Biostimulatory Action of Arbuscular Mycorrhizal Fungi Enhances Productivity, Functional and Sensory Quality in ‘Piennolo del Vesuvio’ Cherry Tomato Landraces

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
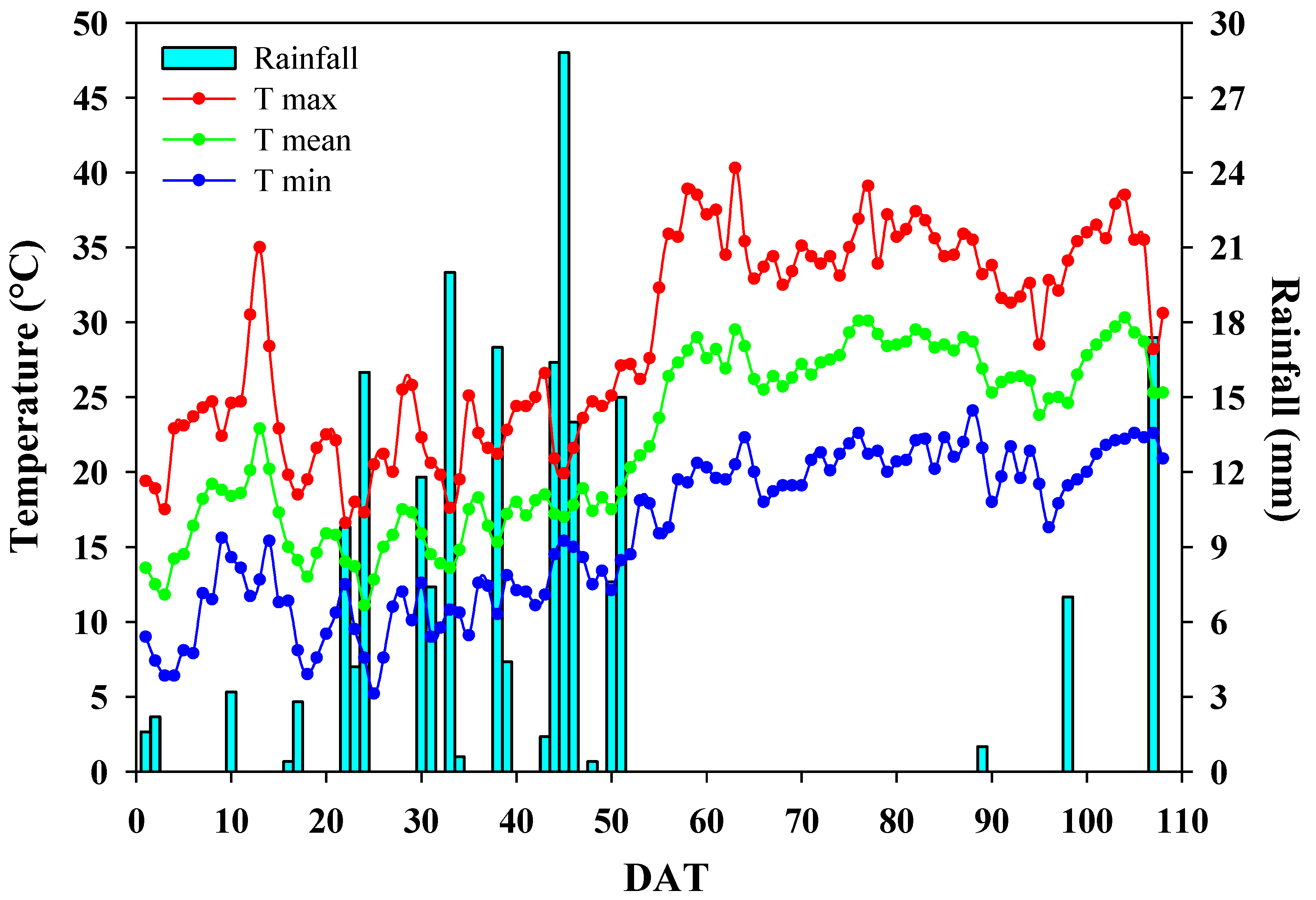
2.1. Growth Conditions, Tomato Landraces, Experimental Design and Cultural Practices
2.2. Yield Measurements, Fruit Quality Sampling and Arbuscular Mycorrhizal Fungi Root Colonization
2.3. Total Soluble Solids and Fruit Dry Matter Content
2.4. Macro and Micro Mineral Content Analysis
2.5. Antioxidant Activity and Bioactive Compounds Analysis
2.6. Starch and Sucrose Analysis
2.7. Amino Acids Analysis
2.8. Statistics
3. Results
3.1. Fungal Concentrations in the Rhizo-Soil and Yield Responses of Two Tomato Landraces as Affected by AMF Inoculation
3.2. Mineral Profile of Two Tomato Landraces as Affected by AMF Inoculation
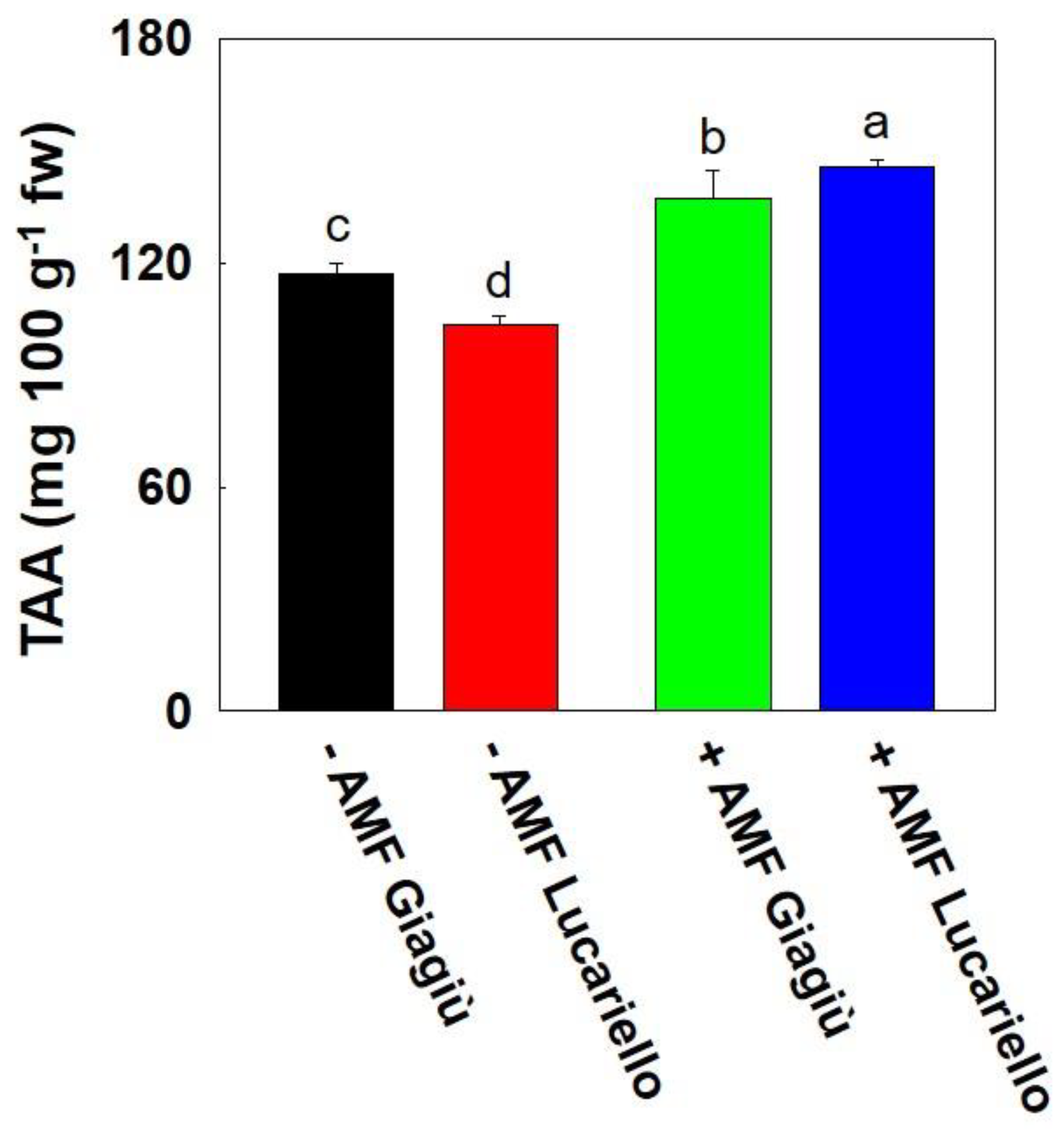
3.3. Fruit Soluble Solids, Dry Matter, Sucrose, Starch, Antioxidant Activity and Bioactive Compounds of Two Tomato Landraces as Affected by AMF Inoculation
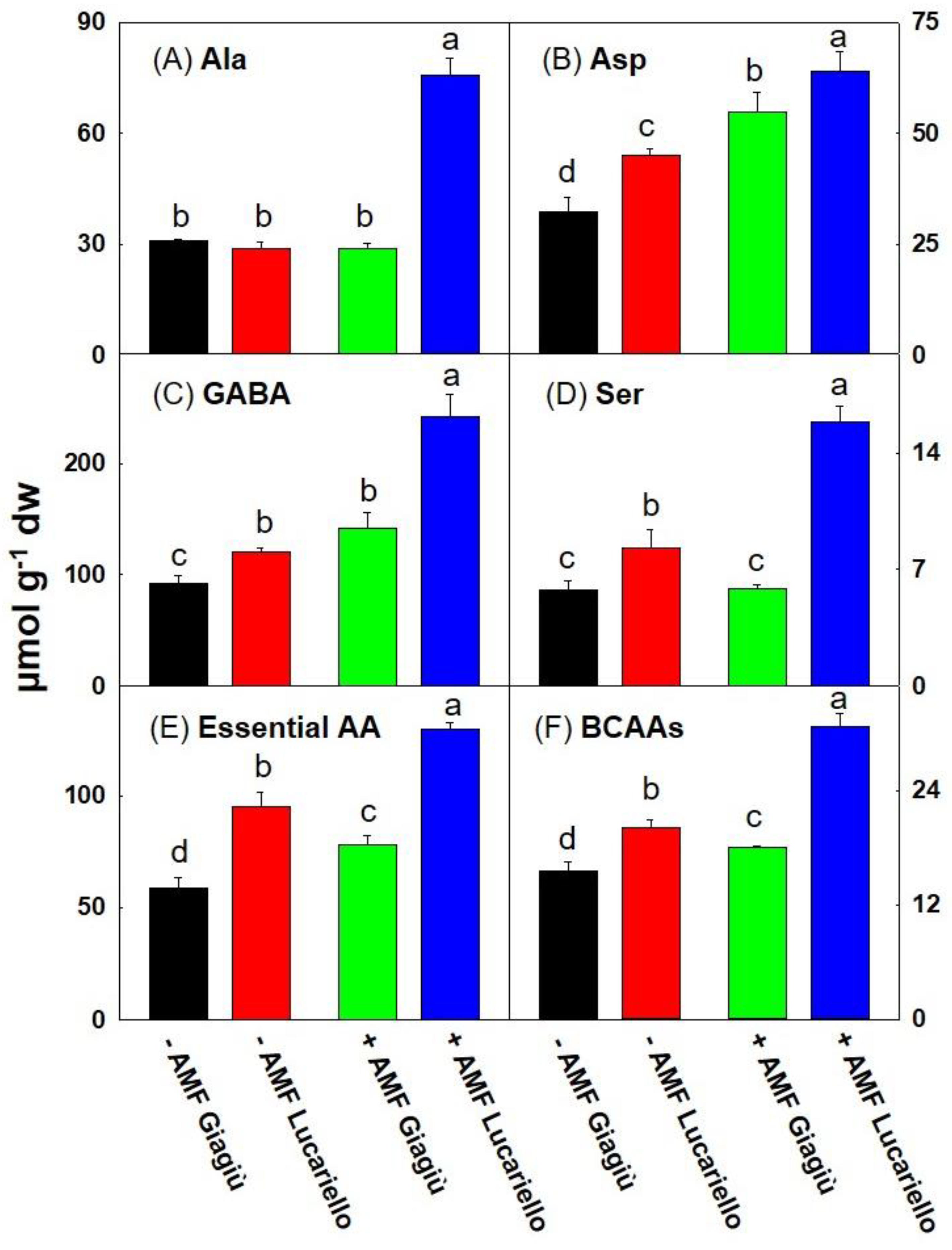
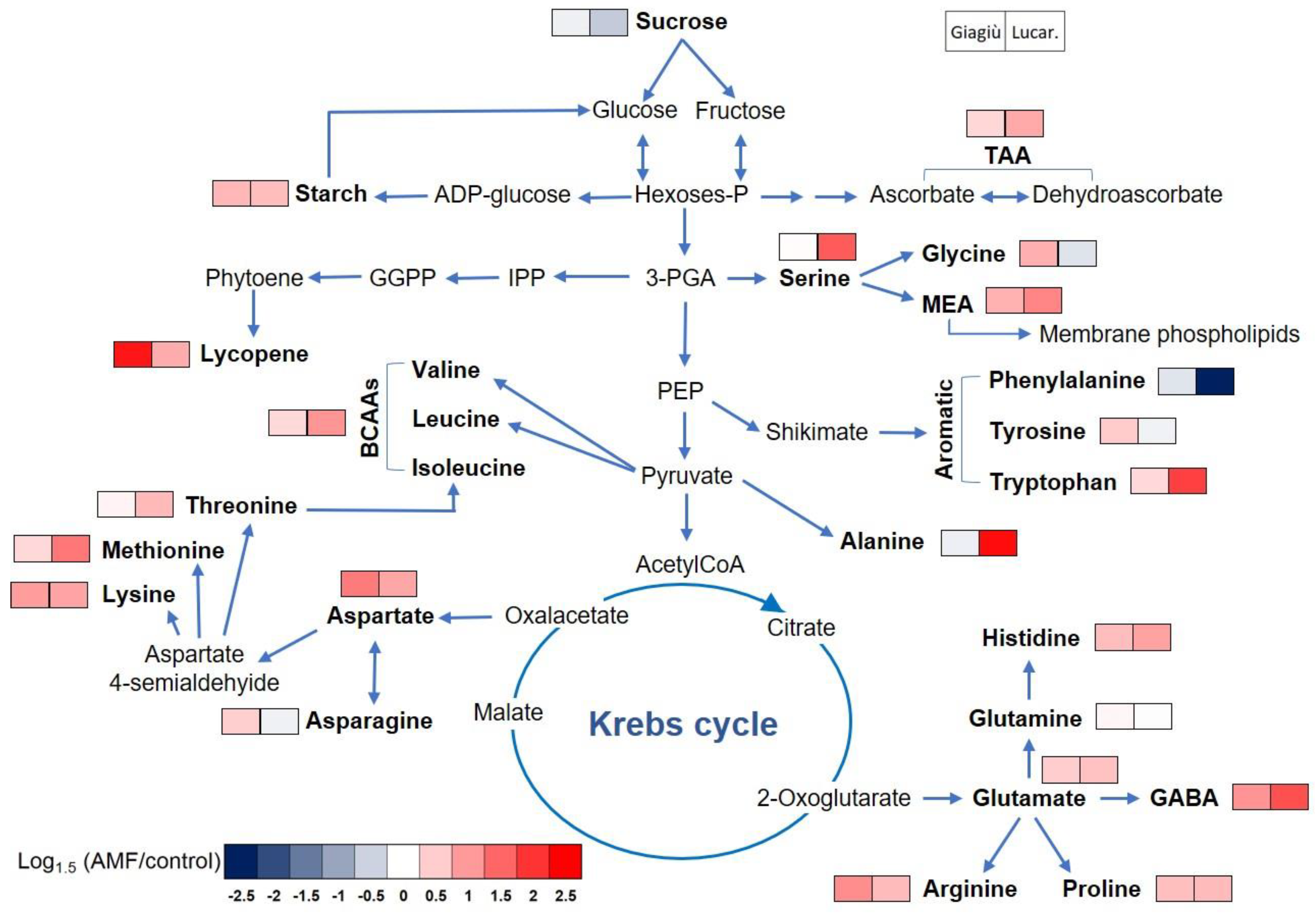
3.4. Amino Acids Profiling of Two Tomato Landraces as Affected by AMF Inoculation
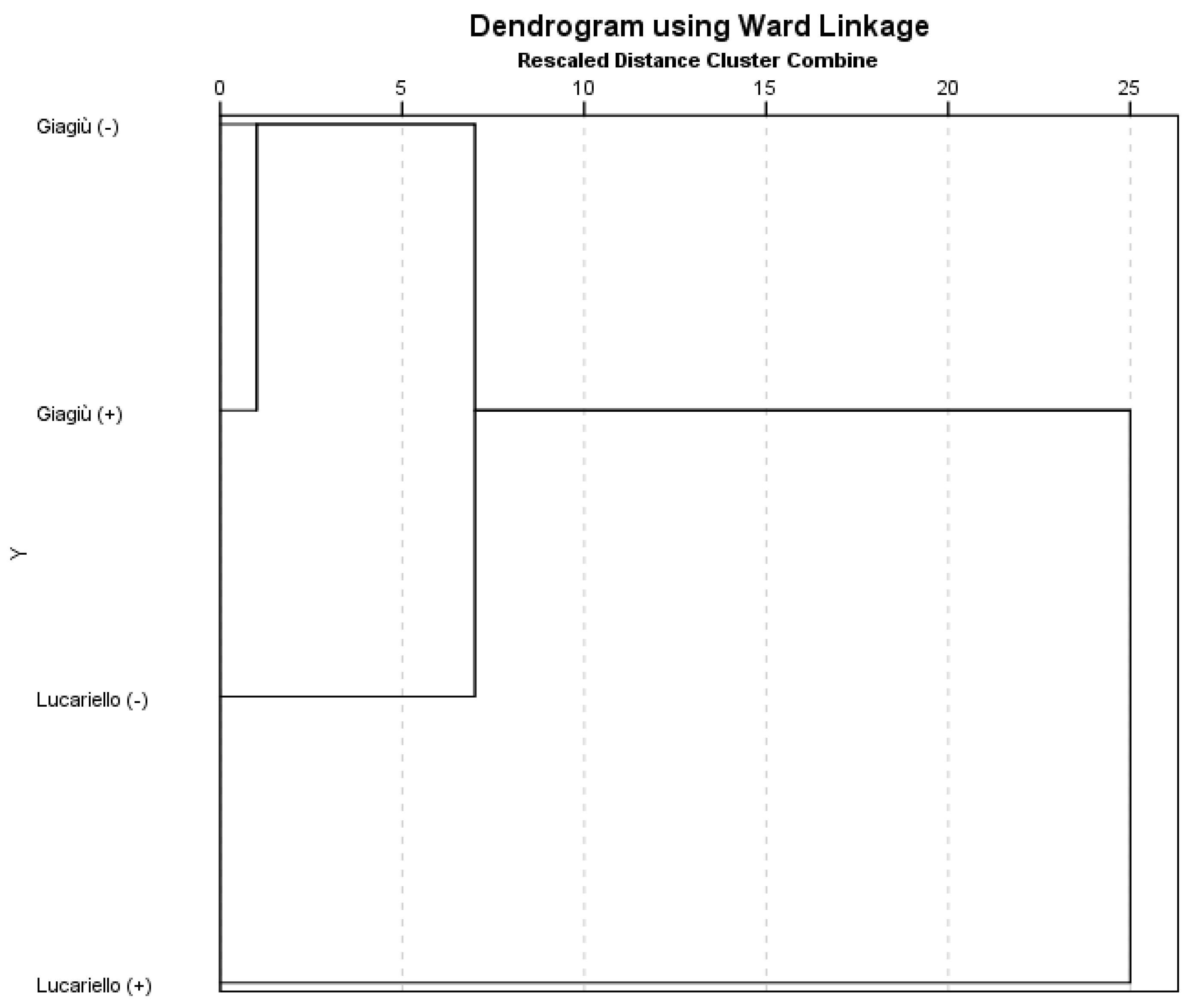
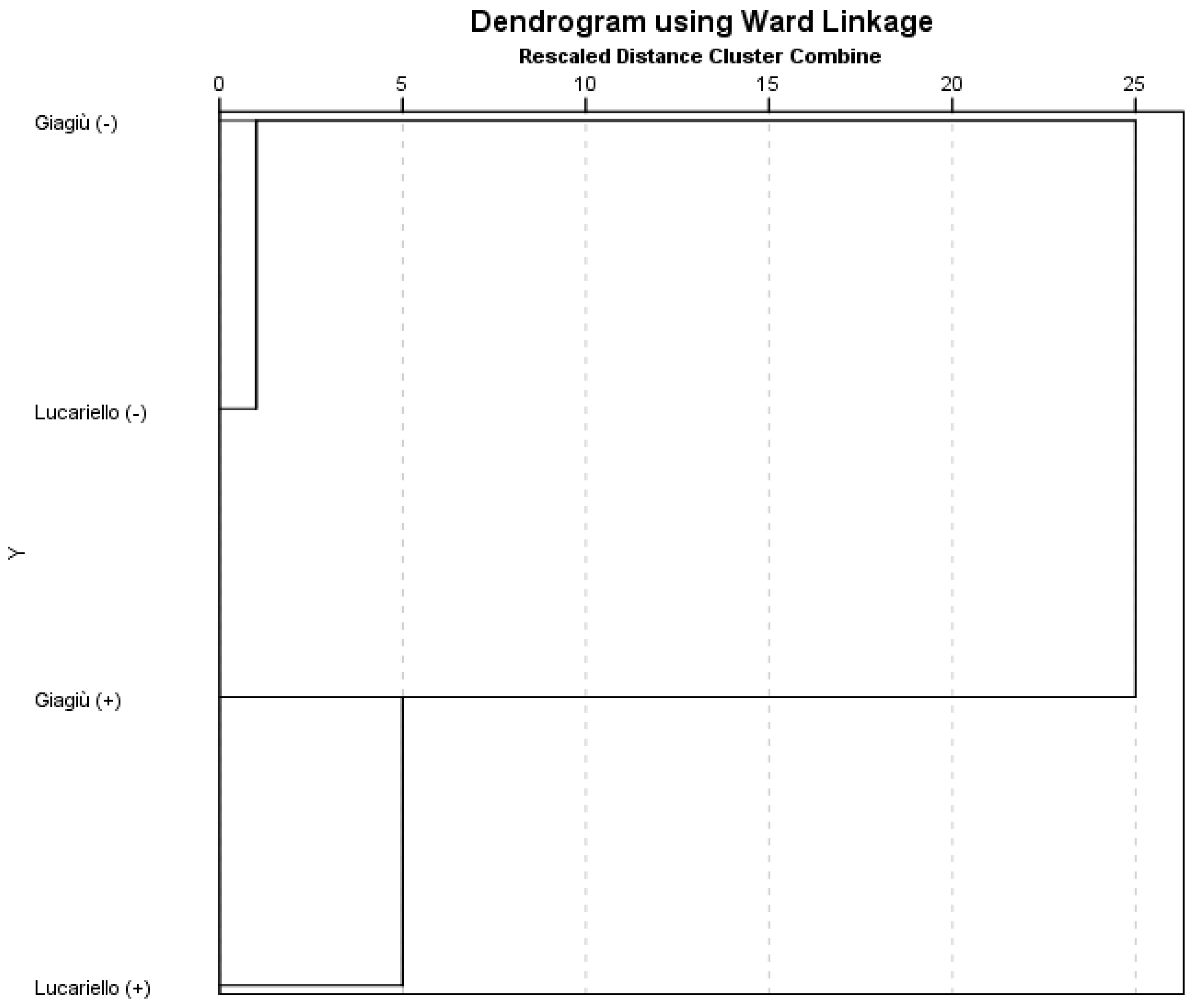
3.5. Hierarchical Cluster Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chakravorty, U.; Fisher, D.K.; Umetsu, C. Environmental effects of intensification of agriculture: Livestock production and regulation. Environ. Econ. Policy Stud. 2007, 8, 315–336. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Dell’Aversana, E.; Ruggiero, A.; Cirillo, V.; Gibon, Y.; Woodrow, P.; Maggio, A.; Carillo, P. Omeprazole Treatment Enhances Nitrogen Use Efficiency Through Increased Nitrogen Uptake and Assimilation in Corn. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quammen, D. Spillover: Animal Infections and the Next Human Pandemic; W.W. Norton & Co.: New York, NY, USA, 2012. [Google Scholar]
- Gosalvez, E. How Habitat Destruction Enables the Spread of Diseases Like COVID-19. Available online: https://cnr.ncsu.edu/news/2020/04/habitat-destruction-covid19/ (accessed on 22 April 2020).
- Pretty, J.; Bharucha, Z.P. Sustainable intensification in agricultural systems. Ann. Bot. 2014, 114, 1571–1596. [Google Scholar] [CrossRef] [PubMed]
- Avio, L.; Turrini, A.; Giovannetti, M.; Sbrana, C. Designing the Ideotype Mycorrhizal Symbionts for the Production of Healthy Food. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; Pascale, S.D.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Castellanos-Morales, V.; Villegas, J.; Wendelin, S.; Vierheilig, H.; Eder, R.; Cárdenas-Navarro, R. Root colonisation by the arbuscular mycorrhizal fungus Glomus intraradices alters the quality of strawberry fruits (Fragaria × ananassa Duch.) at different nitrogen levels. J. Sci. Food Agric. 2010, 90, 1774–1782. [Google Scholar] [CrossRef]
- Mollavali, M.; Bolandnazar, S.A.; Schwarz, D.; Rohn, S.; Riehle, P.; Zaare Nahandi, F. Flavonol Glucoside and Antioxidant Enzyme Biosynthesis Affected by Mycorrhizal Fungi in Various Cultivars of Onion (Allium cepa L.). J. Agric. Food Chem. 2016, 64, 71–77. [Google Scholar] [CrossRef]
- Ahmad, H.; Hayat, S.; Ali, M.; Liu, T.; Cheng, Z. The combination of arbuscular mycorrhizal fungi inoculation (Glomus versiforme) and 28-homobrassinolide spraying intervals improves growth by enhancing photosynthesis, nutrient absorption, and antioxidant system in cucumber (Cucumis sativus L.) under salinity. Ecol. Evol. 2018, 8, 5724–5740. [Google Scholar] [CrossRef] [PubMed]
- Fernández, I.; Merlos, M.; López-Ráez, J.A.; Martínez-Medina, A.; Ferrol, N.; Azcón, C.; Bonfante, P.; Flors, V.; Pozo, M.J. Defense Related Phytohormones Regulation in Arbuscular Mycorrhizal Symbioses Depends on the Partner Genotypes. J. Chem. Ecol. 2014, 40, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula. Agronomy 2020, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; dell’Aversana, E.; D’Amelia, L.; Colla, G.; Caruso, G.; De Pascale, S.; Rouphael, Y. Sensory and functional quality characterization of protected designation of origin ‘Piennolo del Vesuvio’ cherry tomato landraces from Campania-Italy. Food Chem. 2019, 292, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Frusciante, L.; Carli, P.; Ercolano, M.R.; Pernice, R.; Di Matteo, A.; Fogliano, V.; Pellegrini, N. Antioxidant nutritional quality of tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN118. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Volpe, M.G.; Nazzaro, M.; Di Stasio, M.; Siano, F.; Coppola, R.; De Marco, A. Content of micronutrients, mineral and trace elements in some Mediterranean spontaneous edible herbs. Chem. Cent. J. 2015, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef]
- Sadler, G.; Davis, J.; Dezman, D. Rapid Extraction of Lycopene and β-Carotene from Reconstituted Tomato Paste and Pink Grapefruit Homogenates. J. Food Sci. 1990, 55, 1460–1461. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Cacace, D.; De Rosa, M.; De Martino, E.; Cozzolino, C.; Nacca, F.; D’Antonio, R.; Fuggi, A. Process optimisation and physicochemical characterisation of potato powder. Int. J. Food Sci. Technol. 2009, 44, 145–151. [Google Scholar] [CrossRef]
- Woodrow, P.; Ciarmiello, L.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef]
- Newbould, P. The use of nitrogen fertiliser in agriculture. Where do we go practically and ecologically? Plant Soil 1989, 115, 297–311. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant Application with a Tropical Plant Extract Enhances Corchorus olitorius Adaptation to Sub-Optimal Nutrient Regimens by Improving Physiological Parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Colla, G. Role of arbuscular mycorrhizal fungi in alleviating the adverse effects of acidity and aluminium toxicity in zucchini squash. Sci. Hortic. 2015, 188, 97–105. [Google Scholar] [CrossRef]
- Subramanian, K.S.; Santhanakrishnan, P.; Balasubramanian, P. Responses of field grown tomato plants to arbuscular mycorrhizal fungal colonization under varying intensities of drought stress. Sci. Hortic. 2006, 107, 245–253. [Google Scholar] [CrossRef]
- Dasgan, H.; Kusvuran, S.; Ortas, I. Responses of soilless grown tomato plants to arbuscular mycorrhizal fungal (Glomus fasciculatum) colonization in re-cycling and open systems. Afr. J. Biotechnol. 2008, 7, 3606–3613. [Google Scholar]
- Salvioli, A.; Zouari, I.; Chalot, M.; Bonfante, P. The arbuscular mycorrhizal status has an impact on the transcriptome profile and amino acid composition of tomato fruit. BMC Plant Biol. 2012, 12, 44. [Google Scholar] [CrossRef] [Green Version]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Chapter 6—Functions of Macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 135–189. [Google Scholar] [CrossRef]
- Hirschi, K.D. The calcium conundrum. Both versatile nutrient and specific signal. Plant Physiol. 2004, 136, 2438–2442. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Cheng, N.H.; Pittman, J.K.; Yoo, K.S.; Park, J.; Smith, R.H.; Hirschi, K.D. Increased calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol. 2005, 139, 1194–1206. [Google Scholar] [CrossRef] [Green Version]
- de Freitas, S.T.; Padda, M.; Wu, Q.; Park, S.; Mitcham, E.J. Dynamic Alternations in Cellular and Molecular Components during Blossom-End Rot Development in Tomatoes Expressing sCAX1, a Constitutively Active Ca2+/H+ Antiporter from Arabidopsis. Plant Physiol. 2011, 156, 844–855. [Google Scholar] [CrossRef] [Green Version]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit Calcium: Transport and Physiology. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Patni, B.; Shankhdhar, D.; Shankhdhar, S.C. Zinc—An indispensable micronutrient. Physiol. Mol. Biol. Plants 2013, 19, 11–20. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Chapter 7—Function of Nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 191–248. [Google Scholar] [CrossRef]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutts, S. Tomato Fruit Development and Metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazemi, M. Effects of Zn, Fe and their Combination Treatments on the growth and yield of tomato. Bull. Environ. Pharmacol. Life Sci. 2013, 3, 109–114. [Google Scholar]
- Ronaghy, H.A. The role of zinc in human nutrition. World Rev. Nutr. Diet. 1987, 54, 237–254. [Google Scholar] [CrossRef]
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and its importance for human health: An integrative review. J. Res. Med. Sci. 2013, 18, 144–157. [Google Scholar]
- Brown, K.; Wuehler, S.; Peerson, J. The Importance of Zinc in Human Nutrition and Estimation of the Global Prevalence of Zinc Deficiency. Food Nutr. Bull. 2001, 22. [Google Scholar] [CrossRef] [Green Version]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bücking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, D.R.; Hausmann, N.T.; Barrios-Masias, F.H.; Jackson, L.E.; Schachtman, D.P. Transcriptomic and metabolic responses of mycorrhizal roots to nitrogen patches under field conditions. Plant Soil 2012, 350, 145–162. [Google Scholar] [CrossRef]
- Gu, M.; Xu, K.; Chen, A.; Zhu, Y.; Tang, G.; Xu, G. Expression analysis suggests potential roles of microRNAs for phosphate and arbuscular mycorrhizal signaling in Solanum lycopersicum. Physiol. Plant. 2010, 138, 226–237. [Google Scholar] [CrossRef]
- Chaudhari, N.; Pereira, E.; Roper, S.D. Taste receptors for umami: The case for multiple receptors. Am. J. Clin. Nutr. 2009, 90, 738S–742S. [Google Scholar] [CrossRef] [Green Version]
- Galili, G. The aspartate-family pathway of plants: Linking production of essential amino acids with energy and stress regulation. Plant Signal. Behav. 2011, 6, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Cao, Q.; Sun, L.; Yang, X.; Yang, W.; Zhang, H. Stomatal Conductance and Morphology of Arbuscular Mycorrhizal Wheat Plants Response to Elevated CO(2) and NaCl Stress. Front. Plant Sci. 2018, 9, 1363. [Google Scholar] [CrossRef] [PubMed]
- Gout, E.; Bligny, R.; Douce, R. Regulation of intracellular pH values in higher plant cells. Carbon-13 and phosphorus-31 nuclear magnetic resonance studies. J. Biol. Chem. 1992, 267, 13903–13909. [Google Scholar] [PubMed]
- Limami, A.M.; Glévarec, G.; Ricoult, C.; Cliquet, J.B.; Planchet, E. Concerted modulation of alanine and glutamate metabolism in young Medicago truncatula seedlings under hypoxic stress. J. Exp. Bot. 2008, 59, 2325–2335. [Google Scholar] [CrossRef] [Green Version]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P. GABA Shunt in Durum Wheat. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and Temporal Profile of Glycine Betaine Accumulation in Plants Under Abiotic Stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Ruiz, R.; Poirot, E.; Flores-Mosquera, M. GABA, a non-protein amino acid ubiquitous in food matrices. Cogent Food Agric. 2018, 4, 1534323. [Google Scholar] [CrossRef]
- Ngo, D.-H.; Vo, T.S. An Updated Review on Pharmaceutical Properties of Gamma-Aminobutyric Acid. Molecules 2019, 24, 2678. [Google Scholar] [CrossRef] [Green Version]
- Pennathur, S.; Maitra, D.; Byun, J.; Sliskovic, I.; Abdulhamid, I.; Saed, G.M.; Diamond, M.P.; Abu-Soud, H.M. Potent antioxidative activity of lycopene: A potential role in scavenging hypochlorous acid. Free Radic. Biol. Med. 2010, 49, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Sipos, L.; Orbán, C.; Bálint, I.; Csambalik, L.; Divéky-Ertsey, A.; Gere, A. Colour parameters as indicators of lycopene and antioxidant activity traits of cherry tomatoes (Solanum lycopersicum L.). Eur. Food Res. Technol. 2017, 243, 1533–1543. [Google Scholar] [CrossRef]
- Thies, F.; Mills, L.M.; Moir, S.; Masson, L.F. Cardiovascular benefits of lycopene: Fantasy or reality? Proc. Nutr. Soc. 2016, 76, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.V.; Ray, M.R.; Rao, L.G. Lycopene. Adv. Food Nutr. Res. 2006, 51, 99–164. [Google Scholar] [CrossRef]
- Lucarini, M.; Lanzi, S.; D’Evoli, L.; Aguizzi, A.; Lombardi-Boccia, G. Intake of vitamin A and carotenoids from the Italian population—Results of an Italian total diet study. Int. J. Vitam. Nutr. Res. 2006, 76, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Przybylska, S. Lycopene—A bioactive carotenoid offering multiple health benefits: A review. Int. J. Food Sci. Technol. 2020, 55, 11–32. [Google Scholar] [CrossRef]
- Martínez-Valverde, I.; Periago, M.J.; Provan, G.; Chesson, A. Phenolic compounds, lycopene and antioxidant activity in commercial varieties of tomato (Lycopersicum esculentum). J. Sci. Food Agric. 2002, 82, 323–330. [Google Scholar] [CrossRef]
- Mirto, A.; Iannuzzi, F.; Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Fuggi, A. Metabolic characterization and antioxidant activity in sweet cherry (Prunus avium L.) Campania accessions: Metabolic characterization of sweet cherry accessions. Food Chem. 2018, 240, 559–566. [Google Scholar] [CrossRef]
- Ortiz-Espín, A.; Sánchez-Guerrero, A.; Sevilla, F.; Jiménez, A. The Role of Ascorbate in Plant Growth and Development. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M.A., Munné-Bosch, S., Burritt, D.J., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 25–45. [Google Scholar] [CrossRef]
- Gulati, K.; Anand, R.; Ray, A. Chapter 16—Nutraceuticals as Adaptogens: Their Role in Health and Disease. In Nutraceuticals; Gupta, R.C., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 193–205. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Yield | Number of Fruits | Fruit Mean Weight |
|---|---|---|---|
| (g m−2) | (no. m−2) | (g fruit−1) | |
| Mycorrhiza (M) | ** | *** | * |
| Landraces (L) | ** | ns | ** |
| M × L | ns | ns | ns |
| Main effect—AMF | |||
| −AMF | 728 | 67.97 | 10.66 |
| +AMF | 953 | 94.33 | 10.16 |
| Main effect—Landrace | |||
| ‘Giagiù’ | 730 | 81.75 | 8.96 |
| ‘Lucariello’ | 951 | 80.55 | 11.86 |
| Source of Variation | P | K | Ca | Mg | Na | Fe | Cu | Mn | Zn |
|---|---|---|---|---|---|---|---|---|---|
| (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (mg g−1 dw) | (μg g−1 dw) | (μg g−1 dw) | (μg g−1 dw) | (μg g−1 dw) | |
| Mycorrhiza (M) | * | ns | ** | * | *** | ns | * | ns | ** |
| Landraces (L) | ns | ns | ** | ns | *** | ns | * | ns | * |
| M × L | ns | ns | ** | ns | ** | ns | ns | ns | ns |
| Main effect—AMF | |||||||||
| −AMF | 2.83 | 31.86 | 1.10 | 1.09 | 0.02 | 41.48 | 12.58 | 7.55 | 19.95 |
| +AMF | 3.53 | 32.32 | 1.41 | 1.13 | 0.01 | 44.66 | 13.81 | 7.88 | 26.78 |
| Main effect—Landrace | |||||||||
| ‘Giagiù’ | 3.24 | 32.40 | 1.43 | 1.13 | 0.03 | 42.77 | 12.52 | 7.90 | 26.09 |
| ‘Lucariello’ | 3.12 | 31.78 | 1.08 | 1.09 | 0.01 | 43.37 | 13.87 | 7.53 | 20.64 |
| Source of Variation | TSS | DM | LAA | HAA | Starch | TAA | Lycopene | Sucrose |
|---|---|---|---|---|---|---|---|---|
| (°Brix) | (%) | (mmol Trolox 100 g−1 dw) | (mmol AA 100 g−1 dw) | (µmol g−1 dw) | (mg 100 g−1 fw) | (mg 100 g−1 dw) | (µmol g−1 dw) | |
| Mycorrhiza (M) | * | ** | * | * | ** | *** | ** | * |
| Landraces (L) | ns | ns | ** | * | ns | ns | *** | ** |
| M × L | ns | ns | ns | ns | ns | ** | ns | * |
| Main effect—AMF | ||||||||
| −AMF | 8.16 | 9.83 | 17.24 | 10.43 | 15.60 | 110.2 | 26.23 | 13.52 |
| +AMF | 8.95 | 10.78 | 19.31 | 11.33 | 20.50 | 141.5 | 38.48 | 11.56 |
| Main effect—Landrace | ||||||||
| ‘Giagiù’ | 8.67 | 10.37 | 19.75 | 10.67 | 18.22 | 127.0 | 5.89 | 11.59 |
| ‘Lucariello’ | 8.45 | 10.23 | 16.79 | 11.08 | 17.88 | 124.7 | 58.82 | 13.49 |
| Amino Acids (µmol g−1 dw) | Main Effect Mycorrhiza | Main Effect Landraces | Significance | ||||
|---|---|---|---|---|---|---|---|
| −AMF | +AMF | ‘Giagiù’ | ‘Lucariello’ | Mycorrhiza (M) | Landraces (L) | M × L | |
| Ala | 29.86 | 52.22 | 29.80 | 52.27 | *** | *** | *** |
| Arg | 26.10 | 36.82 | 24.86 | 38.06 | ** | *** | ns |
| Asn | 89.54 | 91.37 | 58.75 | 122.16 | ns | ** | ns |
| Asp | 38.61 | 59.36 | 43.50 | 54.47 | *** | * | ns |
| GABA | 106.4 | 191.4 | 116.8 | 181.0 | *** | ** | * |
| Gln | 117.6 | 118.7 | 87.08 | 149.3 | ns | ** | ns |
| Glu | 169.9 | 213.4 | 157.7 | 225.6 | ns | * | ns |
| Gly | 12.00 | 12.66 | 9.93 | 14.73 | ns | * | ns |
| His | 8.06 | 11.15 | 8.04 | 11.18 | ** | * | ns |
| Ile | 7.32 | 10.27 | 7.12 | 10.47 | ** | *** | * |
| Leu | 5.91 | 8.33 | 5.39 | 8.84 | *** | *** | ** |
| Lys | 11.82 | 16.86 | 9.19 | 19.49 | * | ** | ns |
| MEA | 3.89 | 5.82 | 4.48 | 5.23 | * | ns | ns |
| Met | 1.06 | 1.58 | 0.78 | 1.85 | ** | *** | ** |
| Orn | 2.38 | 3.02 | 2.02 | 3.39 | * | ** | ns |
| Phe | 6.02 | 4.73 | 3.62 | 7.13 | * | ** | ns |
| Pro | 13.32 | 17.15 | 15.18 | 15.30 | * | ns | ns |
| Ser | 7.01 | 10.85 | 5.81 | 12.04 | *** | *** | ** |
| Thr | 3.42 | 4.11 | 2.74 | 4.78 | ns | * | ns |
| Trp | 2.45 | 4.11 | 2.22 | 4.33 | ** | ** | ** |
| Tyr | 4.79 | 4.93 | 3.18 | 6.55 | ns | * | ns |
| Val | 4.61 | 5.79 | 4.28 | 6.11 | ** | ** | * |
| Essential AA | 76.77 | 103.7 | 68.26 | 112.3 | *** | *** | ns |
| BCAAs | 17.84 | 24.38 | 16.80 | 25.42 | *** | *** | ** |
| Total AA | 672.1 | 884.7 | 602.53 | 954.3 | ** | *** | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carillo, P.; Kyratzis, A.; Kyriacou, M.C.; Dell’Aversana, E.; Fusco, G.M.; Corrado, G.; Rouphael, Y. Biostimulatory Action of Arbuscular Mycorrhizal Fungi Enhances Productivity, Functional and Sensory Quality in ‘Piennolo del Vesuvio’ Cherry Tomato Landraces. Agronomy 2020, 10, 911. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060911
Carillo P, Kyratzis A, Kyriacou MC, Dell’Aversana E, Fusco GM, Corrado G, Rouphael Y. Biostimulatory Action of Arbuscular Mycorrhizal Fungi Enhances Productivity, Functional and Sensory Quality in ‘Piennolo del Vesuvio’ Cherry Tomato Landraces. Agronomy. 2020; 10(6):911. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060911
Chicago/Turabian StyleCarillo, Petronia, Angelos Kyratzis, Marios C. Kyriacou, Emilia Dell’Aversana, Giovanna Marta Fusco, Giandomenico Corrado, and Youssef Rouphael. 2020. "Biostimulatory Action of Arbuscular Mycorrhizal Fungi Enhances Productivity, Functional and Sensory Quality in ‘Piennolo del Vesuvio’ Cherry Tomato Landraces" Agronomy 10, no. 6: 911. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060911