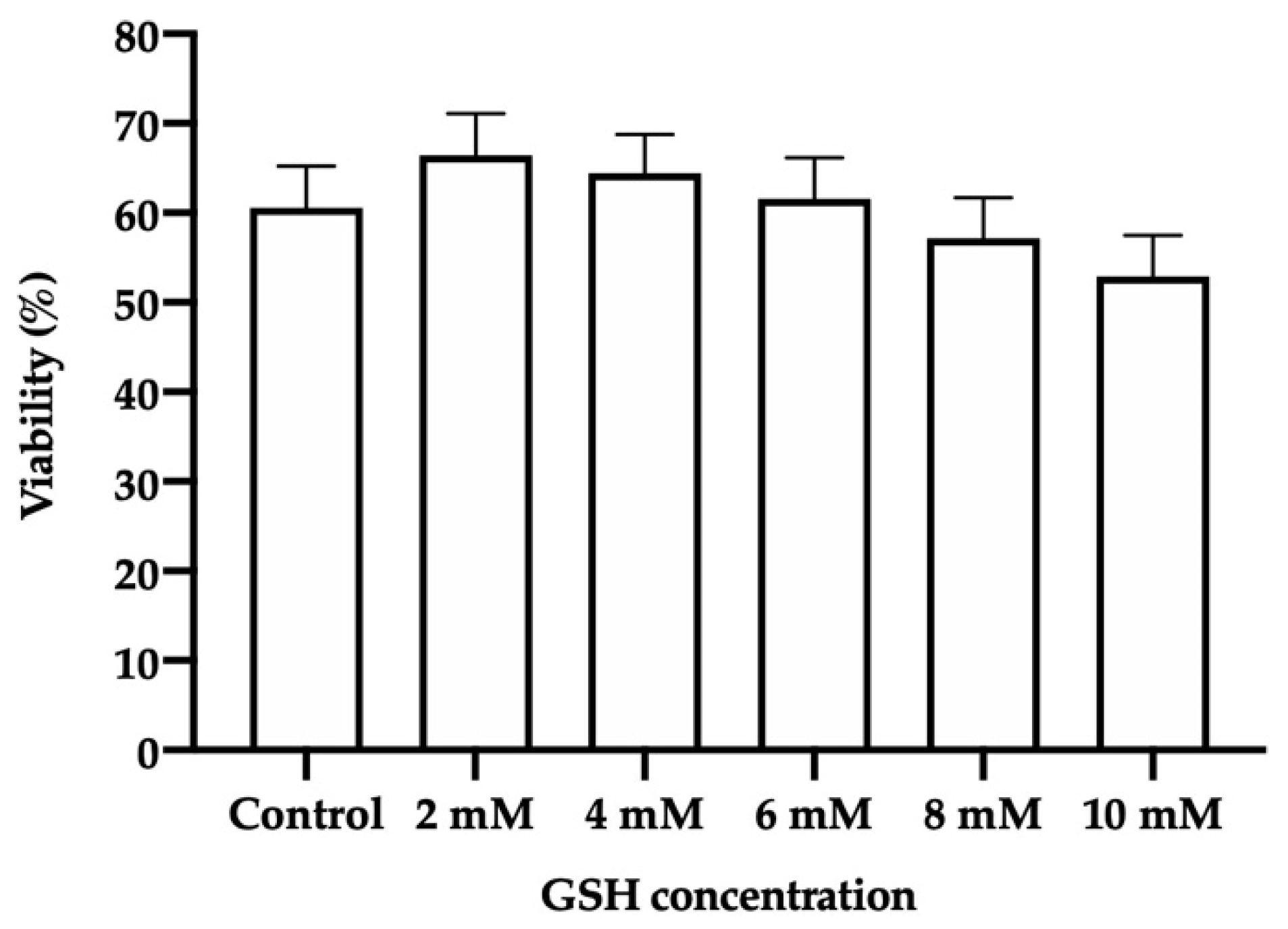
3.3. Sperm Viability
3.4. Sperm Quality
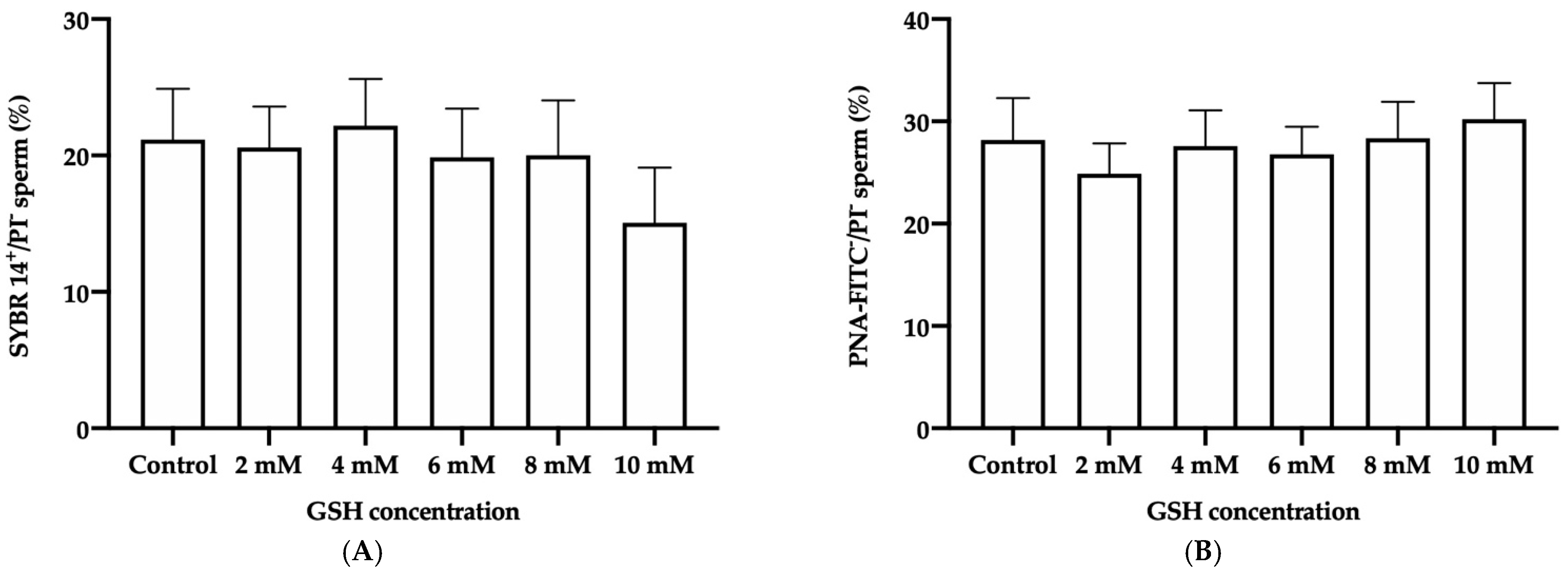
3.4.1. Sperm Membrane Integrity
3.4.2. Acrosome Membrane Integrity
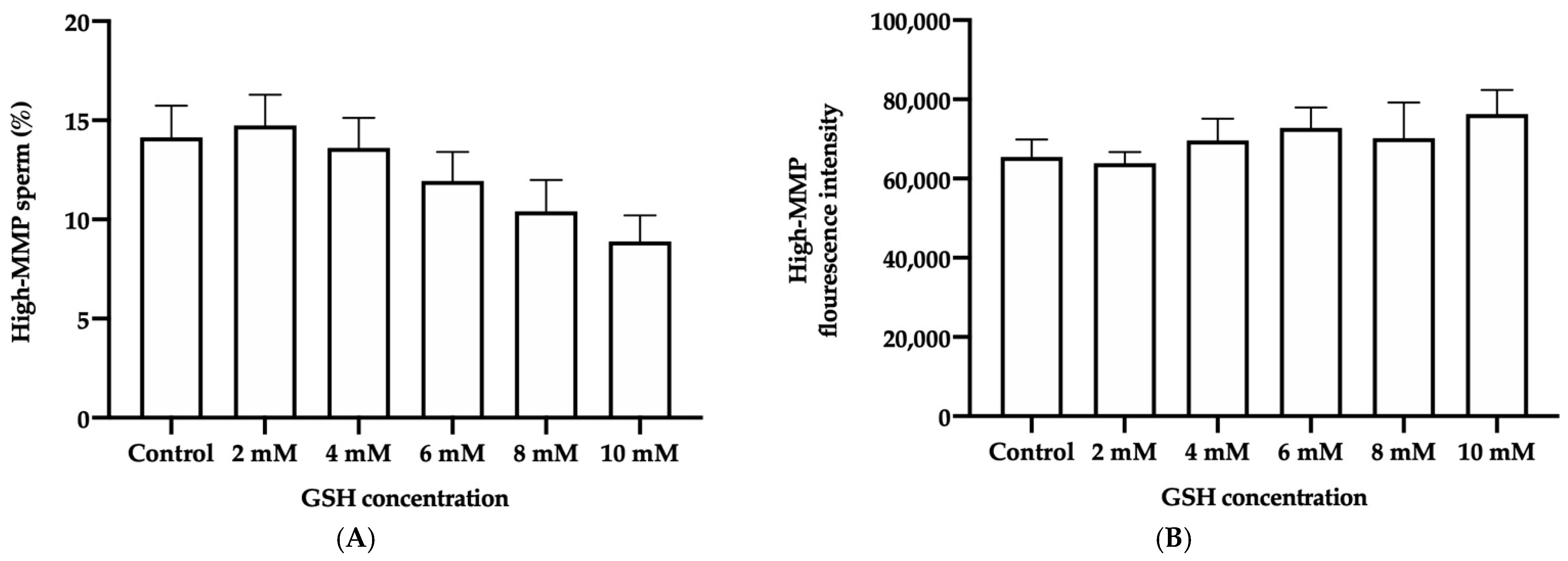
3.4.3. Mitochondrial Membrane Potential (MPP)
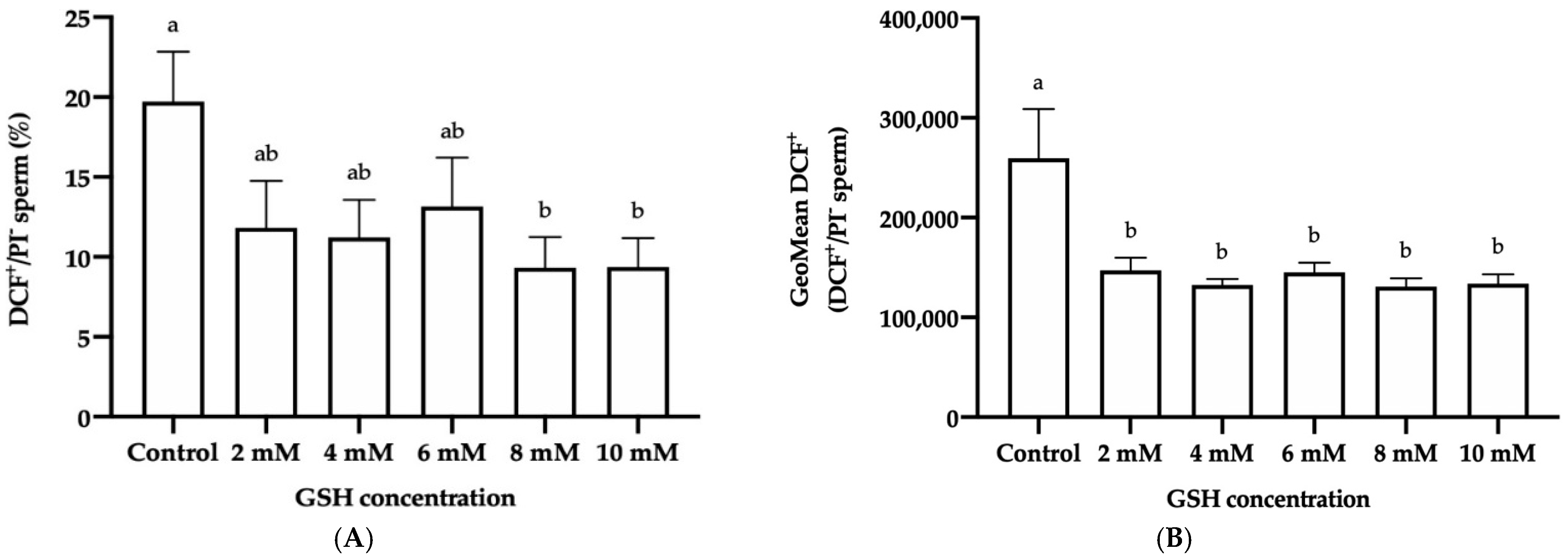
3.4.4. Intracellular Reactive Oxygen Species (ROS) Levels: H2O2 and O2−
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sherman, J.K. Low temperature research on spermatozoa and eggs. Cryobiology 1964, 1, 103–129. [Google Scholar] [CrossRef]
- Loomis, P.R. Advanced methods for handling and preparation of stallion semen. Vet. Clin. N. Am. Equine Pract. 2006, 22, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Fickel, J.; Wagener, A.; Ludwig, A. Semen cryopreservation and the conservation of endangered species. Eur. J. Wildl. Res. 2007, 53, 81–89. [Google Scholar] [CrossRef]
- Camillo, F.; Rota, A.; Biagini, L.; Tesi, M.; Fanelli, D.; Panzani, D. The current situation and trend of donkey industry in Europe. J. Equine Vet. Sci. 2018, 65, 44–49. [Google Scholar] [CrossRef]
- Aranguren-Méndez, J.; Jordana, J.; Gomez, M. Genetic diversity in Spanish donkey breeds using microsatellite DNA markers. Genet. Sel. Evol. 2001, 33, 1–10. [Google Scholar] [CrossRef]
- Trimeche, A.; Renard, P.; Tainturier, D. A procedure for Poitou jackass sperm cryopreservation. Theriogenology 1998, 50, 793–806. [Google Scholar] [CrossRef]
- Vidament, M.; Vincent, P.; Martin, F.X.; Magistrini, M.; Blesbois, E. Differences in ability of jennies and mares to conceive with cooled and frozen semen containing glycerol or not. Anim. Reprod. Sci. 2009, 112, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.V.; Papa, F.O.; Melo-Oña, C.M.; Monteiro, G.A.; Puoli-Filho, J.N.P.; Alvarenga, M.A. New procedures to freeze donkey semen and its influence on mares and jennies fertility. J. Equine Vet. Sci. 2012, 32, 503–504. [Google Scholar] [CrossRef]
- Rota, A.; Panzani, D.; Sabatini, C.; Camillo, F. Donkey jack (Equus asinus) semen cryopreservation: Studies of seminal parameters, post breeding inflammatory response, and fertility in donkey jennies. Theriogenology 2012, 78, 1846–1854. [Google Scholar] [CrossRef]
- Pukazhenthi, B.S.; Johnson, A.; Guthrie, H.D.; Songsasen, N.; Padilla, L.R.; Wolfe, B.A.; Coutinho da Silva, M.; Alvarenga, M.A.; Wildt, D.E. Improved sperm cryosurvival in diluents containing amides versus glycerol in the Przewalski’s horse (Equus ferus przewalskii). Cryobiology 2014, 68, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, I.; Dorado, J.; Morrell, J.M.; Crespo, F.; Gosálvez, J.; Gálvez, M.J.; Acha, D.; Hidalgo, M. Effect of single-layer centrifugation or washing on frozen–thawed donkey semen quality: Do they have the same effect regardless of the quality of the sample? Theriogenology 2015, 84, 294–300. [Google Scholar] [CrossRef]
- Acha, D.; Hidalgo, M.; Ortiz, I.; Gálvez, M.J.; Carrasco, J.J.; Gómez-Arrones, V.; Dorado, J. Freezability of Andalusian donkey (Equus asinus) spermatozoa: Effect of extenders and permeating cryoprotectants. Reprod. Fertil. Dev. 2016, 28, 1990–1998. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.V.; Oliveira, P.V.L.F.; Melo-Oña, C.M.; Guasti, P.N.; Monteiro, G.A.; Sancler da Silva, Y.F.R.; Papa, P.M.; Alvarenga, M.A.; Dell’Aqua, J.A.; Papa, F.O. Strategies to improve the fertility of fresh and frozen donkey semen. Theriogenology 2016, 85, 1267–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottrel, M.; Acha, D.; Ortiz, I.; Hidalgo, M.; Gósalvez, J.; Camisão, J.; Dorado, J. Cryoprotective effect of glutamine, taurine, and proline on post-thaw semen quality and DNA integrity of donkey spermatozoa. Anim. Reprod. Sci. 2018, 189, 128–135. [Google Scholar] [CrossRef]
- Zhang, H.; Ye, H.; Shao, Y.; Wu, S.; Yu, J.; Ji, C.; Wang, S.; Zeng, S. The effects of egg yolk concentration and particle size on donkey semen preservation. J. Equine Vet. Sci. 2018, 65, 19–24. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, R.; Mehta, J.S.; Chaudhary, A.K.; Ravi, S.K.; Chandra Mehta, S.; Ansari, M.M.; Legha, R.A.; Tripathi, B.N.; Talluri, T.R. Ameliorative effect of ascorbic acid and glutathione in combating the cryoinjuries during cryopreservation of exotic jack semen. J. Equine Vet. Sci. 2019, 81, 102796. [Google Scholar] [CrossRef]
- Yu, X.; He, S.; Wang, L.; Kang, M.; Zhu, Y.; Wang, S.; Sun, X. Effects of Vitamin C and Vitamin E on cryopreservation of Guanzhong donkey semen. Pak. J. Zool. 2019, 51, 1777–1781. [Google Scholar] [CrossRef]
- Li, N.; Yu, J.; Yang, F.; Shao, Y.; Wu, S.; Liu, B.; Li, M.; Wang, T.; Li, J.; Zeng, S. L-proline: An effective agent for frozen and post-thawed donkey semen storage. J. Equine Vet. Sci. 2021, 101, 103393. [Google Scholar] [CrossRef] [PubMed]
- Canisso, I.F.; Carvalho, G.R.; Davis Morel, M.; Ker, P.G.; Rodrigues, A.L.; Silva, E.C.; Coutinho Da Silva, M.A. Seminal parameters and field fertility of cryopreserved donkey jack semen after insemination of horse mares. Equine Vet. J. 2011, 43, 179–183. [Google Scholar] [CrossRef]
- Vilés, K.; Rabanal, R.; Rodríguez-Prado, M.; Miró, J. Effect of ketoprofen treatment on the uterine inflammatory response after AI of jennies with frozen semen. Theriogenology 2013, 79, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Miró, J.; Papas, M. Post–Artificial Insemination endometrial inflammation and its control in donkeys. J. Equine Vet. Sci. 2018, 65, 38–43. [Google Scholar] [CrossRef]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; De Iuliis, G.N.; Drevet, J.R. Role of oxidative stress in the etiology of male infertility and the potential therapeutic value of antioxidants. In Oxidants, Antioxidants, and Impact of the Oxidative Status in Male Reproduction; Elsevier: London, UK, 2018; pp. 91–100. [Google Scholar]
- Peña, F.J.; O’Flaherty, C.; Ortiz Rodríguez, J.M.; Martín Cano, F.E.; Gaitskell-Phillips, G.L.; Gil, M.C.; Ortega Ferrusola, C. Redox regulation and oxidative stress: The particular case of the stallion spermatozoa. Antioxidants 2019, 8, 567. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.G.; Parthasarathy, S. Oxidation and the spermatozoa. Semin. Reprod. Endocrinol. 1998, 16, 235–239. [Google Scholar] [CrossRef]
- Papas, M.; Arroyo, L.; Bassols, A.; Catalán, J.; Bonilla-Correal, S.; Gacem, S.; Yeste, M.; Miró, J. Activities of antioxidant seminal plasma enzymes (SOD, CAT, GPX and GSR) are higher in jackasses than in stallions and are correlated with sperm motility in jackasses. Theriogenology 2019, 140, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Papas, M.; Catalán, J.; Recuero, S.; Morrell, J.M.; Yeste, M.; Miró, J. Single layer centrifugation improves the quality of fresh donkey semen and modifies the sperm ability to interact with polymorphonuclear neutrophils. Animals 2020, 10, 2128. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review on evaluation and management. Arab J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M.; Estrada, E.; Casas, I.; Bonet, S.; Rodríguez-Gil, J.E. Good and bad freezability boar ejaculates differ in the integrity of nucleoprotein structure after freeze-thawing but not in ROS levels. Theriogenology 2013, 79, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Kotilainen, T.; Huhtinen, M.; Katila, T. Sperm-induced leukocytosis in the equine uterus. Theriogenology 1994, 41, 629–636. [Google Scholar] [CrossRef]
- Troedsson, M.H.T.; Loset, K.; Alghamdi, A.M.; Dahms, B.; Crabo, B.G. Interaction between equine semen and the endometrium: The inflammatory response to semen. Anim. Reprod. Sci. 2001, 68, 273–278. [Google Scholar] [CrossRef]
- Rodriguez-Martinez, H.; Saravia, F.; Wallgren, M.; Martinez, E.A.; Sanz, L.; Roca, J.; Vazquez, J.M.; Calvete, J.J. Spermadhesin PSP-I/PSP-II heterodimer induces migration of polymorphonuclear neutrophils into the uterine cavity of the sow. J. Reprod. Immunol. 2010, 84, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Zambrano, F.; Carrau, T.; Gärtner, U.; Seipp, A.; Taubert, A.; Felmer, R.; Sanchez, R.; Hermosilla, C. Leukocytes coincubated with human sperm trigger classic neutrophil extracellular traps formation, reducing sperm motility. Fertil. Steril. 2016, 106, 1053–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miró, J.; Marín, H.; Catalán, J.; Papas, M.; Gacem, S.; Yeste, M. Seminal plasma, sperm concentration, and sperm-PMN interaction in the donkey: An in vitro model to study endometrial inflammation at post-insemination. Int. J. Mol. Sci. 2020, 21, 3478. [Google Scholar] [CrossRef]
- Baumber, J.; Vo, A.; Sabeur, K.; Ball, B.A. Generation of reactive oxygen species by equine neutrophils and their effect on motility of equine spermatozoa. Theriogenology 2002, 57, 1025–1033. [Google Scholar] [CrossRef]
- Yánez-Ortiz, I.; Catalán, J.; Mateo-Otero, Y.; Dordas-Perpinyà, M.; Gacem, S.; Yeste, N.; Bassols, A.; Yeste, M.; Miró, J. Extracellular reactive oxygen species (ROS) production in fresh donkey sperm exposed to reductive stress, oxidative stress and NETosis. Antioxidants 2021, 10, 1367. [Google Scholar] [CrossRef]
- Gadea, J.; Molla, M.; Selles, E.; Marco, M.A.; Garcia-Vazquez, F.A.; Gardon, J.C. Reduced glutathione content in human sperm is decreased after cryopreservation: Effect of the addition of reduced glutathione to the freezing and thawing extenders. Cryobiology 2011, 62, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Câmara, D.R.; Mello-Pinto, M.M.C.; Pinto, L.C.; Brasil, O.O.; Nunes, J.F.; Guerra, M.M.P. Effects of reduced glutathione and catalase on the kinematics and membrane functionality of sperm during liquid storage of ram semen. Small Rumin. Res. 2011, 100, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.S.; Rakha, B.A.; Andrabi, S.M.H.; Ullah, N.; Iqbal, R.; Holt, W.V.; Akhter, S. Glutathione-supplemented tris-citric acid extender improves the post-thaw quality and in vivo fertility of buffalo (Bubalus bubalis) bull spermatozoa. Reprod. Biol. 2012, 12, 271–276. [Google Scholar] [CrossRef]
- Oliveira, R.A.; Piersanti, R.L.; Wolf, C.A.; Viu, M.A.O.; Gambarini, M.L. Glutathione for the freezing of cooled equine semen, using different protocols. Anim. Reprod. 2018, 11, 104–109. [Google Scholar]
- Ahmad, E.; Naseer, Z.; Aksoy, M. Glutathione supplementation in semen extender improves rabbit sperm attributes during refrigeration. World Rabbit Sci. 2021, 29, 81. [Google Scholar] [CrossRef]
- Gadea, J.; García-Vazquez, F.; Matás, C.; Gardón, J.C.; Cánovas, S.; Gumbao, D. Cooling and freezing of boar spermatozoa: Supplementation of the freezing media with reduced glutathione preserves sperm function. J. Androl. 2005, 26, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.V.; Soares, A.T.; Batista, A.M.; Almeida, F.C.; Nunes, J.F.; Peixoto, C.A.; Guerra, M.M.P. In vitro and in vivo evaluation of ram sperm frozen in tris egg-yolk and supplemented with superoxide dismutase and reduced glutathione. Reprod. Domest. Anim. 2011, 46, 874–881. [Google Scholar] [CrossRef]
- Oliveira, R.A.; Wolf, C.A.; Viu, M.A.O.; Gambarini, M.L. Addition of glutathione to an extender for frozen equine semen. J. Equine Vet. Sci. 2013, 33, 1148–1152. [Google Scholar] [CrossRef] [Green Version]
- Ogata, K.; Sasaki, A.; Kato, Y.; Takeda, A.; Wakabayashi, M.; Sarentonglaga, B.; Yamaguchi, M.; Hara, A.; Fukumori, R.; Nagao, Y. Glutathione supplementation to semen extender improves the quality of frozen-thawed canine spermatozoa for transcervical insemination. J. Reprod. Dev. 2015, 61, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangwar, C.; Saxena, A.; Patel, A.; Singh, S.P.; Yadav, S.; Kumar, R.; Singh, V. Effect of reduced glutathione supplementation on cryopreservation induced sperm cryoinjuries in Murrah bull semen. Anim. Reprod. Sci. 2018, 192, 171–178. [Google Scholar] [CrossRef]
- Luberda, Z. The role of glutathione in mammalian gametes. Reprod. Biol. 2005, 5, 5–17. [Google Scholar]
- Irvine, D.S. Glutathione as a treatment for male infertility. Rev. Reprod. 1996, 1, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M.; Flores, E.; Estrada, E.; Bonet, S.; Rigau, T.; Rodríguez-Gil, J.E. Reduced glutathione and procaine hydrochloride protect the nucleoprotein structure of boar spermatozoa during freeze-thawing by stabilising disulfide bonds. Reprod. Fertil. Dev. 2013, 25, 1036–1050. [Google Scholar] [CrossRef] [Green Version]
- Miró, J.; Lobo, V.; Quintero-Moreno, A.; Medrano, A.; Peña, A.; Rigau, T. Sperm motility patterns and metabolism in Catalonian donkey semen. Theriogenology 2005, 63, 1706–1716. [Google Scholar] [CrossRef]
- Canisso, I.F.; Panzani, D.; Miró, J.; Ellerbrock, R.E. Key aspects of donkey and mule reproduction. Vet. Clin. N. Am. Equine Pract. 2019, 35, 607–642. [Google Scholar] [CrossRef]
- Gacem, S.; Papas, M.; Catalan, J.; Miró, J. Examination of jackass (Equus asinus) accessory sex glands by B-mode ultrasound and of testicular artery blood flow by colour pulsed-wave Doppler ultrasound: Correlations with semen production. Reprod. Domest. Anim. 2020, 55, 181–188. [Google Scholar] [CrossRef]
- Kenney, M.R. Minimal contamination techniques for breeding mares: Techniques and priliminary findings. Proc. Am. Assoc. Equine Pract. 1975, 327–336. [Google Scholar]
- Gacem, S.; Catalán, J.; Valverde, A.; Soler, C.; Miró, J. Optimization of CASA-mot analysis of donkey sperm: Optimum frame rate and values of kinematic variables for different counting chamber and fields. Animals 2020, 10, 1993. [Google Scholar] [CrossRef]
- Bamba, K. Evaluation of acrosomal integrity of boar spermatozoa by bright field microscopy using an eosin-nigrosin stain. Theriogenology 1988, 29, 1245–1251. [Google Scholar] [CrossRef]
- Lee, J.A.; Spidlen, J.; Boyce, K.; Cai, J.; Crosbie, N.; Dalphin, M.; Furlong, J.; Gasparetto, M.; Goldberg, M.; Goralczyk, E.M.; et al. MIFlowCyt: The minimum information about a flow cytometry experiment. Cytometry 2008, 73, 926–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Rathi, R.; Colenbrander, B.; Bevers, M.M.; Gadella, B.M. Evaluation of in vitro capacitation of stallion spermatozoa. Biol. Reprod. 2001, 65, 462–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega-Ferrusola, C.; Sotillo-Galán, Y.; Varela-Fernández, E.; Gallardo-Bolaños, J.M.; Muriel, A.; González-Fernández, L.; Tapia, J.A.; Peña, F.J. Detection of “apoptosis-like” changes during the cryopreservation process in equine sperm. J. Androl. 2008, 29, 213–221. [Google Scholar] [CrossRef]
- Morrell, J.M.; Winblad, C.; Georgakas, A.; Stuhtmann, G.; Humblot, P.; Johannisson, A. Reactive oxygen species in stallion semen can be affected by season and colloid centrifugation. Anim. Reprod. Sci. 2013, 140, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, H.D.; Welch, G.R. Determination of intracellular reactive oxygen species and high mitochondrial membrane potential in Percoll-treated viable boar sperm using fluorescence-activated flow cytometry. J. Anim. Sci. 2006, 84, 2089–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martí, J.I.; Aparicio, I.M.; García-Herreros, M. Sperm morphometric subpopulations are differentially distributed in rams with different maturity age in cryopreserved ejaculates. Theriogenology 2011, 76, 97–109. [Google Scholar] [CrossRef]
- Meyers, S.A. Sperm physiology. In Equine Breeding Management and Artificial Insemination; Samper, J.C., Ed.; Saunders Elsevier: Philadelphia, PA, USA, 2009; pp. 47–55. [Google Scholar]
- Rodríguez-Martínez, H.; Kvist, U.; Ernerudh, J.; Sanz, L.; Calvete, J.J. Seminal plasma proteins: What role do they play? Am. J. Reprod. Immunol. 2011, 66, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadea, J.; Sellés, E.; Marco, M.A.; Coy, P.; Matás, C.; Romar, R.; Ruiz, S. Decrease in glutathione content in boar sperm after cryopreservation: Effect of the addition of reduced glutathione to the freezing and thawing extenders. Theriogenology 2004, 62, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ferrusola, C.; Martin Muñoz, P.; Ortiz-Rodriguez, J.M.; Anel-López, L.; Balao da Silva, C.; Álvarez, M.; de Paz, P.; Tapia, J.A.; Anel, L.; Silva- Rodríguez, A.; et al. Depletion of thiols leads to redox deregulation, production of 4-hydroxinonenal and sperm senescence: A possible role for GSH regulation in spermatozoa. Biol. Reprod. 2019, 100, 1090–1107. [Google Scholar] [CrossRef]
- Catalán, J.; Papas, M.; Trujillo-Rojas, L.; Blanco-Prieto, O.; Bonilla-Correal, S.; Rodríguez-Gil, J.E.; Miró, J.; Yeste, M. Red LED light acts on the mitochondrial electron chain of donkey sperm and its effects depend on the time of exposure to light. Front. Cell Dev. Biol. 2020, 8, 588621. [Google Scholar] [CrossRef] [PubMed]
- Estrada, E.; Rivera del Álamo, M.M.; Rodríguez-Gil, J.E.; Yeste, M. The addition of reduced glutathione to cryopreservation media induces changes in the structure of motile subpopulations of frozen-thawed boar sperm. Cryobiology 2017, 78, 56–64. [Google Scholar] [CrossRef]
- Estrada, E.; Rodríguez-Gil, J.E.; Rocha, L.G.; Balasch, S.; Bonet, S.; Yeste, M. Supplementing cryopreservation media with reduced glutathione increases fertility and prolificacy of sows inseminated with frozen-thawed boar semen. Andrology 2014, 2, 88–99. [Google Scholar] [CrossRef]
- Ball, B.A.; Vo, A.T.; Baumber, J. Generation of reactive oxygen species by equine spermatozoa. Am. J. Vet. Res. 2001, 62, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, J.F.; Blanchette, S.; Gagnon, C.; Sirard, M.A. Thiols prevent H2O2-mediated loss of sperm motility in cryopreserved bull semen. Theriogenology 2001, 56, 275–286. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide radical and superoxide dismutases. Annu. Rev. Biochem. 1995, 64, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Hammerstedt, R.H. Maintenance of bioenergetic balance in sperm and prevention of lipid peroxidation: A review of the effect on design of storage preservation systems. Reprod. Fertil. Dev. 1993, 5, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Holt, W.V. Mechanisms of sperm storage in the female reproductive tract: An interspecies comparison. Reprod. Domest. Anim. 2011, 46, 68–74. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | GSH Concentration | |||||
|---|---|---|---|---|---|---|
| Control | 2 mM | 4 mM | 6 mM | 8 mM | 10 mM | |
| VCL (µm/s) | 134.66 ± 8.53 a | 152.04 ± 6.74 a | 139.16 ± 7.43 a | 138.69 ± 9.47 a | 114.97 ± 13.76 a | 57.03 ± 18.80 b |
| VSL (µm/s) | 53.16 ± 4.55 a | 60.51 ± 4.09 a | 52.71 ± 4.11 a | 50.47 ± 5.20 a | 30.14 ± 4.76 b | 13.53 ± 5.08 b |
| VAP (µm/s) | 69.42 ± 4.75 a | 76.22 ± 3.75 a | 70.92 ± 2.98 a | 68.26 ± 4.51 a | 53.05 ± 5.95 a | 27.23 ± 8.54 b |
| LIN (%) | 37.02 ± 1.20 a | 37.96 ± 1.26 a | 35.59 ± 1.29 a | 33.96 ± 1.67 a | 23.06 ± 2.61 b | 10.98 ± 3.75 c |
| STR (%) | 70.07 ± 1.91 a | 74.07 ± 2.09 a | 68.06 ± 3.21 a | 67.37 ± 3.21 a | 47.72 ± 5.40 b | 23.19 ± 7.92 c |
| WOB (%) | 51.61 ± 0.39 a | 50.65 ± 0.65 a | 51.96 ± 1.50 a | 49.95 ± 0.88 a | 44.14 ± 4.51 a | 24.70 ± 7.55 b |
| ALH (µm) | 1.81 ± 0.08 a | 2.06 ± 0.08 a | 1.92 ± 0.10 a | 1.93 ± 0.11 a | 1.62 ± 0.18 a | 0.83 ± 0.27 b |
| BCF (Hz) | 28.23 ± 2.28 a | 29.27 ± 2.19 a | 25.84 ± 1.46 a | 23.57 ± 1.91 ab | 15.19 ± 1.90 bc | 8.05 ± 2.52 c |
| Parameter | SP1 | SP2 | SP3 | SP4 | ||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SEM | Range | Mean ± SEM | Range | Mean ± SEM | Range | Mean ± SEM | Range | |
| VCL (µm/s) | 265.51 ± 0.81 | 197.90–429.30 | 69.32 ± 0.35 | 0.00–147.30 | 198.68 ± 0.35 | 127.50–276.00 | 134.67 ± 0.32 | 66.50–218.70 |
| VSL (µm/s) | 115.39 ± 0.54 | 10.80–191.30 | 15.79 ± 0.13 | 0.00–54.70 | 83.56 ± 0.30 | 3.70–152.10 | 49.42 ± 0.22 | 1.80–105.00 |
| VAP (µm/s) | 131.57 ± 0.40 | 78.60–216.50 | 33.31 ± 0.19 | 0.00–91.90 | 99.91 ± 0.22 | 44.30–165.90 | 69.05 ± 0.17 | 25.10–123.90 |
| LIN (%) | 44.13 ± 0.23 | 3.90–75.30 | 24.06 ± 0.19 | 0.00–92.30 | 42.75 ± 0.17 | 1.50–95.60 | 38.03 ± 0.19 | 1.20–100.00 |
| STR (%) | 87.42 ± 0.27 | 7.70–100.00 | 48.01 ± 0.29 | 0.00–99.20 | 83.59 ± 0.23 | 3.70–100.00 | 71.71 ± 0.25 | 2.30–100.00 |
| WOB (%) | 50.09 ± 0.18 | 27.00–85.30 | 48.88 ± 0.17 | 0.00–100.00 | 50.80 ± 0.13 | 19.20–100.00 | 52.32 ± 0.14 | 17.40–100.00 |
| ALH (µm) | 3.36 ± 0.02 | 1.50–6.40 | 1.10 ± 0.00 | 0.00–2.80 | 2.61 ± 0.01 | 0.80–4.00 | 1.83 ± 0.01 | 0.40–3.50 |
| BCF (Hz) | 36.67 ± 0.31 | 0.00–73.50 | 14.39 ± 0.10 | 0.00–52.40 | 35.29 ± 0.19 | 0.00–72.10 | 28.39 ± 0.14 | 0.00–70.80 |
| n (%) | 1776 (9.97%) | 5609 (31.50%) | 4339 (24.36%) | 6085 (34.17) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yánez-Ortiz, I.; Catalán, J.; Delgado-Bermúdez, A.; Carluccio, A.; Miró, J.; Yeste, M. Addition of Reduced Glutathione (GSH) to Freezing Medium Reduces Intracellular ROS Levels in Donkey Sperm. Vet. Sci. 2021, 8, 302. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8120302
Yánez-Ortiz I, Catalán J, Delgado-Bermúdez A, Carluccio A, Miró J, Yeste M. Addition of Reduced Glutathione (GSH) to Freezing Medium Reduces Intracellular ROS Levels in Donkey Sperm. Veterinary Sciences. 2021; 8(12):302. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8120302
Chicago/Turabian StyleYánez-Ortiz, Iván, Jaime Catalán, Ariadna Delgado-Bermúdez, Augusto Carluccio, Jordi Miró, and Marc Yeste. 2021. "Addition of Reduced Glutathione (GSH) to Freezing Medium Reduces Intracellular ROS Levels in Donkey Sperm" Veterinary Sciences 8, no. 12: 302. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8120302