Achievements and Challenges towards a Sustainable Conservation and Use of ‘Galega vulgar’ Olea europaea Variety


1
Instituto de Tecnologia Química e Biológica António Xavier, Universidade Nova de Lisboa, Avenida da República, Estação Agronómica Nacional, 2780-157 Oeiras, Portugal
2
Association BLC3—Technology and Innovation Campus, Centre Bio R&D Unit, Rua Nossa Senhora da Conceição, 2, Lagares, 3405-155 Oliveira do Hospital, Portugal
*
Author to whom correspondence should be addressed.
Agronomy 2020, 10(10), 1467; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101467
Submission received: 31 August 2020
/
Revised: 22 September 2020
/
Accepted: 23 September 2020
/
Published: 25 September 2020
(This article belongs to the Special Issue Plant Genetic Resources Conservation and Utilization)
Abstract
:The cultivated Olea europaea L., or olive tree, is an ancient crop extremely relevant for the bioeconomy of Mediterranean countries, especially for Portugal. With orchard modernization, Portugal has the potential to become the third-largest producer of olive oil over the next decade. In this country, the main national variety is ‘Galega vulgar,’ characterized by an excellent olive oil quality, rusticity and tolerance to drought. Nevertheless, its production has suffered a reduction due to replacement by foreign varieties. The present narrative review offers an overall perspective of present gaps and challenges to the conservation and use of ‘Galega vulgar.’ Existing information about traditional and innovative olive production systems, and the importance of life cycle thinking approaches for a sustainable olive chain were synthesized, focusing particularly in the last 10 years. Furthermore, the olive molecular characterization advances and their breeding importance were also evaluated considering their application to this Portuguese variety. To ensure an efficient and sustainable exploitation of the ‘Galega vulgar,’ we propose that efforts should concentrate on the characterization of the existing variability and development of genotype to phenotype prediction tools, integrating detailed molecular marker genotypic and environmental characterization, to support better informed conservation and breeding decisions in a multi-environment context.
1. Introduction
Olive (Olea europaea L., subsp. europaea, var. europaea) is a multifunctional long-living Mediterranean tree species with an extraordinarily ancient history and tradition. This crop holds an important role both for table olive and oil production and for shaping and protecting the regional landscape. Moreover, it assumes an enormous relevance on human nutrition and rural lifestyle [1]. The olive tree genetic breeding started when human populations domesticated wild olives, selecting the best phenotypes. This selection continued across time, throughout the Mediterranean Basin, leading to the present complex genetic relationships existing among varieties [2,3].
Currently, O. europaea is the most cultivated temperate fruit crop worldwide and is characterized by a vast heritage of clonally propagated traditional varieties [4]. Important socioeconomic changes in several Mediterranean countries led to significant technological improvement in olive cultivation, which increased the risk of genetic erosion on the traditional olive germplasm. This genetic erosion phenomenon is also occurring in Portugal, especially affecting the main national variety ‘Galega vulgar,’ which has been replaced despite its excellent olive oil qualities. Since high-yielding and low-vigor modern varieties, more suitable for mechanically harvested plantations, are preferred to the traditional varieties, the characterization and conservation of existing traditional varieties diversity is essential to ensure their sustainable use in the future [5].
In this narrative review we searched international data bases for English-language articles, and national state technical reports and University dissertations (in Portuguese). Keywords used covered the following topics: Economic and social impact of Olea europaea; Olive Production Systems, LCA, Genetic Diversity, Domestication and origin, Olive germplasm conservation, Conventional and precision breeding techniques in olive, Marker assisted selection, GxE, Genotype-to-phenotype (G2P) models. Search was focused mainly on the last 10 years (2010–2020), with additional older studies scanned from the obtained reference list, giving a special emphasis to ‘Galega vulgar’ variety.
Thus, the present narrative review provides an overview of the millenary culture of O. europaea, the changes that olive production systems have been confronted with over time and how life cycle thinking approaches might tackle these modifications and contribute to a more sustainable olive chain. Furthermore, the olive molecular characterization efforts that have taken place in different olive varieties and their importance for breeding programs are also reviewed. A special emphasis is given to the ‘Galega’ variety, highlighting the existing gaps and future challenges concerning this variety conservation and use.
2. Olive Tree: A Millenary Crop with a Massive Economic Impact Worldwide
The olive tree crop, together with the vine crop, represent the oldest perennial crops for human food [3]. Across time, olive trees assumed a special, even sacred, symbolic meaning in Mediterranean communities. Beyond the immense importance that olives and olive oil hold in gastronomy, considered today as cornerstones of the healthy Mediterranean diet, the olive tree is used during many spiritual celebrations [6]. More recently, olive has been recognized as a functional food due to its various bioactive constituents, particularly the phenolic compounds (such as oleuropein, the major phenolic constituent of O. europaea), and their pharmacological activities [7]. The olive high content of monounsaturated fatty acids (mostly oleic acid, representing 70% to 80% of the total olive oil) plays an important role in preventing cardiovascular diseases when consumed. It is a stimulant of the biliary tract (improving digestion and intestinal function) and encourages the absorption of fat-soluble vitamins (A, D, K and E), being rich in vitamin E [8]. Because of this dietetic value, health benefits and economic relevance, there is a high market demand and a regular increase in olive trees cultivated areas [9].
Mediterranean countries are the main consumers of olive oil, representing around 63% of world intake, which correspond to 1836.7 thousand tons [10]. However, olive oil consumption has been increasing in countries outside this region. This group is led by the United States, with about 11% of world consumption—Figure 1a. Regarding the total intake in the European Union (EU), Spain is the main consumer followed by Italy that together consume almost a third of the worldwide total (Figure 1b).
Considering the production of olive oil, the main producer is Spain (about 56% of world total and 79.08% of EU production, corresponding to 1789.9 thousand tons), followed by Greece and Italy—Figure 2a,b.
In the case of Portugal, in 2017/2018 and 2018/2019, this country ranked seventh in the world production of olive oil (≅4%), and the fourth largest olive producer in Europe, behind Spain, Italy and Greece [10]. However, with the modernization of Portuguese olive groves, this country has the potential to become the third-largest producer of olive oil in the next 10 years. In fact, the olive oil production in Portugal has increased over the last decade, generating in the last three years a turnover 2.5 times higher (€620 million) than the one recorded between 2010 and 2012 [11].
In first decades of the last century, there was an expansion of the olive orchards and the production of olives and olive oil in Portugal. However, during the 1960s, olive cultivation suffered a drastic decrease due to changes in policies in the olive sector, the increase of production costs and the market introduction of other plant oils [12]. It was only in 1986, when Portugal became an EU member, that the modernization in the olive sector started, with the approval of the National Plan for the olive crop. With funds from EU, Portugal was able to renew the production models and started a technological revolution in its orchards, leading to an increased productivity and better quality of Portuguese olive oil [11]. In fact, the Portuguese olives and olive oil production already exceeds the production obtained in 1950/1960 (around 120 thousand tons of olive oil) [11], recording that in 2017, the total olive oil, olives for oil and table olives production was 134.8, 858.4 and 134.8 thousand tons, respectively (Figure 3) [10,13].
In Portugal, there are 361.177 thousand hectares (ha) of olive groves, with around 97.6% of them used for olive oil production. In this way, table olives production occupies only 8773 ha [13]. The main national producing region both for table olives and for olive oil is the “Alentejo” region. This southern territory holds a production of 6.9 thousand tons for table olives and around 551.4 thousand tons for olives for oil (Figure 4a). Concerning the surface occupied in Portugal for olive orchards, “Alentejo” is still the principal region for olive oil (184.936 thousand ha) and for table olives (3258 ha) (Figure 4b) [13].
In Portugal, as in the other traditional olive-growing countries, there is a high number of olive varieties. It is estimated that worldwide this number may exceed 3000 with the particularity that only a small number is grown in more than one region [14].
2.1. ‘Galega Vulgar’, the Most Important Portuguese Olive Variety
In Portugal, the production of olive oil and table olive is mostly concentrated in 22 different varieties: ‘Galega vulgar’ or ‘Galega’, ‘Carrasquenha’, ‘Redondil’, ‘Azeitoneira’ or ‘Azeiteira’, ‘Branquita’ or ‘Blanqueta’, ‘Conserva de Elvas’, ‘Negrinha’, ‘Madural’, ‘Cobrançosa’, ‘Verdeal Transmontana’ or ‘Verdeal de Serpa’, ‘Cordovil de Castelo Branco’, ‘Bical de Castelo Branco’, ‘Maçanilha’ or ‘Maçanilha Fina’, ‘Hojiblanca’ and ‘Gordal’ [15].
The main national variety is ‘Galega vulgar’, also called ‘Galega’ (accounting for about 60% of all olive trees in the country) [16] and is associated with five of the six national PDOs (Protected Designations of Origin) regions [17]. The PDO oils are produced in defined geographical areas, with a characteristic soil and climate combination, and are exclusively made from certain olive varieties. These factors, together with the management of the orchard, harvesting and transport to the mill and the oil-makings conditions, give rise to olive oils with unique chemical and sensory characteristics that allow to distinguish them from other oils [18].
A high rusticity and alternate bearing characterize the ‘Galega’ variety (Figure 5). With a low weight fruit (<2 g), an endocarp with a low to medium weight (<0.30 g to 0.30–0.45 g) and a low flesh to stone ratio (5.6 on average) [15,19,20], it stands out for its excellent quality olive oil, and resistance to drought, despite median productivity, partly due to the sensitivity to pests and diseases [19]. ‘Galega’ has a low yield in olive oil (below 18%); this oil is characterized by a low percentage of linoleic acid and a high stability [19,21]. This olive variety shows a high sensitivity to the anthracnose fungus (Colletotrichum spp.), oleander knot (Pseudomonas savastanoi pv. savastanoi), olive fruit fly (Bactrocera oleae) and the black scale (Saissetia oleae). It also exhibits some susceptibility to olive peacock spot (Spilocaea oleagina) [19].
The ‘Galega’ variety presents a high potential for improvement/breeding due to its intra-varietal diversity [22]. Significant progress in ‘Galega’ research has been attained in last years by several national teams, particularly in the areas of ecophysiology, in vitro culture, genetic resources characterization, improvement in production systems and cultural techniques, phytosanitary protection and technology, quality and market organization (Table 1).
The ‘Galega’ productivity currently ranges between 3 and 5 tones/ha (with up to 200 trees/ha in non-irrigated systems) [71]. Comparatively, foreigner varieties productivity is around 8 to 13 tones/ha in super intensive systems [72]. These data point out the importance and the need to improve the productivity of the ‘Galega’ olive trees, in order to hamper its genetic erosion, by increasing its attractiveness to farmers. Nevertheless, to make ‘Galega’ more competitive regarding foreign varieties, it is very important to maintain its high quality while improving its productivity and adaptation to modern production systems.
2.2. Recent Changes on Olive Production Systems and the Importance of a Life Cycle Assessment
Olive trees naturally grow in relatively dry, rustic environments with a Mediterranean climate, characterized by warm, dry summers and rainy, cool winters [73]; these factors shape traditional olive plantations [5].
Traditional olive varieties are the result of empirical selection processes performed by olive growers in a given combination of soil and climatic conditions, with subsequent preservation by vegetative multiplication [14]. In this way, traditional varieties hold a high genetic variability, mainly because of olive allogamy and predominantly self-incompatible nature, tree longevity and long tradition of cultivation [74]. Consequently, different Mediterranean regions are characterized by different local genetic variability, resulting from a mixture of environments and growing systems [75,76]. However, in recent years, the olive germplasm diversity has been modified drastically because of the changes on the determinant characteristics in the choice of a variety. The improvement of the socioeconomic conditions and the exponential progress of technology worldwide have been the principal boosters in a progressive transformation in olive growing techniques [5,77]. The adaptation to the environment, which was the main selection criterion in the past, has been presently replaced by productivity, higher yield in oil and regularity in production, at reduced costs [14]. Consequently, the traditional, manually harvested non-irrigated olive orchards are being replaced by new high-density irrigated and mechanically harvested modern olive plantations [5].
Countries such as Spain, where modern olive groves have been established intensively, have its olive-growing areas predominantly dominated by only a few varieties, with ‘Picual’, ‘Hojiblanca’ and ‘Arbequina’ as the most common [78]. Indeed, ‘Arbequina’ is not only a variety of choice in Spain, but all over the world, and the globalization of some olive varieties may give rise to the tendency to reduce the genetic variability in the modern olive groves even more [76]. In Portugal, this is also occurring, with the main traditional variety in the country (‘Galega’) being replaced, despite its superior-quality olive oil, probably leading to the loss of alleles or of combinations of alleles, in a clear case of genetic erosion [79,80]. Therefore, it is crucial to characterize, conserve and explore the still-existing traditional olive genetic resources in order to prevent the risk of further genetic erosion, towards an efficient use in breeding programs [76].
The replacement of traditional olive varieties by a few foreign ones not only entails negative impacts related to the loss of genetic variability but could also enhance the negative impacts on the environment, related with the different practices and techniques typically associated with these replacement foreigner varieties. Examples are the depletion of natural resources, land degradation, air emissions, waste generation, soil erosion and water scarcity [81,82]. In order to counteract this negative tendency and ensure the olive sector sustainability, life cycle thinking approaches and assessment methods have been applied, allowing a deeper understanding of the role of the impacts from a life cycle perspective. Additionally, these methodologies are fundamental tools to support decision-making in complex value chains, such as the ones related to bioeconomy [81].
A summary of the existing olive sector Life Cycle Assessment (LCA) studies may be found on Table 2, taking into consideration diverse scenarios at different stages of the olive chain.
From these different studies, the agricultural phase was the issue reported as being the most impactful in the environment (i.e., the principal hotspot in the olive oil cycle), mostly due to the utilization of fertilizers, irrigation and phytosanitary treatments [82]. Moreover, distribution and waste management were also considered crucial issues. However, due to the extremely variation of the distribution phase scenarios, they are often excluded from studies. The impacts of waste treatment are also highly variable between studies. Consequently, they might be considered as “avoided” impacts due to energy or nutrient recovery [82,83].
Although the LCA methodology has been applied to calculate and evaluate the potential environmental impacts of the recent changes on olive production systems from the extraction of the raw materials to the oil mill gate, an integrative methodology that takes into consideration environmental, economic and social constraints is still missing. Particularly in Portugal, LCA studies in the olive sector are almost inexistent hampering a deeper characterization and evaluation of the Portuguese olive sector and its principal impacts. Since the olive sector is one of the most valuable bioeconomic value chains in Portugal, the enrichment of Portuguese inventories and impact assessment methods in olive chain is of utmost importance. These characterizations, which will allow future comparisons and inclusions in international studies, together with a deeper characterization of the diversity of Portuguese olive genetic resources, will allow a better knowledge, management, improvement and conservation of the Portuguese olive heritage.
3. Olea europaea Diversity Evolution
3.1. Domestication and Ancestral Origin of Olive Varieties
The geographic origin of olive tree, its domestication and history of its early use are still under debate [101]. According to archaeological and genetic studies, the cultivated olive tree might have developed by domestication of its wild ancestor “oleaster” (Olea europaea subsp. europaea var. sylvestris), approximately 6000 years ago in the Near East [2,102]. Hence, with the discovery of clonal propagation techniques that allowed the vegetative propagation of the best specimens for human consumption, the olive cultivation was boosted across the Mediterranean Basin [101]. Under this scenario of domestication, probably the wild olives behaved as pollen donors to the primary domesticated varieties, diminishing the deleterious effects that might occur in the inbreeding events and giving rise to locally adapted varieties [103].
However, recent evidences clearly indicate the existence of multiple centers of diversity across the Mediterranean area for cultivated olive, and it remains unclear whether the centers of diversity result from one or two local domestication events (Figure 6) [102]. Nuclear microsatellites characterization disclosed the existence of three main cultivated genepools (Q1, Q2 and Q3, Figure 6b), mostly corresponding to three geographic areas: the west (WW), center and east (WE) Mediterranean Basin (Figure 6a) [102,104,105].
Two simplified scenarios of domestication may be considered (Figure 6c,d), during the Late Pliocene with both western and eastern oleaster genepools diverging, with possible gene flow [2,106]. Later, during the Last Glacial Maximum and the Holocene, a population reduction occurred, followed by successive expansion. On the first scenario (Figure 6c), a primary domestication event (green sphere in the figure) is speculated to have taken place in the Eastern Mediterranean Basin, leading to Q3, while Q2 and Q1 were derived from admixture events between Q3 and WW (for Q2) or Q3 and Q2 (for Q1). On the second scenario (Figure 6d), two independent primary domestication events are thought to have occurred, one for Q3 in the Eastern Mediterranean Basin, and one for Q2 in the Central Mediterranean Basin [102].
Regardless of the primary origin of the olive varieties, since the genotypes were clonally propagated, they could be disseminated via human migration [103]. Although the migration history of olive trees is complex, Phoenicians seemed to have started disseminating the olive cultivation in the Iberian Peninsula, but it was the Roman Empire that was principally responsible for extending and encouraging this crop from east to west through both the northern and southern coasts of the Mediterranean Basin [101,107].
The domestication of olive trees promoted larger fruits and seeds, with more pointed ends in cultivated forms, in comparison with wild forms [108]. Nevertheless, some olive varieties such as ‘Galega’ still show considerable morphological diversity with round to acute endocarps of medium to small weights. This could be a sign of an ongoing domestication process or the effect of a long-term clonal propagation process, reducing the number of generations from the wild ancestor or facilitating the existence of overlapping generations [103]. Corroborating this hypothesis, the ‘Galega’ reference sample at the World Olive Germplasm Bank (WOGB) appears as a blend of West and Central Mediterranean germplasm [109] and cultivated-wild admixture [105], and thus, it is possible that ‘Galega’ still holds undomesticated and interesting novel combinations of traits, which might be beneficial for the selection of interesting phenotypes in breeding programs.
As far as the origin of ‘Galega’ is concerned, little is known. Gemas et al. [22] hypothesized that the morphologically diverse ‘Galega’ had a polyclonal origin, supported by the occurrence of somatic mutations and possible sexual reproduction. Hence, the ‘Galega’ variety might have been originated by a naturalization process or as result of hybridization with wild olive trees. However, the study that led to these conclusions was performed using a small non-representative set of the whole ‘Galega’ diversity in terms of tree ages and geographical locations, and thus, a more comprehensive characterization is still needed to unravel the ancestral origin of the ‘Galega’ variety.
Knowledge on the genetic diversity, associated to the morphological differences, detected among and especially within olive varieties is essential to optimize the exploitation of the existing germplasm resources in a long-term successful breeding program.
3.2. The Use of Molecular Markers on Olive Diversity Characterization
Because of the early domestication and following dissemination throughout the Mediterranean area, the olive crop holds a vast number of varieties [110]. Most of these olive varieties are cultivated at the regional/national level, with few varieties dispersed globally [111], and the presence of several synonyms and homonyms makes their classification and description a difficult task [110]. This great diversity brings with it the need to develop tools to easily and efficiently distinguish the different varieties and characterize their genetic variability [112].
Morphological and agronomic characteristics are often the first choice for germplasm description and classification and their use represent an initial step in plant breeding [113]. In 1985, the Unité pour la Protection des Obtentions Végétales (UPOV) developed the olive descriptor list, with the aim of standardizing germplasm data collection and, currently, together with IPGRI (International Plant Genetic Resources Institute), hold the official identification and description of varieties and rootstocks, based on morphological characteristics. However, this methodology is complex and time-consuming due to the long juvenile phase of the trees and the potential environmental influence on morphology [114]. These drawbacks led to the incorporation of new methodologies into fruit certification schemes and characterization of plant varieties namely at DNA level, with molecular markers, which can accelerate and optimize the identification process, by allowing the identification of each genotype at any stage of development and independently of environmental factors [115].
Molecular marker techniques have also become very powerful for the analysis of olive genetic diversity. Additionally, if certain molecular markers are associated with interesting characteristics, these markers may become valuable tools for accelerating selection in breeding programs [116].
In olive, the lack of sufficient genetic information hampered the use of species-specific markers until the 2000s [117]; however, many studies have been performed with non-species-specific molecular markers. In Table 3, some applications of these initial non-species-specific molecular markers in olive varieties are shown.
However, considering the drawbacks of these kind of molecular markers, such as low reproducibility (RAPDs) or being time consuming or expensive (RFLPs, AFLPs), the use of species-specific markers as SSRs (Simple Sequence Repeats, also called microsatellites) and SNPs (Single-Nucleotide Polymorphisms) have been increasing over the last few years in olive analysis [76,134].
SSRs molecular markers, consisting of tandem repeats of mono-, bi-, tri- or tetra-nucleotides in the eukaryotic genome, present high polymorphism based on the number of the nucleotide motif repeats [135]. SSRs are also characterized by a codominant inheritance, good reproducibility between laboratories and easy detection by PCR. In addition, SSRs can be transferred to related species since the sequences flanking the microsatellites are highly conserved [136,137]. Due to these characteristics, SSRs have been the marker of choice for the molecular characterization, genetic diversity and relationship studies in olive trees [46,105,137,138,139] until the expansion of SNP markers.
SNP markers are individual nucleotide base differences between two DNA sequences, while the polymorphism of microsatellite markers is due to length variations [117,140]. SNPs can be classified according to nucleotide substitution as either transitions or transversions. As a nucleotide base is the smallest unit of inheritance, SNPs provide the ultimate form of molecular marker [140]. Compared with other genetic markers, SNPs are more abundant in the genome, are stably inherited, and can occur in coding regions, occasionally leading to amino acid changes in the polypeptides encoded. They are, in most cases, bi-allelic, codominant, and amenable to high-throughput genotyping and automation [141]. SNP technology is being used to characterize germplasm towards its application in breeding programs. Broad adoption of this technology would be useful also to the plant protection regulatory systems, especially for variety identification and protection purposes. While SSRs are now generally accepted in the courts, some inherent limitations would be overcome using SNP. For instance, high-density oligonucleotide arrays on DNA chips have been developed to analyze genotypes with SNPs. These chips use nucleic acids immobilized on solid-state surfaces, to be hybridized with the sample, without requiring the time-consuming and labor-demanding assays that are needed for most molecular markers (often SSR), essential for size separation of multiple DNA fragments [140]. However, SSRs are still the most used molecular markers for varieties characterization and for diversity studies, since they are cheaper and less technically demanding compared to SNPs.
Table 4 elucidates the growing importance of SSRs and SNPs in olive, by representing the different analysis performed with these markers.
3.3. The Importance of Olea Germplasm Conservation
The recent renewal of old olive orchards in the main olive-producing countries, due to socioeconomic changes, lead to intensive modes of production, with the use of high-yielding and low-vigor varieties. This trend may potentially lead to local olive germplasm erosion, due to the progressive abandonment of several autochthonous and ‘local’ varieties [110,159]. In a climate changing world, with increased frequencies of severe droughts and flooding and the appearance of new diseases and pests, the presence of few varieties in the olive orchards could represent a problem, since monovarietal systems might be less resilient than the more traditional systems [159]. Taking all this into account, the study of traditional varieties and the preservation of its genetic diversity in in situ or ex situ collections are key points, towards the efficient exploitation of these genetic resources, namely by providing breeding material that could improve and diversify modern orchards [110,147,159].
International policies have been taken and projects developed to tackle the continuous loss of olive genetic diversity, due to social, economic and climate changes. For instance, the International Olive Oil Council has been the promoter in 1995 of the European project RESGEN (‘Conservation, characterization, collection and utilization of the olive genetic resources’), which aimed at the collection, characterization and conservation of olive genetic resources as well as at the introduction of germplasm from different countries in national in field ex-situ collections [110]. Moreover, in 2001, the International Treaty on Plant Genetic Resources for Food and Agriculture [160] established a plan for an equitable use of these resources and pictured a global information system to recover and share plant genetic resources [161]. Consequently, prospecting surveys and exchanges of olive varieties between many countries have contributed to the ex situ conservation of this germplasm in several institutions [162]. Currently, important collections exist in all the main olive-growing countries, being the two largest existing ex situ olive germplasm collections, the worldwide olive germplasm Bank of Córdoba, in Spain (WOGBC) and the worldwide olive germplasm Bank of Marrakech, in Morocco (WOGBM) [110,159,162]. In Portugal, a reference collection of olive varieties was set by the Instituto Nacional de Investigação Agrária e Veterinária, I.P (INIAV) in 2012, aiming to conserve and study, under the same edaphoclimatic conditions, the Portuguese olive varieties [16].
In what concerns the ‘Galega’ variety, due to its current replacement in the Portuguese groves, it is essential to prospect and characterize the still available diversity and invest in its conservation, either in situ, through the preservation of ancient isolated individuals or centenary old groves, or ex situ, through the installation of the prospected diversity in comparative field trials.
4. Olive Breeding Worldwide
Across time, olive breeding evolved from an empirical selection performed by olive growers to a more scientific-based selection, using new genomic tools to improve and accelerate breeding programs. This evolution is closely related to the characteristics used to choose a particular variety, which have been changing over time. In fact, presently, breeding objectives include improving yield while including greater quality and value-added traits, whereas adaptation to the environment was the main selection criteria in the past [14,163].
Compared to other fruit crops, olive breeding is in its infancy, since until recently the agronomic evaluation of olive varieties was incomplete and the efforts to explore and characterize true-to-type olive varieties was very fragmented [110,162]. Although some progress has been achieved, it is fundamental to plan more efficient and faster breeding programs, through the use of new approaches [164]. In fact, the majority of the studies performed in wild and cultivated germplasm have focused on describing the existent variability and not in the direct implication in breeding programs. Thus, there is still a gap of knowledge between the available olive germplasm and its effective use for breeding purposes [165].
4.1. From Conventional to Precision Olive Breeding
The origin of olive growing is connected with the discovery of vegetative propagation using cuttings in the Middle East [164], and since then, the selection and propagation were performed by empirical knowledge by the olive growers. Across time, this empirical selection progressively evolved to a more scientific based selection, mainly performed on the direct observation, clonal selection and cross-breeding of individuals with the most desirable phenotypic characteristics by breeders [110]. In fact, most of the traditional olive varieties were selected from singular seedlings with remarkable traits and then clonal propagated, resulting in all current varieties from selected clones [162]. Presently, with the emergence of molecular techniques and the arising of genomic tools, olive breeding is being improved.
Classic methodologies have been applied worldwide to obtain high yield, early bearing varieties, adapted to the new mechanized orchards [5,166], resistant to plagues and diseases [167,168] or to enlarge the basis of genetic variability [168,169,170]. Additionally, in Portugal olive breeding was mostly based on the classic hybridization and selection in the progeny. In 2002 an olive breeding program with ‘Galega’ and ‘Cobrançosa’ varieties was initiated, aiming at more productive varieties, with good quality olive oil and higher levels of resistance to diseases and pests, and with better adaptation to mechanized harvesting [16].
Most of the conventional methods are time consuming and failed to meet several requirements of olive production and the demands of the olive industry, namely in enhancing yield and quality of its products, and accelerate development of new varieties and rootstocks [171]. In order to overcome these limitations, several biotechnological advances are currently used in Olea breeding [172]. In Table 5, several precision breeding approaches and breeding innovations in olive are summarized.
In vitro techniques have been developed and successfully used to propagate olive varieties and, in several cases, the resulting in vitro plants have been transplanted in the field [171,175]. The micropropagated materials have been used for screening resistance to biotic and abiotic stress, pathogen virulence and host interaction with parasites focusing on olive genetic improvement [172]. The use of axillary bud stimulation, organogenesis and somatic embryogenesis techniques are also being used to produce plants on a large scale. These techniques allow the production of pathogen-free genotypes, embryo germination, germplasm preservation, plant regeneration from cell tissues and synthetic seed production [171]. Olive genetic transformation has been used to correct some defects of several varieties, reducing the time and cost of genetic improvement. Currently, this ‘plant gene therapy’ is used with a diversity of objectives, such as improvement of rooting ability, modification of canopy architecture and improvement of the tolerance to biotic and abiotic stresses [172].
Many efforts have also been made in the identification and annotation of genes, through expression analysis studies, based for instance on the identification of ESTs (expressed sequence tags). These studies concerned mainly allergens in olive pollen, characteristics of fruits, olive oil and disease resistance [171,180]. Furthermore, important progress has been accomplished in olive transcriptomic research, with several olive transcriptomes generated from different organs and adaptive responses [190], namely to address flower and fruit development [180,185,190,191], fruit abscission [182], response to abiotic and biotic stresses [181,186,187], plant architecture [188], miRNA [183] and comparative transcriptomics [189].
The entire olive plastome [43,192] provided new information on the olive nucleotide sequence and has been used in different studies such as the assessment of the genetic relationship of olive varieties and the phylogeny of olive [2,102].
Although the described advances in precision breeding techniques, the knowledge about the inheritance of most genes controlling complex agronomical performance and quality traits is still scarce, which consequentially have restricted breeding strategies to clonal or varietal selection [194]. Therefore, understanding the basis of quantitative agronomic traits might help plant breeders to improve important characteristics that are controlled by multiple genes and start a precision olive genetic improvement [171,195].
4.2. Marker Assisted Selection in Olive Breeding
Marker-assisted selection (MAS) for traits controlled by major genes or quantitative trait loci (QTLs) is the primary use of genomics in breeding [164]. However, despite the development and the successful application of different molecular marker systems in olive diversity analysis, their direct application for dissecting agronomic traits or develop trait-associated markers for MAS in breeding has been limited.
The first olive linkage map was developed by De la Rosa et al. [120] using dominant markers (RAPDs and AFLPs) and a limited number of codominant markers (RFLPs and SSRs) in 2003. This pioneer work was followed by others aiming at more saturated, with more markers and longer linkage groups maps [44,125,126,133,149].
All the olive genetic maps available until 2015 were based on non-sequence-based markers (for instance AFLPs) or on low-coverage markers (for example SSRs). However, with the rising of sequencing technologies new high-throughput markers became available, such as SNPs. In fact, genotyping-by-sequencing (GBS), an SNP identification method based on next-generation sequencing (NGS) technologies, showed to be very effective for identify high number of SNP markers and construct high-density genetic linkage maps in olive [156]. Based on this new technology, a high-density genetic linkage map for the olive genome was developed by İpek et al. [157], using 5643 markers (21 SSRs, 203 AFLPs, and 5736 SNPs), representing the most saturated genetic linkage map in olive up to date. Only after this study, it was possible to start locating QTLs for MAS breeding (excepting for some preliminary data from Ben Sadok et al. [149], Atienza et al. [196], Ates [197] and González-Plaza et al. [188]). Additionally, in 2016, Marchese et al. [158] using GBS, constructed a genetic map with 1597 SNP markers, with this linkage map being a useful resource for the study of tree habit and vigor traits segregating in the respective progeny.
Understanding the genetic architecture of important agronomic traits is essential for an effective use of the existent genetic variation in this plant breeding [198,199]. With the development of high-throughput genotyping a transition between the traditional QTL mapping to association or LD (Linkage Desiquilibrium) mapping occurred, which assess the correlation between the phenotype and genotype in populations of unrelated individuals instead of familiar related [199].
Genome-wide association studies (GWAS), can take advantage of many generations of historical recombination in a diverse population and avoid the time-consuming generations of controlled crosses [199,200], providing a higher mapping resolution [201].
Despite the high potential of GWAS, this approach has been almost unused in olive. Kaya et al. [195] published the first olive GWAS study, where some significant markers-trait associations were detected for fruit and stone weight. More recently, GWAS was performed in a diverse panel of olive accessions to identify markers associated with five agronomic traits [201] and using 97 olive varieties, the association of the 5′UTR intron of the FAD2-2 gene with oleic and linoleic acid content [202].
Concerning ‘Galega’, a detailed characterization of the existing genetic variability, with an effective environmental interaction control, using molecular and agro-morphological information is still missing, hampering the application of these genomic approaches. Consequently, in order to start the application of these tools towards a ‘Galega’ breeding program, is important to establish a representative collection of the genetic variability of this variety, under the same environment, to allow phenotypic data screening, which will integrate a future GWAS.
4.3. Genotype to Phenotype (G2P) Prediction Models: The Growing Importance of Integrating Environmental Sensitivities and Crop Growth Models Information
As already highlighted, most of the times, the target traits in olive breeding, such as yield, oil quality, and stress resistance, are complex traits, characterized by the contribution of several genes with relatively small effects, and influenced by genotype-by-environment interactions (G × E) [203]. These complex traits very often difficult in the breeding process [204]. Nevertheless, the basis for the success of a plant breeding program lies on plant genotypes with higher performance (phenotype) in yield and/or quality in different environmental conditions. However, to guarantee this high performance, an understanding of the factors that contribute to the desirable phenotype is needed [205].
Traditional empirical methodologies have been used to identify superior individuals, which involve measuring phenotypic performance in broad segregating populations in different environmental trials, and subsequent application of statistical methods based on quantitative genetics. However, this methodology holds some drawbacks, as interactions with management are not integrated and the G × E might puzzle the selection [206]. Therefore, suitable genotype-to-phenotype (G2P) models that are able to detect genetic and environmental factors, which influence phenotypic variation, and predict the phenotype from genetic and environmental inputs, are needed [207].
The existing models for phenotype prediction hold a blend of statistical, genetic and physiological elements, as is the case of the linear mixed models (LMMs) and crop growth models (CGMs) [203,208]. LMMs can be described with a fixed part, which corresponds to the mean and a random part, composed of variances and covariances [208]. Smith et al. [209] performed a review of the use of mixed models to analyze complex data sets in plant breeding. CGMs integrate genetic and environmental variables in a natural way over time [203] and they have been widely used in plant breeding, namely in the characterization of environments, prediction of consequences of trait variation on yield within a genotype × environment × management context, assessment of hybrid performance and evaluation of breeding strategies [210].
The G2P models are of high relevance, since they can address several design issues, namely in the decision of the optimal environments and genotypes to use for phenotyping, the choice of the most important traits in accordance with the different environmental types and the selection of which phenotyping schedule to use [203]. On the other hand, the use of olive CGM [211] will indicate what is the optimum time to measure the different agronomic traits.
Since ‘Galega’ is present in almost all the Portuguese territory, it is important to phenotype its genetic variability in different environments, possibly with different managements practices, and with a good climate and soil characterizations, for the development of an optimal G2P prediction model. With this model integrating the phenotypic and genotypic diversity, using the soil and climate characteristics as covariates, it will be possible to predict the production and quality of an olive clone in a certain environment or production system.
5. Future Prospects for a Sustainable Conservation and Use of the ‘Galega’ Variety
The traditional varieties of olives due to the many centuries of selection and propagation are not genetically homogenous entities, being composed of different genotypes with different yield and quality potential. Thus, the full exploitation of their potential has to go through the selection of the best intra-varietal genotypes that should provide the highest yields and the best quality, but considering also that from the sustainability point of view, it is important to maintain a certain level of diversity.
From the above, a more efficient and effective selection will be possible through the complementary morpho-agronomic and molecular characterization of the existent diversity, with a clear understanding of the environmental influence on the trait’s expression. This selection will allow the identification of the most interesting clones as the basis for future breeding programs. The development of high-throughput DNA sequencing technologies made available large molecular resources and will support the discovery of controlling genes and regulatory sequences also in olives. In addition, with the assembled draft genome of O. europaea [212], breeding programs will be enhanced, as this genomic tool will facilitate the study of the genotype and its relationship with the phenotype.
However, before the accomplishment of the above, it is essential to start by characterizing the current variability in olive varieties.
In the case of Portugal, since ‘Galega’ is the main variety and is being replaced by other foreign varieties, it is essential to characterize the available diversity and create the indispensable conditions to overcome the agronomic limitations still found in this variety. Therefore, to tackle these limitations we propose that efforts should be made to molecularly characterize the existent genetic diversity at the traditional Portuguese olive orchards, allowing as well to unravel the ‘Galega’ ancestral origin. Furthermore, it will be necessary to create conditions to install, under the same environment, the genetic diversity found, to properly apply association mapping approaches. Thus, the identification of the genetic control of the characteristics to improve within ‘Galega’ will be possible with higher resolution and by developing these molecular tools and associated phenotype predictive models (G2P), a more accurate selection and exploitation of this variety could be accomplished. Hence, the valorization of the Portuguese bioeconomic olive sector will be attained by the introduction of adapted improved ‘Galega’ clones, reinforcing the Portuguese international market niche associated with olive oils and olives of excellent quality.
Author Contributions
Conceptualization, H.S. and M.C.V.P.; writing and draft preparation, H.S.; writing—review and editing, M.C.V.P. and J.N.; funding acquisition, M.C.V.P. and J.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by 3i Bioeconomy project POCI-01-0246-FEDER-026758, funded by the Fundo Europeu de Desenvolvimento Regional (FEDER), under the “Programa Operacional Temático Competitivdade e Internacionalização”—COMPETE 2020, FCT (Fundação para a Ciência e a Tecnologia), MCTES (Ministério da Ciência, Tecnologia e Ensino Superior) and the Association BLC3 through the PhD grant [SFRH/BDE/102401/2014], FCT Investigator 2014 contract (IF/01337/2014), the R&D Unit, UIDB/04551/2020 (GREEN-IT—Bioresources for Sustainability), and the Centre Bio R&D Unit (UID/05083/2020).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rugini, E.; Baldoni, L.; Muleo, R.; Sebastiani, L. Preface. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. v–vi. [Google Scholar] [CrossRef]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; Brunini-Bronzini de Caraffa, V.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. R. Soc. B 2013, 280, 20122833. [Google Scholar] [CrossRef] [Green Version]
- Böhm, J. Domesticação da oliveira na antiguidade na bacia mediterrânica. In O Grande Livro da Oliveira e do Azeite—Portugal Oleícola; Böhm, J., Ed.; Dinalivro Editora: Lisboa, Portugal, 2013; pp. 52–65. ISBN 978-972-576-620-0. [Google Scholar]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2014, 10, 141–155. [Google Scholar] [CrossRef]
- Rallo, L.; Barranco, D.; Castro-García, S.; Connor, D.J.; Gómez del Campo, M.; Rallo, P. High-density olive plantations. Hortic. Rev. 2013, 41, 303–383. [Google Scholar] [CrossRef]
- FAO; Plan Bleu. State of Mediterranean Forests 2018; Food and Agriculture Organization of the United Nations: Rome, Italy; Plan Bleu: Marseille, France, 2018; Available online: https://ec.europa.eu/knowledge4policy/publication/state-mediterranean-forests-2018_en (accessed on 20 June 2020).
- Hashmi, M.A.; Khan, A.; Hanif, M.; Farooq, U.; Perveen, S. Traditional uses, phytochemistry, and pharmacology of Olea europaea (olive). Evid. Based Complement. Altern. Med. 2015, 2015, 541591. [Google Scholar] [CrossRef] [Green Version]
- Saldanha, M.H. Benefícios do Azeite na Saúde Humana. Coleção Estudos e Análises 7; Direção-Geral de Desenvolvimento Rural: Lisboa, Portugal, 1999. Available online: https://www.dgadr.gov.pt/component/jdownloads/send/3-agricultura-e-desenvolvimento-rural/12-beneficios-do-azeite-na-saude-humana (accessed on 20 June 2020).
- Khadari, B.; Breton, C.; Moutier, N.; Roger, J.; Besnard, G.; Bervillé, A.; Dosba, F. The use of molecular markers for germplasm management in a French olive collection. Theor. Appl. Genet. 2003, 106, 521–529. [Google Scholar] [CrossRef]
- International Olive Council (IOC). Economic Affairs & Promotion Unit—Figures. Available online: https://www.internationaloliveoil.org/what-we-do/economic-affairs-promotion-unit/#figures (accessed on 20 June 2020).
- CONSULAI; Juan-Vilar, Consultores Estratégicos. Alentejo: A Liderar a Olivicultura Moderna Internacional. Olivum. 2019. Available online: https://13b249f8-94d9-4c10-832b9c2323cab575.filesusr.com/ugd/a303d9_5993f29b65054e46a54accff8c90cf7f.pdf (accessed on 20 June 2020).
- Reis, P. O Olival em Portugal. Dinâmicas, Tecnologias e Relação com o Desenvolvimento Rural; Animar—Associação Portuguesa para o Desenvolvimento Local: Lisboa, Portugal, 2014; ISBN 978-989-8748-06-5. [Google Scholar]
- Instituto Nacional de Estatística (INE). Estatísticas Agrícolas. Available online: https://www.ine.pt/xportal/xmain?xpid=INE&xpgid=ine_publicacoes&PUBLICACOESpagenumber=1&PUBLICACOEScoleccao=107660&PUBLICACOEStipo=ea&selTab=tab0 (accessed on 20 June 2020).
- Cordeiro, A.M.; Inês, C.; Morais, N. Aspetos gerais da cultura da oliveira: Principais cultivares de oliveira existentes em Portugal. In Boas Práticas no Olival e no Lagar; INIAV, I.P., Ed.; SIG—Sociedade Industrial Gráfica: Lisboa, Portugal, 2014; pp. 37–54. ISBN 978-972-579-041-0. [Google Scholar]
- Leitão, F.; Pontes, M.F.; Calado, M.L.; Almeida, F.J. Descrição de 22 Variedades de Oliveira Cultivadas em Portugal; Ministério da Agricultura, Pescas e Alimentação, Direcção-Geral do Planeamento e Agricultura: Lisboa, Portugal, 1986. Available online: https://www.dgadr.gov.pt/component/jdownloads/send/10-diversos/26-descricao-de-22-variedades-de-oliveira-cultivadas-em-portugal (accessed on 20 June 2020).
- Arias-Calderón, R.; Carvalho, M.T.; Cordeiro, A. A Importância dos Progenitores no Programa de Melhoramento da Oliveira por Hibridação; Instituto Nacional de Investigação Agrária e Veterinária, I.P. (INIAV): Lisboa, Portugal, 2017; Available online: http://www.iniav.pt/fotos/editor2/a_importancia_dos_progenitores_oliveiras.pdf (accessed on 15 July 2020).
- Gouveia, J.; Saldanha, J.; Martins, A.; Modesto, M.; Sobral, V. O Azeite em Portugal; Edições Inapa: Sintra, Portugal, 2002; ISBN 978-972-797-024-7. [Google Scholar]
- Costa, R. Caracterização do Perfil Fenólico e da Ultraestrutura do Pólen de Cultivares de Olea europaea L. como Contributo para uma Classificação Varietal. Master’s Thesis, University of Coimbra, Coimbra, Portugal, 2011. Available online: http://hdl.handle.net/10316/20275 (accessed on 21 June 2020).
- Cordeiro, A.M.; Calado, M.L.; Morais, N.; Miranda, A.; Carvalho, M.T. Variedades de Oliveira: ‘Galega Vulgar’; Direção Regional de Agricultura e Pescas do Centro: Castelo Branco, Portugal, 2010. Available online: https://www.drapc.gov.pt/base/documentos/018_variedades_oliveira_galega_vulgar_nov10.pdf (accessed on 20 June 2020).
- Del Río, C.; Caballero, J.M. Variability and classification of olive cultivars by fruit weight, flesh/stone ratio and oil percentage. Acta Hortic. 2008, 791, 39–44. [Google Scholar] [CrossRef]
- Albuquerque, T.G.; Costa, H.S.; Oliveira, M.B.P.P. An overview of Portuguese olive oils and table olives with protected designation of origin. Eur. J. Lipid Sci. Tech. 2019, 121, 1800129. [Google Scholar] [CrossRef]
- Gemas, V.J.; Almadanim, M.C.; Tenreiro, R.; Martins, A.; Fevereiro, P. Genetic diversity in the olive tree (Olea europaea L. subsp. europaea) cultivated in Portugal revealed by RAPD and ISSR markers. Genet. Resour. Crop Evol. 2004, 51, 501–511. [Google Scholar] [CrossRef]
- Serrano, M.C.F.; Amaral, L. Irrigation effects on some fructification parameters of olive cvs. Olea europaea L. ‘Galega vulgar’, ‘Cordovil de Serpa’ and ‘Verdeal Alentejana’ in Alentejo. Acta Hortic. 2008, 791, 325–331. [Google Scholar] [CrossRef]
- Coelho, R.; Sousa, A.; Rato, A.; Vaz, M. Response to salinity in young olive trees of three Iberian varieties. In Proceedings of the 12th Portuguese-Spanish Symposium on Plant Water Relations, Évora, Portugal, 30 September–3 October 2014; pp. 99–104. [Google Scholar]
- Peixe, A.; Antunes, A.; Hegewald, H.; Costa, C.; Pinto, A.P. Preliminary results on the evaluation of auxin endogenous levels and oxidative enzymes activity, among the rooting of two olive (Olea europaea L.) cultivars ‘Galega vulgar’ and ‘Cobrançosa’. In Atas Portuguesas de Horticultura 14, Proceedings of the 5th Simpósio Nacional de Olivicultura, Santarém, Portugal, 24–26 September 2009; Associação Portuguesa de Horticultura: Lisboa, Portugal, 2011; pp. 13–22. [Google Scholar]
- Lopes, J.P.P.S. Polinização em Oliveira cvs. Galega e Cobrançosa: Avaliação do Vingamento em Ensaios de Polinização Controlada e do Efeito da Aplicação de um Bioestimulante. Master’s Thesis, Instituto Superior de Agronomia, University of Lisboa, Lisboa, Portugal, 2011. Available online: http://hdl.handle.net/10400.5/4122 (accessed on 29 June 2020).
- Azevedo, A.; Bernardes, P. Influência das condições ecológicas no crescimento do olival em regime superintensivo. Reposta da cultivar Galega vulgar à fertilização azotada. In Proceedings of the 1st Congresso Nacional das Escolas Superiores Agrárias, Bragança, Portugal, 2–3 December 2015. [Google Scholar]
- Dias, M.C.; Pinto, D.C.G.A.; Freitas, H.; Santos, C.; Silva, A.M.S. The antioxidant system in Olea europaea to enhanced UV-B radiation also depends on flavonoids and secoiridoids. Phytochemistry 2020, 170, 112199. [Google Scholar] [CrossRef]
- Maia, F.B. Contribuição para o Estudo da Fenologia de Quatro Variedades de Oliveira (Olea europaea L.): ‘Arbequina’, ‘Cobrançosa’, ‘Galega Vulgar’ e ‘Picual’. Master’s Thesis, Instituto Superior de Agronomia, University of Lisboa, Lisboa, Portugal, 2010. Available online: http://hdl.handle.net/10400.5/15199 (accessed on 29 June 2020).
- Cordeiro, A.M.; Martins, P.C.S.; Ramos, A.; Sequeira, P. Characterization of olive cultivars fruit set in self-pollination. In Atas Portuguesas de Horticultura 14, Proceedings of the 5th Simpósio Nacional de Olivicultura, Santarém, Portugal, 24–26 September 2009; Associação Portuguesa de Horticultura: Lisboa, Portugal, 2011; pp. 23–30. [Google Scholar]
- Jimenez-Lopez, J.C.; Morales, S.; Castro, A.J.; Volkmann, D.; Rodríguez-García, M.I.; Alche, J.D. Characterization of profilin polymorphism in pollen with a focus on multifunctionality. PLoS ONE 2012, 7, e30878. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, H.; Calado, L.; Cunha, M.; Abreu, I. Evaluation of pollen viability and germination from 20 varieties of Olea europaea L. grown in Portugal. In Atas Portuguesas de Horticultura 14, Proceedings of the 5th Simpósio Nacional de Olivicultura, Santarém, Portugal, 24–26 September 2009; Associação Portuguesa de Horticultura: Lisboa, Portugal, 2011; pp. 39–46. [Google Scholar]
- Velada, I.; Grzebelus, D.; Lousa, D.; Soares, C.M.; Macedo, E.S.; Peixe, A.; Arnholdt-Schmitt, B.; Cardoso, H.G. AOX1-subfamily gene members in Olea europaea cv. “Galega Vulgar”—gene characterization and expression of transcripts during IBA-induced in vitro adventitious rooting. Int. J. Mol. Sci. 2018, 19, 597. [Google Scholar] [CrossRef] [Green Version]
- Velada, I.; Cardoso, H.; Porfirio, S.; Peixe, A. Expression profile of PIN-formed auxin efflux carrier genes during IBA-induced in vitro adventitious rooting in Olea europaea L. Plants 2020, 9, 185. [Google Scholar] [CrossRef] [Green Version]
- Macedo, E.; Vieira, C.; Carrizo, D.; Porfirio, S.; Hegewald, H.; Arnholdt-Schmitt, B.; Calado, M.L.; Peixe, A. Adventitious root formation in olive (Olea europaea L.) microshoots: Anatomical evaluation and associated biochemical changes in peroxidase and polyphenol oxidase activities. J. Hortic. Sci. Biotech. 2013, 88, 53–59. [Google Scholar] [CrossRef]
- Porfirio, S.; Calado, M.L.; Noceda, C.; Cabrita, M.J.; Silva, M.G.; Azadi, P.; Peixe, A. Tracking biochemical changes during adventitious root formation in olive (Olea europaea). Sci. Hortic. 2016, 204, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, A.V.A.P. Simplificação do Processo de Multiplicação in vitro da Oliveira “Olea europaea L.”. Master’s Thesis, University of Évora, Évora, Portugal, 2016. Available online: http://hdl.handle.net/10174/20702 (accessed on 29 June 2020).
- Pires, R.N. Ensaios com Vista à Indução de Embriogénese Somática em Oliveira (Olea europaea L.). Master’s Thesis, University of Évora, Évora, Portugal, 2018. Available online: http://hdl.handle.net/10174/23981 (accessed on 29 June 2020).
- Rallo, P.; Dorado, G.; Martin, A. Development of simple sequence repeats in olive tree (Olea europaea L.). Theor. Appl. Genet. 2000, 101, 984–989. [Google Scholar] [CrossRef]
- Díaz, A.; De la Rosa, R.; Martín, A.; Rallo, P. Development, characterization and inheritance of new microsatellites in olive (Olea europaea L.) and evaluation of their usefulness in cultivar identification and genetic relationship studies. Tree Genet. Genomes 2006, 2, 165–175. [Google Scholar] [CrossRef]
- Martins-Lopes, P.; Lima-Brito, J.; Gomes, S.; Meirinhos, J.; Santos, L.; Guedes-Pinto, H. RAPD and ISSR molecular markers in Olea europaea L.: Genetic variability and molecular cultivar identification. Genet. Resour. Crop Evol. 2007, 54, 117–128. [Google Scholar] [CrossRef]
- Gomes, S.; Martins-Lopes, P.; Lopes, J.; Guedes-Pinto, H. Assessing genetic diversity in Olea europaea L. using ISSR and SSR markers. Plant Mol. Biol. Rep. 2009, 27, 365–373. [Google Scholar] [CrossRef]
- Mariotti, R.; Cultrera, N.G.M.; Díez, C.M.; Baldoni, L.; Rubini, A. Identification of new polymorphic regions and differentiation of cultivated olives (Olea europaea L.) through plastome sequence comparison. BMC Plant Biol. 2010, 10, 211. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-García, M.C.; Belaj, A.; De la Rosa, R.; Šatović, Z.; Heller-Uszynska, K.; Kilian, A.; Martín, A.; Atienza, S.G. Development of DArT markers in olive (Olea europaea L.) and usefulness in variability studies and genome mapping. Sci. Hortic. 2012, 136, 50–60. [Google Scholar] [CrossRef]
- Parra-Lobato, M.C.; Delgado-Martinez, F.J.; Gomez-Jimenez, M.C. Morphological traits and RAPD markers for characterization and identification of minor Spanish olive cultivars from the Extremadura region. Genet. Mol. Res. 2012, 11, 2401–2411. [Google Scholar] [CrossRef] [PubMed]
- Lazović, B.; Adakalić, M.; Pucci, C.; Perović, T.; Bandelj, D.; Belaj, A.; Mariotti, R.; Baldoni, L. Characterizing ancient and local olive germplasm from Montenegro. Sci. Hortic. 2016, 209, 117–123. [Google Scholar] [CrossRef]
- Veloso, M.; Simões-Costa, M.C.; Carneiro, L.C.; Guimarães, J.B.; Mateus, C.; Fevereiro, P.; Pinto-Ricardo, C. Olive tree (Olea europaea L.) diversity in traditional small farms of Ficalho, Portugal. Diversity 2018, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.S.; Mendonça, D.; Sefc, K.M.; Gil, F.S.; Da Câmara Machado, A. Genetic evidence of intra-cultivar variability within Iberian olive cultivars. HortScience 2004, 39, 1562–1565. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, A.I.; Sanchez-Sevilla, J.F.; Alvarez-Tinaut, M.C.; Gomez-Jimenez, M.C. Genetic diversity assessment in Portugal accessions of Olea europaea by RAPD markers. Biol. Plant. 2008, 52, 642–647. [Google Scholar] [CrossRef]
- Fevereiro, P.; Leitão, F.; Potes, F.; Gemas, V.; Alves, M.; Favoretto, P. The Portuguese olive (Olea europaea subsp. europaea) germplasm. Acta Hortic. 2011, 924, 291–298. [Google Scholar] [CrossRef]
- Figueiredo, E.; Canhoto, J.; Ribeiro, M.M. Fingerprinting and genetic diversity of Olea europaea L. ssp. europaea accessions from the cultivar Galega using RAPD markers. Sci. Hortic. 2013, 156, 24–28. [Google Scholar] [CrossRef]
- Cultrera, N.G.M.; Sarri, V.; Lucentini, L.; Ceccarelli, M.; Alagna, F.; Mariotti, R.; Mousavi, S.; Ruiz, C.G.; Baldoni, L. High levels of variation within gene sequences of Olea europaea L. Front. Plant Sci. 2019, 9, 1932. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.R. Diversidade Genómica e Genética Intra-Varietal em Oliveira ‘Galega Vulgar’. Master’s Thesis, Instituto Superior de Agronomia, University of Lisboa, Lisboa, Portugal, 2019. Available online: http://hdl.handle.net/10400.5/18359 (accessed on 29 June 2020).
- Dias, A.B.; Pimentel, R.S.; Pinheiro, A.; Peça, J.O. Resultados da avaliação da poda com máquina de discos num olival intensivo da variedade “Galega vulgar”. In Actas de Horticultura 73, Proceedings of the 1st Congreso Ibérico de Olivicultura, 5th Jornadas Nacionales del Grupo de Olivicultura de la Sociedad Española de Ciencias Hortícolas (SECH) and 7th Simpósio Nacional de Olivicultura da Associação Portuguesa de Horticultura (APH), Badajoz-Elvas, Spain, Portugal, 13–15 April 2016; Sociedad Española de Ciencias Hortícolas: Córdoba, Spain, 2018; pp. 194–201. [Google Scholar]
- Goldental-Cohen, S.; Biton, I.; Many, Y.; Ben-Sason, S.; Zemach, H.; Avidan, B.; Ben-Ari, G. Green olive browning differ between cultivars. Front. Plant Sci. 2019, 10, 1260. [Google Scholar] [CrossRef]
- Landum, M.C.; Félix, M.R.; Alho, J.; Garcia, R.; Cabrita, M.J.; Rei, F.; Varanda, C.M.R. Antagonistic activity of fungi of Olea europaea L. against Colletotrichum acutatum. Microbiol. Res. 2016, 183, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Materatski, P.; Varanda, C.; Carvalho, T.; Dias, A.B.; Campos, M.D.; Rei, F.; Félix, M.R. Spatial and temporal variation of fungal endophytic richness and diversity associated to the phyllosphere of olive cultivars. Fungal Biol. 2019, 123, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Varanda, C.M.R.; Materatski, P.; Landum, M.; Campos, M.D.; Félix, M.R. Fungal communities associated with peacock and cercospora leaf spots in olive. Plants 2019, 8, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M.D.; Zellama, M.S.; Varanda, C.; Materatski, P.; Peixe, A.; Chaouachi, M.; Félix, M.R. Establishment of a sensitive qPCR methodology for detection of the olive-infecting viruses in Portuguese and Tunisian orchards. Front. Plant Sci. 2019, 10, 694. [Google Scholar] [CrossRef]
- Gouvinhas, I.; Martins-Lopes, P.; Carvalho, T.; Barros, A.; Gomes, S. Impact of Colletotrichum acutatum pathogen on olive phenylpropanoid metabolism. Agriculture 2019, 9, 173. [Google Scholar] [CrossRef] [Green Version]
- Materatski, P.; Varanda, C.; Carvalho, T.; Dias, A.B.; Campos, M.D.; Gomes, L.; Nobre, T.; Rei, F.; Félix, M.R. Effect of long-term fungicide applications on virulence and diversity of Colletotrichum spp. associated to olive anthracnose. Plants 2019, 8, 311. [Google Scholar] [CrossRef] [Green Version]
- Azevedo-Nogueira, F.; Gomes, S.; Carvalho, T.; Martins-Lopes, P. Development of high-throughput real-time PCR assays for the Colletotrichum acutatum detection on infected olive fruits and olive oils. Food Chem. 2020, 317, 126417. [Google Scholar] [CrossRef]
- Alves, E.; Melo, T.; Barros, M.P.; Domingues, M.R.M.; Domingues, P. Lipidomic profiling of the olive (Olea europaea L.) fruit towards its valorisation as a functional food: In-depth identification of triacylglycerols and polar lipids in Portuguese olives. Molecules 2019, 24, 2555. [Google Scholar] [CrossRef] [Green Version]
- Ferro, M.D.; Santos, S.A.O.; Silvestre, A.J.D.; Duarte, M.F. Chromatographic separation of phenolic compounds from extra virgin olive oil: Development and validation of a new method based on a biphenyl HPLC column. Int. J. Mol. Sci. 2019, 20, 201. [Google Scholar] [CrossRef] [Green Version]
- Peres, M.F.; Henriques, L.R.; Simões-Lopes, P.; Pinheiro-Alves, M.C. Azeites da ‘Galega Vulgar’—efeito do loteamento e do armazenamento. In Atas Portuguesas de Horticultura 13, Proceedings of the 3rd Simpósio Nacional de Olivicultura, Castelo Branco, Porttugal, 29–31 October 2003; Associação Portuguesa de Horticultura: Lisboa, Portugal, 2009; pp. 186–191. [Google Scholar]
- Marcelo, M.E.; Lopes, J.I.; Soares, F.M.; Centeno, M.S.L.; Cordeiro, A.M.; Vasconcelos, P.M. Effect of fertilization on polyphenols, sterols and waxes contents of the olive oil from six cultivars in Portugal. In Atas Portuguesas de Horticultura 14, Proceedings of the 5th Simpósio Nacional de Olivicultura, Santarém, Portugal, 24–26 September 2009; Associação Portuguesa de Horticultura: Lisboa, Portugal, 2011; pp. 141–150. [Google Scholar]
- Pita, D.; Vitorino, M.C.; Gouveia, C.; Peres, F. Extraction of monovarietal olive oils with natural microtalc. In Atas Portuguesas de Horticultura 14, Proceedings of the 5th Simpósio Nacional de Olivicultura, Santarém, Portugal, 24–26 September 2009; Associação Portuguesa de Horticultura: Lisboa, Portugal, 2011; pp. 151–157. [Google Scholar]
- Quintero-Flórez, A.; Nieva, L.S.; Sánchez-Ortíz, A.; Beltrán, G.; Perona, J.S. The fatty acid composition of virgin olive oil from different cultivars is determinant for foam cell formation by macrophages. J. Agric. Food Chem. 2015, 63, 6731–6738. [Google Scholar] [CrossRef]
- Gouvinhas, I.; Domínguez-Perles, R.; Machado, N.; Carvalho, T.; Matos, C.; Barros, A.I.R.N.A. Effect of agro-environmental factors on the mineral content of olive oils: Categorization of the three major Portuguese cultivars. J. Am. Oil Chem. Soc. 2016, 93, 813–822. [Google Scholar] [CrossRef]
- Pires-Cabral, P.; Barros, T.; Nunes, P.; Quintas, C. Physicochemical, nutritional and microbiological characteristics of traditional table olives from Southern Portugal. Emir. J. Food Agric. 2018, 30, 611–620. [Google Scholar] [CrossRef]
- Saramago, I.S.L. Olival em Modo de Produção Biológico. Master’s Thesis, Escola Superior Agrária, Instituto Politécnico de Beja, Beja, Portugal, 2009. Available online: http://hdl.handle.net/10400.26/3979 (accessed on 15 September 2020).
- Tous, J.; Romero, A.; Hermoso, J.F.; Mallén, N. Sistemas de producción del olivo en seto. Experiencias en Cataluña. Agricultura 2007, 896, 360–367. [Google Scholar]
- Guerrero Maldonado, N.; López, M.J.; Caudullo, G.; de Rigo, D. Olea europaea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publ. Off. EU: Luxembourg, 2016; p. e01534b+. [Google Scholar]
- Beghè, D.; Ferrarini, A.; Ganino, T.; Fabbri, A. Molecular characterization and identification of a group of local Olea europaea L. varieties. Tree Genet. Genomes 2011, 7, 1185–1198. [Google Scholar] [CrossRef]
- Marra, F.P.; Caruso, T.; Costa, F.; Di Vaio, C.; Mafrica, R.; Marchese, A. Genetic relationships, structure and parentage simulation among the olive tree (Olea europaea L. subsp. europaea) cultivated in Southern Italy revealed by SSR markers. Tree Genet. Genomes 2013, 9, 961–973. [Google Scholar] [CrossRef]
- Belaj, A.; Veral, M.G.; Sikaoui, H.; Moukhli, A.; Khadari, B.; Mariotti, R.; Baldoni, L. Olive genetic resources. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 27–54. [Google Scholar] [CrossRef]
- Lavee, S. Evaluation of the need and present potential of olive breeding indicating the nature of the available genetic resources involved. Sci. Hortic. 2013, 161, 333–339. [Google Scholar] [CrossRef]
- Belaj, A.; Muñoz-Diez, C.; Baldoni, L.; Šatović, Z.; Barranco, D. Genetic diversity and relationships of wild and cultivated olives at regional level in Spain. Sci. Hortic. 2010, 124, 323–330. [Google Scholar] [CrossRef]
- Maxted, N.; Guarino, L. Genetic erosion and genetic pollution of crop wild relatives. In Proceedings of the PGR Forum Workshop 5, Genetic Erosion and Pollution Assessment Methodologies, Terceira Island, Autonomous Region of the Azores, Portugal, 8–11 September 2004; pp. 35–45. [Google Scholar]
- Brown, A.H.D.; Hodgkin, T. Indicators of Genetic Diversity, Genetic Erosion, and Genetic Vulnerability for Plant Genetic Resources. In Genetic Diversity and Erosion in Plants. Sustainable Development and Biodiversity; Ahuja, M., Jain, S., Eds.; Springer: Cham, Switerzland, 2015; Volume 7, pp. 25–53. [Google Scholar] [CrossRef]
- Salomone, R.; Cappelletti, G.M.; Malandrino, O.; Mistretta, M.; Neri, E.; Nicoletti, G.; Notarnicola, B.; Pattara, C.; Russo, C.; Saija, G. Life cycle assessment in the olive oil sector. In Life Cycle Assessment in the Agri-Food Sector; Notarnicola, B., Salomone, R., Petti, L., Renzulli, P., Roma, R., Cerutti, A., Eds.; Springer: Cham, Switzerland, 2015; pp. 57–121. [Google Scholar] [CrossRef]
- Espadas-Aldana, G.; Vialle, C.; Belaud, J.P.; Vaca-Garcia, C.; Sablayrolles, C. Analysis and trends for Life Cycle Assessment of olive oil production. Sustain. Prod. Consum. 2019, 19, 216–230. [Google Scholar] [CrossRef] [Green Version]
- Salomone, R.; Ioppolo, G. Environmental impacts of olive oil production: A life cycle assessment case study in the province of Messina (Sicily). J. Clean. Prod. 2012, 28, 88–100. [Google Scholar] [CrossRef]
- Rajaeifar, M.A.; Akram, A.; Ghobadian, B.; Rafiee, S.; Heidari, M.D. Energy economic life cycle assessment (LCA) and greenhouse gas emissions analysis of olive oil production in Iran. Energy 2014, 66, 139–149. [Google Scholar] [CrossRef]
- Proietti, S.; Sdringola, P.; Regni, L.; Evangelisti, N.; Brunori, A.; Ilarioni, L.; Nasini, L.; Proietti, P. Extra virgin olive oil as carbon negative product: Experimental analysis and validation of results. J. Clean. Prod. 2017, 166, 550–562. [Google Scholar] [CrossRef]
- Tsarouhas, P.; Achillas, C.; Aidonis, D.; Folinas, D.; Maslis, V. Life cycle assessment of olive oil production in Greece. J. Clean. Prod. 2015, 93, 75–83. [Google Scholar] [CrossRef]
- El Hanandeh, A.; Gharaibeh, M.A. Environmental efficiency of olive oil production by small and micro-scale farmers in northern Jordan: Life cycle assessment. Agric. Syst. 2016, 148, 169–177. [Google Scholar] [CrossRef]
- Pattara, C.; Salomone, R.; Cichelli, A. Carbon footprint of extra virgin olive oil: A comparative and driver analysis of different production processes in Centre Italy. J. Clean. Prod. 2016, 127, 533–547. [Google Scholar] [CrossRef]
- Accorsi, R.; Cascini, A.; Ferrari, E.; Manzini, R.; Pareschi, A.; Versari, L. Life cycle assessment of an extra-virgin olive oil supply chain. In Proceedings of the 18th Summer School “Francesco Turco”—Industrial Mechanical Plants, Senigallia, Italy, 11–13 September 2013; pp. 172–178. [Google Scholar]
- Guiso, A.; Parenti, A.; Masella, P.; Guerrini, L.; Baldi, F.; Spugnoli, P. Environmental impact assessment of three packages for high-quality extra-virgin olive oil. Agric. Eng. 2016, 47, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.; Puig, R.; Martí, E.; Bala, A.; Fullana-i Palmer, P. Tackling the relevance of packaging in life cycle assessment of virgin olive oil and the environmental consequences of regulation. Environ. Manag. 2018, 62, 277–294. [Google Scholar] [CrossRef]
- Chatzisymeon, E.; Foteinis, S.; Mantzavinos, D.; Tsoutsos, T. Life cycle assessment of advanced oxidation processes for olive mill wastewater treatment. J. Clean. Prod. 2013, 54, 229–234. [Google Scholar] [CrossRef] [Green Version]
- El Hanandeh, A. Energy recovery alternatives for the sustainable management of olive oil industry waste in Australia: Life cycle assessment. J. Clean. Prod. 2015, 91, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Christoforou, E.A.; Fokaides, P.A. Life cycle assessment (LCA) of olive husk torrefaction. Renew. Energy 2016, 90, 257–266. [Google Scholar] [CrossRef]
- Parascanu, M.M.; Gamero, M.P.; Sánchez, P.; Soreanu, G.; Valverde, J.L.; Sanchez-Silva, L. Life cycle assessment of olive pomace valorisation through pyrolysis. Renew. Energy 2018, 122, 589–601. [Google Scholar] [CrossRef]
- Parascanu, M.M.; Sánchez, P.; Soreanu, G.; Valverde, J.L.; Sanchez-Silva, L. Environmental assessment of olive pomace valorization through two different thermochemical processes for energy production. J. Clean. Prod. 2018, 186, 771–781. [Google Scholar] [CrossRef]
- Mohamad, R.S.; Verrastro, V.; Cardone, G.; Bteich, M.R.; Favia, M.; Moretti, M.; Roma, R. Optimization of organic and conventional olive agricultural practices from a life cycle assessment and life cycle costing perspectives. J. Clean. Prod. 2014, 70, 78–89. [Google Scholar] [CrossRef]
- De Luca, A.I.; Falcone, G.; Stillitano, T.; Iofrida, N.; Strano, A.; Gulisano, G. Evaluation of sustainable innovations in olive growing systems: A life cycle sustainability assessment case study in southern Italy. J. Clean. Prod. 2018, 171, 1187–1202. [Google Scholar] [CrossRef]
- Romero-Gámez, M.; Castro-Rodríguez, J.; Suárez-Rey, E.M. Optimization of olive growing practices in Spain from a life cycle assessment perspective. J. Clean. Prod. 2017, 149, 25–37. [Google Scholar] [CrossRef]
- Bernardi, B.; Falcone, G.; Stillitano, T.; Benalia, S.; Strano, A.; Bacenetti, J.; De Luca, A.I. Harvesting system sustainability in Mediterranean olive cultivation. Sci. Total Environ. 2018, 625, 1446–1458. [Google Scholar] [CrossRef]
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidences from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Terral, J.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef] [Green Version]
- Díez, C.M.; Moral, J.; Barranco, D.; Rallo, L. Genetic diversity and conservation of olive genetic resources. In Genetic Diversity and Erosion in Plants. Sustainable Development and Biodiversity; Ahuja, M., Jain, S., Eds.; Springer: Cham, Switerzland, 2015; Volume 8, pp. 337–356. [Google Scholar] [CrossRef]
- Besnard, G.; El Bakkali, A.; Haouane, H.; Baali-Cherif, D.; Moukhli, A.; Khadari, B. Population genetics of Mediterranean and Saharan olives: Geographic patterns of differentiation and evidence for early-generations of admixture. Ann. Bot. 2013, 112, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Díez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Besnard, G.; Rubio de Casas, R.; Christin, P.A.; Vargas, P. Phylogenetics of Olea (Oleaceae) based on plastid and nuclear ribosomal DNA sequences: Tertiary climatic shifts and lineage differentiation times. Ann. Bot. 2009, 104, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Ladisa, G.; Calabrese, E.; Perrino, E.V. The origin and distribution of olive tree and olive crop. In Study on Biodiversity in Century-Old Olive Groves; Calabrese, G., Tartaglini, N., Ladisa, G., Eds.; CIHEAM—Mediterranean Agronomic Institute of Bari: Bari, Italy, 2012; pp. 1–3. [Google Scholar]
- Fuller, D.Q. Long and attenuated: Comparative trends in the domestication of tree fruits. Veg. Hist. Archaeobotany 2018, 27, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belaj, A.; Dominguez-García, M.C.; Atienza, S.G.; Urdíroz, N.M.; De la Rosa, R.; Šatović, Z.; Martín, A.; Kilian, A.; Trujillo, I.; Valpuesta, V.; et al. Developing a core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 2012, 8, 365–378. [Google Scholar] [CrossRef]
- Fabbri, A.; Lambardi, M.; Ozden-Tokatli, Y. Olive Breeding. In Breeding Plantation Tree Crops: Tropical Species; Jain, S.M., Priyadarshan, P.M., Eds.; Springer: New York, NY, USA, 2009; pp. 423–465. ISBN 978-038-771-199-7. [Google Scholar]
- Doveri, S.; Baldoni, L. Olive. In Genome Mapping and Molecular Breeding in Plants—Fruits and Nuts; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 4, pp. 253–256. ISBN 978-354-034-531-2. [Google Scholar]
- Alves, M.L.V. Caracterização e Estrutura Genéticas da Cultivar de Oliveira ‘Cobrançosa’ e Sua Relação com o Zambujeiro. Master’s Thesis, University of Lisboa, Lisboa, Portugal, 2007. Available online: http://hdl.handle.net/10451/1358 (accessed on 30 June 2020).
- Belaj, A.; León, L.; Šatović, Z.; De la Rosa, R. Variability of wild olives (Olea europaea subsp. europaea var. sylvestris) analyzed by agro-morphological traits and SSR markers. Sci. Hortic. 2011, 129, 561–569. [Google Scholar] [CrossRef]
- Bouhadida, M.; Casas, A.M.; Moreno, M.A.; Gogorcena, Y. Molecular characterization of Miraflores peach variety and relatives using SSRs. Sci. Hortic. 2007, 111, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Wünsch, A.; Hormaza, J.I. Cultivar identification and genetic fingerprinting of temperate fruit tree species using DNA markers. Euphytica 2002, 125, 59–67. [Google Scholar] [CrossRef]
- Hammer, K.; Arrowsmith, N.; Gladis, T. Agrobiodiversity with emphasis on plant genetic resources. Naturwissenschaften 2003, 90, 241–250. [Google Scholar] [CrossRef]
- Pasqualone, A.; Montemurro, C.; Di Rienzo, V.; Summo, C.; Paradiso, V.M.; Caponio, F. Evolution and perspectives of cultivar identification and traceability from tree to oil and table olives by means of DNA markers. J. Sci. Food Agric. 2016, 96, 3642–3657. [Google Scholar] [CrossRef]
- Lazović, B.; Bosković, R.; James, C.; Tobutt, K.R.; Gasic, K. Genetic diversity of olives grown along the coast of Montenegro. Acta Hortic. 2002, 586, 167–170. [Google Scholar] [CrossRef]
- Besnard, G.; Baradat, P.; Chevalier, D.; Tagmount, A.; Bervillé, A. Genetic differentiation in the olive complex (Olea europaea L.) revealed by RAPDs and RFLPs in the rRNA genes. Genet. Resour. Crop Evol. 2001, 48, 165–182. [Google Scholar] [CrossRef]
- De la Rosa, R.; Angiolillo, A.; Rallo, L.; Guerrero, C.; Pellegrini, M.; Besnard, G.; Bervillé, A.; Martin, A.; Baldoni, L. A first linkage map of olive (Olea europaea L.) cultivars using RAPD, AFLP, RFLP and SSR markers. Theor. Appl. Genet. 2003, 106, 1273–1282. [Google Scholar] [CrossRef]
- Belaj, A.; Šatović, Z.; Trujillo, I.; Rallo, L. Genetic relationships of Spanish olive cultivars using RAPD markers. HortScience 2004, 39, 948–951. [Google Scholar] [CrossRef]
- Brake, M.; Migdadi, H.; Al-Gharaibeh, M.; Ayoub, S.; Haddad, N.; El Oqlah, A. Characterization of Jordanian olive cultivars (Olea europaea L.) using RAPD and ISSR molecular markers. Sci. Hortic. 2014, 176, 282–289. [Google Scholar] [CrossRef]
- Linos, A.; Nikoloudakis, N.; Katsiotis, A.; Hagidimitriou, M. Genetic structure of the Greek olive germplasm revealed by RAPD, ISSR and SSR markers. Sci. Hortic. 2014, 175, 33–43. [Google Scholar] [CrossRef]
- Besnard, G.; Bervillé, A. Multiple origins for Mediterranean olive (Olea europaea L. ssp europaea) based upon mitochondrial DNA polymorphisms. Comptes Rendus Acad. Sci. 2000, 323, 173–181. [Google Scholar] [CrossRef]
- Wu, S.B.; Collins, G.; Sedgley, M. A molecular linkage map of olive (Olea europaea L.) based on RAPD, microsatellite, and SCAR markers. Genome 2004, 47, 26–35. [Google Scholar] [CrossRef]
- Charafi, J.; Zine El Aabidine, A.; Grout, C.; Rahioui, B.; El Meziane, A.; Moukhli, A.; Boulouha, B.; El Modafar, C.; Khadari, B. A genetic linkage map of Olea europaea L. using a pseudo-test cross- mapping strategy based on SSR, AFLP, ISSR, RAPD and SCAR markers. Acta Hortic. 2009, 814, 609–614. [Google Scholar] [CrossRef]
- Terzopoulos, P.J.; Kolano, B.; Bebeli, P.J.; Kaltsikes, P.J.; Metzidakis, I. Identification of Olea europaea L. cultivars using inter-simple sequence repeat markers. Sci. Hortic. 2005, 105, 45–51. [Google Scholar] [CrossRef]
- Essadki, M.; Ouazzani, N.; Lumaret, R.; Moumni, M. ISSR variation in olive-tree cultivars from Morocco and other western countries of the Mediterranean basin. Genet. Resour. Crop Evol. 2006, 53, 475–482. [Google Scholar] [CrossRef]
- Owen, C.A.; Bita, E.C.; Banilas, G.; Hajjar, S.E.; Sellianakis, V.; Aksoy, U.; Hepaksoy, S.; Chamoun, R.; Talhook, S.N.; Metzidakis, I.; et al. AFLP reveals structural details of genetic diversity within cultivated olive germplasm from the Eastern Mediterranean. Theor. Appl. Genet. 2005, 110, 1169–1176. [Google Scholar] [CrossRef]
- Ercisli, S.; Barut, E.; Ipek, A. Molecular characterization of olive cultivars using amplified fragment length polymorphism markers. Genet. Mol. Res. 2009, 8, 414–419. [Google Scholar] [CrossRef]
- Ipek, M.; Seker, M.; Ipek, A.; Gul, M.K. Identification of molecular markers associated with fruit traits in olive and assessment of olive core collection with AFLP markers and fruit traits. Genet. Mol. Res. 2015, 14, 2762–2774. [Google Scholar] [CrossRef] [PubMed]
- El Aabidine, A.Z.; Charafi, J.; Grout, C.; Doligez, A.; Santoni, S.; Moukhli, A.; Jay-Allemand, C.; El Modafar, C.; Khadari, B. Construction of a genetic linkage map for the olive based on AFLP and SSR markers. Crop Sci. 2010, 50, 2291–2302. [Google Scholar] [CrossRef]
- Khadari, B.; El Aabidine, A.Z.; Grout, C.; Ben Sadok, I.; Doligez, A.; Moutier, N.; Santoni, S.; Costes, E. A genetic linkage map of olive based on amplified fragment length polymorphism, intersimple sequence repeat and simple sequence repeat markers. J. Am. Soc. Hortic. Sci. 2010, 135, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ayed, R.; Ennouri, K.; Ben-Hassen, H.; Triki, M.A.; Rebaï, A. Comparison between DNA-based, pomological and chemical markers accomplished by bioinformatic tools to distinguish within Tunisian olive cultivars. J. Fundam. Appl. Sci. 2015, 7, 408–421. [Google Scholar] [CrossRef]
- Morgante, M.; Olivieri, A.M. PCR-amplified microsatellites as markers in plant genetics. Plant J. 1993, 3, 175–182. [Google Scholar] [CrossRef]
- De la Rosa, R.; James, C.M.; Tobutt, K.R. Isolation and characterization of polymorphic microsatellites in olive (Olea europaea L.) and their transferability to other genera in the Oleaceae. Mol. Ecol. Notes 2002, 2, 265–267. [Google Scholar] [CrossRef]
- Rallo, P.; Tenzer, I.; Gessler, C.; Baldoni, L.; Dorado, G.; Martín, A. Transferability of olive microsatellite loci across the genus Olea. Theor. Appl. Genet. 2003, 107, 940–946. [Google Scholar] [CrossRef]
- Belaj, A.; Cipriani, G.; Testolin, R.; Rallo, L.; Trujillo, I. Characterization and identification of the main Spanish and Italian olive cultivars by simple-sequence-repeat markers. HortScience 2004, 39, 1557–1561. [Google Scholar] [CrossRef]
- Salimonti, A.; Simeone, V.; Cesari, G.; Lamaj, F.; Cattivelli, L.; Perri, E.; Desiderio, F.; Fanizzi, F.P.; Del Coco, L.; Zelasco, S. A first molecular investigation of monumental olive trees in Apulia region. Sci. Hortic. 2013, 162, 204–212. [Google Scholar] [CrossRef]
- Xu, Y. Molecular Plant Breeding; CAB International: Oxfordshire, UK, 2010; pp. 195–247. ISBN 978-184-593-392-0. [Google Scholar]
- Reale, S.; Doveri, S.; Díaz, A.; Angiolillo, A.; Lucentini, L.; Pilla, F.; Martín, A.; Donini, P.; Lee, D. SNP-based markers for discriminating olive (Olea europaea L.) cultivars. Genome 2006, 49, 1193–1205. [Google Scholar] [CrossRef]
- Carriero, F.; Fontanazza, G.; Cellini, F.; Giorio, G. Identification of simple sequence repeats (SSRs) in olive (Olea europaea L.). Theor. Appl. Genet. 2002, 104, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Baldoni, L.; Cultrera, N.G.; Mariotti, R.; Riccioloni, C.; Arcioni, S.; Vendramin, G.G.; Buonamici, A.; Porceddu, A.; Sarri, V.; Ojeda, M.A.; et al. A consensus list of microsatellite markers for olive genotyping. Mol. Breed. 2009, 24, 213–231. [Google Scholar] [CrossRef]
- Belaj, A.; Muñoz-Diez, C.; Baldoni, L.; Porceddu, A.; Barranco, D.; Šatović, Z. Genetic diversity and population structure of wild olives from the North-western Mediterranean by SSR markers. Ann. Bot. 2007, 100, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipek, A.; Barut, E.; Gulen, H.; Ipek, M. Assessment of inter-and intra-cultivar variations in olive using SSR markers. Sci. Agric. 2012, 69, 327–335. [Google Scholar] [CrossRef]
- Caruso, T.; Marra, F.P.; Costa, F.; Campisi, G.; Macaluso, L.; Marchese, A. Genetic diversity and clonal variation within the main Sicilian olive cultivars based on morphological traits and microsatellite markers. Sci. Hortic. 2014, 180, 130–138. [Google Scholar] [CrossRef]
- Abdessemed, S.; Muzzalupo, I.; Benbouza, H. Assessment of genetic diversity among Algerian olive (Olea europaea L.) cultivars using SSR marker. Sci. Hortic. 2015, 192, 10–20. [Google Scholar] [CrossRef]
- Lazović, B.; Klepo, T.; Adakalić, M.; Šatović, Z.; Arbeiter, A.B.; Hladnik, M.; Strikić, F.; Liber, Z.; Bandelj, D. Intra-varietal variability and genetic relationships among the homonymic East Adriatic olive (Olea europaea L.) varieties. Sci. Hortic. 2018, 236, 175–185. [Google Scholar] [CrossRef]
- Ben Sadok, I.; Celton, J.M.; Essalouh, L.; El Aabidine, A.Z.; Garcia, G.; Martinez, S.; Grati-Kamoun, N.; Rebai, A.; Costes, E.; Khadari, B. QTL mapping of flowering and fruiting traits in olive. PLoS ONE 2013, 8, e62831. [Google Scholar] [CrossRef] [Green Version]
- Mookerjee, S.; Guerin, J.; Collins, G.; Ford, C.; Sedgley, M. Paternity analysis using microsatellite markers to identify pollen donors in an olive grove. Theor. Appl. Genet. 2005, 111, 1174–1182. [Google Scholar] [CrossRef]
- De la Rosa, R.; Belaj, A.; Muñoz-Mérida, A.; Trelles, O.; Ortíz-Martín, I.; González-Plaza, J.J.; Valpuesta, V.; Beuzón, C.R. Development of EST-derived SSR markers with long-core repeat in olive and their use for paternity testing. J. Am. Soc. Hortic. Sci. 2013, 138, 290–296. [Google Scholar] [CrossRef]
- Hakim, I.R.; Grati-Kammoun, N.; Makhloufi, E.; Rebaï, A. Discovery and potential of SNP markers in characterization of Tunisian olive germplasm. Diversity 2010, 2, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-García, M.C.; Laib, M.; De La Rosa, R.; Belaj, A. Characterisation and identification of olive cultivars from North-eastern Algeria using molecular markers. J. Hortic. Sci. Biotechnol. 2012, 87, 95–100. [Google Scholar] [CrossRef]
- Biton, I.; Doron-Faigenboim, A.; Jamwal, M.; Mani, Y.; Eshed, R.; Rosen, A.; Sherman, A.; Ophir, R.; Lavee, S.; Avidan, B.; et al. Development of a large set of SNP markers for assessing phylogenetic relationships between the olive cultivars composing the Israeli olive germplasm collection. Mol. Breed. 2015, 35, 107–120. [Google Scholar] [CrossRef]
- Zhu, S.; Niu, E.; Shi, A.; Mou, B. Genetic diversity analysis of olive germplasm (Olea europaea L.) with Genotyping-by-Sequencing Technology. Front. Genet. 2019, 10, 755. [Google Scholar] [CrossRef] [Green Version]
- Baldoni, L.; Khadari, B.; De la Rosa, R. Genetic mapping and detection of Quantitative Trait Loci. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 65–74. [Google Scholar] [CrossRef]
- İpek, A.; Yılmaz, K.; Sıkıcı, P.; Tangu, N.A.; Öz, A.T.; Bayraktar, M.; İpek, M.; Gülen, H. SNP discovery by GBS in olive and the construction of a high-density genetic linkage map. Biochem. Genet. 2016, 54, 313–325. [Google Scholar] [CrossRef]
- Marchese, A.; Marra, F.P.; Caruso, T.; Mhelembe, K.; Costa, F.; Fretto, S.; Sargent, D.J. The first high-density sequence characterized SNP-based linkage map of olive (Olea europaea L. subsp. europaea) developed using genotyping by sequencing. Aust. J. Crop Sci. 2016, 10, 857. [Google Scholar] [CrossRef]
- El Bakkali, A.; Essalouh, L.; Tollon, C.; Rivallan, R.; Mournet, P.; Moukhli, A.; Zaher, H.; Mekkaoui, A.; Hadidou, A.; Sikaoui, L.; et al. Characterization of worldwide olive germplasm banks of Marrakech (Morocco) and Córdoba (Spain): Towards management and use of olive germplasm in breeding programs. PLoS ONE 2019, 14, e0223716. [Google Scholar] [CrossRef] [Green Version]
- FAO. International Treaty on Plant Genetic Resources for Food and Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; Available online: http://www.fao.org/plant-treaty/overview/en/ (accessed on 15 July 2020).
- Sardaro, R.; Bozzo, F.; Petrontino, A.; Fucilli, V. Community preferences in support of a conservation programme for olive landraces in the Mediterranean area. Acta Hortic. 2018, 1199, 183–188. [Google Scholar] [CrossRef]
- Rallo, L.; Barranco, D.; Díez, C.M.; Rallo, P.; Suárez, M.P.; Trapero, C.; Pliego-Alfaro, F. Strategies for olive (Olea europaea L.) breeding: Cultivated genetic resources and crossbreeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2018; Volume 3, pp. 536–600. [Google Scholar]
- Baenziger, P.S.; Russell, W.K.; Graef, G.L.; Campbell, B.T. Improving lives: 50 years of crop breeding, genetics, and cytology (C-1). Crop Sci. 2006, 46, 2230–2244. [Google Scholar] [CrossRef]
- Rallo, L.; Caruso, T.; Díez, C.M.; Campisi, G. Olive growing in a time of change: From empiricism to genomics. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 55–64. [Google Scholar] [CrossRef]
- Baldoni, L.; Belaj, A. Olive. In Oil Crops. Handbook of Plant Breeding; Vollmann, J., Rajcan, I., Eds.; Springer: New York, NY, USA, 2009; Volume 4, pp. 397–421. [Google Scholar] [CrossRef]
- Rallo, L. Breeding oil and table olives for mechanical harvesting in Spain. Hortech 2014, 24, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Arias-Calderón, R.; Rodríguez-Jurado, D.; León, L.; Bejarano-Alcázar, J.; De la Rosa, R.; Belaj, A. Pre-breeding for resistance to Verticillium wilt in olive: Fishing in the wild relative gene pool. Crop Prot. 2015, 75, 25–33. [Google Scholar] [CrossRef]
- Trapero, C.; Rallo, L.; López-Escudero, F.J.; Barranco, D.; Díez, C.M. Variability and selection of verticillium wilt resistant genotypes in cultivated olive and in the Olea genus. Plant Pathol. 2015, 64, 890–900. [Google Scholar] [CrossRef]
- Klepo, T.; Toumi, A.; De la Rosa, R.; León, L.; Belaj, A. Agronomic evaluation of seedlings from crosses between the main Spanish olive cultivar ‘Picual’ and two wild olive trees. J. Hortic. Sci. Biotechnol. 2014, 89, 508–512. [Google Scholar] [CrossRef]
- Caceres, M.E.; Ceccarelli, M.; Pupilli, F.; Sarri, V.; Mencuccini, M. Obtainment of inter-subspecific hybribs in olive (Olea europaea L.). Euphytica 2015, 201, 307–319. [Google Scholar] [CrossRef]
- Rugini, E.; Cristofori, V.; Silvestri, C. Genetic improvement of olive (Olea europaea L.) by conventional and in vitro biotechnology methods. Biotechnol. Adv. 2016, 34, 687–696. [Google Scholar] [CrossRef]
- Rugini, E.; De Pace, C. Olive breeding with classical and modern approaches. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 163–193. [Google Scholar] [CrossRef]
- Leva, A.R. Innovative protocol for “ex vitro rooting” on olive micropropagation. Cent. Eur. J. Biol. 2011, 6, 352–358. [Google Scholar] [CrossRef]
- Benelli, C.; De Carlo, A.; Englmen, F. Recent advances in the cryopreservation of shoot-derived germplasm of economically important fruit trees of Actinidia, Diospyros, Malus, Olea, Prunus, Pyrus and Vitis. Biotechnol. Adv. 2013, 31, 175–185. [Google Scholar] [CrossRef]
- Rugini, E.; Silvestri, C.; Ceccarelli, M.; Muleo, R.; Cristofori, V. Mutagenesis and biotechnology techniques as tools for selecting new stable diploid and tetraploid olive genotypes and their dwarfing agronomical characterization. HortScience 2016, 51, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Capelo, A.M.; Silva, S.; Brito, G.; Santos, C. Somatic embryogenesis induction in leaves and petioles of a mature wild olive. Plant Cell Tissue Org. 2010, 103, 237–242. [Google Scholar] [CrossRef]
- Toufik, I.; Guenoun, F.; Belkoura, I. Embryogenesis expression from somatic explants of olive (Olea europaea L.) cv Picual. Moroc. J. Biol. 2014, 11, 17–25. [Google Scholar]
- Torreblanca, R.; Cerezo, S.; Palomo-Ríos, E.; Mercado, J.A.; Pliego-Alfaro, F. Development of a high throughput system for genetic transformation of olive (Olea europaea L.) plants. Plant Cell Tissue Org. 2010, 103, 61–69. [Google Scholar] [CrossRef]
- Titouh, K.; Khelifi, L.; Moussa, K.T.; Cerezo-Medina, S.; Mercado, J.A.; Pliego-Alfaro, F. Evaluation of the effect of phosphinothricin, as selection agent, on the growth of olive somatic embryos. Acta Hortic. 2014, 1057, 533–542. [Google Scholar] [CrossRef]
- Ozgenturk, N.O.; Oruç, F.; Sezerman, U.; Kuçukural, A.; Korkut, S.V.; Toksoz, F.; Un, C. Generation and analysis of expressed sequence tags from Olea europaea L. Comp. Funct. Genom. 2010, 2010, 757512. [Google Scholar] [CrossRef]
- Bazakos, C.; Manioudaki, M.E.; Therios, I.; Voyiatzis, D.; Kafetzopoulos, D.; Awada, T.; Kalaitzis, P. Comparative transcriptome analysis of two olive cultivars in response to NaCl-stress. PLoS ONE 2012, 7, e42931. [Google Scholar] [CrossRef] [PubMed]
- Parra, R.; Paredes, M.A.; Sanchez-Calle, I.M.; Gomez-Jimenez, M.C. Comparative transcriptional profiling analysis of olive ripe-fruit pericarp and abscission zone tissues shows expression differences and distinct patterns of transcriptional regulation. BMC Genom. 2013, 14, 866. [Google Scholar] [CrossRef] [Green Version]
- Yanik, H.; Turktas, M.; Dundar, E.; Hernandez, P.; Dorado, G.; Unver, T. Genome-wide identification of alternate bearing-associated microRNAs (miRNAs) in olive (Olea europaea L.). BMC Plant Biol. 2013, 13, 10. [Google Scholar] [CrossRef] [Green Version]
- Cabanás, C.G.; Schilirò, E.; Valverde-Corredor, A.; Mercado-Blanco, J. Systemic responses in a tolerant olive (Olea europaea L.) cultivar upon root colonization by the vascular pathogen Verticillium dahliae. Front. Microbiol. 2015, 6, 928. [Google Scholar] [CrossRef] [Green Version]
- Carmona, R.; Zafra, A.; Seoane, P.; Castro, A.J.; Guerrero-Fernández, D.; Castillo-Castillo, T.; Medina-García, A.; Cánovas, F.M.; Aldana-Montes, J.F.; Navas-Delgado, I.; et al. ReprOlive: A database with linked data for the olive tree (Olea europaea L.) reproductive transcriptome. Front. Plant Sci. 2015, 6, 625. [Google Scholar] [CrossRef] [Green Version]
- Leyva-Pérez, M.O.; Valverde-Corredor, A.; Valderrama, R.; Jiménez-Ruiz, J.; Muñoz-Merida, A.; Trelles, O.; Barroso, J.B.; Mercado-Blanco, J.; Luque, F. Early and delayed long-term transcriptional changes and short-term transient responses during cold acclimation in olive leaves. DNA Res. 2015, 22, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Giampetruzzi, A.; Morelli, M.; Saponari, M.; Loconsole, G.; Chiumenti, M.; Boscia, D.; Saldarelli, P. Transcriptome profiling of two olive cultivars in response to infection by the OQDS strain of Xylella fastidiosa ssp. pauca. BMC Genom. 2016, 17, 1. [Google Scholar] [CrossRef] [Green Version]
- González-Plaza, J.J.; Ortiz-Martín, I.; Muñoz-Mérida, A.; García-López, C.; Sánchez-Sevilla, J.F.; Luque, F.; Trelles, O.; Bejarano, E.R.; De la Rosa, R.; Valpuesta, V.; et al. Transcriptomic analysis using olive varieties and breeding progenies identifies candidate genes involved in plant architecture. Front. Plant Sci. 2016, 7, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarah, G.; Homa, F.; Pointet, S.; Contreras, S.; Sabot, F.; Nabholz, B.; Santoni, S.; Sauné, L.; Ardisson, M.; Chantret, N.; et al. A large set of 26 new reference transcriptomes dedicated to comparative population genomics in crops and wild relatives. Mol. Ecol. Resour. 2017, 17, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Zafra, A.; Carmona, R.; Traverso, J.A.; Hancock, J.T.; Goldman, M.H.S.; Claros, M.G.; Hiscock, S.J.; Alche, J.D. Identification and functional annotation of genes differentially expressed in the reproductive tissues of the olive tree (Olea europaea L.) through the generation of subtractive libraries. Front. Plant Sci. 2017, 8, 1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guodong, R.; Jianguo, Z.; Xiaoxia, L.; Ying, L. Identification of putative genes for polyphenol biosynthesis in olive fruits and leaves using full-length transcriptome sequencing. Food Chem. 2019, 300, 125246. [Google Scholar] [CrossRef]
- Besnard, G.; Hernández, P.; Khadari, B.; Dorado, G.; Savolainen, V. Genomic profiling of plastid DNA variation in the Mediterranean olive tree. BMC Plant Biol. 2011, 11, 80. [Google Scholar] [CrossRef] [Green Version]
- Kaya, E.; Vatansever, R.; Filiz, E. Assessment of the genetic relationship of Turkish olives (Olea europaea subsp. europaea) cultivars based on cpDNA trnL-F regions. Acta Bot. Croat. 2018, 77, 88–92. [Google Scholar] [CrossRef]
- Rugini, E.; De Pace, C.; Gutiérrez-Pesce, P.; Muleo, R. Olea. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 79–144. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.B.; Cetin, O.; Kaya, H.S.; Sahin, M.; Sefer, F.; Tanyolac, B. Association mapping in Turkish olive cultivars revealed significant markers related to some important agronomic traits. Biochem. Genet. 2016, 54, 506–533. [Google Scholar] [CrossRef]
- Atienza, S.G.; De La Rosa, R.; León, L.; Martín, A.; Belaj, A. Identification of QTL for agronomic traits of importance for olive breeding. Mol. Breed. 2014, 34, 725–737. [Google Scholar] [CrossRef]
- Ates, D. Identification of QTLs controlling genes of ripening time, flesh detachment from stone and firmness in olive (Olea europaea L.). In Proceedings of the 24th Plant and Animal Genome Conference, San Diego, CA, USA, 9–13 January 2016; p. P1163. [Google Scholar]
- Mauricio, R. Mapping quantitative trait loci in plants: Uses and caveats for evolutionary biology. Nat. Rev. Genet. 2001, 2, 370–381. [Google Scholar] [CrossRef]
- Morrell, P.L.; Buckler, E.S.; Ross-Ibarra, J. Crop genomics: Advances and applications. Nat. Rev. Genet. 2012, 13, 85–96. [Google Scholar] [CrossRef]
- Yu, J.; Buckler, E.S. Genetic association mapping and genome organization of maize. Curr. Opin. Biotechnol. 2006, 17, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.B.; Akdemir, D.; Lozano, R.; Cetin, O.; Kaya, H.S.; Sahin, M.; Smith, J.L.; Tanyolac, B.; Jannink, J.L. Genome wide association study of 5 agronomic traits in olive (Olea europaea L.). Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Salimonti, A.; Carbone, F.; Romano, E.; Pellegrino, M.; Benincasa, C.; Micali, S.; Tondelli, A.; Conforti, F.L.; Perri, E.; Ienco, A.; et al. Association study of the 5′ UTR intron of the FAD2-2 gene with oleic and linoleic acid content in Olea europaea L. Front. Plant Sci. 2020, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Van Eeuwijk, F.A.; Bustos-Korts, D.; Millet, E.J.; Boer, M.P.; Kruijer, W.; Thompson, A.; Malosetti, M.; Iwata, H.; Quiroz, R.; Kuppe, C.; et al. Modelling strategies for assessing and increasing the effectiveness of new phenotyping techniques in plant breeding. Plant Sci. 2019, 282, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Podlich, D.W.; Winkler, C.R.; Cooper, M. Mapping as you go. An effective approach for marker assisted selection of complex traits. J. Crop Sci. 2004, 44, 1560–1571. [Google Scholar] [CrossRef] [Green Version]
- Malosetti, M.; Ribaut, J.M.; Van Eeuwijk, F.A. The statistical analysis of multi-environment data: Modeling genotype-by-environment interaction and its genetic basis. Front. Physiol. 2013, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Hammer, G.; Messina, C.; Van Oosterom, E.; Chapman, S.; Singh, V.; Borrell, A.; Jordan, D.; Cooper, M. Molecular breeding for complex adaptive traits: How integrating crop ecophysiology and modelling can enhance efficiency. In Crop Systems Biology; Yin, X., Struik, P., Eds.; Springer: Cham, Switzerland, 2016; pp. 147–162. [Google Scholar] [CrossRef]
- Van Eeuwijk, F.A.; Bink, M.C.A.M.; Chenu, K.; Chapman, S.C. Detection and use of QTL for complex traits in multiple environments. Curr. Opin. Plant Biol. 2010, 13, 193–205. [Google Scholar] [CrossRef]
- Bustos-Korts, D.; Malosetti, M.; Chapman, S.; Van Eeuwijk, F. Modelling of genotype by environment interaction and prediction of complex traits across multiple environments as a synthesis of crop growth modelling, genetics and statistics. In Crop Systems Biology; Yin, X., Struik, P., Eds.; Springer: Cham, Switzerland, 2016; pp. 55–82. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.B.; Cullis, B.R.; Thompson, R. The analysis of crop cultivar breeding and evaluation trials: An overview of current mixed model approaches. J. Agric. Sci. 2005, 143, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Technow, F.; Messina, C.D.; Totir, L.R.; Cooper, M. Integrating crop growth models with whole genome prediction through approximate bayesian computation. PLoS ONE 2015, 10, e0130855. [Google Scholar] [CrossRef] [Green Version]
- López-Bernal, Á.; Morales, A.; García-Tejera, O.; Testi, L.; Orgaz, F.; De Melo-Abreu, J.P.; Villalobos, F.J. OliveCan: A process-based model of development, growth and yield of olive orchards. Front. Plant Sci. 2018, 9, 632. [Google Scholar] [CrossRef] [Green Version]
- Cruz, F.; Julca, I.; Gómez-Garrido, J.; Loska, D.; Marcet-Houben, M.; Cano, E.; Galán, B.; Frias, L.; Ribeca, P.; Derdak, S.; et al. Genome sequence of the olive tree, Olea europaea. GigaScience 2016, 5, 29. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Consumption of olive oil. Provisory data from the campaign year 2018/2019: (a) World consumption; (b) European consumption (data from International Olive Council (IOC) [10]).
Figure 1.
Consumption of olive oil. Provisory data from the campaign year 2018/2019: (a) World consumption; (b) European consumption (data from International Olive Council (IOC) [10]).

Figure 2.
Production of olive oil. Provisory data from the campaign year 2018/2019: (a): World production; (b): European production (data from IOC [10]).
Figure 2.
Production of olive oil. Provisory data from the campaign year 2018/2019: (a): World production; (b): European production (data from IOC [10]).

Figure 3.
Evolution of production of olive oil and olives in the Portuguese olive sector since the 1990/1991 campaign year (data from IOC [10] and Instituto Nacional de Estatística (INE) [13]).

Figure 4.
Production (thousand tons) and surface (ha) of olives for oil and table olives by NUTS (nomenclature of territorial units for statistics). Provisory data from the campaign year 2018/2019: (a) Portuguese production of olives for oil and table olives; (b) surface of olives for olive oil and table olives occupied in Portugal (data from INE [13]).
Figure 4.
Production (thousand tons) and surface (ha) of olives for oil and table olives by NUTS (nomenclature of territorial units for statistics). Provisory data from the campaign year 2018/2019: (a) Portuguese production of olives for oil and table olives; (b) surface of olives for olive oil and table olives occupied in Portugal (data from INE [13]).

Figure 5.
‘Galega’ olive tree, the main Portuguese variety: (a) ‘Galega’ in a traditional olive orchard in Portugal; (b) Abaxial and adaxial surfaces of ‘Galega’ leaves; (c) Detail of the ‘Galega’ inflorescence before the pollination; (d) Flowers of ‘Galega’; (e) Drupes in ‘Galega’ tree; (f) Endocarp inside the drupe and the mesocarp; (g) Endocarp of ‘Galega.’ Photographs by Hélia Sales.
Figure 5.
‘Galega’ olive tree, the main Portuguese variety: (a) ‘Galega’ in a traditional olive orchard in Portugal; (b) Abaxial and adaxial surfaces of ‘Galega’ leaves; (c) Detail of the ‘Galega’ inflorescence before the pollination; (d) Flowers of ‘Galega’; (e) Drupes in ‘Galega’ tree; (f) Endocarp inside the drupe and the mesocarp; (g) Endocarp of ‘Galega.’ Photographs by Hélia Sales.

Figure 6.
Single or multiple independent domestications for cultivated olive trees, the unanswered question: (a,b) distribution olive genepools in the Mediterranean Basin, accordingly to Besnard et al. [104] and Díez et al. [105], respectively; (c,d) simplified scenarios of domestication; (c) one single primary domestication event; (d) two independent primary domestication events. Green spheres indicate the primary domestication events, horizontal arrows indicate the possible occurrence of gene flow and the dashed horizontal connections indicate the admixture events (schematic representation made in ©BioRender—biorender.com).
Figure 6.
Single or multiple independent domestications for cultivated olive trees, the unanswered question: (a,b) distribution olive genepools in the Mediterranean Basin, accordingly to Besnard et al. [104] and Díez et al. [105], respectively; (c,d) simplified scenarios of domestication; (c) one single primary domestication event; (d) two independent primary domestication events. Green spheres indicate the primary domestication events, horizontal arrows indicate the possible occurrence of gene flow and the dashed horizontal connections indicate the admixture events (schematic representation made in ©BioRender—biorender.com).


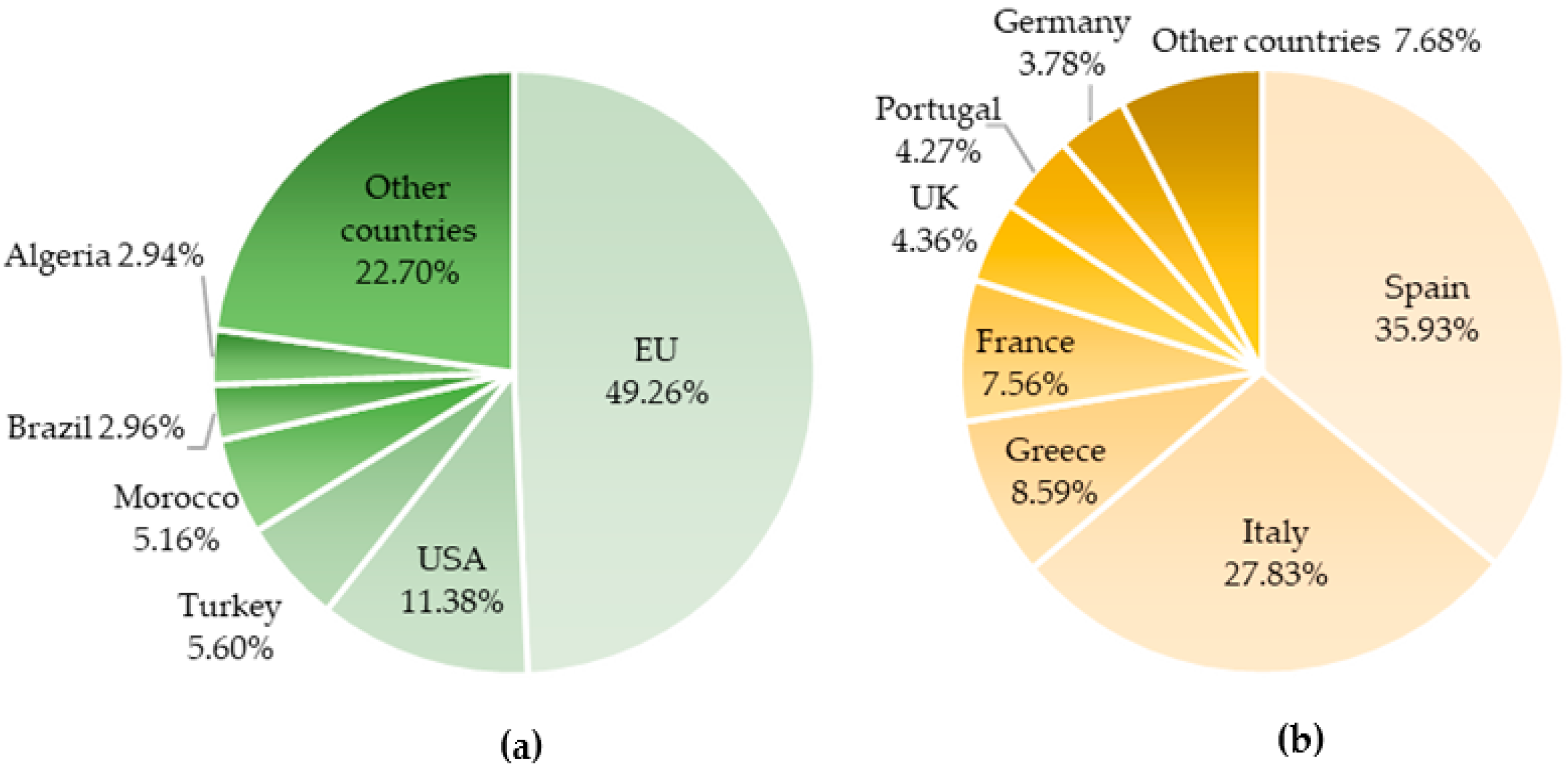{kind=link}
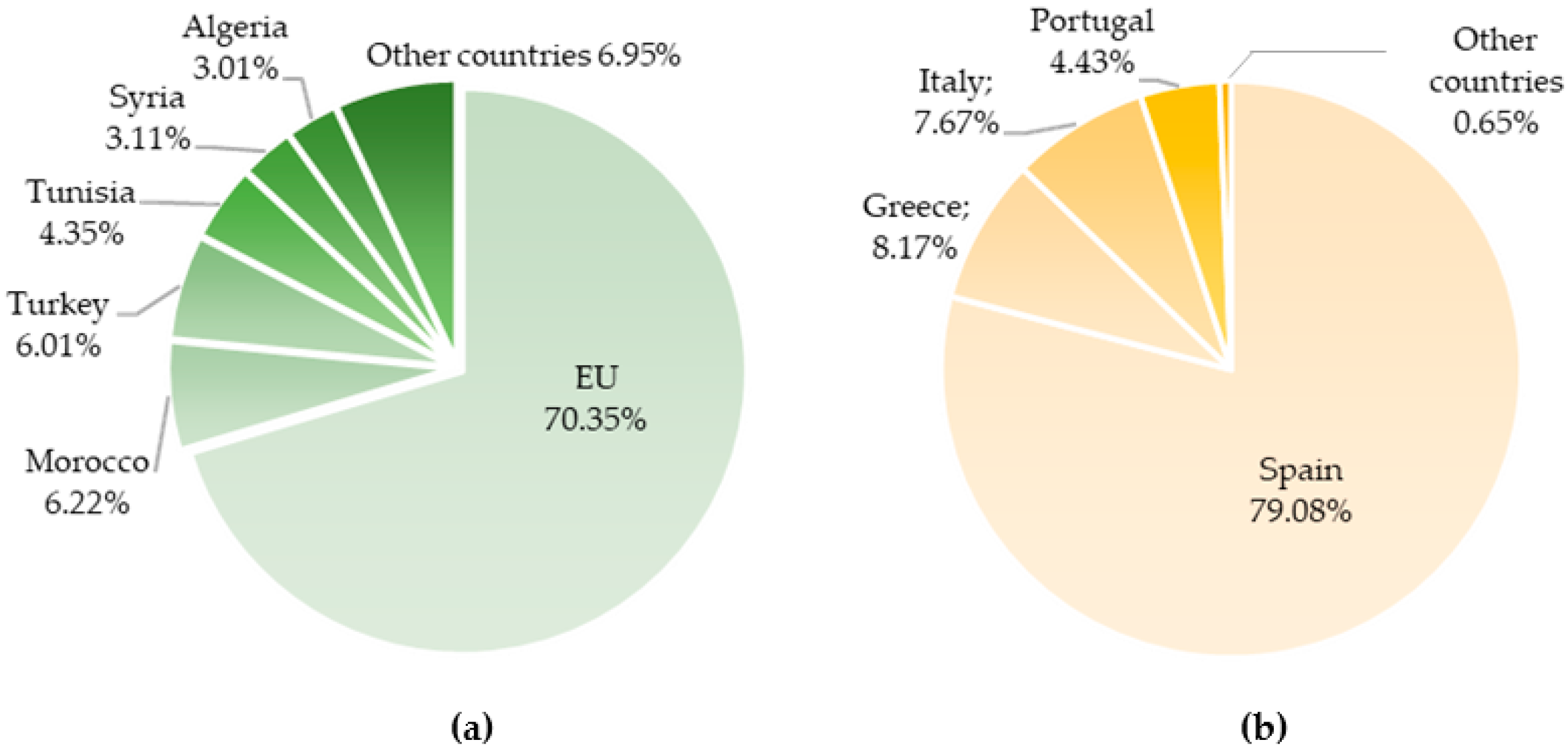{kind=link}
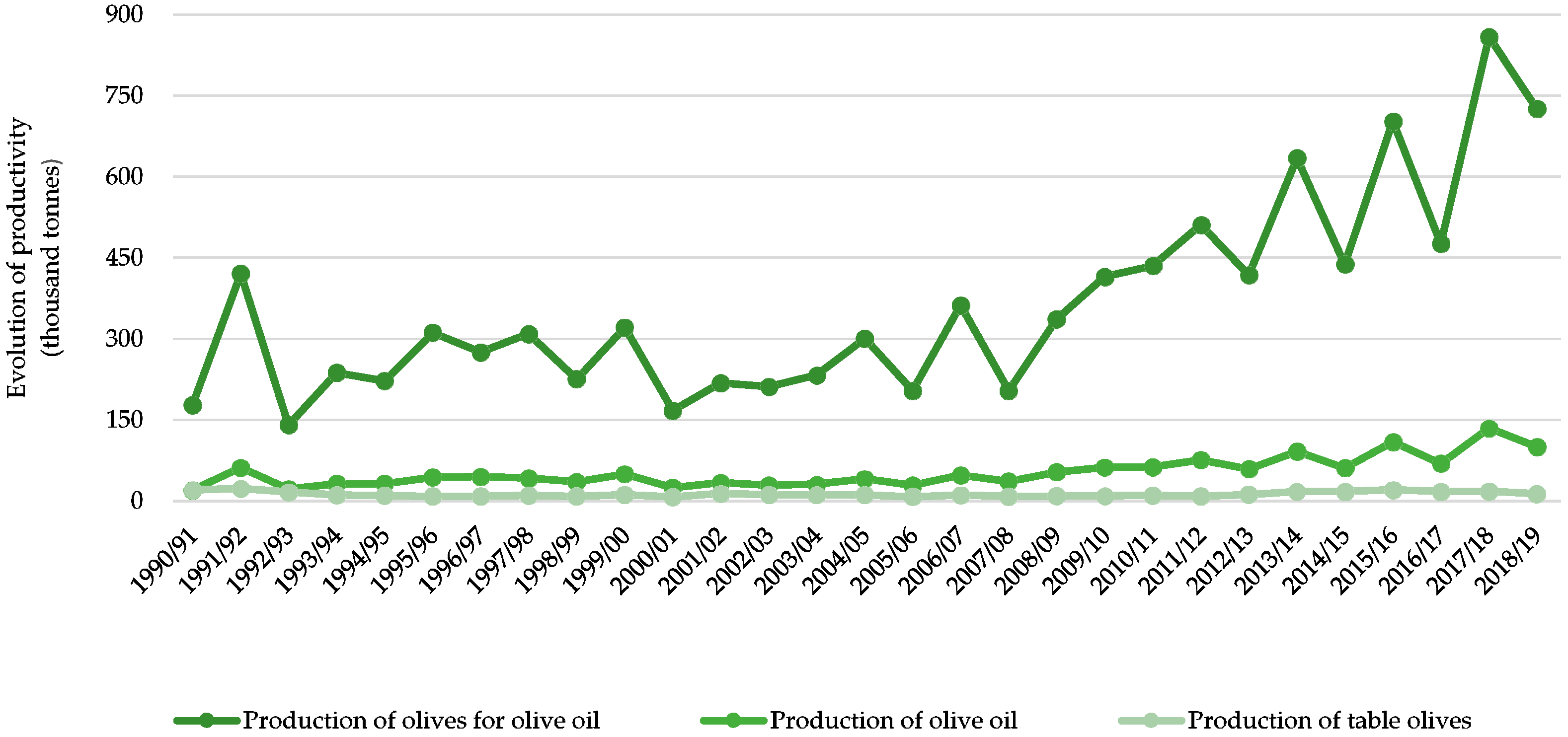{kind=link}
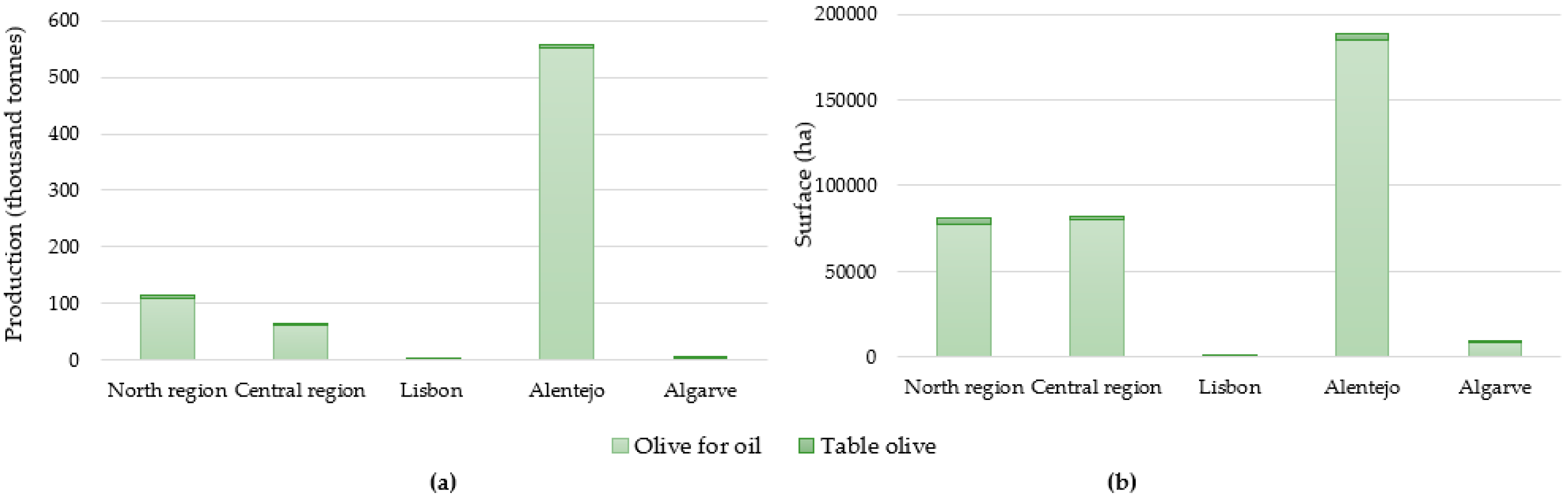{kind=link}
{kind=link}
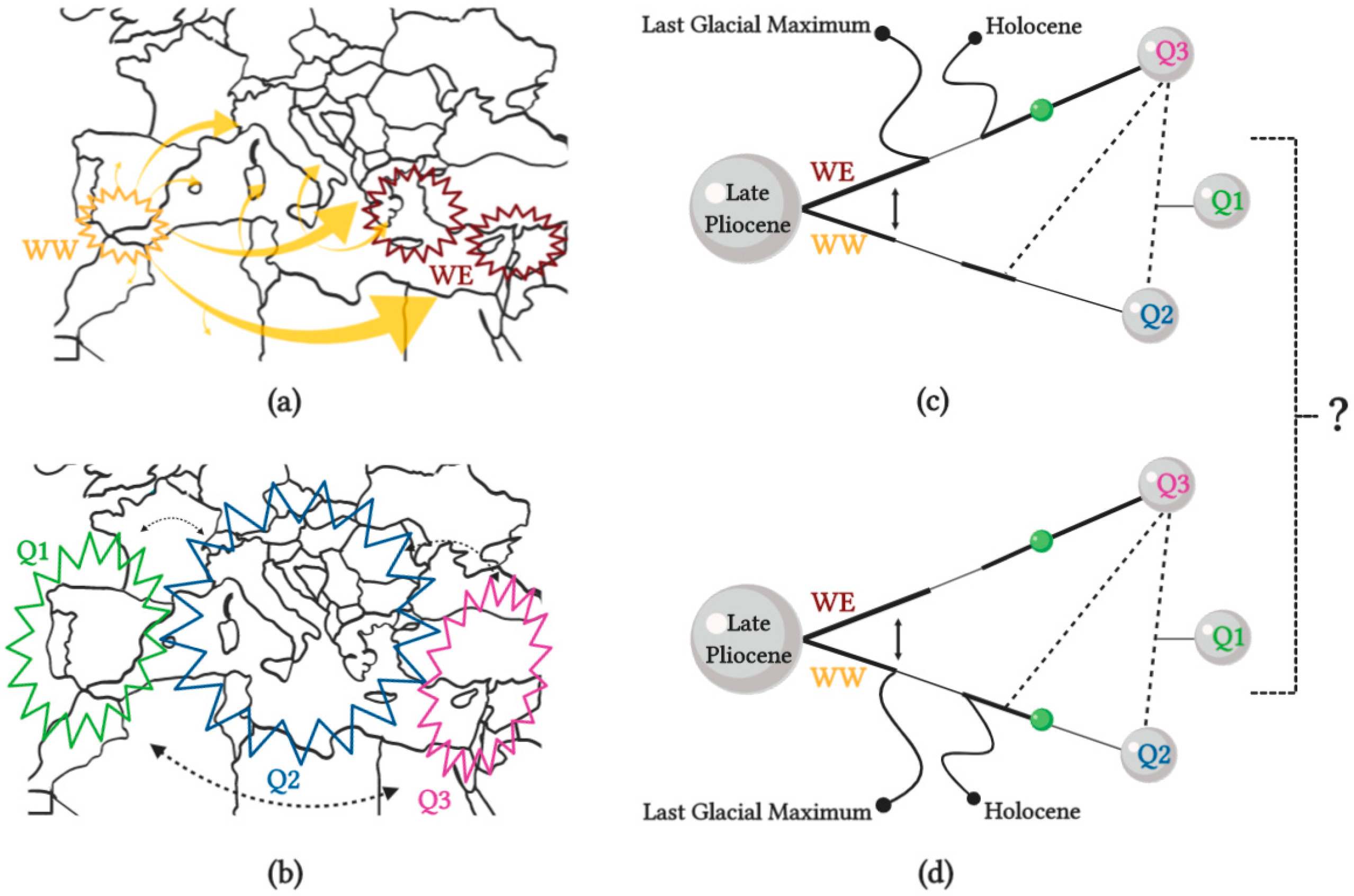{kind=link}
Table 1.
Research developed considering the ‘Galega’ variety in the last years.
| Area | Applications | References |
|---|---|---|
| Ecophysiology | Water use efficiency; salinity stress | [23,24] |
| Rooting | [25] | |
| Effect of chemical and natural compounds application; influence UV-B radiation | [26,27,28] | |
| Phenology studies | [29] | |
| Different types of pollination; profilin polymorphism | [26,30,31] | |
| Pollen viability and germination | [32] | |
| In vitro culture | Gene characterization and expression | [33,34] |
| Adventitious root formation | [35,36] | |
| Micropropagation | [37,38] | |
| Genetic resources | Olive varieties identification and genetic relationships | [39,40,41,42,43,44,45,46,47] |
| Genetic variability; genotyping and genetic mapping | [22,44,48,49,50,51,52,53] | |
| Improvement in production systems and cultural techniques | Mechanical pruning and harvesting; effect of rejuvenation pruning | [54,55] |
| Phytosanitary protection | Richness and diversity of fungal communities | [56,57,58] |
| Susceptibility to fungal diseases; response to pathogens attack | [59,60,61,62] | |
| Technology, quality and markets | Analytical characterization of olive oil compounds; certification and olive oil traceability | [63,64] |
| Effect of blending and storage time of olive oil | [65] | |
| Effect of fertilization on olive oil polyphenol, sterol and wax content | [66] | |
| Extraction of monovarietal olive oils with natural compounds | [67] | |
| Physicochemical, nutritional and microbiological characterization; health effects; agro-environmental factors on the olive oil mineral content | [68,69,70] |
Table 2.
Life Cycle Assessment (LCA) studies performed in the olive sector in the last 10 years.
| Aim of the Application of LCA Methodology | FU 1 | Country | References |
|---|---|---|---|
| olive oil production | 1 ton of olives | Italy | [83] |
| 1 ha | Iran | [84] | |
| 1 L EVOO 2 | Italy | [85] | |
| 1 L EVOO | Greece | [86] | |
| 1 kg of olive oil | Jordan | [87] | |
| 5 L EVOO | Italy | [88] | |
| distribution processes of olive oil supply chain and packaging | 1 L EVOO | Italy | [89] |
| 1 L of bottling capacity | Italy | [90] | |
| 0.5 L bottle of VOO 3 | Spain | [91] | |
| olive oil industry waste treatments | 1 L olive mill waste | Greece | [92] |
| 1 mg of olive solid waste | Australia | [93] | |
| 1 ton of torrefied olive husk | Cyprus | [94] | |
| 100 kg olive pomace | Spain | [95] | |
| 1MJ of energy production | Spain | [96] | |
| field agricultural practices | 1 ha | Italy | [97,98] |
| 1 ton of olives | Spain | [99] | |
| olive-harvesting practices | 1 ha; 1 kg | Italy | [100] |
1 FU—Functional Unit 2 EVOO—Extra virgin olive oil. 3 VOO—virgin olive oil.
Table 3.
Applications of some non-species-specific molecular markers in olive varieties.
| Method | Applications | References |
|---|---|---|
| Isoenzymes | Olive varieties identification; genetic diversity; genetic relationships | [118] |
| RFLP (Restriction Fragment Length Polymorphism) | Genetic distance/genetic relationships | [119] |
| Genetic linkage map | [120] | |
| RAPD (Randomly Amplified Polymorphic DNA) | Olive varieties identification; genetic diversity; genetic relationships | [22,41,45,49,51,53,118,121,122,123] |
| Genetic distance estimation between wild and cultivated olive genotypes | [119,124] | |
| Genetic linkage map | [120,125,126] | |
| ISSRs (InterSimple Sequence Repeats) | Olive varieties identification; genetic diversity; genetic relationships | [22,41,42,53,122,123,127,128] |
| AFLPs (Amplified Fragment Length Polymorphism) | Olive varieties identification; genetic diversity; genetic relationships | [129,130,131] |
| Genetic linkage map | [120,126,132,133] |
Table 4.
Applications of SSRs and SNPs in olive varieties.
| Method | Applications | References |
|---|---|---|
| SSRs | Identification/characterization of markers | [39,40,136,142,143] |
| Olive varieties identification; genetic diversity; genetic relationships | [4,40,42,46,47,48,50,75,105,112,123,138,144,145,146,147,148] | |
| Genetic linkage map | [120,125,126,132,133,149] | |
| Paternity analysis | [150,151] | |
| SNPs | Olive varieties identification; genetic diversity; genetic relationships | [52,141,152,153] |
| Genetic linkage map | [154,155,156,157,158] |
Table 5.
Precision breeding approaches and breeding innovations in Olea sp.
| Area | References |
|---|---|
| In vitro techniques for supporting unconventional methods of genetic improvement | [33,34,35,37,38,173,174,175] |
| Plant regeneration from in vitro cultured tissues | [176,177] |
| Genetic transformation and plant recovery | [178,179] |
| Transcriptome analysis | [180,181,182,183,184,185,186,187,188,189,190,191] |
| Plastome analysis | [2,43,192,193] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sales, H.; Nunes, J.; Vaz Patto, M.C. Achievements and Challenges towards a Sustainable Conservation and Use of ‘Galega vulgar’ Olea europaea Variety. Agronomy 2020, 10, 1467. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101467
AMA Style
Sales H, Nunes J, Vaz Patto MC. Achievements and Challenges towards a Sustainable Conservation and Use of ‘Galega vulgar’ Olea europaea Variety. Agronomy. 2020; 10(10):1467. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101467
Chicago/Turabian StyleSales, Hélia, João Nunes, and Maria Carlota Vaz Patto. 2020. "Achievements and Challenges towards a Sustainable Conservation and Use of ‘Galega vulgar’ Olea europaea Variety" Agronomy 10, no. 10: 1467. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10101467
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.