
Effects of Spring Warming and Drought Events on the Autumn Growth of Larix kaempferi Seedlings



1
Department of Environmental Science and Ecological Engineering, Korea University, Seoul 02841, Korea
2
Ojeong Resilience Institute, Korea University, Seoul 02841, Korea
3
Institute of Life Science and Natural Resources, Korea University, Seoul 02841, Korea
*
Author to whom correspondence should be addressed.
Water 2022, 14(12), 1962; https://0-doi-org.brum.beds.ac.uk/10.3390/w14121962
Submission received: 18 May 2022
/
Revised: 16 June 2022
/
Accepted: 17 June 2022
/
Published: 19 June 2022
(This article belongs to the Special Issue Implications of Climate Change on the Sustainable Management of the Water–Forest Nexus)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:High temperatures and droughts following winter dormancy can negatively affect seedling growth and mortality. An open-field experiment was conducted to study the growth and mortality of Larix kaempferi seedlings in response to spring warming and drought treatments and to determine whether seedlings could regain their growth capability once the treatments were discontinued. In May 2020, 1-year-old seedlings were exposed to four treatments: control, warming-only, drought-only, and the combined warming and drought. Drought treatment reduced the seedling height and root collar diameter and increased the mortality rate. The combined warming and drought treatments had the highest mortality rates, followed by the drought, control, and warming treatments. However, after the cessation of the treatments, the combined warming and drought treatments increased seedling height, root collar diameter, and individual seedling biomass because the high mortality rate relaxed competition among seedlings. This suggests that the effects of low competition on the surviving seedlings may mitigate the negative effects of warming and drought on seedling growth. Our study demonstrates that despite the high mortality and decreased growth during the treatment period, seedlings subjected to combined high temperature and drought stress showed short-term high levels of growth compared to seedlings subjected to a single stress.
1. Introduction
Human-induced warming reached approximately 1 °C above the pre-industrial global temperature levels in 2017, increasing at a rate of 0.2 °C per decade [1]. At the same time, climate change has generally accelerated the hydrological processes, resulting in more intense and frequent drought events [2]. The Intergovernmental Panel on Climate Change (IPCC) predicted that the increase in global temperature and drought events will gradually deteriorate, especially in terrestrial regions [3]. Those changes in climate will trigger a wide range of impacts on forest ecosystems, including direct impacts on trees such as increased mortality, decrease in growth rate, and reduced regeneration success [4,5,6].
As spring is the growing season, plants are more sensitive to environmental stresses compared to other seasons. Changes in spring temperature and precipitation substantially impact the structure and functioning of ecosystems by affecting plant phenology and growth [7,8]. Warmer springs may cause the positive or negative lagged effects on plants’ productivity during the following summer and autumn [9]. Richardson et al. [10] who investigated relationships between surface-atmosphere CO2 fluxes and interannual variation in a boreal-northern forest, showed that the earlier spring might accelerate N mineralization, and thus, photosynthetic activities could be increased as a result of higher foliar N concentrations over the entire growing season. In contrast, warmer springs could trigger a decrease in the productivity of plants in the subsequent seasons through the water deficit [11]. In addition, drier springs could affect leaf life span, resulting in earlier autumn senescence [12,13]. The impact of spring drought on plants would be greater because spring droughts have increased their intensity due to climate change in recent years [14].
To understand the response of trees to environmental change, researchers have manipulated air temperature and precipitation in numerous field experiments [15,16,17]. Climate manipulation experiments can be useful tools to figure out the responses of trees to climate change since the manipulative experiments allow researchers to pinpoint the mechanisms between environmental factors and ecosystems [18]. Jo et al. [19] and Chang et al. [20] reported that the experimental +3 °C warming using an infrared heater might induce a decrease in photosynthetic activities of coniferous seedlings, potentially resulting in an increase in mortality rate. Using a mobile rainout shelter, Misson et al. [21] simulated drought in the Mediterranean forest and showed that spring drought can inhibit foliar and floral development of Quercus ilex by affecting the phenophase progression. In the experiment of Rizhsky et al. [22], Arabidopsis plants could open their stomata due to the extremely high leaf temperature under the combined warming and drought treatment.
Larix kaempferi is a major plantation species with fast-growing characteristics and accounts for 32% of the total plantation area in South Korea [14,23]. The production of L. kaempferi seedlings is steadily increasing as demand for timber has increased [14,23]. However, L. kaempferi is known for being highly vulnerable to climate change [24]. In the experiment of An et al. [25], the +3 °C warming treatment significantly limited leaf gas exchange of 2-year-old L. kaempferi seedlings. Kwon et al. [26] showed that the mortality rate of L. kaempferi increased under the increased temperature and decreased precipitation, and the seedling quality index was low in the increased precipitation treated group. In addition, Bhusal et al. [14] revealed that the maximum photosynthetic rate of L. kaempferi seedlings can be decreased by approximately 48% compared to that of the control under drought stresses. However, few studies have examined the effects of warming and drought on L. kaempferi in the spring season, and even fewer have examined the effects on L. kaempferi for an extended period after the cessation of the treatments. It is necessary to understand how excessively high temperatures and drought might influence L. kaempferi seedlings in the spring and how their response is altered after a period of warming and drought.
In this study, an open-field manipulation system was used to simulate the spring warming and drought of Seoul in the 2070s for L. kaempferi seedlings. Among all growing stages, the seedling stages are particularly susceptible to changes in environmental conditions due to the immature state of their roots and stems [27]. Regarding the long-term performance of tree species which is highly related to the seedling stage, it is important to determine the response of seedlings under the changing environments to understand the overall performance of trees [28]. The objectives of this study were to figure out the growth and mortality of L. kaempferi seedlings under the spring warming and drought treatment, and to determine whether the seedlings can recover from the heat and drought stresses during the subsequent summer and autumn.
2. Materials and Methods
2.1. Experimental Warming and Drought Treatments
The experimental site was located in an open-field nursery site at the Korea University Green Campus in Seoul, South Korea (37°35′36″ N, 127°1′31″ E). The area has a moderate climate, with annual mean precipitation of 1450.5 mm (61% of which occurs during June through August) and an annual mean temperature of 12.5 °C [29]. The soil texture was sandy loam. The acidity (pH) of the soil was 6.52, carbon content was 0.22%, and nitrogen content was 0.05%.
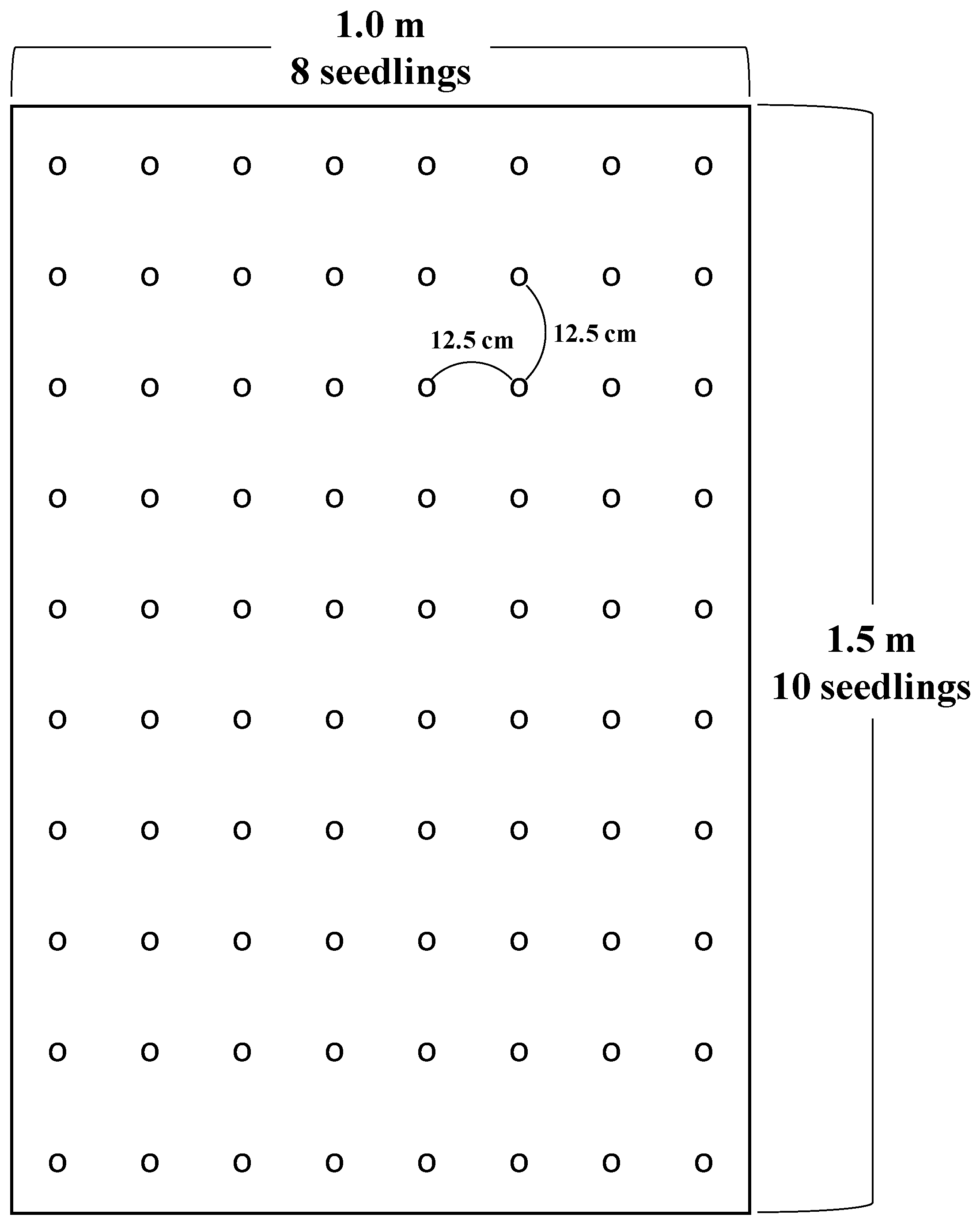
In March 2020, the experimental plots were established in March 2020. In April, seedlings had a rest period for a month to develop a root system and get accustomed to the soil. In May, 1-year-old L. kaempferi seedlings were exposed to two different temperature conditions (ambient temperature and a 4 °C increase in the second and fourth weeks of May) and two different precipitation conditions (ambient precipitation and precipitation blocking in all four weeks in May). Five 1.0 m × 1.5 m plots were established for each of the four treatments (a total of twenty plots): control, warming-only, drought-only, and the combined warming and drought. A total of 80 1-year-old L. kaempferi seedlings were planted in each plot (Figure 1). The seedlings were chosen based on the quality standard of forest seeds by the National Forest Seed and Variety Center and were genetically homogeneous.
Environmental factors were manipulated to simulate the spring environment of Seoul in the 2070s. Under the Representative Concentration Pathway (RCP) 8.5 scenario which predicted climate change until 2100 in Seoul, Korea, the average spring temperature is predicted to increase from 12.6 °C (2011–2020) to 16.7 °C (2071–2080), and the maximum consecutive dry days is projected to increase from 29.3 days (2011–2020) to 29.8 days (2071–2080) [29]. Accordingly, the warming treatment was designed to increase the air temperature by 4 °C and the drought treatment was designed to simulate 30 days of precipitation reduction [1,29].
Infrared heaters hanging 60 cm above the average seedling height were established to increase the temperature in the warming plots. As infrared heaters cast shadows over plots, two dummies with the same shape and size as the infrared heater were also established in the control and the warming-only plots. An automatic rainout shelter was installed 1.6 m above the ground to completely block rain in the drought plots. The rainout shelters were coiled onto a beam when there is no rain and automatically opened when an electronic controller detects rain. Infrared thermometers (SI-111; Campbell Scientific Inc., Logan, UT, USA) and soil temperature moisture sensors (CS655; Campbell Scientific Inc., Logan, UT, USA) were placed in each plot to measure air temperature, soil temperature, and soil moisture content. These environmental data were collected every half-hour by using a data logger (CR1000X; Campbell Scientific Inc., Logan, UT, USA).
2.2. Seedling Growth and Mortality Rate
We measured all the seedlings’ height and root collar diameter on 30 April (right before the treatment started), 1 June (right after the treatment ceased), and 30 October (when the growing season ended). Seedling height was measured using a folding ruler. Root collar diameter was measured 1 cm above the ground using a digital caliper. At the end of the growing season, five randomly chosen seedlings from each plot were harvested, and their biomasses were measured. Manual excavation was used to retrieve as many roots as possible from the seedlings. Seedlings were collected; partitioned into stems, leaves, and roots; dried at 65 °C, and weighed to the nearest 0.01 g. The mortality rate was quantified at the end of the growing season (1).
2.3. Data Analysis
The effects of artificial warming and drought and their interaction on air temperature, soil temperature, soil moisture content, seedling height, root collar diameter, biomass, and mortality were analyzed using two-way ANOVA. Tukey’s test was used to determine whether individual treatments were significantly different at a 0.05 probability level. Additionally, linear regression was used to analyze relationships between mortality rate and biomass of leaves, stems, and roots, and the total biomass. SAS version 9.4 was used for all statistical analyses (SAS Institute Inc., Cary, NC, USA).
3. Results
3.1. Experimental Warming and Drought Treatments
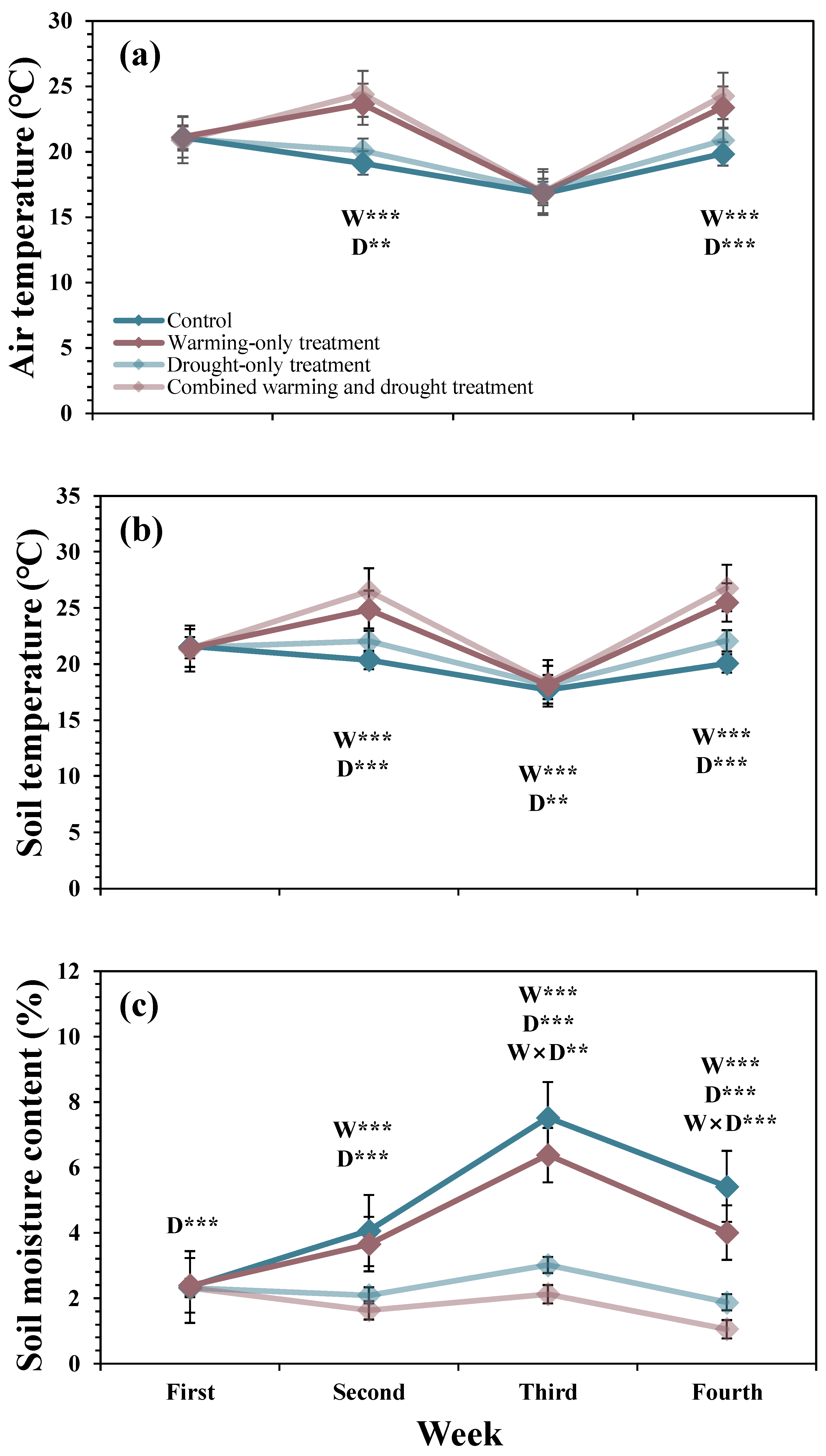
During the second and fourth weeks in May when infrared heaters were activated, both warming and drought treatments increased air temperature (Figure 2a). Air temperature in the control, warming-only, drought-only, and combined warming and drought plots were 19.1, 23.5, 20.1, and 24.3 °C in the second week and 19.8, 23.1, 20.8, and 24.0 °C in the fourth week, respectively. On average, the increase in air temperature by the warming and drought treatments were 4.0 and 1.0 °C, respectively. Additionally, the warming and drought treatments elevated soil temperature significantly during the warming treatment period (Figure 2b). Soil temperatures in the control, warming-only, drought-only, and combined warming and drought plots were 20.4, 24.9, 22.0, and 26.5 °C in the second week and 20.0, 25.5, 22.1, and 27.2 °C in the fourth week, respectively. On average, the increase in soil temperature by the warming and drought treatments were 5.0 and 1.9 °C, respectively. Although the infrared heaters were not activated in the third week, soil temperature in the warming-only plot was higher than that in the control plot by 0.5 °C.
The mean soil moisture content of the control plot in the first, second, third, and fourth weeks in May were 2.4, 4.1, 7.5, and 5.4%, respectively (Figure 2c). In the first week, the drought treatment significantly decreased soil moisture content by 0.04%. In the second week, the warming and drought treatments decreased soil moisture content by 2.0 and 0.4%, respectively. The warming and drought treatments decreased soil moisture content, but the warming treatment decreased the decreasing impact of the drought treatment on soil moisture content in the third and fourth weeks. In the third week, the drought treatment without warming decreased soil moisture content by 4.5%, but the drought treatment with warming decreased soil moisture content by only 4.3%. Similarly, in the fourth week, the drought treatment without warming decreased soil moisture content by 3.5%, but the drought treatment with warming decreased soil moisture content by only 3.0% in the third week.
3.2. Seedling Height and Root Collar Diameter Response
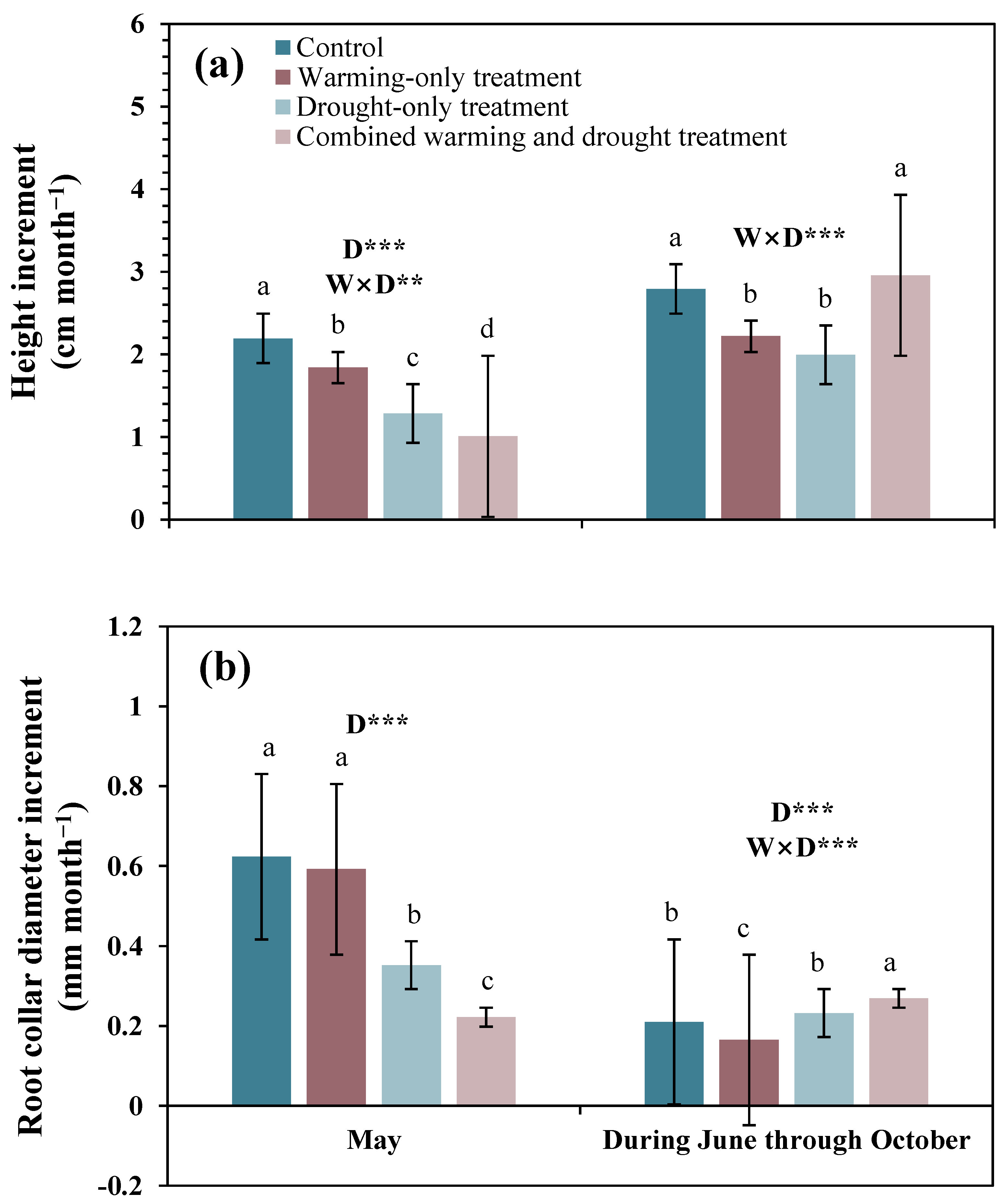
In the treatment period (i.e., in May), the height increment was largest in the control plots by 2.2 cm month−1 and lowest in the combined warming and drought plots by 3.0 cm month−1 (Figure 3a). In this period, the drought treatment significantly decreased height increment; however, there was no significant effect of warming on height increment, and there was a significant interaction effect between warming and drought. The drought treatment without warming decreased height increment by 0.91 cm month−1, but the drought treatment with warming decreased height increment only by 0.83 cm month−1. However, until the end of the growing season after the treatment period (i.e., during June through October), there was no significant difference in height increments in the control and the combined warming and drought plots. Moreover, there were no significant impacts of warming and drought treatments on height increment, yet there was a significant interaction effect between these two treatments.
In the treatment period, root collar diameter increment was largest in the control plots by 0.6 cm month−1 and lowest in the combined warming and drought plots by 0.2 cm month−1 (Figure 3b). In this period, the drought treatment significantly decreased root collar diameter increment. However, until the end of the growing season after the treatment period (i.e., during June through October), the root collar diameter increment in the combined warming and drought plots was larger than that in the control plots. Moreover, the drought treatment significantly affected root collar diameter increment, while there was no significant effect of the warming treatment. Additionally, there was a significant interaction effect between warming and drought treatment; the drought treatment without warming increased height increment by 0.02 cm month−1, but the drought treatment with warming decreased height increment only by 0.10 cm month−1.
3.3. Biomass and Mortality Rate
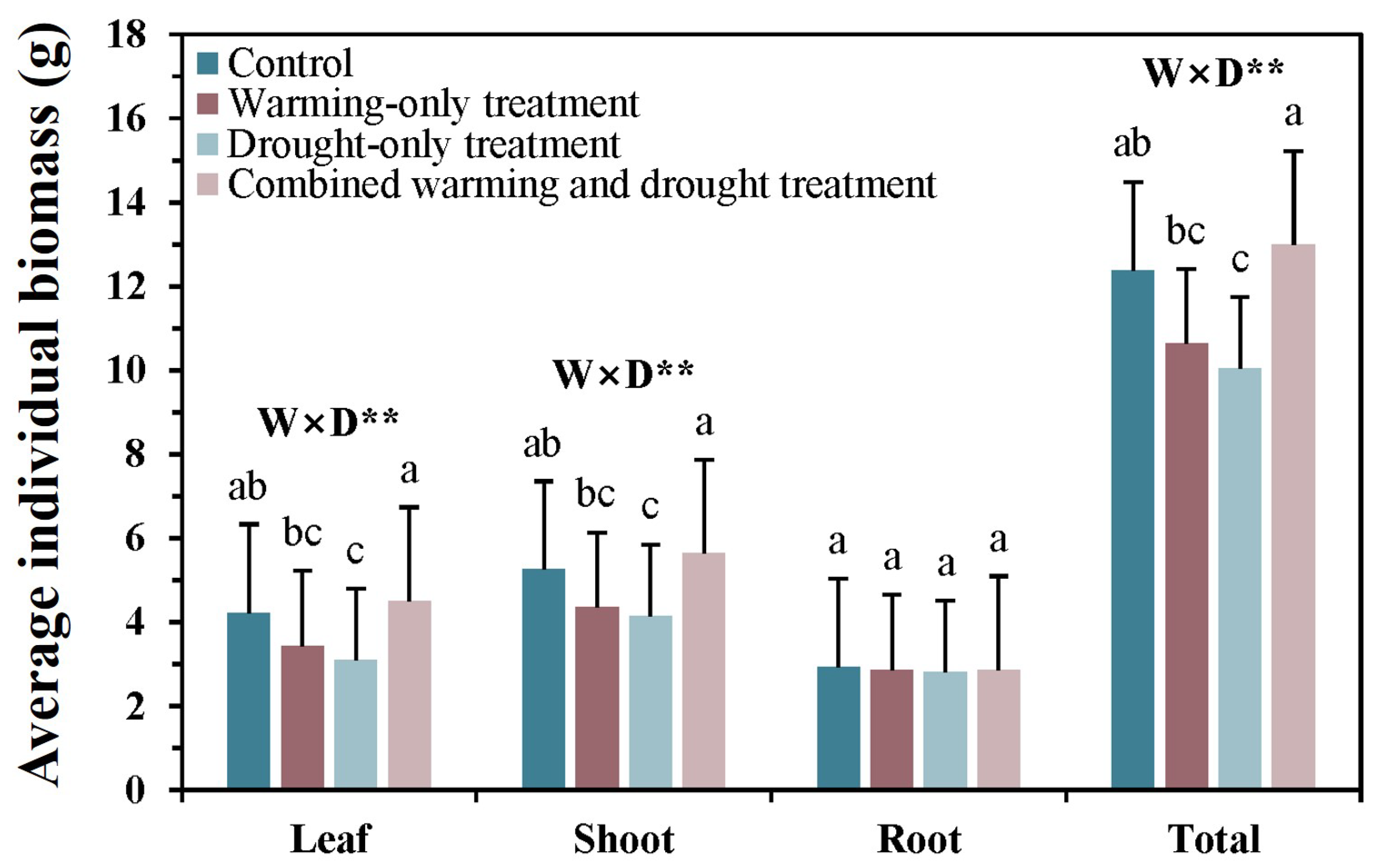
The average individual biomass of leaf, shoot, root, and the total in the control plots were 4.4, 5.6, 3.0, and 12.9 g, respectively (Figure 4). Leaf, shoot, and total biomass were greater in the combined warming and drought treatment than in the warming-only and drought-only plots. Meanwhile, there were no significant effects of warming and drought on the leaf, shoot, and total biomass, but there was a significant interaction effect between warming and drought treatments. However, there was no significant difference in root biomass among treatments nor significant effects of warming and drought treatments on root biomass.
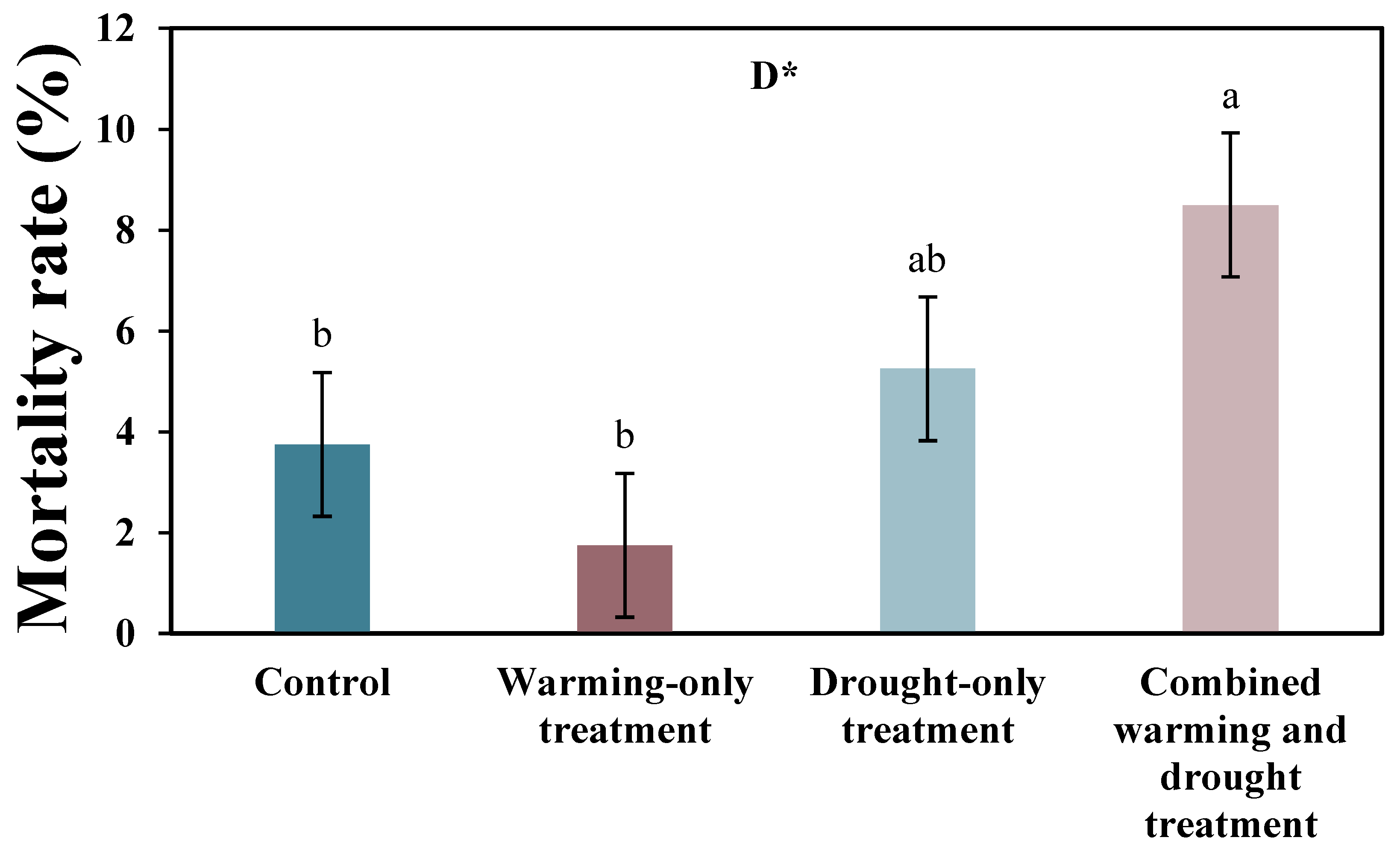
The average mortality rate at the end of the growing season in the control, warming-only, drought-only, and combined warming and drought plots were 3.8, 1.8, 5.3, and 8.5, respectively (Figure 5). In particular, the drought treatment significantly increased the mortality rate, and the mortality rate in the combined warming and drought plots was significantly higher than in the other plots.
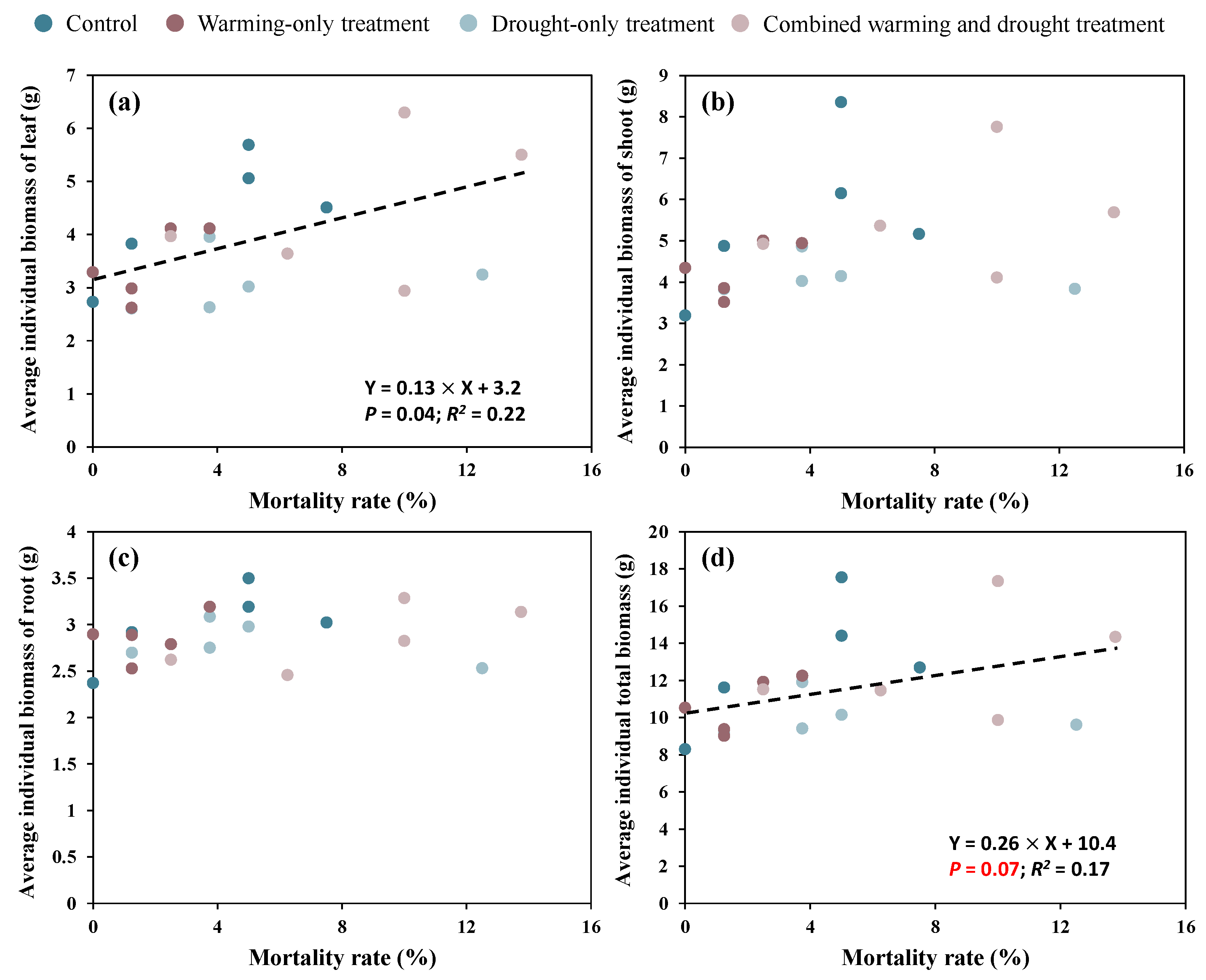
There was a significant linear relationship (p = 0.04) between the mortality rate at the end of the growing season and the average individual biomass of leaf (Figure 6a). The leaf biomass increased by 1.3 g per 10% increase in mortality rate. Additionally, there was a quasi-significant linear relationship (p = 0.07) between the mortality rate and the average individual total biomass, showing a 2.6 g increase in total biomass per 10% increase in the mortality rate (Figure 6d). However, there were no significant linear relationships between the mortality rate and the average individual biomass of shoot and roots (Figure 6c,d).
4. Discussion
The increase in air temperature by 4.0 °C under the warming treatment and the decrease in soil moisture content under the drought treatment showed that our climate manipulation system was operating properly (Figure 2). Meanwhile, our findings showed that the drought treatment increased the mortality rate (Figure 5). These results are consistent with those of prior research, which showed that the 1982–1983 El Niño-related drought was associated with increased seedling mortality rates [30]. In general, soil moisture conditions are significant for the survival of seedlings in the early stage. In the case of anisohydric plants, stomatal closure does not occur even during the drying period. Therefore, the leaf moisture potential remains high [26]. Our findings suggested that L. kaempferi exhibits anisohydric characteristics [26]. Under the reduced moisture conditions, the survival rate was lowered by continuing gas exchange without closing the stomata until death due to water loss.
However, the warming treatment did not significantly affect the mortality rate by itself (Figure 5). Similar effects of increased temperature on the mortality rate of seedlings have been previously reported; the seedling survival rate of Pinus densiflora showed no significant difference when experimental warming increased by 3 °C [26,31]. The increased air temperature in this study (i.e., 4 °C) may have been insufficient to induce substantial changes in the mortality rate. In addition, the drought treatment was carried out for four consecutive weeks, whereas the warming treatment had a rest period in the middle (i.e., the third week in May).
Our results are in line with previous studies where warming and drought treatments constrain seedling height and root collar diameter increments but only immediately after treatment. In our experiment, seedling height and root collar diameter increments decreased under drought treatments, consistent with previous findings with Abies fabri [32]. This suggests that our drought treatments induced heat and water stress in L. kaempferi seedlings. However, the analyses of seedling height, root collar diameter, and biomass five months after treatment revealed that seedlings exposed to the combined treatment had the highest mortality rates but showed the highest growth. Geange et al. [33] reported similar results in Aciphylla glacialis. Environmental stress significantly increased seedling mortality, but the surviving seedlings grew better than their counterparts.
The positive effects of environmental stress on plant growth have been poorly discussed in the literature, and the mechanisms remain unknown [34]. Moreover, few studies have shown that high temperature and drought stress can trigger a series of physio-biochemical compensations in plants. The higher growth in the combined warming–drought treatment might be due to overcompensation [35] for the disappearance of the stressful treatments, the low competition effect [36] of the surviving seedlings, and the development of adaptive mechanisms by slowly increasing stress. Numerous variables, such as genotype, nutritional status, light, competition with other plants, plant phenology, plant size, severity and length of drought, and rehydration duration, impact the physiological and morphological characteristics of re-watered plants [37].
Overcompensation is typically associated with a hormetic effect, which occurs when the homeostasis of an organism is disturbed [38,39,40,41]. If cells are exposed to stresses that disrupt homeostasis, the unfolded protein response, endoplasmic reticulum-associated degradation, and the heat shock response will all become active to aid recovery [42]. It has been shown that activating the heat shock response results in increased amounts of heat shock proteins, which alleviate a load of aberrant proteins by enabling them to refold their original shape [43]. Cells also demonstrate higher concentrations of components from the ubiquitin-proteasome system after being exposed to stress, and as a result, they have a greater ability to destroy proteins that cannot be refolded as a result [44]. The modulation of heat shock protein levels and overexpression of ubiquitin-proteasome system components may be stated to be a reliable molecular basis for the positive effects of hormesis [42]. Wei et al. [45] found that the net photosynthetic rate, stomatal conductance, soluble protein, malondialdehyde, and chlorophyll content increased at 6–9 days after high-temperature stress, which was a significant driving factor for the compensatory growth of Brassica rapa. Zhou et al. [46] discovered that after 6 days of drought stress and rehydration, proline, soluble protein, superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX) activity significantly increased. High levels of SOD and CAT may successfully eliminate reactive oxygen species and APX enzyme activity could mitigate membrane damage and lipid peroxidation [47].
Another possibility is that seedlings in the combined warming–drought treatment experienced decreased intraspecific competition as the mortality rate increased. Several researchers have observed that intraspecific competition reduces plant biomass [48,49]. An increased plant density may result in an asymmetrical frequency distribution of plants, resulting in size variations among individual plants [50]. For instance, if there is competition for water, individuals with more extensive roots may reduce the water available to ones with smaller roots, thereby suppressing their development. In addition, in a symmetrical competitive response, the biomass production of each individual declines uniformly [49]. Horner et al. [51] discovered that high-density stands were dominated by thin trees. In contrast, low-density stands generated distributions with a broad range of stem sizes and a high mean and maximum stem diameter.
Slowly increasing stresses may produce physiological adaptations that protect plants against the growth inhibition and damage that result from suddenly applied environmental stresses [52]. The stress imposed in our experiment could correspond to a slowly increasing stress, so a mechanism protecting the plants may have been at work. The exposure of plants to sub-lethal high temperatures can enhance their thermotolerance [53]. Potential modes of the reaction include the formation of heat-shock proteins, isoprene, and antioxidants to safeguard the photosynthetic apparatus and cellular metabolism [52]. Seedlings previously subjected to water stress often experience less inhibition of development and other processes upon transplantation than seedlings not previously subjected to water stress [54]. Typically, controlled wetting and drying cycles promote early bud break, dormancy, and drought resistance [55]. In many species, improved drought tolerance is connected with osmotic adjustment, including the buildup of osmotically active chemicals after such cycles [52].
However, the drought-only treated seedlings, which had a high mortality rate due to drought stress, did not show as much recovery as the combined warming–drought treatment. This might be due to the combined stress inducing different, and sometimes antagonistic, signaling pathways [56]. In particular, plants growing in the field are subjected to diverse co-occurring abiotic stimuli that most likely cannot be extrapolated from the sum of individual stresses that affect plant metabolism in unique ways [57]. Similar findings have been obtained for Pinus sylvestris [58] under heat and drought conditions and under herbivory and competitive conditions [59]. These findings demonstrate superior stress resistance and resilience of L. kaempferi even when the mortality rate increases.
5. Conclusions
In our study, warming and drought treatments increased the mortality rate and decreased growth during the treatment period. However, after the cessation of the treatment, seedlings in the combined warming and drought plots showed higher individual biomass growth compared to seedlings that experienced either warming or drought treatment. We suggest that this is because the higher mortality rate under the combined warming and drought plots might result in relaxation in seedling competition for resources, such as water, light, and nutrients, which can lead to an increase in individual biomass growth. However, these mechanisms do not necessarily lead to a positive effect during early spring warming and drought stress in L. kaempferi seedlings. Extreme and persistent stresses are likely to result in a reduction in growth and high mortality rates. Therefore, even if 1-year-old L. kaempferi seedlings experience spring warming and drought events at a similar level to this study in the future, it is expected that the reduction in growth will sufficiently recover when they are exposed to the average temperature and precipitation in summer and autumn. To accurately predict stress response dynamics under climate change, further studies should be conducted on various stress levels, molecular-based protection mechanisms, such as the antioxidant systems, and distance independent competition indices. In addition, it is necessary to collect information on lateral spread roots or foliage and whether the crowns were in contact to understand the response to competition better.
Author Contributions
Conceptualization, H.K., H.J. and G.-J.K.; methodology, H.K., H.J. and G.-J.K.; software, H.K., H.J. and G.-J.K.; investigation, H.K., H.J. and G.-J.K.; resources, H.K., H.J. and G.-J.K.; writing—original draft preparation, H.K.; writing—review and editing, H.K., H.J., G.-J.K., H.-S.K. and Y.S.; visualization, H.K.; supervision, Y.S.; project administration, H.K. and G.-J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the R&D Program for Forest Science Technology (Project No. “2020181B10-2222-BB01”) provided by Korea Forest Service (Korea Forestry Promotion Institute). This research was also supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2021R1A6A1A10045235).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This study was carried out with the support of the Korea Forest Service Government (KFSG) as a graduate school specialization in carbon sinks.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Intergovernmental Panel on Climate Change (IPCC). Summary for Policymakers. In Global Warming of 1.5 °C; IPCC: Geneva, Switzerland, 2018; pp. 51–54. [Google Scholar]
- Mukherjee, S.; Mishra, A.; Trenberth, K.E. Climate change and drought: A perspective on drought indices. Curr. Clim. Chang. Rep. 2018, 4, 145–163. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2021; pp. 1–2913. [Google Scholar]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Li, C. Some aspects of leaf water relations in four provenances of Eucalyptus Microtheca Seedlings. For. Ecol. Manag. 1998, 111, 303–308. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.; Estiarte, M.; Peñuelas, J. Drought advances spring growth phenology of the Mediterranean shrub Erica multiflora. Plant Biol. 2011, 13, 252–257. [Google Scholar] [CrossRef]
- Kim, H.; Jo, H.; Kim, G.J.; Kim, H.S.; Son, Y. Short-term physiological responses of Larix Kaempferi Seedlings Spring Warm. Drought Manip. For. Sci. Technol. 2021, 17, 197–205. [Google Scholar]
- Buermann, W.; Forkel, M.; O’sullivan, M.; Sitch, S.; Friedlingstein, P.; Haverd, V.; Jain, A.K.; Kato, E.; Kautz, M.; Lienert, S.; et al. Widespread seasonal compensation effects of spring warming on northern plant productivity. Nature 2018, 562, 110–114. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Andy Black, T.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef] [Green Version]
- Buermann, W.; Bikash, P.R.; Jung, M.; Burn, D.H.; Reichstein, M. Earlier springs decrease peak summer productivity in North American boreal forests. Environ. Res. Lett. 2013, 8, 024027. [Google Scholar] [CrossRef]
- Casper, B.; Forseth, I.; Kempenich, H.; Seltzer, S.; Xavier, K. Drought prolongs leaf life span in the herbaceous desert perennial Cryptantha flava. Funct. Ecol. 2001, 15, 740–747. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.S.; Campioli, M.; Vitasse, Y.; De Boeck, H.J.; Van den Berge, J.; AbdElgawad, H.; Asard, H.; Piao, S.; Deckmyn, G.; Janssens, I.A. Variation in leaf flushing date influences autumnal senescence and next year’s flushing date in two temperate tree species. Proc. Natl. Acad. Sci. USA 2014, 111, 7355–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus Sargentii Larix Kaempferi Seedlings Using Morphol. Physiol. Parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Carey, J.C.; Tang, J.; Templer, P.H.; Kroeger, K.D.; Crowther, T.W.; Burton, A.J.; Dukes, J.S.; Emmett, B.; Frey, S.D.; Heskel, M.A.; et al. Temperature response of soil respiration largely unaltered with experimental warming. Proc. Natl. Acad. Sci. USA 2016, 113, 13797–13802. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.; Woodin, S.J. Climate change in the Arctic: Using plant functional types in a meta-analysis of field experiments. Funct. Ecol. 2002, 16, 4–17. [Google Scholar] [CrossRef]
- Shaver, G.R.; Jonasson, S. Response of Arctic ecosystems to climate change: Results of long-term field experiments in Sweden and Alaska. Polar Res. 1999, 18, 245–252. [Google Scholar] [CrossRef]
- Rustad, L.E. From transient to steady-state response of ecosystems to atmospheric CO2-enrichment and global climate change: Conceptual challenges and need for an integrated approach. Plant Ecol. 2006, 182, 43–62. [Google Scholar]
- Jo, H.; Chang, H.; An, J.; Cho, M.S.; Son, Y. Species specific physiological responses of Pinus Densiflora Larix Kaempferi Seedlings Open-Field Exp. Warm. Precip. Manip. For. Sci. Technol. 2019, 15, 44–50. [Google Scholar]
- Chang, H.; An, J.; Roh, Y.; Son, Y. Experimental warming and drought treatments reduce physiological activities and increase mortality of Pinus Koraiensis Seedlings. Plant Ecol. 2020, 221, 515–527. [Google Scholar] [CrossRef]
- Misson, L.; Degueldre, D.; Collin, C.; Rodriguez, R.; Rocheteau, A.; OURCIVAL, J.M.; Rambal, S. Phenological responses to extreme droughts in a Mediterranean forest. Glob. Chang. Biol. 2011, 17, 1036–1048. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Korea Forest Service. Start Production of Cloned Larch Seedlings for Economic Forest Development. Available online: https://nifos.forest.go.kr/kfsweb/cop/bbs/selectBoardArticle.do;jsessionid=aIMJ8WfBNqtyuhk7ttW6wJsMpyuUB2jDi1CC7LPVOljdWlsq4gG3CpCJOqd9wEs7.frswas02_servlet_engine5?nttId=3145455&bbsId=BBSMSTR_1036&pageUnit=10&pageIndex=1&searchtitle=title&searchcont=&searchkey=&searchwriter=&searchWrd=&ctgryLrcls=CTGRY150&ctgryMdcls=&ctgrySmcls=&ntcStartDt=&ntcEndDt=&mn=UKFR_03_03&orgId=kfri (accessed on 6 January 2021).
- NIFoS. Research Report on Korea Forest Research Institute. National Institute of Forest Science. 2012. Economic Tree Species4; NIFos: Seoul, Korea, 2012; p. 16. [Google Scholar]
- An, J.; Chang, H.; Park, M.J.; Han, S.H.; Hwang, J.; Cho, M.S.; Son, Y. Effect of Experimental Warming on Physiological and Growth Responses of Larix Kaempferi Seedlings. J. Clim. Change Res. 2016, 7, 77–84. [Google Scholar] [CrossRef]
- Kwon, B.; Cho, M.S.; Yang, A.R.; Chang, H.; An, J.; Son, Y. The early growth performances of Pinus Densiflora Larix Kaempferi Seedlings Open-Field Exp. Warm. Precip. Manip. J. Korean Soc. For. Sci. 2020, 109, 31–40. [Google Scholar]
- Fisichelli, N.; Wright, A.; Rice, K.; Mau, A.; Buschena, C.; Reich, P.B. First-year seedlings and climate change: Species-specific responses of 15 North American tree species. Oikos 2014, 123, 1331–1340. [Google Scholar] [CrossRef]
- Haase, D.L. Understanding forest seedling quality: Measurements and interpretation. Tree Plant. Notes 2008, 52, 24–30. [Google Scholar]
- Climate Information Portal. Available online: http://www.climate.go.kr/home/CCS/contents/33_2_areapoint_basic.php (accessed on 14 October 2020).
- Gilbert, G.S.; Harms, K.E.; Hamill, D.N.; Hubbell, S.P. Effects of seedling size, El Niño drought, seedling density, and distance to nearest conspecific adult on 6-year survival of Ocotea Whitei Seedlings Panamá. Oecologia 2001, 127, 509–516. [Google Scholar] [CrossRef]
- Cho, M.S.; Hwang, J.; Yang, A.R.; Han, S.; Son, Y. Seed germination and seedling survival rate of Pinus Densiflora Abies Holophylla Open-Field Exp. Warm. Using Infrared Lamp. J. Korean Soc. For. Sci. 2014, 103, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, G.; Yang, L.; Guo, J. Effects of drought and warming on biomass, nutrient allocation, and oxidative stress in Abies Fabri East. Tibet. Plateau. J. Plant Growth Regul. 2013, 32, 298–306. [Google Scholar] [CrossRef]
- Geange, S.R.; Holloway-Phillips, M.M.; Briceño, V.F.; Nicotra, A.B. Aciphylla Glacialis Mortality, Growth Frost Resist. A Field Warm. Exp. Aust. J. Bot. 2020, 67, 599–609. [Google Scholar] [CrossRef]
- Lin, R.; Wang, X.; Luo, Y.; Du, W.; Guo, H.; Yin, D. Effects of soil cadmium on growth, oxidative stress and antioxidant system in wheat seedlings (Triticum Aestivum L.). Chemosphere 2007, 69, 89–98. [Google Scholar] [CrossRef]
- Correia, B.; Hancock, R.D.; Amaral, J.; Gomez-Cadenas, A.; Valledor, L.; Pinto, G. Combined drought and heat activates protective responses in Eucalyptus globulus that are not activated when subjected to drought or heat stress alone. Front. Plant Sci. 2018, 9, 819. [Google Scholar] [CrossRef] [Green Version]
- Alexander, H.M.; Mihail, J.D. Seedling disease in an annual legume: Consequences for seedling mortality, plant size, and population seed production. Oecologia 2000, 122, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Correia, B.; Pintó-Marijuan, M.; Neves, L.; Brossa, R.; Dias, M.C.; Costa, A.; Castro, B.B.; Araújo, C.; Santos, C.; Chaves, M.M.; et al. Water stress and recovery in the performance of two Eucalyptus globulus clones: Physiological and biochemical profiles. Physiol. Plant. 2014, 150, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Aina, R.; Labra, M.; Fumagalli, P.; Vannini, C.; Marsoni, M.; Cucchi, U.; Bracale, M.; Sgorbati, S.; Citterio, S. Thiol-peptide level and proteomic changes in response to cadmium toxicity in Oryza Sativa L. Roots. Environ. Exp. Bot. 2007, 59, 381–392. [Google Scholar] [CrossRef]
- Von Zglinicki, T.; Edwall, C.; Ostlund, E.; Lind, B.; Nordberg, M.; Ringertz, N.; Wroblewski, J. Very low cadmium concentrations stimulate DNA synthesis and cell growth. J. Cell Sci. 1992, 103, 1073–1081. [Google Scholar] [CrossRef]
- Beyersmann, D.; Hechtenberg, S. Cadmium, gene regulation, and cellular signalling in mammalian cells. Toxicol. Appl. Pharmacol. 1997, 144, 247–261. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.; Peñuelas, J. Chlorophyll hormesis: Are chlorophylls major components of stress biology in higher plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef]
- Wiegant, F.; Poot, S.d.; Boers-Trilles, V.; Schreij, A. Hormesis and cellular quality control: A possible explanation for the molecular mechanisms that underlie the benefits of mild stress. Dose-Response 2013, 11, 413–430. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, R.I.; Kline, M.P.; Bimston, D.N.; Cotto, J.J. The heat-shock response: Regulation and function of heat-shock proteins and molecular chaperones. Essays Biochem. 1997, 32, 17–29. [Google Scholar]
- Amm, I.; Sommer, T.; Wolf, D.H. Protein quality control and elimination of protein waste: The role of the ubiquitin–proteasome system. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2014, 1843, 182–196. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Chen-xi, S.; He-li, Z.; Qi, H.; Qing-zhou, Z.; Xing-lin, S. Compensatory ability and defense mechanism of Chinese cabbage under high temperature stress. Chin. J. Agrometeorol. 2018, 39, 119. [Google Scholar]
- Zhou, X.; Wang, G.; Liang, Z.; Han, R. Effects of drought stress and rehydration on physiological characteristics and glycyrrhizin accumulation of Glycyrrhiza uralensis fisch. Seedling. Acta Agric. Boreali-Occident. Sin. 2011, 20, 64–70. [Google Scholar]
- Dong, S.; Jiang, Y.; Dong, Y.; Wang, L.; Wang, W.; Ma, Z.; Yan, C.; Ma, C.; Liu, L. A study on soybean responses to drought stress and rehydration. Saudi J. Biol. Sci. 2019, 26, 2006–2017. [Google Scholar] [CrossRef] [PubMed]
- Kleunen, M.v.; Fischer, M.; Schmid, B. Effects of intraspecific competition on size variation and reproductive allocation in a clonal plant. Oikos 2001, 94, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.W.; Showalter, A.M.; Ungar, I.A. Effects of intraspecific competition on growth and photosynthesis of Atriplex Prostrata. Aquat. Bot. 2005, 83, 187–192. [Google Scholar] [CrossRef]
- Drake, D.R.; Ungar, I.A. Effects of salinity, nitrogen, and population density on the survival, growth, and reproduction of Atriplex Triangularis (Chenopodiaceae). Am. J. Bot. 1989, 76, 1125–1135. [Google Scholar] [CrossRef]
- Horner, G.J.; Baker, P.J.; Mac Nally, R.; Cunningham, S.C.; Thomson, J.R.; Hamilton, F. Mortality of developing floodplain forests subjected to a drying climate and water extraction. Glob. Chang. Biol. 2009, 15, 2176–2186. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Levitt, J. Response of plants to environmental stresses: Chilling, freezing, and high temperature stresses. Physiol. Ecol. A Ser. Monogr. Texts Treatises 1980, 1, 23–64. [Google Scholar]
- Ladjal, M.; Epron, D.; Ducrey, M. Effects of drought preconditioning on thermotolerance of photosystem II and susceptibility of photosynthesis to heat stress in cedar seedlings. Tree Physiol. 2000, 20, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Mexal, J.G.; South, D.B. Bareroot seedling culture. For. Regen. Man. 1991, 89–115. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Rivero, R.M.; Martínez, V.; Gómez-Cadenas, A.; Arbona, V. Tolerance of citrus plants to the combination of high temperatures and drought is associated to the increase in transpiration modulated by a reduction in abscisic acid levels. BMC Plant Biol. 2016, 16, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehschuh, R.; Ruehr, N.K. Unrevealing water and carbon relations during and after heat and hot drought stress in Pinus sylvestris. BioRxiv 2021. [Google Scholar] [CrossRef]
- Edenius, L.; Danell, K.; Bergström, R. Impact of herbivory and competition on compensatory growth in woody plants: Winter browsing by moose on Scots pine. Oikos 1993, 66, 286–292. [Google Scholar] [CrossRef]
Figure 1.
Seedling arrangement and spacing per plot.

Figure 2.
Mean weekly changes in air temperature (a), soil temperature (b), and soil moisture content (c) in May 2020. Error bars are the standard errors of the means, and asterisks (*) indicate significant differences between the warming and drought treatments (** p < 0.01; *** p < 0.001). W: warming effect; D: drought effect; W × D: interaction effect of warming and drought.
Figure 2.
Mean weekly changes in air temperature (a), soil temperature (b), and soil moisture content (c) in May 2020. Error bars are the standard errors of the means, and asterisks (*) indicate significant differences between the warming and drought treatments (** p < 0.01; *** p < 0.001). W: warming effect; D: drought effect; W × D: interaction effect of warming and drought.

Figure 3.
Height increment (a) and root collar diameter increment (b) of Larix kaempferi seedlings in 2020. Bars with different letters are significantly different among treatments at p < 0.05. Error bars are the standard errors of the means, and asterisks (*) indicate significant differences between the warming and drought treatments (** p < 0.01; *** p < 0.001). W: warming effect; D: drought effect; W × D: interaction effect of warming and drought.
Figure 3.
Height increment (a) and root collar diameter increment (b) of Larix kaempferi seedlings in 2020. Bars with different letters are significantly different among treatments at p < 0.05. Error bars are the standard errors of the means, and asterisks (*) indicate significant differences between the warming and drought treatments (** p < 0.01; *** p < 0.001). W: warming effect; D: drought effect; W × D: interaction effect of warming and drought.

Figure 4.
Average individual biomass of Larix kaempferi seedlings as influenced by warming and drought treatments in November 2020. Bars with different letters are significantly different among treatments at p < 0.05. Error bars are the standard errors of the means, and asterisks (*) indicate significant differences between the warming and drought treatments (** p < 0.01). W: warming effect; D: drought effect; W × D: interaction effect of warming and drought.
Figure 4.
Average individual biomass of Larix kaempferi seedlings as influenced by warming and drought treatments in November 2020. Bars with different letters are significantly different among treatments at p < 0.05. Error bars are the standard errors of the means, and asterisks (*) indicate significant differences between the warming and drought treatments (** p < 0.01). W: warming effect; D: drought effect; W × D: interaction effect of warming and drought.

Figure 5.
Mortality of Larix kaempferi seedlings in 2020. Bars with different letters are significantly different among treatments at p < 0.05. Error bars represent standard errors of the mean. The asterisk (*) indicates a significant difference in the drought treatment (* p < 0.05). D: drought effect.
Figure 5.
Mortality of Larix kaempferi seedlings in 2020. Bars with different letters are significantly different among treatments at p < 0.05. Error bars represent standard errors of the mean. The asterisk (*) indicates a significant difference in the drought treatment (* p < 0.05). D: drought effect.

Figure 6.
Relationships between the mortality rate at the end of the growing season and the average individual biomass of leaf (a), shoot (b), root (c), and the total biomass (d). Black dashed lines indicate linear relation ships between two variables.
Figure 6.
Relationships between the mortality rate at the end of the growing season and the average individual biomass of leaf (a), shoot (b), root (c), and the total biomass (d). Black dashed lines indicate linear relation ships between two variables.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, H.; Jo, H.; Kim, G.-J.; Kim, H.-S.; Son, Y. Effects of Spring Warming and Drought Events on the Autumn Growth of Larix kaempferi Seedlings. Water 2022, 14, 1962. https://0-doi-org.brum.beds.ac.uk/10.3390/w14121962
AMA Style
Kim H, Jo H, Kim G-J, Kim H-S, Son Y. Effects of Spring Warming and Drought Events on the Autumn Growth of Larix kaempferi Seedlings. Water. 2022; 14(12):1962. https://0-doi-org.brum.beds.ac.uk/10.3390/w14121962
Chicago/Turabian StyleKim, Hyeonji, Heejae Jo, Gwang-Jung Kim, Hyung-Sub Kim, and Yowhan Son. 2022. "Effects of Spring Warming and Drought Events on the Autumn Growth of Larix kaempferi Seedlings" Water 14, no. 12: 1962. https://0-doi-org.brum.beds.ac.uk/10.3390/w14121962
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.