
Diatoms from the Spring Ecosystems Selected for the Long-Term Monitoring of Climate-Change Effects in the Berchtesgaden National Park (Germany)

 ,
,  ,
, 
 , and
, and 
Abstract
:1. Introduction
- -
- Value for nature conservation. Cantonati et al. [11] highlighted the potential of Diatom Red Lists [6,12] which allow a characterisation of the ecological integrity and of the diatom diversity of inland water ecosystems (also of practical importance to designate the most relevant habitats for conservation purposes), including a clear assessment of the threat status of the habitat. Furthermore, they offer ample possibilities to track the effects of stressors and of environmental change. The cumulative proportion of diatom species belonging to threat categories is a good indicator of spring-habitat ecological integrity as it consistently decreases with changes such as the increase in groundwater nitrates or spring-morphology alteration [5,11]. By comparison with an extensive stream-diatom database, we could recently show [13] that, in densely populated and exploited areas, springs are the last high integrity refugium for Red-List and sensitive (oligotraphentic) diatom species (Least-Impaired Habitat Relicts concept—LIHRe, [5]).
- -
- Water quality, including factors and parameters that are particularly relevant in springs (e.g., nitrates, often used as proxies of aquifer contamination in spring studies).
- -
- Contamination by metals and trace elements. Cantonati et al. [14] showed peculiar valve deformities of the widespread species Achnanthidium minutissimum (Kütz.) Czarn. could be used as indicators of copper, zinc, and antimony natural or anthropogenic contamination.
- -
- Spring classification. Spring types can be straightforwardly identified using diatoms [10] as many species respond with high sensitivity to the main parameters used in most spring classifications (current velocity, mineral content, substrata, light conditions, etc.).
- -
- Flow variability. Diatoms live not only in the wetted perimeter of springs, but also in microhabitats that are only periodically (discharge fluctuations) or intermittently (spray zones) provided with water. Thus, the composition of diatom assemblages can provide information on hydroperiod and flow variability and persistence (e.g., [10]), which is very useful in particular in explorative hydrogeological studies.
- -
- Springs-dependent species. Though no diatom species found exclusively in springs have been identified, there are species that occur mainly in the spring-fed headwaters of carbonate streams [15], and can thus even be used as indicators of “spring conditions” as opposed to “stream conditions”.
- -
- Geological substratum. The distribution of some diatom species is influenced by the geological substratum of the aquifer feeding the spring. Achnanthidium dolomiticum Cantonati et Lange-Bert. [16], for instance, is found in springs (and other aquatic habitats) with drainage basins formed by rocks that confer to the water above-average magnesium values as in the case of dolomites and ophiolites.
2. Materials and Methods
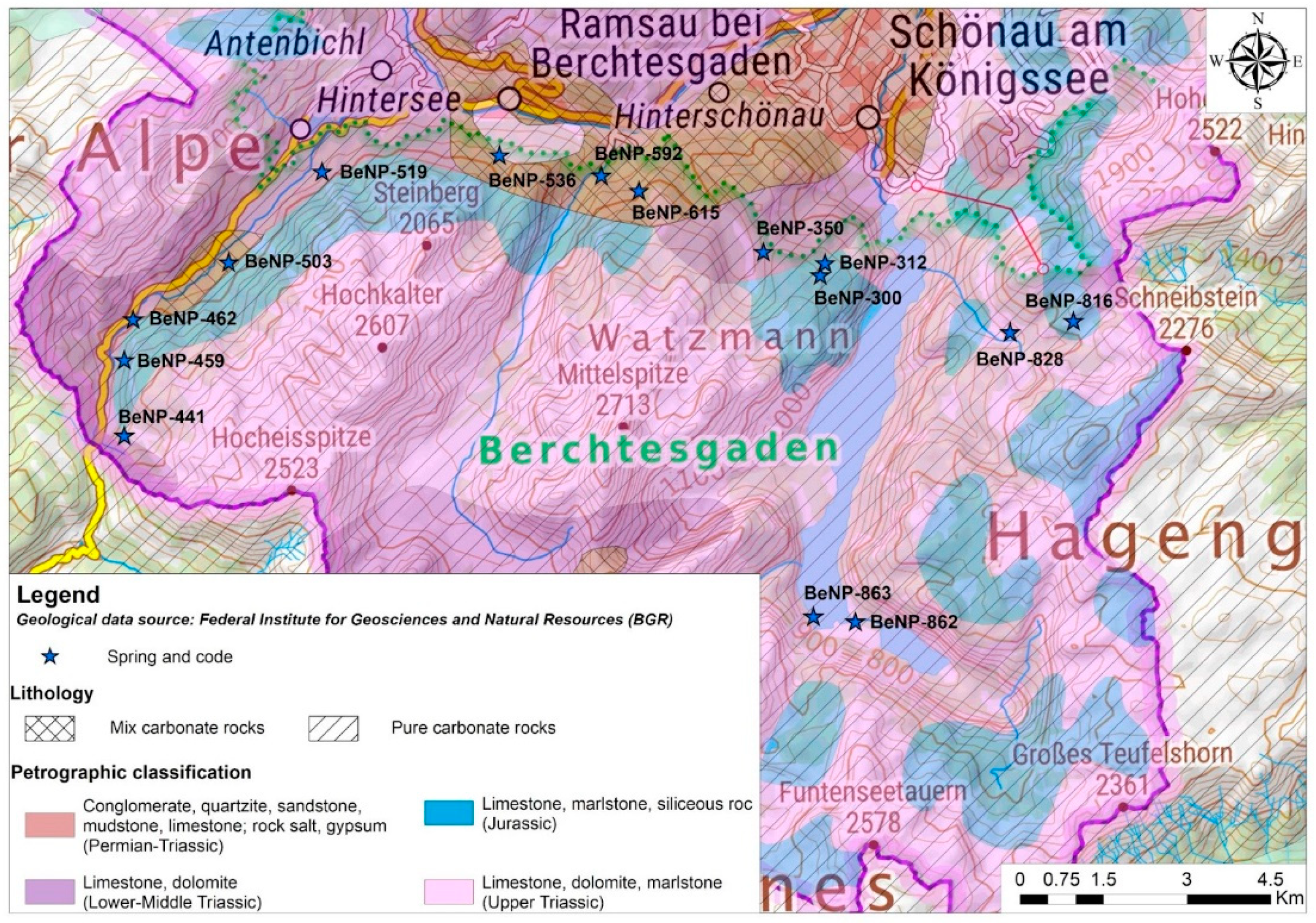
2.1. Study Area
2.2. Field Work & Sampling
2.3. Geology and Hydrogeology
2.4. Hydrochemistry
2.5. Diatom Sampling, Identification, and Quantification
2.6. Bryophytes
2.7. Data Processing and Statistical Analyses
3. Results
3.1. Morphological, Physical, and Chemical Characterization of the Springs Studied
3.2. Bryophytes (Sampled to Study Epiphytic Diatoms)
3.3. Diatom Species Found in the Different Spring Types and on the Different Substrata
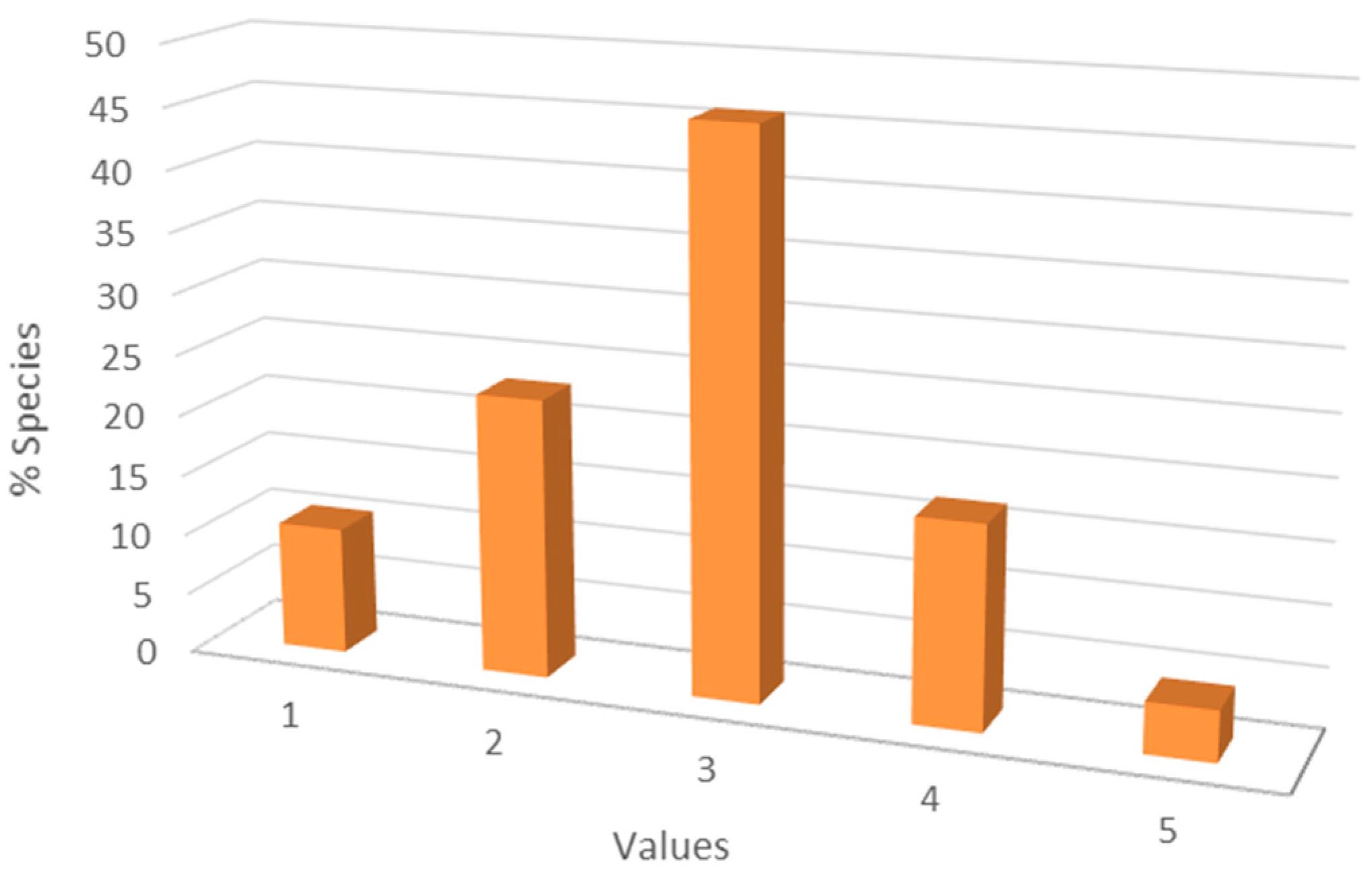
3.4. Red-List Species
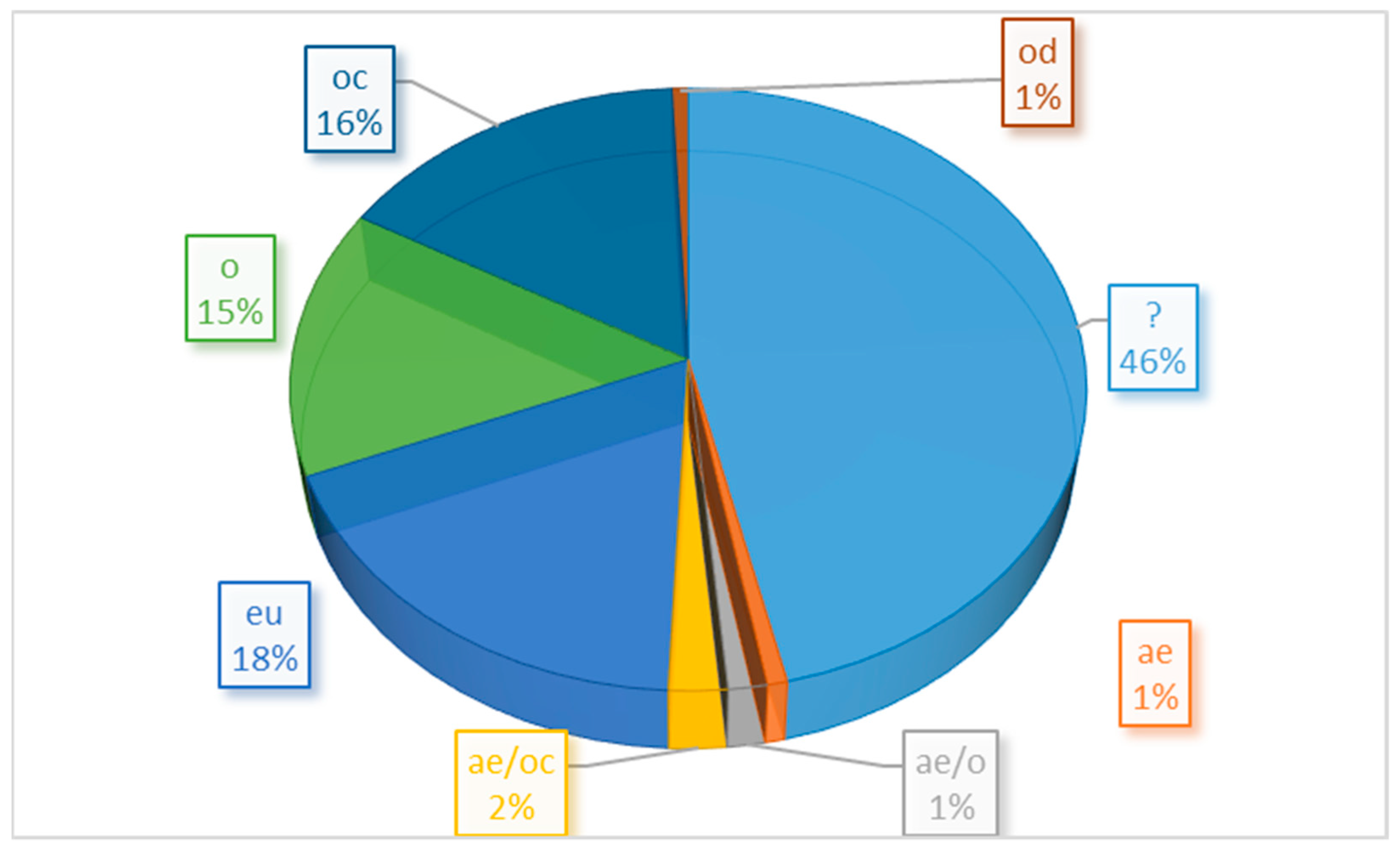
3.5. Ecological Preferences of the Species Found
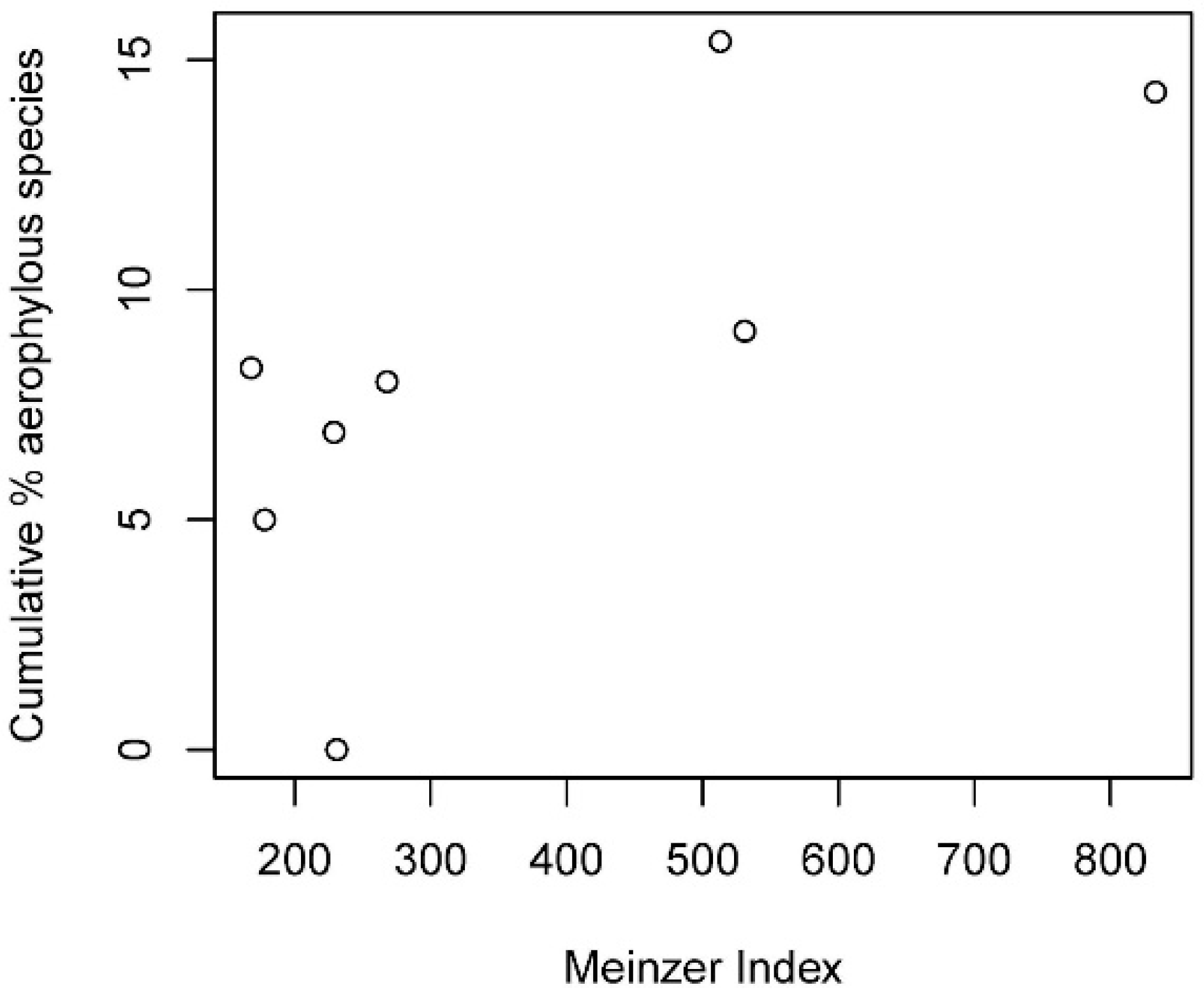
3.6. Relations with Environmental Factors
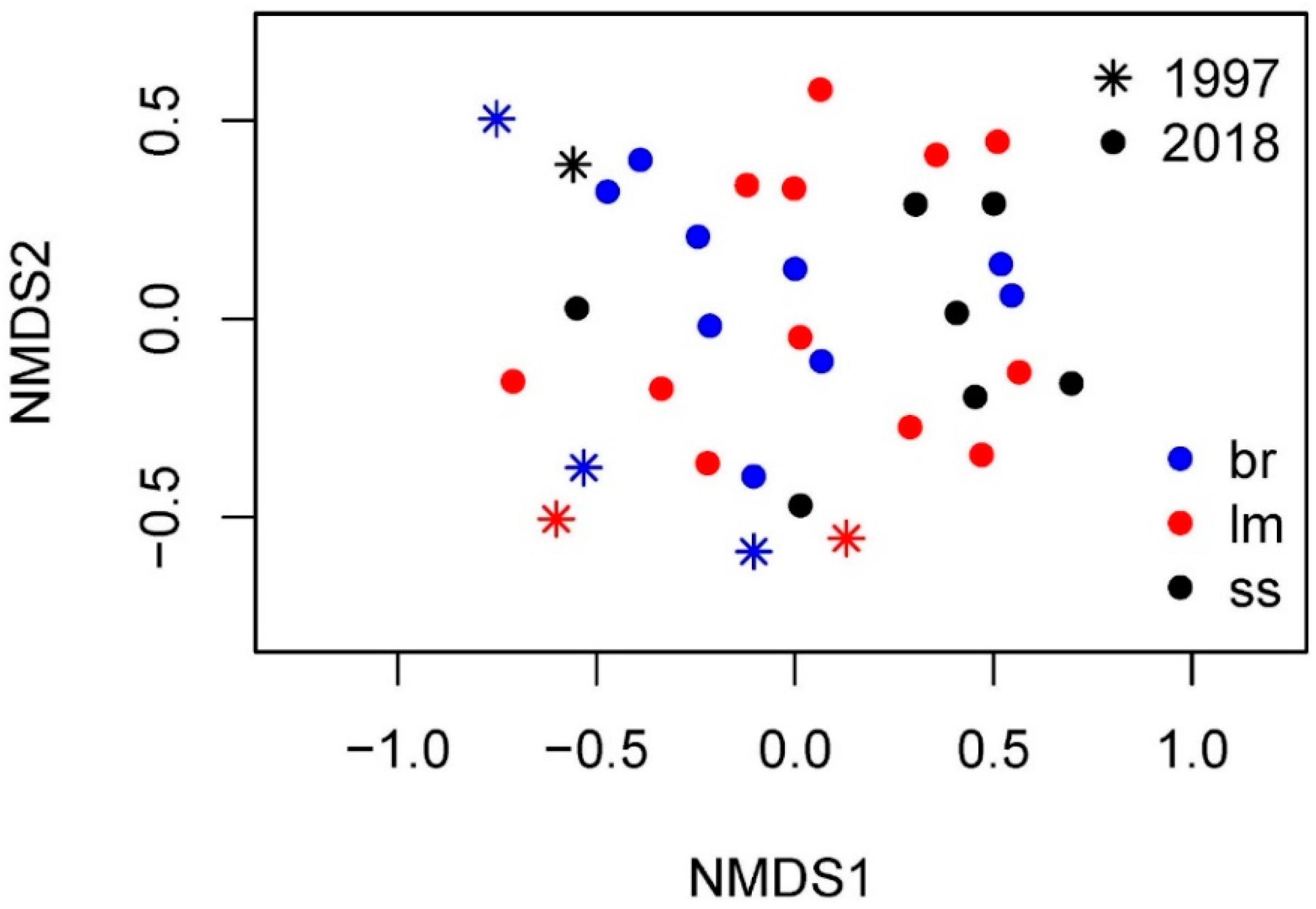
3.7. Comparison with the Data of 1997–1998
4. Discussion
- -
- a relevant occurrence of eutraphentic diatom species and reduction in taxa number and diversity in some springs (compare [20]), resulting from increased nitrate concentrations, likely due to diffuse airborne pollution or local impacts such as forest management, game, and cattle;
- -
- -
- a higher species richness on bryophytes (compare [10]), suggesting this substratum to be best suitable for studies focussing on biodiversity inventories (whilst stones should be preferred to investigate relationships between diatom assemblages and hydrochemistry).
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mann, D.G.; Vanormelingen, P. An Inordinate Fondness? The Number, Distributions, and Origins of Diatom Species. J. Eukaryot. Microbiol. 2013, 60, 414–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, D.G. The species concept in diatoms. Phycological. Reviews 18. Phycologia 1999, 38, 437–495. [Google Scholar] [CrossRef] [Green Version]
- Smol, J.P.; Stoermer, E.F. The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010; pp. 1–667. [Google Scholar]
- Lange-Bertalot, H.; Metzeltin, D. Indicators of Oligotrophy. 2. In Iconogr. Diatomol; Lange-Bertalot, H., Ed.; Koeltz Scientific Books: Königstein, Germany, 1996; pp. 1–390. [Google Scholar]
- Cantonati, M.; Füreder, L.; Gerecke, R.; Jüttner, I.; Cox, E.J. Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 2012, 31, 463–480. [Google Scholar] [CrossRef]
- Hofmann, G.; Lange-Bertalot, H.; Werum, M.; Klee, R.; unter Mitarbeit von König, C.; Kusber, W.-H.; Metzeltin, D.; Reichardt, E. Rote Liste und Gesamtartenliste der limnischen Kieselalgen (Bacillariophyta) Deutschlands. In Rote Liste Gefährdeter Tiere, Pflanzen und Pilze Deutschlands; Metzing, D., Hofbauer, N., Ludwig, G., Matzke Hajek, G., Eds.; Band 7; Naturschutz und Biologische Vielfalt 70; Landwirtschaftsverlag: Münster, Germany, 2018; pp. 601–708. [Google Scholar]
- Moser, G.; Lange-Bertalot, H.; Metzeltin, D. Insel der Endemiten Geobotanisches Phänomen Neukaledonien (Island of endemics New Caledonia—A geobotanical phenomenon); Bibliotheca Diatomologica 38; J. Cramer: Berlin/Stuttgart, Germany, 1998; pp. 1–464. [Google Scholar]
- Kociolek, J.P.; Stoermer, E.F. Oligotrophy: The forgotten end of an ecological spectrum. Proceedings of the 20th International Diatom Symposium (Dubrovnik, Croatia). Acta Bot. Croat. 2009, 68, 465–472. [Google Scholar]
- Lowe, W.H.; Likens, G.E. Moving headwater streams to the head of the class. BioScience 2005, 55, 196–197. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Angeli, N.; Bertuzzi, E.; Spitale, D.; Lange-Bertalot, H. Diatoms in springs of the Alps: Spring types, environmental determinants, and substratum. Freshw. Sci. 2012, 31, 499–524. [Google Scholar] [CrossRef]
- Cantonati, M.; Hofmann, G.; Spitale, D.; Werum, M.; Lange-Bertalot, H. Diatom Red Lists: Important tools to assess and preserve biodiversity and habitats in the face of direct impacts and environmental change. Biodivers. Conserv. 2021, in press. [Google Scholar] [CrossRef]
- Lange-Bertalot, H. Rote Liste der Limnischen Kieselalgen (Bacillariophyceae) Deutschlands; 28. Schriftenreihe für Vegetationskunde; Bundesamt fuür Naturschutz: Bonn, Germany, 1996; pp. 633–677.
- Taxböck, L.; Linder, H.P.; Cantonati, M. To what extent are Swiss springs refugial habitats for sensitive and endangered diatom taxa? Water 2017, 9, 967. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Virtanen, L.; Angeli, N.; Wojtal, A.; Gabrieli, J.; Falasco, E.; Lavoie, I.; Morin, S.; Marchetto, A.; Fortin, C.; et al. Achnanthidium minutissimum (Bacillariophyta) valve deformities as indicators of metal enrichment in diverse widely-distributed freshwater habitats. Sci. Total Environ. 2014, 475, 201–215. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H.; Scalfi, A.; Angeli, N. Cymbella tridentina sp. nov. (Bacillariophyta), a crenophilous diatom from carbonate springs of the Alps. J. N. Am. Benthol. Soc. 2010, 29, 775–788. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H. Achnanthidium dolomiticum sp. nov. (Bacillariophyta) from oligotrophic mountain springs and lakes fed by dolomite aquifers. J. Phycol. 2006, 42, 1184–1188. [Google Scholar] [CrossRef]
- Lichtenwöhrer, K.; Leonhardt, G.; Seifert, L.; Hotzy, R.; Schubert, E.; Gerecke, R.; Cantonati, M.; Lotz, A. Erfassung von Klimawandelfolgen an Quellen in Bayern. Leitfaden für eine langfristige Beobachtung von Quellen zur Erfassung von Klimawandelfolgen in Bayern. Nationalpark Berchtesgaden und Nationalpark Bayerischer Wald. Auf Vorgabe des Bayerischen Umweltministeriums. Entwurf zur praktischen Evaluierung und wissenschaftlichen Kommentierung. Bayern, Germany. 2020, pp. 1–71. Available online: https://www.nationalpark-bayerischer-wald.bayern.de/forschung/projekte/doc/leitfaden_quellbeobachtung.pdf (accessed on 7 November 2021).
- Lotter, A.F.; Pienitz, R.; Schmidt, R. Diatoms as indicators of environmental change in subarctic and alpine regions. In The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Smol, J.P., Stoermer, E.F., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 231–248. [Google Scholar] [CrossRef]
- Bahls, L.; Pierce, J.; Apfelbeck, R.; Olsen, L. Encyonema droseraphilum sp. nov. (Bacillariophyta) and other rare diatoms from undisturbed floating-mat fens in the northern Rocky Mountains, USA. Phytotaxa 2013, 127, 32–48. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Lange-Bertalot, H. Diatom biodiversity of springs in the Berchtesgaden National Park (North-Eastern Alps, Germany), with the ecological and morphological characterization of two species new to science. Diatom Res. 2010, 25, 251–280. [Google Scholar] [CrossRef]
- Gerecke, R.; Cantonati, M.; Spitale, D.; Stur, E.; Wiedenbrug, S. The challenges of long-term ecological research in springs in the northern and southern Alps: Indicator groups, habitat diversity, and medium term change. J. Limnol. 2011, 70 (Suppl. 1), 168–187. [Google Scholar] [CrossRef]
- Meinzer, O.E. Outline of Ground-Water Hydrology; Water Supply Paper; Department of the Interior, U.S. Geological Survey. Washington Government Printing Office: Washington, DC, USA, 1923; p. 494.
- Standard Methods for the Examination of Water and Wastewater, 20th ed.; APHA, AWWA & WEF, American Public Health Association: Washington, DC, USA, 2000; pp. 1–541.
- Cantonati, M.; Kelly, M.G.; Lange-Bertalot, H. (Eds.) Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Koeltz Botanical Books: Schmitten-Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 29 December 2021).
- Kociolek, J.P.; Balasubramanian, K.; Blanco, S.; Coste, M.; Ector, L.; Liu, Y.; Kulikovskiy, M.; Lundholm, N.; Ludwig, T.; Potapova, M.; et al. DiatomBase. 2021. Available online: http://www.diatombase.org (accessed on 29 December 2021).
- Spaulding, S.A.; Bishop, I.W.; Edlund, M.B.; Lee, S.; Furey, P.; Jovanovska, E.; Potapova, M. Diatoms of North America. 2020. Available online: https://diatoms.org (accessed on 30 December 2021).
- Jüttner, I.; Carter, C.; Chudaev, D.; Cox, E.J.; Ector, L.; Jones, V.; Kelly, M.G.; Kennedy, B.; Mann, D.G.; Turner, J.A.; et al. Freshwater Diatom Flora of Britain and Ireland. Amgueddfa Cymru–National Museum Wales. 2021. Available online: https://naturalhistory.museumwales.ac.uk/diatoms (accessed on 30 December 2021).
- Hodgetts, N.G.; Hodgetts, N.G.; Soderstrom, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schrock, C.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Caspari, S.; Dürhammer, O.; Sauer, M.; Schmidt, C. Rote Liste und Gesamtartenliste der Moose (Anthocerotophyta, Marchantiophyta und Bryophyta) Deutschlands. In Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands; Band 7: Pflanzen; Metzing, D., Hofbauer, N., Ludwig, G., Matzke-Hajek, G., Eds.; Münster (Landwirtschaftsverlag): Münster, Germany, 2018; Volume 70, pp. 361–489. [Google Scholar]
- Ludwig, G.; Düll, R.; Philippi, G.; Ahrens, M.; Caspari, S.; Koperski, M.; Lütt, S.; Schulz, F.; Schwab, G. Rote Liste der Moose (Anthocerophyta et Bryophyta) Deutschlands. In Rote Listen gefährdeter Pflanzen Deutschlands; Ludwig, G., Schnittler, M., Eds.; 28. Schriftenreihe für Vegetationskunde; Bundesamt fuür Naturschutz: Bonn, Germany, 1996; pp. 189–306. [Google Scholar]
- Hill, M.O.; Preston, C.D.; Bosanquet, S.D.S.; Roy, D.B. BRYOATT: Attributes of British and Irish Mosses, Liverworts and Hornworts; Software (updated 2017); Centre for Ecology and Hydrology: Lancaster, UK, 2007. [Google Scholar]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Neth. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar]
- Rott, E.; Pipp, E.; Pfister, P.; Van Dam, H.; Ortler, K.; Binder, N.; Pall, K. Indikationslisten für Aufwuchsalgen in österreichischen Fliessgewässern. Teil 2: Trophieindikation (Sowie Geochemische Präferenzen, Taxonomische und Toxikologische Anmerkungen). Wasserwirtschaftskataster, Bundesministerium f. Land-u; Forstwirtschaft: Vienna, Austria, 1999; pp. 1–248. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 37–42. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 7 March 2003).
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan, R Package Version 2.5–6; Community Ecology Package; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulißen, D. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 1991, 18, 1–248. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 1. Teil: Naviculaceae. In Süsswasserflora von Mitteleuropa, Band 2/1; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Jena, Germany, 1986; pp. 1–876. [Google Scholar]
- Krammer, K. Cymbella. In Diatoms of Europe; Lange-Bertalot, H., Ed.; J. Cramer: Berlin/Stuttgart, Germany, 2002; Volume 3, pp. 1–584. [Google Scholar]
- Cantonati, M.; Spitale, D. The role of environmental variables in structuring epiphytic and epilithic diatom assemblages in springs and streams of the Dolomiti Bellunesi National Park (south-eastern Alps). Fundam. Appl. Limnol. Arch. Hydrobiol. 2009, 1742, 117–133. [Google Scholar] [CrossRef]
- Lai, G.G.; Ector, L.; Lugliè, A.; Sechi, N.; Wetzel, C.E. Sellaphora gologonica sp. nov. (Bacillariophyta, Sellaphoraceae), a new diatom species from a Mediterranean karst spring (Sardinia, Italy). Phytotaxa 2018, 356, 145–157. [Google Scholar] [CrossRef]
- Cantonati, M.; Scola, S.; Angeli, N.; Guella, G.; Frassanito, R. Environmental controls of epilithic diatom depth-distribution in an oligotrophic lake characterised by marked water-level fluctuations. Eur. J. Phycol. 2009, 44, 15–29. [Google Scholar] [CrossRef]
- Vidaković, D.; Cantonati, M.; Mogna, M.; Jakovljević, O.; Šovran, S.; Lazović, V.; Stojanović, K.; Đorđević, J.; Krizmanić, J. Additional information on the distribution and ecology of the recently described diatom species Geissleria gereckei. Oceanol. Hydrobiol. Stud. 2017, 46, 18–23. [Google Scholar] [CrossRef]
- Cantonati, M.; Kelly, M.G.; Demartini, D.; Angeli, N.; Dörflinger, G.; Papatheodoulou, A.; Armanini, D. Overwhelming role of hydrology-related variables and river types in driving diatom species distribution and community assemblage in streams in Cyprus. Ecol. Indic. 2020, 117, 106690. [Google Scholar] [CrossRef]
- Rondón, J.C.D.; Aragón, Y.A.A. Factors driving diversity and succession of diatom assemblages in a Neotropical rainforest stream. Ann. Limnol. Int. J. Lim. 2018, 54, 30. [Google Scholar] [CrossRef]
- Taxböck, L.; Karger, D.N.; Kessler, M.; Spitale, D.; Cantonati, M. Diatom species richness in Swiss Springs Increases with Habitat Complexity and Elevation. Water 2020, 12, 449. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Segadelli, S.; Spitale, S.; Gabrieli, J.; Gerecke, R.; Angeli, N.; De Nardo, M.T.; Ogata, K.; Wehr, J.D. Geological and hydrochemical prerequisites of unexpectedly high biodiversity in spring ecosystems at the landscape level. Sci. Total Environ. 2020, 740, 140157. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SPRING CODE | BeNP-300 | BeNP-312 | BeNP-350 | BeNP-441 | BeNP-459 | BeNP-462 | BeNP-503 | BeNP-519 | BeNP-536 | BeNP-592 | BeNP-615 | BeNP-816 | BeNP-828 | BeNP-862 | BeNP-863 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Coordinates | 47°57′73.92″ N | 47°57′93.04″ N | 47°58′16.47″ N | 47°55′71.19″ N | 47°56′93.39″ N | 47°57′58.25″ N | 47°58′42.57″ N | 47°59′81.83″ N | 47°59′93.69″ N | 47°59′52.7″ N | 47°59′24.89″ N | 47°56′78.85″ N | 47°56′66.04″ N | 47°52′11.97″ N | 47°52′23.71″ N |
| 12°97′10.68″ E | 12°97′23.28″ E | 12°95′78.32″ E | 12°80′34.1″ E | 12°80′42.86″ E | 12°80′68.39″ E | 12°83′02.67″ E | 12°85′36.37″ E | 12°89′60.88″ E | 12°92′00.8″ E | 12°92′88.67″ E | 13°03′07.77″ E | 13°01′55.53″ E | 12°97′51.52″ E | 12°96′52.67″ E | |
| Altitude, m.a.s.l. | 1250 | 1150 | 1170 | 1270 | 1100 | 960 | 860 | 800 | 905 | 730 | 880 | 1575 | 1200 | 604 | 604 |
| Shading, sc. 1–5 | 2 | 1–2 | 3–4 | 3–4 | 2 | 4 | 4–5 | 3 | 5 | 4–5 | 4–5 | 1 | 3–4 | 2 | 1 |
| Spring type | Rheo-helo | Helo | Rheo | Rheo-helo | Rheo | Rheo-helo | Rheo | Rheo-helo | Rheo-helo | Rheo | Rheo-helo | Rheo-helo | Rheo-helo | Rheo-limno | Rheo-limno |
| Lithology | PCR (L, M) | PCR (L, M) | PCR (L, D) | PCR (L, D) | PCR (L, M) | PCR (L, D) | MCR (CC, Q, CM) | PCR (L, D) | MCR (CC, Q, CM) | MCR (CC, Q, CM) | MCR (CC, Q, CM) | PCR(L, M) | PCR (L, D) | PCR (L, D) | PCR (L, D) |
| Disch., L s−1 | 0.13 (0.01/0.70) | 0.03 (0.00/0.25) | 12.89 (2.00/25.0) | - | 23.75 (10.00/50.00) | - | 200 (70.00/500.00) | 0.19 (0.00/0.75) | 0.32 (0.01/0.75) | 3.29 (7.50/70.00) | 1.83 (0.10/5.00) | 0.38 (0.05/2.00) | - | - | 3.92 (1.00/10.00) |
| MVID | 531 | 833 | 178 | - | 168 | - | 215 | 395 | 231 | 1900 | 268 | 513 | - | - | 229 |
| Water T, °C | 6.85 (5.30/9.40) | 8.02 (5.70/10.30) | 5.02 (4.80/5.20) | - | 4.23 (4.00/4.50) | - | 4.68 (4.50/4.90) | 9.4 (6.70/12.70) | 8.24 (6.90/9.77) | 5.41 (5.11/5.92) | 6.98 (6.01/7.72) | 6.21 (5.50/6.80) | - | - | 6.47 (5.88/7.86) |
| MVIT | 60.00 | 57.00 | 8.00 | - | 12.00 | - | 8.50 | 64.00 | 35.00 | 15.00 | 24.50 | 21.00 | - | - | 31.00 |
| Cond., µS cm−1 | 341 (295/382) | 333 (324/346) | 273 (271/276) | 347 | 164 (143/183) | - | 155 (129/177) | 309 (276/331) | 307 (285/326) | 192 (168/217) | 336 (320/351) | 377 (357/404) | 283 | 260 | 152 (119/168) |
| MVIC | 26 | 7 | 2 | - | 24 | - | 31 | 18 | 13 | 26 | 9 | 13 | - | - | 32 |
| pH | 7.72 (7.37/8.14) | 7.91 (7.74/8.31) | 7.85 (7.74/8.02) | 7.69 | 8.14 (7.97/8.35) | - | 8.19 (8.02/8.37) | 8.18 (7.82/8.35) | 7.81 (7.56/8.07) | 8.2 (7.95/8.54) | 7.92 (7.71/8.03) | 7.65 (7.39/7.89) | 7.66 | 7.73 | 8.39 (8.07/8.94) |
| % O2 sat. | 87.18 (76.90/94.90) | 85.54 (73.30/96.80) | 98.52 (93.00/101.90) | 92 | 100.13 (96.80/104.90) | - | 100.48 (97.00/103.20) | 97.01 (92.10/101.00) | 96.95 (92.80/99.50) | 98.33 (93.20/103.60) | 98.2 (93.70/101.40) | 95.14 (92.80/101.30) | 93 | 63 | 106.72 (99.70/121.00) |
| Mg2+, mg L−1 | 2.46 (1.98/2.89) | 1.93 (1.62/2.25) | 4.34 (3.76/5.03) | 8.13 | 4.96 (3.53/7.68) | - | 487 (3.86/6.15) | 3.53 (2.43/5.06) | 6.28 (4.98/7.58) | 3.92 (2.13/6.28) | 18.37 (15.57/21.35) | 8.7 (7.87/9.30) | 3.18 | 3.14 | 1.15 (0.55/1.66) |
| Ca2+, mg L−1 | 72.15 (60.01/83.49) | 69.1 (67.34/71.35) | 53.23 (50.84/55.08) | 61.57 | 28.46 (22.94/34.03) | - | 26.94 (21.60/32.28) | 62.18 (55.23/70.31) | 57.23 (51.41/61.09) | 35.62 (29.95/47.02) | 47.1 (43.54/54.10) | 73.3 (64.11/88.29) | 63.47 | 56.87 | 30.51 (24.47/34.69) |
| Na+, mg L−1 | 0.39 (0.14/0.63) | 0.66 (0.22/1.19) | 0.51 (0.45/0.54) | 0.231 | 0.1 (0.04/0.14) | - | 0.09 (0.08/0.12) | 0.65 (0.44/1.06) | 0.24 (0.17/0.31) | 0.14 (0.06/0.31) | 0.23 (0.10/0.32) | 0.43 (0.19/0.52) | 0.53 | 0.59 | 0.11 (0.04/0.20) |
| K+, mg L−1 | 0.15 (0.05/0.34) | 0.19 (0.00/0.52) | 0.19 (0.16/0.23) | 0.08 | 0.06 (0.00/0.11) | - | 0.07 (0.00/0.15) | 0.4 (0.22/0.66) | 0.16 (0.09/0.29) | 0.11 (0.04/0.29) | 0.17 (0.08/0.26) | 0.38 (0.20/0.48) | 0.46 | 0.12 | 0.06 (0.00/0.13) |
| Cl−, mg L−1 | 0.33 (0.15/0.58) | 0.42 (0.35/0.57) | 0.42 (0.23/0.71) | 0.33 | 0.2 (0.09/0.42) | - | 0.19 (0.07/0.39) | 0.39 (0.22/0.65) | 0.4 (0.19/0.90) | 0.25 (0.09/0.85) | 0.42 (0.33/0.78) | 0.24 (0.08/0.39) | 0.26 | 0.26 | 1.13 (0.10/0.18) |
| NO3−, mg L−1 | 3.65 (1.33/10.14) | 1.35 (0.68/2.49) | 3.52 (2.59/6.38) | 2.29 | 2.22 (1.42/4.68) | - | 2.1 (1.60/3.84) | 7.14 (4.70/10.89) | 5.38 (3.49/9.38) | 3.37 (1.43/9.18) | 5.13 (3.89/11.32) | 1.29 (0.02/2.49) | 3.1 | 2.60 | 1.6 (0.67/3.34) |
| SO42−, mg L−1 | 3.79 (2.56/7.16) | 3.96 (3.04/6.08) | 5.14 (3.78/9.09) | 0.91 | 1.24 (0.67/2.62) | - | 1.3 (0.89/2.42) | 3.87 (2.67/7.59) | 2.66 (1.31/5.86) | 1.53 (0.61/3.29) | 3.11 (2.29/4.36) | 1.05 (0.01/1.73) | 2.19 | 1.26 | 0.71 (0.34/1.11) |
| CODE | BeNP-312 | BeNP-350 | BeNP-459 | |||
|---|---|---|---|---|---|---|
| Year | 2018 | 1997 | 2018 | 1997 | 2018 | 1997 |
| Disch., L s−1 | 0.03 (0.00/0.25) | 0.01 | 13 (2/25) | 10 (2/20) | 24 (10/50) | 25 (10/40) |
| Water T, °C | 8.0 (5.7/10.3) | 7.7 (5.7/10.3) | 5.0 (4.8/5.2) | 5 (4.8/5.1) | 4.2 (4.0/4.5) | 4.2 (4.1/4.5) |
| Cond., µS cm−1 | 333 (324/346) | 332 (324–346) | 273 (271/276) | 273 (271–276) | 164 (143/183) | 170 (149–183) |
| pH | 7.91 (7.74/8.31) | 7.8 (7.7–7.9) | 7.85 (7.74/8.02) | 7.8 (7.7–7.9) | 8.14 (7.97/8.35) | 8.14 (7.97–8.35) |
| Mg2+, mg L−1 | 1.9 (1.6/2.2) | 1.9 (1.6–2.1) | 4.3 (3.8/5.03) | 4.4 (3.8–5.0) | 5.0 (3.53/7.68) | 4.9 (4.6–5.9) |
| Ca2+, mg L−1 | 69.1 (67.3/71.3) | 69.0 (67.3–71.4) | 53.2 (50.8/55.1) | 53.7 (52.8–55.1) | 28.5 (22.9/34.0) | 29 (25.5–34) |
| Na+, mg L−1 | 0.7 (0.2/1.2) | 0.6 (0.6–0.7) | 0.5 (0.4/0.5) | 0.5 | 0.1 (0.04/0.14) | 0.1 |
| K+, mg L−1 | 0.19 (0.00/0.52) | 0.3 (0.1–0.5) | 0.19 (0.16/0.23) | 0.2 | 0.06 (0.00/0.11) | 0.1 |
| Cl−, mg L−1 | 0.4 (0.3/0.6) | 0.4 (0.4–0.6) | 0.4 (0.2/0.7) | 0.4 (0.2–0.7) | 0.2 (0.1/0.4) | 0.2 (0.1–0.4) |
| NO3−, mg L−1 | 1.35 (0.68/2.49) | 1.0 (0.8–1.9) | 3.52 (2.59/6.38) | 2.8 (2.7–6.4) | 2.22 (1.42/4.68) | 2.0 (1.4–4.7) |
| SO42−, mg L−1 | 4.0 (3.0/6.1) | 3.7 (3.6–4.0) | 5.1 (3.8/9.1) | 4.3 (3.9–9.1) | 1.2 (0.7/2.6) | 1.2 (1.0–2.6) |
| Ellenberg Values | Red List | Be NP-300 | Be NP-312 | Be NP-350 | Be NP-441 | Be NP-459 | Be NP-462 | Be NP-519 | Be NP-536 | Be NP-592 | Be NP-615 | Be NP-816 | Be NP-828 | Be NP-862 | Be NP-863 | tot occ | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | L | M | R | N | HM | (′18) | (′96) | |||||||||||||||
| Brachythecium rivulare Schimp. | 6 | 8 | 6 | 5 | 0 | * | * | 1 | 1 | |||||||||||||
| Bryum pseudotriquetrum (Hedw.) G.Gaertn. et al. | 8 | 9 | 6 | 3 | 0 | * | ** | 1 | 1 | 2 | ||||||||||||
| Calliergonella cuspidata (Hedw.) Loeske | 7 | 7 | 7 | 4 | 1 | * | ** | 1 | 1 | |||||||||||||
| Cratoneuron filicinum (Hedw.) Spruce | 6 | 8 | 7 | 5 | 2 | * | * | 1 | 1 | 2 | ||||||||||||
| Palustriella commutata (Hedw.) Ochyra | 6 | 9 | 8 | 2 | 0 | V | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | |||||
| Palustriella decipiens (De Not.) Ochyra | 7 | 9 | 6 | 2 | 0 | 3 | 3 | 1 | 1 | |||||||||||||
| Palustriella falcata (Brid.) Hedenäs | 8 | 9 | 6 | 2 | 0 | G | D | 1 | 1 | 2 | ||||||||||||
| Plagiomnium medium (Bruch & Schimp.) T.J.Kop. | 6 | 7 | 5 | 3 | 0 | * | 3 | 1 | 1 | |||||||||||||
| Rhizomnium punctatum (Hedw.) T.J. Kop. | 5 | 8 | 5 | 4 | 1 | * | * | 1 | 1 | 2 | ||||||||||||
| Species List | RL (′18) | RL (′96) | br | lm | ss | Ec |
|---|---|---|---|---|---|---|
| Achnanthidium affine (Grunow) Czarn. | * | * | + | - | - | ? |
| A. atomoides Monnier, Lange-Bert. et Ector | * | - | - | + | - | ? |
| A. dolomiticum Cantonati et Lange-Bert. | 2 | - | + | + | + | ? |
| A. jackii Rabenh. | D | D | + | + | + | ? |
| A. lineare W.Smith | G | ♦ | - | + | + | eu |
| A. minutissimum (Kütz.) Czarn. | * | ** | +++ | +++ | +++ | ? |
| A. pfisteri Lange-Bert. | D | - | + | ++ | + | ? |
| A. pyrenaicum (Hust.) Kobayasi | * | ** | + | ++ | + | ? |
| A. rostropyrenaicum Jüttner et Cox | (G) | + | + | - | - | |
| A. sublineare Van de Vijver, Jarlman et Ector | (R) | - | ++ | - | - | |
| Adlafia bryophila (J.B.Petersen) Lange-Bert. | * | V | + | + | + | ? |
| A. minuscula (Grunow) Lange-Bert. | * | * | + | + | ++ | ? |
| Amphipleura pellucida (Kütz.) Kütz. | * | * | + | + | + | ? |
| Amphora copulata (Kütz.) Schoeman et Archibald | * | ** | - | - | + | ? |
| A. eximia J.R. Carter | R | R | + | + | + | o |
| A. inariensis Krammer | * | 3 | - | + | - | o |
| A. indistincta Levkov | * | - | + | + | + | ? |
| A. micra Levkov | Do | + | + | - | - | |
| A. pediculus (Kütz.) Grunow | * | ** | + | + | + | ? |
| A. pellucida W.Greg. | + | + | + | - | ||
| Brachysira neoexilis Lange-Bert. | * | * | + | - | + | o |
| Caloneis constans E.Reichardt | R | - | + | - | - | oc |
| C. fontinalis (Grunow) Lange-Bert. et E.Reichardt | * | - | ++ | + | + | ? |
| C. lancettula (Schulz-Danzing) Lange-Bert. et Witkowski | * | ♦ | + | + | - | eu |
| C. langebertalotioides E.Reichardt | G | - | + | - | - | oc |
| C. schumanniana (Grunow) Cleve | - | + | - | o | ||
| C. tenuis (W.Greg.) Krammer | 3 | G | + | - | - | o |
| Cavinula jaernefeltii (Hust.) D.G. Mann et Stickle | 3 | 3 | - | - | + | ? |
| Cocconeis euglypta Ehrenb. | * | ** | + | + | + | ? |
| C. lineata Ehrenb. | * | ** | + | + | + | ? |
| C. pseudolineata (Geitler) Lange-Bert. | * | D | + | + | + | ? |
| Cymbella affinis Kütz. | 2 | ♦ | + | + | + | ? |
| C. tridentina Lange-Bert., Cantonati et A.Scalfi | 2 | - | - | + | - | oc |
| C. vulgata Krammer | 3 | - | + | - | - | ? |
| Cymbopleura austriaca (Grunow) Krammer | 2 | V | + | - | - | ae/oc |
| Cymbella diminuta (Grunow) Krammer | (3) | + | - | - | o | |
| C. korana Krammer | Do | - | - | - | + | oc |
| C. naviculiformis (Auersw. ex Heib.) Krammer | * | * | - | - | + | ? |
| C. subaequalis (Grunow) Krammer | 3 | ♦ | + | - | - | o |
| Cyclotella sp. (Kütz.) Bréb. | + | - | - | - | ||
| Delicata delicatula (Kütz.) Krammer | 3 | G | - | - | + | oc |
| D. minuta Krammer | G | - | + | + | - | ae/oc |
| Denticula tenuis Kütz. | * | * | ++ | +++ | ++ | o |
| Diploneis krammeri Lange-Bert. et E.Reichardt | V | ♦ | + | + | + | oc |
| D. oculata (Bréb.) Cleve | * | * | + | + | + | ? |
| D. petersenii Hust. | 3 | 3 | - | + | - | o |
| D. separanda Lange-Bert. | D | ♦ | + | + | + | oc |
| D. tirolensis Lange-Bert. | D | - | + | - | - | oc |
| Diploneis sp. (Ehrenb.) Cleve | + | - | - | - | ||
| Ellerbeckia arenaria (Moore ex Ralfs) Crawford | * | ** | - | - | + | ? |
| Encyonema alpinum (Grunow) D.G.Mann D.G.Mann | G | G | + | + | - | ae/oc |
| E. auerswaldii Rabenh. | D | - | - | + | - | ? |
| E. lange-bertalotii Krammer | * | - | + | + | ++ | ? |
| E. minutum (Hilse) D.G. Mann | * | * | + | ++ | + | ? |
| E. silesiacum (Bleisch) D.G. Mann | * | ♦ | - | - | + | ? |
| E. sublangebertalotii Lange-Bert. & Cantonati | G | - | + | + | + | oc |
| E. ventricosum (Agardh) Grunow | * | ♦ | + | + | - | ? |
| Encyonopsis cesatii (Rabenh.) Krammer | V | ♦ | + | + | + | o |
| E. falaisensis (Grunow) Krammer | G | G | + | - | - | o |
| E. fonticola (Hust.) Krammer | (3) | + | + | - | - | |
| E. hibernica Kennedy, Buckley et Allott | + | - | - | - | ||
| E. krammeri E.Reichardt | G | - | + | + | + | oc |
| E. minuta Krammer et E.Reichardt | D | - | + | + | + | ? |
| E. subminuta Krammer et E.Reichardt | G | - | + | - | + | o |
| Encyonopsis sp. Krammer | + | - | - | - | ||
| Eucocconeis laevis (Østrup) Lange-Bert. | V | * | + | + | ++ | o |
| Eunotia arcubus Nörpel et Lange-Bert. | 2 | 2 | + | + | - | oc |
| E. bilunaris (Ehrenb.) Schaarschm. | * | ** | + | - | - | ? |
| E. glacialispinosa Cantonati et Lange-Bert. | G | - | + | - | - | o |
| Fallacia lenzii (Hust.) Lange-Bert. | * | 3 | + | - | - | ? |
| F. subhamulata (Grunow) D.G.Mann D.G.Mann | * | * | - | - | + | eu |
| Fragilaria gracilis Østrup | * | * | - | - | + | ? |
| F. vaucheriae (Kütz.) J.B.Petersen | * | ** | - | + | + | eu |
| Geissleria gereckei Cantonati & Lange-Bert. | (2) | - | + | - | - | |
| Gomphonema angustum C. Agardh | G | V | ++ | ++ | + | oc |
| G. elegantissimum E.Reichardt et Lange-Bert. | * | - | + | ++ | - | oc |
| G. hebridense W.Greg. | V | V | - | + | - | ? |
| G. innocens E.Reichardt | * | - | - | + | + | ? |
| G. longiceps Ehrenb. | D | ♦ | + | - | - | ? |
| G. minutum (C.Agardh) C.Agardh | * | ** | - | + | - | eu |
| G. micropus Kütz. | * | ♦ | ++ | - | + | ? |
| G. pala E.Reichardt | G | - | + | - | - | o |
| G. parvulum (Kütz.) Kütz. | * | ** | + | - | - | ? |
| G. sarcophagus W.Greg. | (R) | + | - | - | ? | |
| G. subclavatum (Grunow) M. Schmidt | * | ♦ | + | - | - | ? |
| G. utae Lange-Bertalot et E.Reichardt | * | D | + | - | - | ? |
| Gyrosigma acuminatum (Kütz.) Rabenh. | * | ♦ | - | - | + | eu |
| G. attenuatum (Kütz.) Rabenh. | * | * | - | + | - | ? |
| Humidophila contenta (Grunow) Lowe, Kociolek, Johansen, Van de Vijver, Lange-Bert. et Kopalová | D | ♦ | + | + | - | ae |
| H. paracontenta (Lange-Bert. & Werum) Lowe, Kociolek, Johansen, Van de Vijver, Lange-Bert. et Kopalová | Do | - | - | + | + | ae/o |
| H. perpusilla (Grunow) R.L.Lowe, Kociolek, J.R.Johans., Van de Vijver, Lange-Bert. et Kopalová | * | ** | + | + | - | ae/o |
| Karayevia clevei (Grunow) Bukht. | * | * | + | + | + | eu |
| Lindavia radiosa (Grunow) De Toni et Forti | - | + | + | ? | ||
| Luticola frequentissima Levkov, Metzeltin et A.Pavlov | D | 0 | + | - | - | ? |
| Meridion circulare (Gréville) C. Agardh | * | ** | + | + | + | ? |
| Navicula antonii Lange-Bert. | * | ** | + | + | ++ | eu |
| N. sp. aff. metareichardtiana Lange-Bert. et Kusber | + | - | + | - | ||
| N. cataracta-rheni Lange-Bert. | G | R | + | + | + | oc |
| N. cryptocephela Kütz. | * | ** | + | + | +++ | eu |
| N. cryptotenella Lange-Bert. | * | - | +++ | ++ | +++ | ? |
| N. dealpina Lange-Bert. | 2 | V | + | + | - | oc |
| N. lanceolata (C.Agardh) Ehrenb. | * | ** | - | + | + | eu |
| N. leistikowii Lange-Bert. | G | G | + | + | + | oc |
| N. radiosa Kütz. | * | ** | + | - | - | ? |
| N. recens (Lange-Bert.) Lange-Bert. | * | * | + | - | - | eu |
| N. subalpina E.Reichardt | 3 | V | - | + | - | oc |
| N. upsaliensis (Grunow) Perag. | * | R | - | - | + | eu |
| N. wildii Lange-Bert. | 2 | 3 | + | - | - | oc |
| N. wygaschii Lange-Bert. | G | - | - | - | + | oc |
| Naviculadicta sp. Lange-Bert. | + | - | - | - | ||
| Neidiomorpha binodiformis (Krammer) Cantonati, Lange-Bert. et N.Angeli | G | G | - | + | - | oc |
| Neidium cuneatiforme Levkov | R+ | - | - | + | - | |
| Nitzschia acidoclinata Lange-Bert. | V | * | + | - | + | ? |
| N. alpina Hust. | 3 | G | - | - | + | o |
| N. dissipata (Kütz.) Grunow | * | ** | + | + | +++ | eu |
| N. fonticola Grunow | * | ** | ++ | + | ++ | eu |
| N. linearis (C. Agardh) W. Smith | * | ** | + | - | + | eu |
| N. cf. palea (Kütz.) W.Smith | * | ** | - | - | + | eu |
| N. perminuta (Grunow) H.Perag. | * | * | + | - | + | ? |
| N. puriformis Hlúbiková & Ector | D | ♦ | + | - | - | eu |
| N. sigmoidea (Nitzsch) W. Smith | * | ** | - | + | + | eu |
| N. sublinearis Hust. | * | * | + | - | + | ? |
| N. tenuis W.Smith | * | * | - | - | + | ? |
| Odontidium mesodon (Ehrenb.) Ralfs | * | * | +++ | ++ | + | ? |
| O. neomaximum Jüttner, D.M.Williams, Levkov, E.Falasco, M.Battegazzore, Cantonati, Van de Vijver, C.Angele et Ector | (3) | + | + | - | - | |
| Pinnularia viridiformis Krammer | D | G | + | - | - | o |
| Pinnularia sp. “small” 18 str/10 µm | - | + | - | - | ||
| Placoneis paraelginensis Lange-Bert. | D | - | + | - | - | ? |
| P. undulata(Østrup) Lange-Bert. | * | - | + | + | - | ? |
| Planothidium dubium (Grunow) Round et Bukht. | * | * | - | + | ++ | eu |
| P. frequentissimum (Lange-Bert.) Lange-Bert. | * | ** | + | + | + | eu |
| P. lanceolatum (Bréb. ex Kütz.) Lange-Bert. | * | ** | +++ | +++ | +++ | ? |
| P. reichardtii Lange-Bert. et Werum | D | - | + | + | +++ | ? |
| Platessa montana (Krasske) Lange-Bert. | 3 | 3 | - | + | - | o |
| Psammothidium sp. Bukht. et Round | + | - | - | - | ||
| P. bioretii (Germain) Bukht. et Round | * | V | - | + | + | ? |
| P. grischunum (Wuthrich) Bukht. et Round | V | - | + | + | ++ | ? |
| Pseudostaurosira parasitica (W.Smith) E.Morales | * | ** | - | - | + | eu |
| P. robusta (Fusey) D.M. Williams et Round | G | * | + | - | - | ? |
| Reimeria capitata (Cleve-Euler) Levkov & Ector | (R) | - | + | - | - | |
| R. fontinalis Levkov et Ector | (R) | + | + | + | - | |
| R. ovata (Hust.) Levkov et Ector | (R) | - | + | - | - | |
| R. sinuata (W.Greg.) Kociolek et Stoermer | * | ♦ | - | + | - | ? |
| Rossithidium petersenii (Hust.) Round et Bukht. | 3 | +++ | + | - | o | |
| Sellaphora bacillum (Ehrenb.) D.G. Mann | * | V | + | - | - | eu |
| S. gologonica Lai, Ector et C.E.Wetzel | (G) | + | + | + | - | |
| S. nigri (De Not.) C.E.Wetzel et Ector | - | + | + | - | ||
| S. pseudopupula (Krasske) Lange-Bert. | G | G | + | + | - | od |
| S. pupula (Kütz.) Mereschk. | D | ** | - | + | + | eu |
| S. aff. schadei (Krasske) C.E.Wetzel, Ector, Van de Vijver, Compère et D.G.Mann | 2 | 2 | + | - | - | o |
| S. seminulum (Grunow) D.G.Mann | * | ** | + | + | + | ? |
| S. stroemii (Hust.) D.G.Mann | 2 | 3 | - | + | + | oc |
| S. cf. labernardierei A.Beauger, C.E.Wetzel et Ector | (G) | - | - | + | - | |
| S. aff. circumborealis (Lange-Bert.) C.E.Wetzel, Ector, Van de Vijver, Compère et D.G.Mann | - | + | - | - | ||
| Stauroforma exiguiformis (Lange-Bert.) Flower, Jones et Round | 3 | + | - | - | ? | |
| Stauroneis separanda Lange-Bertalot et Werum | V | - | - | - | + | oc |
| S. smithii Grunow | R | ♦ | + | - | + | eu |
| Staurosira venter (Ehrenb.) Grunow | * | ** | + | + | + | ? |
| S. leptostauron (Ehrenb.) Kulikovskiy et Genkal | - | - | + | ? | ||
| S. leptostauron var. dubia (Grunow) M.B.Edlund | + | - | - | ? | ||
| Staurosirella neopinnata E.A.Morales, C.E.Wetzel, Haworth et Ector | D | + | + | + | - | |
| Surirella angusta Kütz. | * | * | - | - | + | eu |
| Df | SS | MS | F | R2 | P | |
|---|---|---|---|---|---|---|
| Year | 1 | 0.586 | 0.586 | 2.109 | 0.061 | 0.032 |
| Substratum | 2 | 0.611 | 0.306 | 1.1 | 0.064 | 0.351 |
| Residuals | 30 | 8.337 | 0.278 | 0.874 | ||
| Total | 33 | 9.534 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantonati, M.; Bilous, O.; Spitale, D.; Angeli, N.; Segadelli, S.; Bernabè, D.; Lichtenwöhrer, K.; Gerecke, R.; Saber, A.A. Diatoms from the Spring Ecosystems Selected for the Long-Term Monitoring of Climate-Change Effects in the Berchtesgaden National Park (Germany). Water 2022, 14, 381. https://0-doi-org.brum.beds.ac.uk/10.3390/w14030381
Cantonati M, Bilous O, Spitale D, Angeli N, Segadelli S, Bernabè D, Lichtenwöhrer K, Gerecke R, Saber AA. Diatoms from the Spring Ecosystems Selected for the Long-Term Monitoring of Climate-Change Effects in the Berchtesgaden National Park (Germany). Water. 2022; 14(3):381. https://0-doi-org.brum.beds.ac.uk/10.3390/w14030381
Chicago/Turabian StyleCantonati, Marco, Olena Bilous, Daniel Spitale, Nicola Angeli, Stefano Segadelli, Dimitri Bernabè, Kurt Lichtenwöhrer, Reinhard Gerecke, and Abdullah A. Saber. 2022. "Diatoms from the Spring Ecosystems Selected for the Long-Term Monitoring of Climate-Change Effects in the Berchtesgaden National Park (Germany)" Water 14, no. 3: 381. https://0-doi-org.brum.beds.ac.uk/10.3390/w14030381