
A Simulation of the Use of High Throughput Sequencing as Pre-Screening Assay to Enhance the Surveillance of Citrus Viruses and Viroids in the EPPO Region


Abstract
:1. Introduction
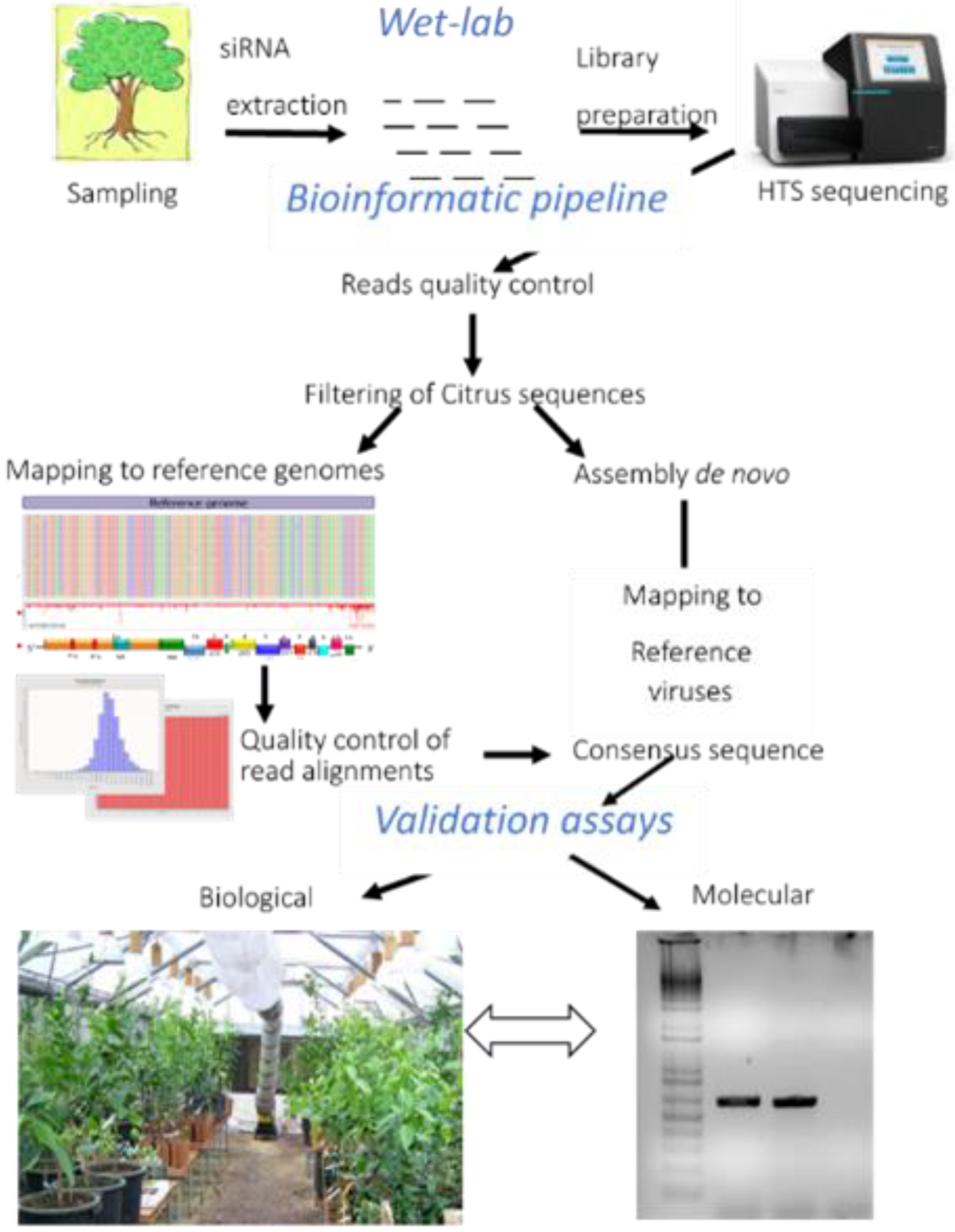
2. Materials and Methods
2.1. Source Plants and Small-RNA Libraries
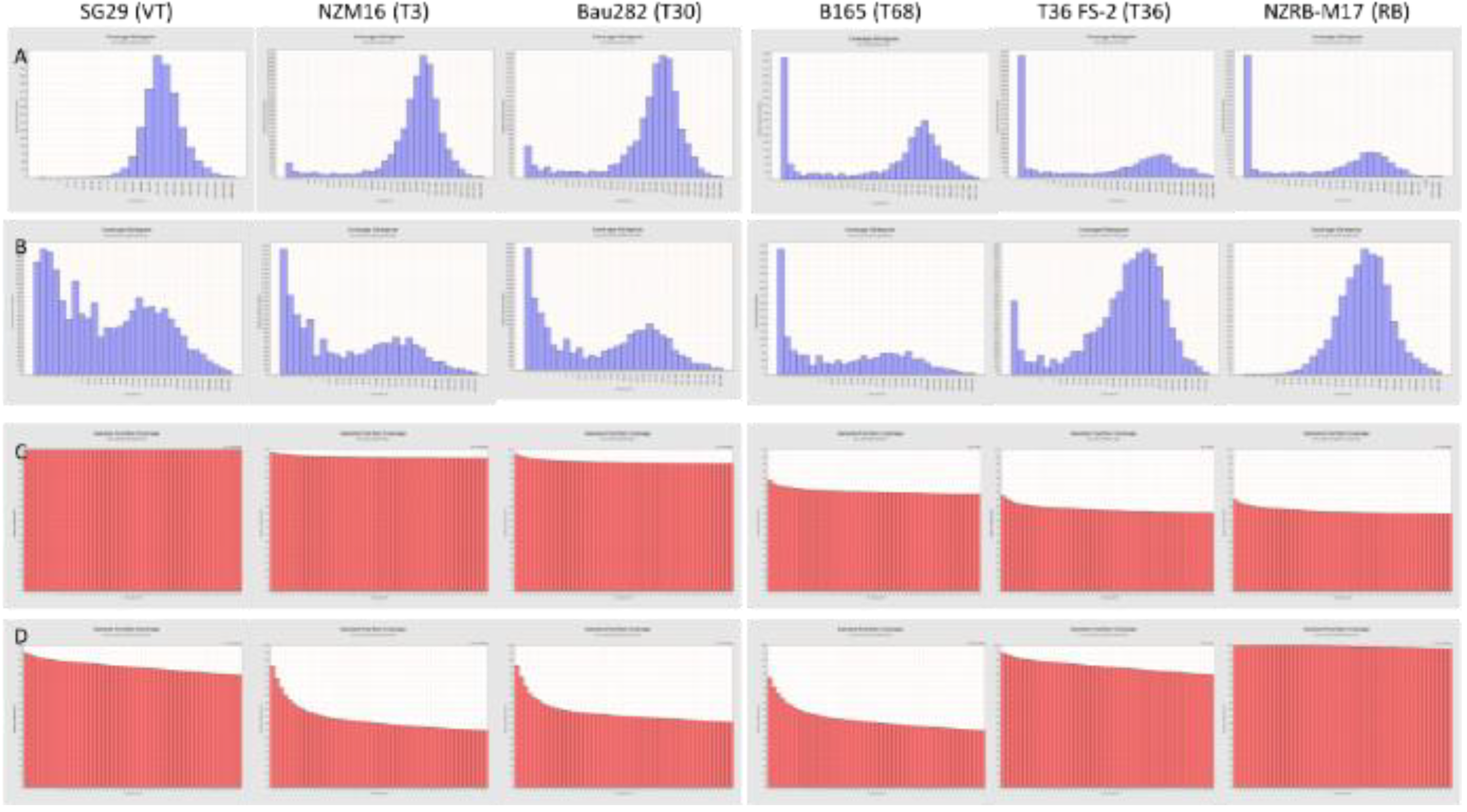
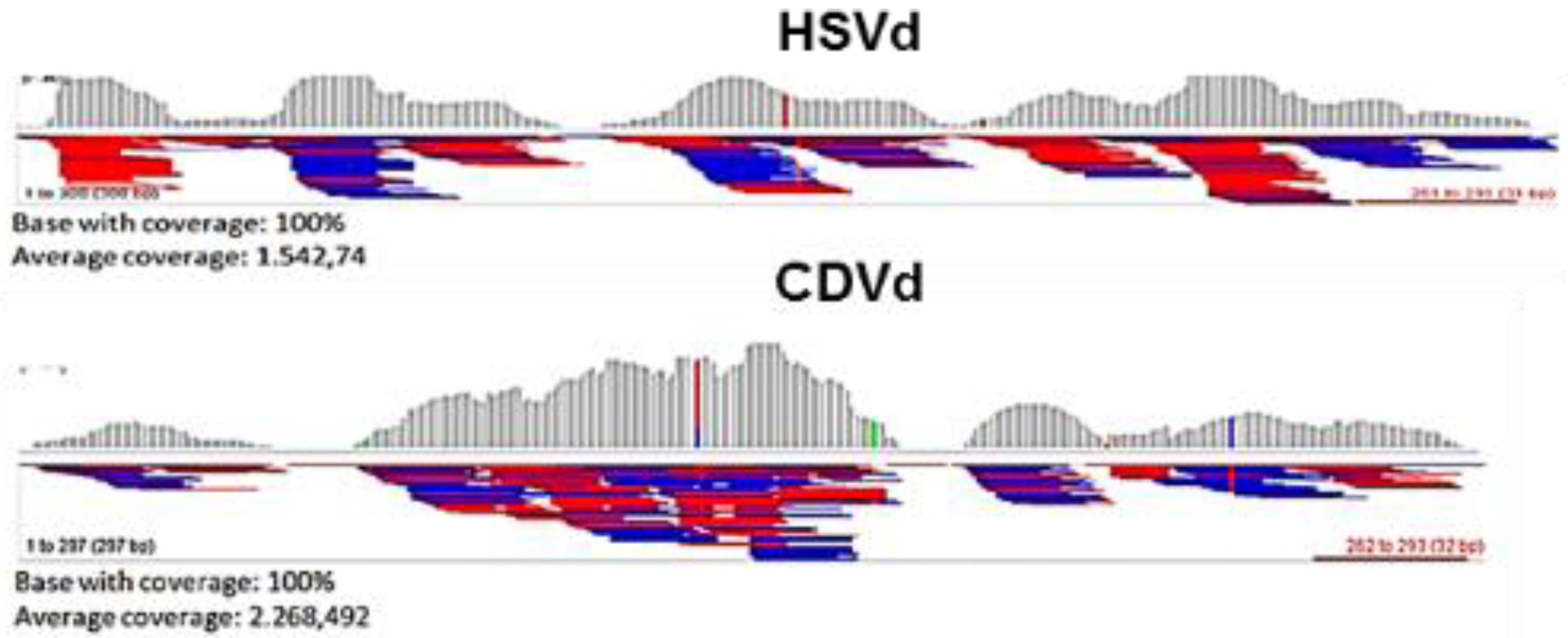
2.2. Data Evaluation by Bioinformatics
2.3. Biological Indexing
2.4. RT-PCR and Real-Time RT-PCR
3. Results
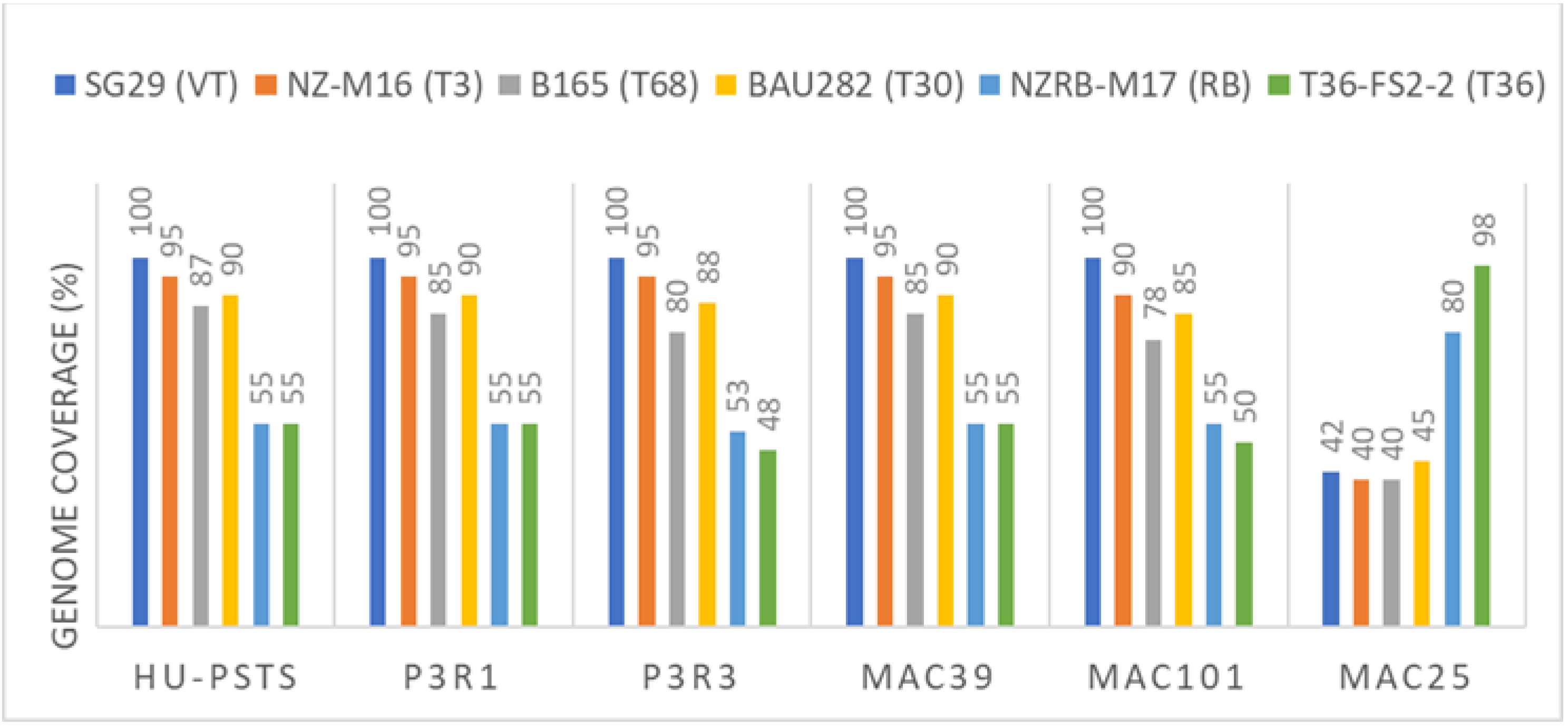
3.1. Searching for Non-EU CTV Isolates
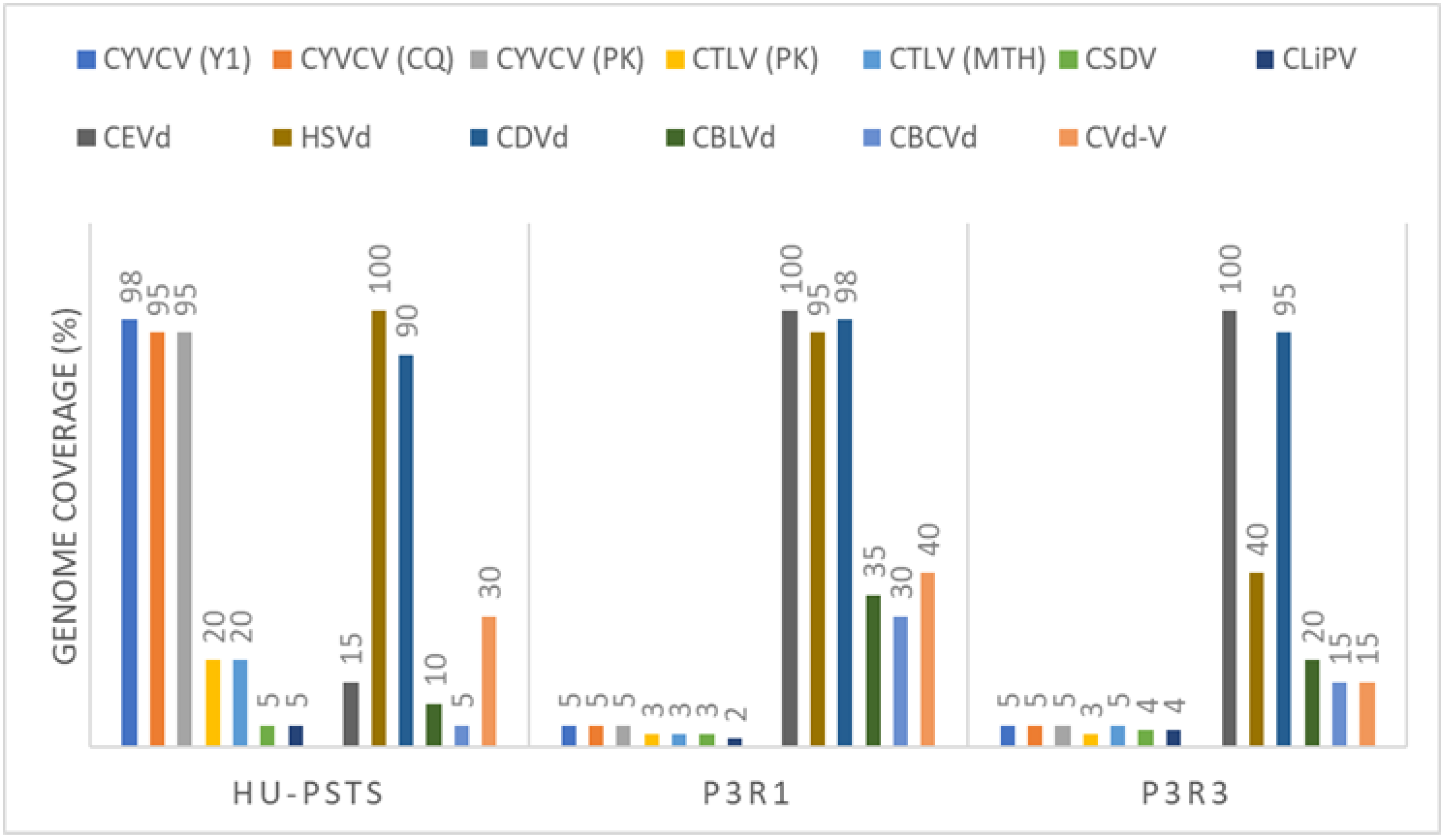
3.2. Searching for Non-EU Virus and Viroids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, C.; Da Graça, J.V.; Freitas-Astúa, J.; Vidalakis, G.; Duran-Vila, N.; Lavagi, I. Citrus viruses and viroids. In The Genus Citrus, 2nd ed.; Talon, M., Caruso, M., Gmitter, F., Jr., Eds.; Woodhead Publishing: Cambridge, UK; Elsevier: Amsterdam, The Netherlands, 2020; pp. 391–410. [Google Scholar]
- International Plant Protection Convention (IPPC); Food and Agriculture Organization of the United Nations (FAO). Surveillance ISPM 6; International Plant Protection Convention (IPPC): Rome, Italy; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2018. [Google Scholar]
- Ochoa-Corona, F.M. Biosecurity, microbial forensics and plant pathology: Education challenges, overlapping disciplines and research needs. Australas. Plant Pathol. 2011, 40, 335–338. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Schrader, G.; Camilleri, M.; Diakaki, M.; Vos., S. Pest survey card on non-European isolates of citrus tristeza virus. Survey card on non-EU isolates of Citrus tristeza virus. EFSA Support. Publ. 2019, EN-1600. [Google Scholar] [CrossRef] [Green Version]
- OEPP/EPPO. EPPO A1 and A2 Lists of Pests Recommended for Regulation as Quarantine Pests (2020). 2019. Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/A1_list (accessed on 23 March 2020).
- EFSA PLH Panel (EFSA Panel on Plant Health). Scientific opinion of pest categorisation of “blight and blight-like” diseases of citrus. EFSA J. 2018, 16, 5248. [Google Scholar]
- Tan, S.-H.; Osman, F.; Bodaghi, S.; Dang, T.; Greer, G.; Huang, A.; Hammado, S.; Abu-Hajar, S.; Campos, R.; Vidalakis, G. Full genome characterization of 12 citrus tatter leaf virus isolates for the development of a detection assay. PLoS ONE 2019, 14, e0223958. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Hartung, J.S. Cloning and sequence analysis of an infectious clone of citrus yellow mosaic virus that can infect sweet orange via agrobacterium-mediated inoculation. J. Gen. Virol. 2001, 82, 2549–2558. [Google Scholar] [CrossRef]
- Ahlawat, Y.S.; Varma, A.; Lockhart, B.E.; Srivastava, M.; Chakraborty, N.K.; Varma, A. Association of a badnavirus with citrus mosaic disease in India. Plant Dis. 1996, 80, 590–592. [Google Scholar] [CrossRef]
- EFSA PLH Panel (EFSA Panel on Plant Health). Scientific opinion on the pest categorisation of Citrus tristeza virus (non-European isolates). EFSA J. 2017, 15, 5031. [Google Scholar]
- Jakse, J.; Radisek, S.; Pokorn, T.; Matousek, J.; Javornik, B. Deep-Sequencing revealed citrus bark cracking viroid (CBCVd) as a highly aggressive pathogen on hop. Plant Pathol. 2015, 64, 831–842. [Google Scholar] [CrossRef]
- Bani Hashemian, S.M.; Serra, P.; Barbosa, C.J.; Juárez, J.; Aleza, P.; Corvera, J.M.; Lluch, A. The effect of a field-source mixture of citrus viroids on the performance of ‘Nules’ clementine and ‘Navelina’ sweet orange trees grafted on carrizo citrange. Plant Dis. 2009, 93, 699–707. [Google Scholar] [CrossRef]
- Murcia, N.; Bani Hashemian, S.M.; Serra, P.; Pina, J.A.; Duran-Vila, N. Citrus viroids: Symptom expression and performance of Washington navel sweet orange trees grafted on carrizo citrange. Plant Dis. 2015, 99, 125–136. [Google Scholar] [CrossRef]
- Liu, C.; Liu, H.; Hurst, J.; Timko, M.P.; Zhou, C. Recent advances on citrus yellow vein clearing virus in citrus. Hort. Plant J. 2020, 6, 216–222. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, S.; Navarro, B.; Gisel, A.; Peña, L.; Navarro, L.; Moreno, P.; Di Serio, F.; Flores, R. Citrus tristeza virus infection induces the accumulation of viral small RNAs (21–24-nt) mapping preferentially at the 3′-terminal region of the genomic RNA and affects the host small RNA profile. Plant Mol. Biol. 2011, 75, 607–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, S.J. Citrus tristeza virus: Evolution of complex and varied genotypic groups. Front. Microbiol. 2013, 4, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokomi, R.K.; Selvaraj, V.; Maheshwari, Y.; Saponari, M.; Giampetruzzi, A.; Chiumenti, M.; Hajeri, S. Identification and characterization of Citrus tristeza virus isolates breaking resistance in trifoliate orange in California. Phytopathology 2017, 107, 901–908. [Google Scholar] [CrossRef] [Green Version]
- Zablocki, O.; Pietersen, G. Characterization of a novel citrus tristeza virus genotype within three cross-protecting source GFMS12 sub-isolates in South Africa by means of illumina sequencing. Arch. Virol. 2014, 159, 2133–2139. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, E.E.; Coletta-Filho, H.D.; Nouri, S.; Falk, B.W.; Nerva, L.; Oliveira, T.S.; Dorta, S.O.; Machado, M.A. Deep sequencing analysis of RNAs from citrus plants grown in a citrus sudden death-affected area reveals diverse known and putative novel viruses. Viruses 2017, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhu, D.; Tan, Y.; Zong, X.; Wei, H.; Liu, Q. First report of citrus leaf blotch virus in sweet cherry. Plant Dis. 2015, 100, 1027. [Google Scholar] [CrossRef]
- Jo, Y.; Choi, H.; Kim, S.M.; Kim, S.L.; Lee, B.C.; Cho, W.K. Integrated analyses using RNA-seq data reveal viral genomes, single nucleotide variations, the phylogenetic relationship, and recombination for apple stem grooving virus. BMC Genom. 2016, 17, 579. [Google Scholar] [CrossRef] [Green Version]
- Loconsole, G.; Onelge, N.; Potere, O.; Giampetruzzi, A.; Bozan, O.; Satar, S.; De Stradis, A.; Savino, V.; Yokomi, R.K.; Saponari, M. Identification and characterization of citrus yellow vein clearing virus, a putative new member of the genus mandarivirus. Phytopathology 2012, 102, 1168–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loconsole, G.; Saldarelli, P.; Doddapaneni, H.; Savino, V.; Martelli, G.P.; Saponari, M. Identification of a single-stranded DNA virus associated with citrus chlorotic dwarf disease, a new member in the family Geminiviridae. Virology 2012, 432, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Vives, M.C.; Velázquez, K.; Pina, J.A.; Moreno, P.; Guerri, J.; Navarro, L. Identification of a new enamovirus associated with citrus vein enation disease by deep sequencing of small RNAs. Phytopathology 2013, 103, 1077–1086. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Stone, A.; Otero-Colina, G.; Wei, G.; Choudhary, N.; Achor, D.; Shao, J.; Levy, L.; Nakhla, M.K.; Hollingsworth, C.R.; et al. Genome assembly of Citrus leprosis virus nuclear type reveals a close association with Orchid fleck virus. Genome Announc. 2013, 1, e00519-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Shao, J.; Hartung, J.S.; Schneider, W.; Brlansky, R.H. A case study on discovery of novel Citrus leprosis virus cytoplasmic type 2 utilizing small RNA libraries by next generation sequencing and bioinformatic analyses. J. Data Min. Genom. Proteom. 2013, 4, 129. [Google Scholar] [CrossRef]
- Navarro, B.L.; Minutolo, M.; Stradis, A.D.; Palmisano, F.; Alioto, D.; Di Serio, F. The first phlebo-like virus infecting plants: A case study on the adaptation of negative-stranded RNA viruses to new hosts. Mol. Plant Pathol. 2018, 19, 1075–1089. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, S.; Wu, J.; Yang, F.; Zhou, Y.; Zhou, C.; Cao, M. Complete genome sequences and recombination analysis of three divergent satsuma dwarf virus isolates. Trop. Plant Pathol. 2021, 46, 26–30. [Google Scholar] [CrossRef]
- Dang, T.; Menchaca, I.; Bodaghi, S.; Lavagi, I.; Osman, F.; Vidalakis, G. Next generation sequencing as a routine diagnostic tool for citrus variety introduction at the citrus clonal protection program. J. Citrus Pathol. 2019, 6, 6. [Google Scholar]
- Chambers, G.; Englezou, A.; Webster, J.; Bogema, D.; Donovan, N. Using next generation sequencing (NGS) to characterize Australia’s living pathogen collection. J. Citrus Pathol. 2019, 6, 4–5. [Google Scholar]
- Olmos, A.; Boonham, N.; Candresse, T.; Gentit, P.; Giovani, B.; Kutnjak, D.; Liefting, L.; Maree, H.J.; Minafra, A.; Moreira, A.; et al. High-Throughput sequencing technologies for plant pest diagnosis: Challenges and opportunities. Bull. OEPP EPPO Bull. 2018, 48, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Bester, R.; Cook, G.; Maree, H.J. Citrus tristeza virus genotype detection using high-throughput sequencing. Viruses 2021, 13, 168. [Google Scholar] [CrossRef]
- Scuderi, G.; Russo, M.; Davino, S.; Ferraro, R.; Catara, A.; Licciardello, G. Occurrence of the T36 genotype of Citrus tristeza virus in citrus orchards in Sicily, Italy. Plant Dis. 2016, 100, 1253. [Google Scholar] [CrossRef]
- Bar-Joseph, M.; Catara, A.F.; Licciardello, G. The puzzling phenomenon of seedling yellows recovery and natural spread of asymptomatic infections of citrus tristeza virus: Two sides of the same coin. Hortic. Rev. 2021, 48, 339–362. [Google Scholar]
- Licciardello, G.; Scuderi, G.; Ferraro, R.; Russo, M.; Dai, S.M.; Catara, A.; Deng, Z.N. High throughput sequencing of a stem pitting Citrus tristeza virus isolate from Hunan province (P.R. China). J. Citrus Pathol. Under review.
- Flores, R.; Delgado, S.; Gas, M.E.; Carbonell, A.; Molina, D.; Gago, S.; De la Pena, M. Viroids: The minimal non-coding RNAs with autonomous replication. FEBS Lett. 2004, 567, 42–48. [Google Scholar] [CrossRef]
- Chambers, G.A.; Donovan, N.J.; Bodaghi, S.; Jelinek, S.M.; Vidalakis, G. A novel citrus viroid found in Australia, tentatively named citrus viroid VII. Arch. Virol. 2018, 163, 215–218. [Google Scholar] [CrossRef]
- Maree, H.J.; Fox, A.; Al Rwahnih, M.; Boonham, N.; Candresse, T. Application of HTS for routine plant virus diagnostics: State of the art and challenges. Front. Plant Sci. 2018, 9, 1082. [Google Scholar] [CrossRef] [Green Version]
- Massart, S.; Chiumenti, M.; De Jonghe, K.; Glover, R.; Haegeman, A.; Koloniuk, I.; Komínek, P.; Kreuze, J.; Kutnjak, D.; Lotos, L.; et al. Virus detection by high-throughput sequencing of small RNAs: Large-scale performance testing of sequence analysis strategies. Phytopathology 2019, 109, 488–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliogka, V.I.; Minafra, A.; Saldarelli, P.; Ruiz-García, A.B.; Glasa, M.; Katis, N.; Olmos, A. Recent advances on detection and characterization of fruit tree viruses using high-throughput sequencing technologies. Viruses 2018, 10, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Serio, F.; Cao, M. Identification of novel viruses and viroids in citrus hosts by high-throughput sequencing: Methodologies, opportunities, challenges and impact. J. Citrus Pathol. 2019, 6, 10–11. [Google Scholar]
- Licciardello, G.; Ferraro, R.; Scuderi, G.; Russo, M.; Catara, A. Next generation sequencing of small RNAs from citrus plants with single and mixed infections of viruses and viroids. J. Citrus Pathol. 2019, 6, 5–6. [Google Scholar]
- Licciardello, G.; Scuderi, G.; Ferraro, R.; Giampetruzzi, A.; Russo, M.; Lombardo, A.; Raspagliesi, D.; Bar-Joseph, M.; Catara, A. Deep sequencing and analysis of small RNAs in sweet orange grafted on sour orange infected with two Citrus tristeza virus isolates prevalent in Sicily. Arch. Virol. 2015, 160, 2583–2589. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Gen. Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 15, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnsey, S.M.; Civerolo, E.L.; Gumpf, D.J.; Paul, C.; Hilf, M.E.; Lee, R.F.; Brlansky, R.H.; Yokomi, R.K.; Hartung, J.S. Biological characterization of an international collection of Citrus tristeza virus (CTV) isolates. In Proceedings of the 16th Conference of the International Organization of Citrus Virologist, Monterrey, Mexico, 7–12 November 2004; Hilf, M.E., Duran-Vila, N., Rocha-Peña, M.A., Eds.; International Organization of Citrus Virologist (IOCV): Riverside, CA, USA, 2005; pp. 75–93. [Google Scholar]
- Roy, A.; Ananthakrishnan, G.; Hartung, J.S.; Brlansky, R.H. Development and application of a multiplex reverse-transcription polymerase chain reaction assay for screening a global collection of Citrus tristeza virus isolates. Phytopathology 2010, 100, 1077–1088. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruiz, S.; Moreno, P.; Guerri, J.; Ambrós, S. Discrimination between mild and severe Citrus tristeza virus isolates with a rapid and highly specific real-time reverse transcription-polymerase chain reaction method using TaqMan LNA probes. Phytopathology 2009, 99, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Yokomi, R.K.; Saponari, M.; Sieburth, P.J. Rapid differentiation and identification of potential severe strains of Citrus tristeza virus by real-time reverse transcription-polymerase chain reaction assays. Phytopathology 2010, 100, 319–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatineni, S.; Afunian, M.R.; Hilf, M.E.; Gowda, S.; Dawson, W.O.; Garnsey, S.M. Molecular characterization of Citrus tatter leaf virus historically associated with meyer lemon trees: Complete genome sequence and development of biologically active in vitro transcripts. Phytopathology 2008, 99, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Papayiannis, L.C. Diagnostic real-time RT-PCR for the simultaneous detection of Citrus exocortis viroid and hop stunt viroid. J. Virol. Methods 2014, 96, 93–99. [Google Scholar] [CrossRef]
- Rizza, S.; Nobile, G.; Tessitori, M.; Catara, A.; Conte, E. Real time RT-PCR assay for quantitative detection of citrus viroid III in plant tissues. Plant Pathol. 2009, 58, 181–185. [Google Scholar] [CrossRef]
- Bernard, L.; Duran-Vila, N. A novel RT-PCR approach for detection and characterization of citrus viroids. Mol. Cell. Probes 2006, 20, 105–113. [Google Scholar] [CrossRef]
- Serra, P.; Eiras, M.; Bani-Hashemian, S.M.; Murcia, N.; Kitajima, E.W.; Daros, J.A.; Flores, R.; Duran-Vila, N. Citrus viroid V: Occurrence, host range, diagnosis, and identification of new variants. Phytopathology 2008, 98, 1199–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Garcìa, A.B.; Bester, R.; Olmos, A.; Maree, H.J. Bioinformatic tools and genome analysis of Citrus tristeza virus. In Citrus Tristeza Virus: Methods in Molecular Biology; Catara, A., Bar-Joseph, M., Licciardello, G., Eds.; Humana: New York, NY, USA, 2019; Volume 2015, pp. 163–178. [Google Scholar]
- Duran-Vila, N.; Semancik, J. Citrus viroids. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, Australia, 2003; pp. 178–194. [Google Scholar]
- Duran-Vila, N. Citrus exocortis viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 169–179. [Google Scholar]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Islas, C.; Rowhani, A. Comparison of next-generation sequencing versus biological indexing for the optimal detection of viral pathogens of grapevines. Phytopathology 2015, 105, 758–763. [Google Scholar] [CrossRef] [Green Version]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Mumford, R.; Jackeviciene, E.; Navalin-Skiene, M.; Samuitiene, M.; Boonham, N. Next generation sequencing and metagenomic analysis: A universal diagnostic tool in plant virology. Mol. Plant Pathol. 2009, 10, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Massart, S.; Olmos, A.; Jijakli, H.; Candresse, T. Current impact and future directions of high throughput sequencing in plant virus diagnostics. Virus Res. 2014, 188, 90–96. [Google Scholar] [CrossRef]
- Licciardello, G.; Xiao, C.; Russo, M.; Dai, S.M.; Daden, M.; Deng, Z.N.; Catara, A.F. Genetic structure of Citrus tristeza virus in Hunan province (P.R. China). Acta Hortic. 2015, 1065, 781–790. [Google Scholar] [CrossRef]
- Santala, J.; Valkonen, J.P.T. Sensitivity of small RNA-based detection of plant viruses. Front. Microbiol. 2018, 9, 939. [Google Scholar] [CrossRef] [Green Version]
- Bukowska-O’sko, I.; Perlejewski, K.; Nakamura, S.; Motooka, D.; Stokowy, T.; Kosinska, J.; Popiel, M.; Płoski, R.; Horban, A.; Lipowski, D. Sensitivity of next-generation sequencing metagenomic analysis for detection of RNA and DNA viruses in cerebrospinal fluid: The confounding effect of background contamination. In Respiratory Treatment and Prevention; Pokorski, M., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 53–62. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Genus | Family | Host | Country | Reference |
|---|---|---|---|---|---|
| Citrus tristeza virus (CTV) | Closterovirus | Closteroviridae | Sweet orange, Mexican lime, sour orange | Spain | [15] |
| C. sinensis, C. macrophylla | Florida | [16] | |||
| C. sinensis, C. reticulata | California | [17] | |||
| Grapefruit | South Africa | [18] | |||
| Citrus sudden death-associated virus (CSDaV) | Marafivirus | Tymoviridae | Sweet orange | Brazil | [19] |
| Citrus leaf blotch virus (CLBV) | Citrivirus | Betaflexiviridae | Sweet Cherry | China | [20] |
| Citrus psorosis ophiovirus (CPV) | Ophiovirus | Aspiviridae | Sweet orange | China | [unpublished] |
| Citrus tatter leaf virus (CTLV) | Capillovirus | Betaflexiviridae | Apple | China | [21] |
| Citrus yellow vein clearing virus (CYVCV) | Mandarivirus | Alphaflexiviridae | Lemon | Turkey | [22] |
| Citrus chlorotic dwarf-associated virus (CCDaV) | Geminivirus | Geminiviridae | Lemon | Turkey | [23] |
| Citrus vein enation virus (CVEV) | Enamovirus | Luteoviridae | Etrog citron | Spain | [24] |
| Citrus leprosis virus-N (CiLV-N) | Dichoravirus | Rhabdoviridae | Mexico | [25] | |
| Citrus leprosis virus-C2 (CiLV-2) | Cilevirus | Unassigned | Colombia | [26] | |
| Citrus concave gum-associated virus (CCGaV) | Phlebovirus | Phenuiviridae | Tarocco sweet orange | Italy | [27] |
| Citrus jingmen-like virus (CJLV) | Flavivirus | Flaviviridae | Valencia sweet orange | Brazil | [19] |
| Citrus virga-like virus (CVLV) | Virgavirus | Virgaviridae | Valencia sweet orange | Brazil | [19] |
| Satsuma dwarf virus (SDV) | Sadwavirus | Secoviridae | Sour orange and tangor | China | [28] |
| Virus Isolate | Field Plant | Origin | Age | Tissue Source |
|---|---|---|---|---|
| HU-PSTS | Sweet orange/P. trifoliata | Hunan, China | ≈30 years | Bark, sour orange |
| P3R1 | Sweet orange/sour orange | Sicily, Italy | ≈22 years | Bark, sweet orange |
| P3R3 | Sweet orange/sour orange | Sicily, Italy | ≈22 years | Bark, sweet orange |
| Mac39 | Alemow seedling | Sicily, Italy | ≈2 years | Bark, sour orange |
| Mac101 | Alemow seedling | Sicily, Italy | ≈2 years | Bark, sour orange |
| Mac25 | Alemow seedling | Sicily, Italy | ≈2 years | Bark, sour orange |
| Virus | Genotype | Accession | Isolate | Origin |
|---|---|---|---|---|
| Citrus tristeza virus | VT | KC748392 | SG29 | Italy |
| T3 | EU857538 | NZ-M16 | New Zealand | |
| T30 | KC748391 | Bau282 | Italy | |
| T68 | EU076703 | B165 | India | |
| RB | FJ525435 | NZRB-M17 | New Zealand | |
| T36 | EU937521 | T36 FS2-2 | Florida | |
| Citrus yellow vein clearing | JX040635 | Y1 | Turkey | |
| NC_026592 | CQ | China | ||
| KP313241 | PK | Pakistan | ||
| Citrus tatter leaf virus | KC588948 | MTH | China | |
| JX416228 | PK | Taiwan | ||
| Citrus sudden death-associated virus | AY884005 | Brazil | ||
| Citrus leprosis virus C | RNA1 | NC_008169 | Brazil | |
| RNA2 | NC_008170 | Brazil | ||
| Citrus yellow mosaic virus | DQ875213 | India | ||
| Viroids | ||||
| Citrus exocortis viroid | AB054599 | MA | Japan | |
| Hop stunt viroid | AB211242 | EX1 | Japan | |
| Citrus bent leaf viroid | NC001651 | CBLVd-225 | Israel | |
| Citrus dwarfing viroid isolate | EU934031 | E822 | California | |
| Citrus viroid V | NC010165 | Spain | ||
| Citrus bark cracking viroid | NC003539 | Israel |
| Read Count (RC) | ||||||
|---|---|---|---|---|---|---|
| Viroid | Mac39 | Mac101 | Mac25 | P3R1 | P3R3 | HU-PSTS |
| Citrus exocortis viroid | 29 | 208 | 459 | 57,780 | 23,522 | 602 |
| Hop stunt viroid | 42 | 498 | 738 | 15,400 | 5146 | 21,939 |
| Citrus bent leaf viroid | 74 | 237 | 472 | 6859 | 3600 | 829 |
| Citrus dwarfing viroid | 178 | 118 | 228 | 32,317 | 17,250 | 32,393 |
| Citrus viroid V | 191 | 309 | 132 | 7497 | 5058 | 5374 |
| Citrus bark cracking viroid | 16 | 115 | 227 | 11,036 | 4932 | 199 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Licciardello, G.; Ferraro, R.; Scuderi, G.; Russo, M.; Catara, A.F. A Simulation of the Use of High Throughput Sequencing as Pre-Screening Assay to Enhance the Surveillance of Citrus Viruses and Viroids in the EPPO Region. Agriculture 2021, 11, 400. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11050400
Licciardello G, Ferraro R, Scuderi G, Russo M, Catara AF. A Simulation of the Use of High Throughput Sequencing as Pre-Screening Assay to Enhance the Surveillance of Citrus Viruses and Viroids in the EPPO Region. Agriculture. 2021; 11(5):400. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11050400
Chicago/Turabian StyleLicciardello, Grazia, Rosario Ferraro, Giuseppe Scuderi, Marcella Russo, and Antonino F. Catara. 2021. "A Simulation of the Use of High Throughput Sequencing as Pre-Screening Assay to Enhance the Surveillance of Citrus Viruses and Viroids in the EPPO Region" Agriculture 11, no. 5: 400. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11050400