
The Role of Peat-Free Organic Substrates in the Sustainable Management of Soilless Cultivations


Abstract
:1. Introduction
2. Soilless Systems and Circular Horticulture
3. Overview of the Investigated Materials
4. The Role of Organic Substrates in Plant Nutrition and Irrigation of Soilless Crops
4.1. Biochar
4.2. Coir
4.3. Green Compost
4.4. Wood Fiber
5. The Role of Organic Substrates in Crop Tolerance to Abiotic and Biotic Stresses of Soilless Crops
5.1. Biochar
5.2. Coir
5.3. Green Compost
5.4. Wood Fibers
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Putra, P.A.; Yuliando, H. Soilless culture system to support water use efficiency and product quality: A review. Agric. Agric. Sci. Proc. 2015, 3, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Raviv, M.; Lieth, J.H.; Bar-Tal, A. Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems—A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Majsztrik, J.C.; Ristvey, A.G.; Lea-Cox, J.D. Water and nutrient management in the production of container-grown ornamentals. Hortic. Rev. 2011, 38, 253–297. [Google Scholar]
- Massa, D.; Magán, J.J.; Montesano, F.F.; Tzortzakis, N. Minimizing water and nutrient losses from soilless cropping in southern Europe. Agric. Water Manag. 2020, 241, 106395. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant Sci. 2018, 9, 1–6. [Google Scholar] [CrossRef]
- Atzori, G.; Mancuso, S.; Masi, E. Seawater potential use in soilless culture: A review. Sci. Hortic. 2019, 249, 199–207. [Google Scholar] [CrossRef]
- Atzori, G.; Guidi Nissim, W.; Caparrotta, S.; Masi, E.; Azzarello, E.; Pandolfi, C.; Vignolini, P.; Gonnelli, C.; Mancuso, S. Potential and constraints of different seawater and freshwater blends as growing media for three vegetable crops. Agric. Water Manag. 2016, 176, 255–262. [Google Scholar] [CrossRef]
- Bar Yosef, B. Advances in fertigation. Adv. Agron. 1999, 65, 1–77. [Google Scholar]
- Savvas, D.; Gruda, N. Application of soilless culture technologies in the modern greenhouse industry—A review. Eur. J. Hortic. Sci. 2018, 83, 280–293. [Google Scholar] [CrossRef]
- Papadopoulos, A.P.; Bar-Tal, A.; Silber, A.; Saha, U.K.; Raviv, M. Inorganic and synthetic organic components of soilless culture and potting mixes. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2008; pp. 505–543. [Google Scholar]
- Raviv, M. Composts in growing media: What’s new and what’s next? Acta Hortic. 2013, 982, 39–52. [Google Scholar] [CrossRef]
- Atzori, G.; Nissim, W.G.; Rodolfi, L. Algae and Bioguano as promising source of organic fertilizers. J. Appl. Phycol. 2020, 32, 3971–3981. [Google Scholar] [CrossRef]
- Sakai, S.I.; Yoshida, H.; Hirai, Y.; Asari, M.; Takigami, H.; Takahashi, S.; Tomoda, K.; Peeler, M.V.; Wejchert, J.; Schmid-Unterseh, T.; et al. International comparative study of 3R and waste management policy developments. J. Mater. Cycles Waste Manag. 2011, 13, 86–102. [Google Scholar] [CrossRef] [Green Version]
- Carlile, W.R.; Raviv, M.; Prasad, M. Organic Soilless Media Components; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 9780444636966. [Google Scholar]
- Schmilewski, G. Growing media constituents used in the EU in 2013. Acta Hortic. 2017, 1168, 85–92. [Google Scholar] [CrossRef]
- Bourguignon, D. Closing the Loop: New Circular Economy Package. Available online: https://www.europarl.europa.eu/RegData/etudes/BRIE/2016/573899/EPRS_BRI(2016)573899_EN.pdf (accessed on 10 April 2021).
- Kiss, K.; Ruszkai, C.; Takacs-Gyorgy, K. Examination of short supply chains based on circular economy and sustainability aspects. Resources 2019, 8, 161. [Google Scholar] [CrossRef] [Green Version]
- Verain, M.C.D.; Sijtsema, S.J.; Antonides, G. Consumer segmentation based on food-category attribute importance: The relation with healthiness and sustainability perceptions. Food Qual. Prefer. 2016, 48, 99–106. [Google Scholar] [CrossRef]
- Schmilewski, G. Producing growing media responsibly to help sustain horticulture. Acta Hortic. 2014, 1034, 299–306. [Google Scholar] [CrossRef]
- Raviv, M.; Lieth, J.H. Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Bullock, C.H.; Collier, M.J.; Convery, F. Land use policy peatlands, their economic value and priorities for their future management—The example of Ireland. Land Use Policy 2012, 29, 921–928. [Google Scholar] [CrossRef]
- Cleary, J.; Roulet, N.T.; Moore, T.R. Greenhouse gas emissions from Canadian peat extraction, 1990–2000: A life-cycle analysis. AMBIO J. Human Environ. 2005, 34, 456–461. [Google Scholar] [CrossRef]
- Gruda, N.S. Increasing sustainability of growing media constituents and stand-alone substrates in soilless culture systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Mondejar, R.; Bernal-Vicente, A.; Ros, M.; Tittarelli, F.; Canali, S.; Intrigiolo, F.; Pascual, J.A. Utilisation of citrus compost-based growing media amended with Trichoderma harzianum T-78 in Cucumis melo L. seedling production. Bioresour. Technol. 2010, 101, 3718–3723. [Google Scholar] [CrossRef]
- Vandecasteele, B.; Debode, J.; Willekens, K.; Delm, T. Van Recycling of P and K in circular horticulture through compost application in sustainable growing media for fertigated strawberry cultivation. Eur. J. Agron. 2018, 96, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Zulfiqar, F.; Allaire, S.E.; Akram, N.A.; Méndez, A.; Younis, A.; Peerzada, A.M.; Shaukat, N.; Wright, S.R. Challenges in organic component selection and biochar as an opportunity in potting substrates: A review. J. Plant Nutr. 2019, 42, 1386–1401. [Google Scholar] [CrossRef]
- Jindo, K.; Sánchez-Monedero, M.A.; Mastrolonardo, G.; Audette, Y.; Higashikawa, F.S.; Silva, C.A.; Akashi, K.; Mondini, C. Role of biochar in promoting circular economy in the agriculture sector. Part 2: A review of the biochar roles in growing media, composting and as soil amendment. Chem. Biol. Technol. Agric. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Sohi, S.P.; Krull, E.; Lopez-Capel, E.; Bol, R. A review of biochar and its use and function in soil. Adv. Agron. 2010, 105, 47–82. [Google Scholar]
- Evans, M.R.; Jackson, B.E.; Popp, M.; Sadaka, S. Chemical properties of biochar materials manufactured from agricultural products common to the southeast united states. Horttechnology 2017, 27, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Massa, D.; Bonetti, A.; Cacini, S.; Faraloni, C.; Prisa, D.; Tuccio, L.; Petruccelli, R. Soilless tomato grown under nutritional stress increases green biomass but not yield or quality in presence of biochar as growing medium. Hortic. Environ. Biotechnol. 2019, 60, 871–881. [Google Scholar] [CrossRef]
- Zaccheo, P.; Crippa, L.; Bedussi, F. Biochar can enhance potassium availability during cyclamen cultivation. Acta Hortic. 2019, 1266, 35–42. [Google Scholar] [CrossRef]
- Prasad, M.; Chrysargyris, A.; McDaniel, N.; Kavanagh, A.; Gruda, N.S.; Tzortzakis, N. Plant nutrient availability and pH of biochars and their fractions, with the possible use as a component in a growing media. Agronomy 2020, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Noguera, P.; Abad, M.; Puchades, R.; Maquieira, A.; Noguera, V. Influence of particle size on physical and chemical properties of coconut coir dust as container medium. Commun. Soil Sci. Plant Anal. 2003, 34, 593–605. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Efficacy of organic amendments used in containerized plant production: Part 1—Compost-based amendments. Sci. Hortic. 2020, 266, 108856. [Google Scholar] [CrossRef]
- Carlile, W.R.; Cattivello, C.; Zaccheo, P. Organic growing media: Constituents and properties. Vadose Zone J. 2015, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Abad, M.; Noguera, P.; Puchades, R.; Maquieira, A.; Noguera, V. Physico-chemical and chemical properties of some coconut coir dusts for use as a peat substitute for containerised ornamental plants. Bioresour. Technol. 2002, 82, 241–245. [Google Scholar] [CrossRef]
- Mariotti, B.; Martini, S.; Raddi, S.; Tani, A.; Jacobs, D.F.; Oliet, J.A.; Maltoni, A. Coconut coir as a sustainable nursery growing media for seedling production of the ecologically diverse quercus species. Forests 2020, 11, 522. [Google Scholar] [CrossRef]
- Poulter, R. Quantifying differences between treated and untreated coir substrate. Acta Hortic. 2014, 1018, 557–564. [Google Scholar] [CrossRef]
- Hongpakdee, P.; Ruamrungsri, S. Water use efficiency, nutrient leaching, and growth in potted marigolds affected by coconut coir dust amended in substrate media. Hortic. Environ. Biotechnol. 2015, 56, 27–35. [Google Scholar] [CrossRef]
- Xiong, J.; Tian, Y.; Wang, J.; Liu, W.; Chen, Q. Comparison of coconut coir, rockwool, and peat cultivations for tomato production: Nutrient balance, plant growth and fruit quality. Front. Plant. Sci. 2017, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Abad, M.; Fornes, F.; Carrión, C.; Noguera, V.; Noguera, P.; Maquieira, Á.; Puchades, R. Physical properties of various coconut coir dusts compared to peat. HortScience 2005, 40, 2138–2144. [Google Scholar] [CrossRef] [Green Version]
- Michel, J.C. Wettability of organic growing media used in horticulture: A review. Vadose Zone J. 2015, 14, 1–6. [Google Scholar] [CrossRef]
- Shara, S.A.; Zaharah, S.S.; Radziah, O.; Puteri, E.M.W. Physical, chemical and microbiological properties of different combination of soilless media and their effect on the vegetative component and nutrient content of hempedu bumi (Andrographis paniculata). Pertanika J. Trop. Agric. Sci. 2017, 40, 35–52. [Google Scholar]
- Fascella, G.; Zizzo, G.V.; Agnello, S. Soilless cultivation of mother plants of Euphorbia x lomi hybrids on different substrates. Acta Hortic. 2006, 718, 507–513. [Google Scholar] [CrossRef]
- Fascella, G.; Zizzo, G. Efficient propagation technique of Euphorbia ×lomi thai hybrids. HortScience 2009, 44, 495–498. [Google Scholar] [CrossRef] [Green Version]
- Nappi, P.; Barberis, R. Compost as growing medium: Chemical, physical and biological aspects. Acta Hortic. 1993, 342, 249–256. [Google Scholar] [CrossRef]
- Farrell, M.; Jones, D.L. Food waste composting: Its use as a peat replacement. Waste Manag. 2010, 30, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Barker, A.V.; Bryson, G.M. Comparisons of composts with low or high nutrient status for growth of plants in containers. Commun. Soil Sci. Plant. Anal. 2006, 37, 1303–1319. [Google Scholar] [CrossRef]
- Cendón, Y.; Moldes, A.; Barral, M.T. Evaluation of municipal solid waste compost as a growing media component for potted plant production. Acta Hortic. 2008, 779, 591–598. [Google Scholar] [CrossRef]
- Ostos, J.C.; Lopez-Garrido, R.; Murillo, J.M.; Lopez, R. Substitution of peat for municipal solid waste- and sewage sludge-based composts in nursery growing media: Effects on growth and nutrition of the native shrub Pistacia lentiscus L. Bioresour. Technol. 2008, 99, 1793–1800. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Saridakis, C.; Tzortzakis, N. Use of municipal solid waste compost as growing medium component for melon seedlings production. Plant. Biol. Soil Health 2013, 1, 1–5. [Google Scholar]
- Zoes, V.; Paré, T.; Dinel, H.; Dumontet, S.; Pasquale, V.; Scopa, A. Growth and yield of tomato cultivated on composted duck excreta enriched wood shavings and source-separated municipal solid waste. Ital. J. Agron. 2011, 6, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Sciubba, L.; Cavani, L.; Grigatti, M.; Ciavatta, C.; Marzadori, C. Relationships between stability, maturity, water-extractable organic matter of municipal sewage sludge composts and soil functionality. Environ. Sci. Pollut. Res. 2015, 22, 13393–13403. [Google Scholar] [CrossRef] [PubMed]
- Eklind, Y.; Rämert, B.; Wivstad, M. Evaluation of growing media containing farmyard manure compost, household waste compost or chicken manure for the propagation of lettuce (Lactuca sativa L.) transplants. Biol. Agric. Hortic. 2001, 19, 157–181. [Google Scholar] [CrossRef]
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Bioresource technology composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef]
- Bustamante, M.A.; Paredes, C.; Moral, R.; Agullo, E.; Pérez-Murcia, M.D.; Abad, M. Composts from distillery wastes as peat substitutes for transplant production. Resour. Conserv. Recycl. 2008, 52, 792–799. [Google Scholar] [CrossRef]
- Kritsotakis, I.K.; Kabourakis, E.M. Grape vine waste and giant reed biomass composts as peat and mineral fertilizer substitutes for producing organic tomato transplants. J. Crop. Improv. 2011, 25, 664–679. [Google Scholar] [CrossRef]
- Massa, D.; Malorgio, F.; Lazzereschi, S.; Carmassi, G.; Prisa, D.; Burchi, G. Evaluation of two green composts for peat substitution in geranium (Pelargonium zonale L.) cultivation: Effect on plant growth, quality, nutrition, and photosynthesis. Sci. Hortic. 2018, 228, 213–221. [Google Scholar] [CrossRef]
- Brinton, W.F. Compost Quality Standards and Guidelines; Woods End Laboratories Inc.: Mount Vernon, ME, USA, 2000. [Google Scholar]
- St. Martin, C.C.G.; Brathwaite, R.A.I. Compost and compost tea: Principles and prospects as substrates and soil-borne disease management strategies in soil-less vegetable production. Biol. Agric. Hortic. 2012, 28, 1–33. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 2014, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, V.; Di Meo, V.; Monda, H.; Spaccini, R.; Piccolo, A. The molecular characteristics of compost affect plant growth, arbuscular mycorrhizal fungi, and soil microbial community composition. Biol. Fertil. Soils 2016, 52, 15–29. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Monda, H.; Cozzolino, V.; Vinci, G.; Drosos, M.; Savy, D.; Piccolo, A. Molecular composition of the Humeome extracted from different green composts and their biostimulation on early growth of maize. Plant. Soil 2018, 429, 407–424. [Google Scholar] [CrossRef]
- Olszewski, M.W.; Trego, T.A.; Kuper, R. Effects of peat moss substitution with arboretum and greenhouse waste compost for use in container media. Compost Sci. Util. 2009, 17, 151–157. [Google Scholar] [CrossRef]
- Lopez-Cuadrado, M.C.; Ruiz-Fernandez, J.; Masaguer, A.; Moliner, A. Utilization of different organic wastes from Madrid as growth media for Pelargonium zonale. Acta Hortic. 2008, 779, 623–630. [Google Scholar] [CrossRef]
- Eklind, Y.; Salomonsson, L.; Wivstad, M.; Rämert, B. Use of herbage compost as horticultural substrate and source of plant nutrients. Biol. Agric. Hortic. 1998, 16, 269–290. [Google Scholar] [CrossRef]
- Pane, C.; Celano, G.; Piccolo, A.; Villecco, D.; Spaccini, R.; Palese, A.M.; Zaccardelli, M. Effects of on-farm composted tomato residues on soil biological activity and yields in a tomato cropping system. Chem. Biol. Technol. Agric. 2015, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Liguori, L.; Pane, C.; Albanese, D.; Celano, G.; Zaccardelli, M.; Di Matteo, M. Compost and compost tea management of mini watermelon cultivations affects the chemical, physical and sensory assessment of the fruits. Agric. Sci. 2015, 6, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Mugnai, S.; Pasquini, T.; Azzarello, E.; Pandolfi, C.; Mancuso, S. Evaluation of composted green waste in ornamental container-grown plants: Effects on growth and plant water relations. Compost Sci. Util. 2007, 15, 283–287. [Google Scholar] [CrossRef]
- Massa, D.; Prisa, D.; Lazzereschi, S.; Cacini, S.; Burchi, G. Heterogeneous response of two bedding plants to peat substitution by two green composts. Hort. Sci. 2018, 45, 164–172. [Google Scholar] [CrossRef]
- Aleandri, M.P.; Chilosi, G.; Muganu, M.; Vettraino, A.; Marinari, S.; Paolocci, M.; Luccioli, E.; Vannini, A. On farm production of compost from nursery green residues and its use to reduce peat for the production of olive pot plants. Sci. Hortic. 2015, 193, 301–307. [Google Scholar] [CrossRef]
- Prasad, M.; Maher, M.J. The use of composted green waste (CGW) as a growing medium component. Acta Hortic. 2001, 549, 107–113. [Google Scholar] [CrossRef]
- Benito, M.; Masaguer, A.; Antonio, R.D.; Moliner, A. Use of pruning waste compost as a component in soilless growing media. Bioresour. Technol. 2005, 96, 597–603. [Google Scholar] [CrossRef]
- Brito, L.M.; Reis, M.; Mourão, I.; Coutinho, J. Use of Acacia waste compost as an alternative component for horticultural substrates. Commun. Soil Sci. Plant. Anal. 2015, 46, 1814–1826. [Google Scholar] [CrossRef]
- Marble, S.C.; Fain, G.B.; Gilliam, C.H.; Runion, G.B.; Prior, S.A.; Torbert, H.A.; Wells, D.E. Landscape establishment of woody ornamentals grown in alternative wood-based container substrates. J. Environ. Hort 2012, 30, 13–16. [Google Scholar] [CrossRef]
- Maher, M.; Prasad, M.; Raviv, M. Organic soilless media components. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2008; pp. 459–504. [Google Scholar]
- Gruda, N.; Schnitzler, W.H. Suitability of wood fiber substrates for production of vegetable transplants II. The effect of wood fiber substrates and their volume weights on the growth of tomato transplants. Sci. Hortic. 2004, 100, 333–340. [Google Scholar] [CrossRef]
- Schmilewski, G.; Nordzieke, B. Researched, developed and commercialized: GreenFiber. Acta Hortic. 2019, 1266, 361–368. [Google Scholar] [CrossRef]
- Michel, J.C.; Jackson, B.E.; Fonteno, W.C. Classification of organic substrates’ wettability from contact angle measurements and hydration efficiency tests. Acta Hortic. 2017, 1168, 199–206. [Google Scholar] [CrossRef]
- Michel, J.C.; Kerloch, E. Evolution of hydraulic properties and wettability of organic growing media during cultivation according to irrigation strategies. Sci. Hortic. 2017, 217, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Apaolaza, L.; Gascó, A.M.; Gascó, J.M.; Guerrero, F. Reuse of waste materials as growing media for ornamental plants. Bioresour. Technol. 2005, 96, 125–131. [Google Scholar] [CrossRef]
- Rosen, C.J.; Halbach, T.R.; Swanson, B.T. Horticultural uses of municipal solid waste composts. Horttechnology 1993, 3, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Yargicoglu, E.N.; Yamini, B.; Reddy, K.R.; Spokas, K. Physical and chemical characterization of waste wood derived biochars. Waste Manag. 2015, 36, 256–268. [Google Scholar] [CrossRef]
- Kaudal, B.B.; Chen, D.; Madhavan, D.B.; Downie, A.; Weatherley, A. An examination of physical and chemical properties of urban biochar for use as growing media substrate. Biomass Bioenergy 2016, 84, 49–58. [Google Scholar] [CrossRef]
- Benito, M.; Masaguer, A.; Moliner, A.; De Antonio, R. Chemical and physical properties of pruning waste compost and their seasonal variability. Bioresour. Technol. 2006, 97, 2071–2076. [Google Scholar] [CrossRef] [PubMed]
- Domeño, I.; Irigoyen, I.; Muro, J. New wood fibre substrates characterization and evaluation in hydroponic tomato culture. Eur. J. Hortic. Sci. 2010, 75, 89–94. [Google Scholar]
- Abad, M.; Noguera, P.; Burés, S. National inventory of organic wastes for use as growing media for ornamental potted plant production: Case study in Spain. Bioresour. Technol. 2001, 77, 197–200. [Google Scholar] [CrossRef]
- Kern, J.; Tammeorg, P.; Shanskiy, M.; Sakrabani, R.; Knicker, H.; Kammann, C.; Tuhkanen, E.M.; Smidt, G.; Prasad, M.; Tiilikkala, K.; et al. Synergistic use of peat and charred material in growing media—An option to reduce the pressure on peatlands? J. Environ. Eng. Landsc. Manag. 2017, 25, 160–174. [Google Scholar] [CrossRef]
- NiChualain, D.; Hynes, C.; Carlile, W.C. Physical properties and water uptake in peat-based growing media containing green compost. Acta Hortic. 2014, 1034, 233–240. [Google Scholar] [CrossRef]
- Gruda, N. Current and future perspective of growing media in Europe. Acta Hortic. 2012, 960, 37–43. [Google Scholar] [CrossRef]
- Michel, J. The physical properties of peat: A key factor for modern growing media. Mires Peat 2010, 6, 1–6. [Google Scholar]
- Alsanius, B.W.; Wohanka, W. Root zone microbiology of soilless cropping systems. In Soilless Culture: Theory and Practice, 2nd ed.; Raviv, M., Heinrich Lieth, J., Bar-Tal, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 149–194. [Google Scholar]
- Dewi, T.K.; Mubarok, W.Z.; Antonius, S. Study of plant growth promoting bacteria from coconut coir dust. In Proceedings of the IOP Conference Series: Earth and Environmental Science, International Symposium of Innovative Bio-Production Indonesia on Biotechnology and Bioengineering, Tangerang, Indonesia, 23–24 October 2019. [Google Scholar]
- Hasan, Z.A.E.; Mohd Zainudin, N.A.I.; Aris, A.; Ibrahim, M.H.; Yusof, M.T. Biocontrol efficacy of Trichoderma asperellum-enriched coconut fibre against Fusarium wilts of cherry tomato. J. Appl. Microbiol. 2020, 129, 991–1003. [Google Scholar] [CrossRef]
- Raviv, M. Recent advances in soil-borne disease control using suppressive media. Acta Hortic. 2009, 819, 125–134. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Efficacy of organic amendments used in containerized plant production: Part 2—Non-compost-based amendments. Sci. Hortic. 2020, 260, 108855. [Google Scholar] [CrossRef]
- Fuchs, J.G.; Hedrich, T.; Hofer, V.; Koller, M.; Oberhaensli, T.; Ribera Regal, J.; Tamm, L.; Thuerig, B.; Schwarze, F.W.M.R.; Herforth-Rahmé, J. Development of disease-suppressive organic growing media. Acta Hortic. 2017, 1164, 181–188. [Google Scholar] [CrossRef]
- Hautsalo, J.; Vestberg, M.; Parikka, P.; Kukkonen, S.; Karhu, S.; Tahvonen, R. Biological control of strawberry crown rot is substrate dependent phenomenon. J. Berry Res. 2016, 6, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Edwards, L.C. The Improvement of Plant Performance in Reduced Peat Growing Media Using Arbuscular Mycorrhizal Fungi. Ph.D. Thesis, Royal Holloway, University of London, London, UK, 2017. [Google Scholar]
- Schmilewski, G. The role of peat in assuring the quality of growing media. Mires Peat 2008, 3, 1–8. [Google Scholar]
- Dunlop, S.J.; Arbestain, M.C.; Bishop, P.A.; Wargent, J.J. Closing the loop: Use of biochar produced from tomato crop green waste as a substrate for soilless, hydroponic tomato production. HortScience 2015, 50, 1572–1581. [Google Scholar] [CrossRef]
- Awad, Y.M.; Lee, S.E.; Ahmed, M.B.M.; Vu, N.T.; Farooq, M.; Kim, I.S.; Kim, H.S.; Vithanage, M.; Usman, A.R.A.; Al-Wabel, M.; et al. Biochar, a potential hydroponic growth substrate, enhances the nutritional status and growth of leafy vegetables. J. Clean. Prod. 2017, 156, 581–588. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.R.; El-Nemr, M.A.; Tantawy, A.S.; Habib, H.A. Alleviation of salinity effects on green bean plants using some environmental friendly materials. J. Appl. Sci. Res. 2010, 6, 871–878. [Google Scholar]
- Ben-Laouane, R.; Baslam, M.; Ait-El-mokhtar, M.; Anli, M.; Boutasknit, A.; Ait-Rahou, Y.; Toubali, S.; Mitsui, T.; Oufdou, K.; Wahbi, S.; et al. Potential of native arbuscular mycorrhizal fungi, rhizobia, and/or green compost as alfalfa (Medicago sativa) enhancers under salinity. Microorganisms 2020, 8, 1695. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Baslam, M.; Ben-Laouane, R.; Anli, M.; Boutasknit, A.; Mitsui, T.; Wahbi, S.; Meddich, A. Alleviation of detrimental effects of salt stress on date palm (Phoenix dactylifera L.) by the application of arbuscular mycorrhizal fungi and/or compost. Front. Sustain. Food Syst. 2020, 4, 1–19. [Google Scholar] [CrossRef]
- Incrocci, L.; Massa, D.; Pardossi, A. New trends in the fertigation management of irrigated vegetable crops. Horticulturae 2017, 3, 37. [Google Scholar] [CrossRef]
- Altland, J.E.; Locke, J.C. Biochar affects macronutrient leaching from a soilless substrate. HortScience 2012, 47, 1136–1140. [Google Scholar] [CrossRef]
- Mukherjee, A.; Lal, R. The biochar dilemma. Soil Res. 2014, 52, 217–230. [Google Scholar] [CrossRef]
- Guo, Y.; Niu, G.; Starman, T.; Volder, A.; Gu, M. Poinsettia growth and development response to container root substrate with biochar. Horticulturae 2018, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Sun, X.; Li, S.; Wang, H.; Wang, L.; Cao, J.; Zhang, L. Biochar made from green waste as peat substitute in growth media for Calathea rotundifola cv. Fasciata. Sci. Hortic. 2012, 143, 15–18. [Google Scholar] [CrossRef]
- Liu, Z.; Demisie, W.; Zhang, M. Simulated degradation of biochar and its potential environmental implications. Environ. Pollut. 2013, 179, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Petruccelli, R.; Bonetti, A.; Traversi, M.L.; Faraloni, C.; Valagussa, M.; Pozzi, A. Influence of biochar application on nutritional quality of tomato (Lycopersicon esculentum). Crop. Pasture Sci. 2015, 66, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Suthar, R.G.; Wang, C.; Nunes, M.C.N.; Chen, J.; Sargent, S.A.; Bucklin, R.A.; Gao, B. Bamboo biochar pyrolyzed at low temperature improves tomato plant growth and fruit quality. Agriculture 2018, 8, 153. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; Iapichino, G.; Mauro, R.P.; Consentino, B.B.; Pasquale, C.D. Poplar biochar as an alternative substrate for curly endive cultivated in a soilless system. Appl. Sci. 2020, 10, 1258. [Google Scholar] [CrossRef] [Green Version]
- Agegnehu, G.; Srivastava, A.K.; Bird, M.I. The role of biochar and biochar-compost in improving soil quality and crop performance: A review. Appl. Soil Ecol. 2017, 119, 156–170. [Google Scholar] [CrossRef]
- Graber, E.R.; Harel, Y.M.; Kolton, M.; Cytryn, E.; Silber, A.; David, D.R.; Tsechansky, L.; Borenshtein, M.; Elad, Y. Biochar impact on development and productivity of pepper and tomato grown in fertigated soilless media. Plant. Soil 2010, 337, 481–496. [Google Scholar] [CrossRef]
- Fascella, G.; Mammano, M.M.; D’Angiolillo, F.; Rouphael, Y. Effects of conifer wood biochar as a substrate component on ornamental performance, photosynthetic activity, and mineral composition of potted Rosa rugosa. J. Hortic. Sci. Biotechnol. 2018, 93, 519–528. [Google Scholar] [CrossRef]
- Fascella, G.; Mammano, M.M.; D’Angiolillo, F.; Pannico, A.; Rouphael, Y. Coniferous wood biochar as substrate component of two containerized Lavender species: Effects on morpho-physiological traits and nutrients partitioning. Sci. Hortic. 2020, 267, 109356. [Google Scholar] [CrossRef]
- Altland, J.E.; Locke, J.C. High rates of gasified rice hull biochar affect geranium and tomato growth in a soilless substrate. J. Plant. Nutr. 2017, 40, 1816–1828. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Prasad, M.; Kavanagh, A.; Tzortzakis, N. Biochar type and ratio as a peat additive/partial peat replacement in growing media for cabbage seedling production. Agronomy 2019, 9, 693. [Google Scholar] [CrossRef] [Green Version]
- Bedussi, F.; Zaccheo, P.; Crippa, L. Pattern of pore water nutrients in planted and non-planted soilless substrates as affected by the addition of biochars from wood gasification. Biol. Fertil. Soils 2015, 51, 625–635. [Google Scholar] [CrossRef]
- Choi, H.S.; Zhao, Y.; Dou, H.; Cai, X.; Gu, M.; Yu, F. Effects of biochar mixtures with pine-bark based substrates on growth and development of horticultural crops. Hortic Environ. Biotechnol 2018, 59, 345–354. [Google Scholar] [CrossRef]
- Judd, L.A.; Jackson, B.E.; Hesterberg, D.L.; Boyette, M.D.; Evans, M.R.; Fonteno, W.C. Assessing biochar as a lime replacement for peat substrates. Acta Hortic. 2019, 1266, 21–25. [Google Scholar] [CrossRef]
- Zaccheo, P.; Crippa, L.; Cattivello, C. Liming power of different particle fractions of biochar. Acta Hortic. 2014, 363–368. [Google Scholar] [CrossRef]
- Jahromi, N.B.; Fulcher, A.; Walker, F.; Altland, J.; Wright, W.; Eash, N. Evaluating on-demand irrigation systems for container-grown woody plants grown in biochar-amended pine bark. HortScience 2018, 53, 1891–1896. [Google Scholar] [CrossRef] [Green Version]
- Fornes, F.; Belda, R.M.; Fernández de Córdova, P.; Cebolla-Cornejo, J. Assessment of biochar and hydrochar as minor to major constituents of growing media for containerized tomato production. J. Sci. Food Agric. 2017, 97, 3675–3684. [Google Scholar] [CrossRef] [PubMed]
- Camposeco-Montejo, N.; Robledo-Torres, V.; Ramírez-Godina, F.; Mendoza-Villarreal, R.; Pérez-Rodríguez, M.Á.; Cabrera-de la Fuente, M. Response of bell pepper to rootstock and greenhouse cultivation in coconut fiber or soil. Agronomy 2018, 8, 111. [Google Scholar] [CrossRef] [Green Version]
- Jordan, R.A.; Ribeiro, E.F.; de Oliveira, F.C.; Geisenhoff, L.O.; Martins, E.A.S. Yield of lettuce grown in hydroponic and aquaponic systems using different substrates. Rev. Bras. Eng. Agric. Ambient. 2018, 22, 525–529. [Google Scholar] [CrossRef] [Green Version]
- Beozzi, S.; Cabral, F.; Vasconcelos, E.; Ribeiro, H.M. Organic production of potted parsley and coriander in coconut coir amended with compost. Acta Hortic. 2017, 1168, 295–302. [Google Scholar] [CrossRef]
- Kingston, P.H.; Scagel, C.F.; Bryla, D.R.; Strik, B. Suitability of sphagnum moss, coir, and douglas fir bark as soilless substrates for container production of highbush blueberry. HortScience 2017, 52, 1692–1699. [Google Scholar] [CrossRef]
- Paradiso, R.; De Pascale, S. Effects of coco fibre addition to perlite on growth and yield of cut gerbera. Acta Hortic. 2008, 779, 529–534. [Google Scholar] [CrossRef]
- Paillat, L.; Guénon, R.; Huché-Thélier, L.; Barraud, F.; Cannavo, P. Growing media and organic fertilizers affect microbial activities and nutrient availability in soilless horticulture. Acta Hortic. 2020, 1296, 791–798. [Google Scholar] [CrossRef]
- Xing, J.; Gruda, N.; Xiong, J.; Liu, W. Influence of organic substrates on nutrient accumulation and proteome changes in tomato-roots. Sci. Hortic. 2019, 252, 192–200. [Google Scholar] [CrossRef]
- Van Labeke, M.C.; Dambre, P. Gerbera cultivation on coir with recirculation of the nutrient solution: A comparison with rockwool culture. Acta Hortic. 1998, 458, 357–362. [Google Scholar] [CrossRef]
- Del Amor, F.M.; Gómez-López, M.D. Agronomical response and water use efficiency of sweet pepper plants grown in different greenhouse substrates. HortScience 2009, 44, 810–814. [Google Scholar] [CrossRef] [Green Version]
- Linderman, R.G.; Davis, E.A. Arbuscular mycorrhiza and growth responses of several ornamental plants grown in soilless peat-based medium amended with coconut dust (coir). Horttechnology 2003, 13, 482–487. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, I.; Konieczny, A.; Gąstoł, M.; Sady, W.; Hanus-Fajerska, E. Effect of mycorrhiza and phosphorus content in nutrient solution on the yield and nutritional status of tomato plants grown on rockwool or coconut coir. Agric. Food Sci. 2015, 24, 39–51. [Google Scholar] [CrossRef]
- Boyer, L.R.; Feng, W.; Gulbis, N.; Hajdu, K.; Harrison, R.J.; Jeffries, P.; Xu, X. The use of arbuscular mycorrhizal fungi to improve strawberry production in coir substrate. Front. Plant. Sci. 2016, 7, 1–9. [Google Scholar]
- Pant, A.P.; Radovich, T.J.K.; Hue, N.V.; Paull, R.E. Biochemical properties of compost tea associated with compost quality and effects on pak choi growth. Sci. Hortic. 2012, 148, 138–146. [Google Scholar] [CrossRef]
- Vandecasteele, B.; Boogaerts, C.; Vandaele, E. Combining woody biomass for combustion with green waste composting: Effect of removal of woody biomass on compost quality. Waste Manag. 2016, 58, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Hurley, S.; Shrestha, P.; Cording, A. Nutrient leaching from compost: Implications for bioretention and other green stormwater infrastructure. J. Sustain. Water Built Environ. 2017, 3, 04017006. [Google Scholar] [CrossRef]
- Pane, C.; Maria, A.; Spaccini, R.; Piccolo, A.; Celano, G.; Zaccardelli, M. Enhancing sustainability of a processing tomato cultivation system by using bioactive compost teas. Sci. Hortic. 2016, 202, 117–124. [Google Scholar] [CrossRef]
- Chong, C. Experiences with wastes and composts in nursery substrates. Horttechnology 2005, 15, 739–747. [Google Scholar] [CrossRef]
- Pane, C.; Palese, A.M.; Celano, G.; Zaccardelli, M. Effects of compost tea treatments on productivity of lettuce and kohlrabi systems under organic cropping management. Ital. J. Agron. 2014, 9, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Zaccardelli, M.; Pane, C.; Villecco, D.; Palese, A.M.; Celano, G. Compost tea spraying increases yield performance of pepper (Capsicum annuum L.) grown in greenhouse under organic farming system. Ital. J. Agron. 2018, 13, 229–234. [Google Scholar] [CrossRef]
- Copetta, A.; Bardi, L.; Bertolone, E.; Berta, G. Fruit production and quality of tomato plants (Solanum lycopersicum L.) are affected by green compost and arbuscular mycorrhizal fungi. Plant. Biosyst. 2011, 145, 106–115. [Google Scholar] [CrossRef]
- Morales-Corts, M.R.; Gómez-Sánchez, M.Á.; Pérez-Sánchez, R. Evaluation of green/pruning wastes compost and vermicompost, slumgum compost and their mixes as growing media for horticultural production. Sci. Hortic. 2014, 172, 155–160. [Google Scholar] [CrossRef]
- Bernal-Vicente, A.; Ros, M.; Tittarelli, F.; Intrigliolo, F.; Pascual, J.A. Citrus compost and its water extract for cultivation of melon plants in greenhouse nurseries. Evaluation of nutriactive and biocontrol effects. Bioresour. Technol. 2008, 99, 8722–8728. [Google Scholar] [CrossRef]
- Spiers, T.M.; Fietje, G. Green waste compost as a component in soilless growing media. Compost Sci. Util. 2000, 8, 19–23. [Google Scholar] [CrossRef]
- Rainbow, A. The use of green compost in the production of container nursery stock in the UK: Challenges and opportunities. Acta Hortic. 2009, 819, 27–32. [Google Scholar] [CrossRef]
- Witcher, A.L.; Pickens, J.M.; Blythe, E.K. Container color and compost substrate affect root zone temperature and growth of “green Giant” arborvitae. Agronomy 2020, 10, 484. [Google Scholar] [CrossRef] [Green Version]
- Graceson, A.; Hare, M.; Monaghan, J.; Hall, N. The water retention capabilities of growing media for green roofs. Ecol. Eng. 2013, 61, 328–334. [Google Scholar] [CrossRef]
- Gruda, N.; Schnitzler, W.H. Suitability of wood fiber substrate for production of vegetable transplants: I. Physical properties of wood fiber substrates. Sci. Hortic. 2004, 100, 309–322. [Google Scholar] [CrossRef]
- Boyer, C.R.; Gilliam, C.H.; Fain, G.B.; Gallagher, T.V.; Torbert, H.A.; Sibley, J.L. Production of woody nursery crops in clean chip residual substrate. J. Environ. Hortic. 2009, 27, 56–62. [Google Scholar] [CrossRef]
- Jackson, B.E.; Wright, R.D.; Browder, J.F.; Harris, J.R.; Niemiera, A.X. Effect of fertilizer rate on growth of azalea and holly in pine bark and pine tree substrates. HortScience 2008, 43, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Domeño, I.; Irigoyen, N.; Muro, J. Evolution of organic matter and drainages in wood fibre and coconut fibre substrates. Sci. Hortic. 2009, 122, 269–274. [Google Scholar] [CrossRef]
- Gruda, N.; von Tucher, S.; Schnitzler, W.H. N-immobilization by wood fibre substrates in the production of tomato transplants (Lycopersicon lycopersicum (L.) Karst. ex Farw.). Angew. Bot. 2000, 74, 32–37. [Google Scholar]
- Burgel, L.; Hartung, J.; Graeff-Hönninger, S. Impact of different growing substrates on growth, yield and cannabinoid content of two Cannabis sativa L. genotypes in a pot culture. Horticulturae 2020, 6, 62. [Google Scholar] [CrossRef]
- Frangi, P.; Amoroso, G.; Ferrini, F.; Fini, A. Growth of ornamental shrubs in wood fibre-based growing media. Acta Hortic. 2008, 801, 1571–1575. [Google Scholar] [CrossRef]
- Handreck, K.; Black, N. Growing Media for Ornamental Plants and Turf, 3rd ed.; University of New South Wales Press: Sydney, NSW, Australia, 2002. [Google Scholar]
- Caron, J.; Elrick, D.E.; Beeson, R.; Boudreau, J. Defining Critical Capillary Rise Properties for Growing Media in Nurseries. Soil Sci. Soc. Am. J. 2005, 69, 794–806. [Google Scholar] [CrossRef]
- Jahromi, N.B.; Walker, F.; Fulcher, A.; Altland, J.; Wright, W.C. Growth response, mineral nutrition, and water utilization of containergrown woody ornamentals grown in biochar-amended pine bark. HortScience 2018, 53, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Olle, M.; Ngouajio, M.; Siomos, A. Vegetable quality and productivity as influenced by growing medium: A review. Zemdirbyste 2012, 99, 399–408. [Google Scholar]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant. Sci. 2018, 8, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant. Sci. 2020, 11, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Rouphael, Y. Biostimulants in horticulture. Sci. Hortic. 2015, 196, 1–2. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systems. Italus Hortus 2018, 25, 27–36. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Andersen, M.N.; Liu, F. Biochar mitigates salinity stress in potato. J. Agron. Crop. Sci. 2015, 201, 368–378. [Google Scholar] [CrossRef]
- Pane, C.; Villecco, D.; Campanile, F.; Zaccardelli, M. Novel strains of Bacillus, isolated from compost and compost-amended soils, as biological control agents against soil-borne phytopathogenic fungi. Biocontrol Sci. Technol. 2012, 22, 1373–1388. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Sorrentino, R.; Caputo, M.; Scotti, R.; De Falco, E.; Pane, C. Stepwise-selected Bacillus amyloliquefaciens and B. Subtilis strains from composted aromatic plant waste able to control soil-borne diseases. Agriculture 2020, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Chilosi, G.; Aleandri, M.P.; Luccioli, E.; Stazi, S.R.; Marabottini, R.; Morales-Rodríguez, C.; Vettraino, A.M.; Vannini, A. Suppression of soil-borne plant pathogens in growing media amended with espresso spent coffee grounds as a carrier of Trichoderma spp. Sci. Hortic. 2020, 259, 108666. [Google Scholar] [CrossRef]
- Schnitzler, W.H. Pest and disease management of soilless culture. Acta Hortic. 2004, 648, 191–203. [Google Scholar] [CrossRef]
- Postma, J.; Willemsen-de Klein, M.J.E.I.M.; Rattink, H.; van Os, E.A. Disease suppressive soilless culture systems; Characterisation of its micloflora. Acta Hortic. 2001, 554, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Duffy, B.K.; Défago, G. Macro- and microelement fertilizers influence the severity of Fusarium crown and root rot of tomato in a soilless production system. HortScience 1999, 34, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Abro, M.A.; Syed, R.N.; Yasmin, A.; Abro, S.; Kumar, A.; Nicot, P.C. Different nitrogen fertilization regimes influences the susceptibility of lettuce plants to Botrytis cinerea and Sclerotinia sclerotiorum. Sci. Int. 2014, 26, 325–329. [Google Scholar]
- Martínez, F.; Castillo, S.; Carmona, E.; Avilés, M. Dissemination of Phytophthora cactorum, cause of crown rot in strawberry, in open and closed soilless growing systems and the potential for control using slow sand filtration. Sci. Hortic. 2010, 125, 756–760. [Google Scholar] [CrossRef]
- Segarra, G.; Casanova, E.; Avilés, M.; Trillas, I. Trichoderma asperellum strain T34 controls Fusarium wilt disease in tomato plants in soilless culture through competition for iron. Microb. Ecol. 2010, 59, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Pane, C.; Spaccini, R.; Piccolo, A.; Scala, F.; Bonanomi, G. Compost amendments enhance peat suppressiveness to Pythium ultimum, Rhizoctonia solani and Sclerotinia minor. Biol. Control. 2011, 56, 115–124. [Google Scholar] [CrossRef]
- Bonanomi, G.; Antignani, V.; Pane, C.; Scala, F. Suppression of soilborn fungal disease with organic amendments. J. Plant. Pathol. 2007, 89, 311–324. [Google Scholar]
- Pane, C.; Piccolo, A.; Spaccini, R.; Celano, G.; Villecco, D.; Zaccardelli, M. Agricultural waste-based composts exhibiting suppressivity to diseases caused by the phytopathogenic soil-borne fungi Rhizoctonia solani and Sclerotinia minor. Appl. Soil Ecol. 2013, 65, 43–51. [Google Scholar] [CrossRef]
- Pane, C.; Chiantese, C.; Scala, F.; Bonanomi, G. Assessment of gardening growing media suppressiveness against Rhizoctonia damping-off disease. J. Plant. Pathol. 2013, 95, 401–405. [Google Scholar]
- Tenuta, M.; Lazarovits, G. Ammonia and nitrous acid from nitrogenous amendments kill the microsclerotia of Verticillium dahliae. Phytopathology 2002, 92, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linderman, R.G.; Davis, E.A.; Masters, C.J. Response of conifer seedlings to potting medium amendment with meadowfoam seed meal. Issues New Crop. New Uses 2007, 138–142. [Google Scholar]
- Ayanbimpe, G.M.; Ojo, T.K.; Afolabi, E.; Opara, F.; Orsaah, S.; Ojerinde, O.S. Evaluation of extracts of Jatropha curcas and Moringa oleifera in culture media for selective inhibition of saprophytic fungal contaminants. J. Clin. Lab. Anal. 2009, 23, 161–164. [Google Scholar] [CrossRef]
- Fields, J.S.; Owen, J.S.; Altland, J.E.; van Iersel, M.W.; Jackson, B.E. Soilless substrate hydrology can be engineered to influence plant water status for an ornamental containerized crop grown within optimal water potentials. J. Am. Soc. Hortic. Sci. 2018, 143, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Farrell, C.; Cao, C.T.N.; Ang, X.Q.; Rayner, J.P. Use of water-retention additives to improve performance of green roof substrates. Acta Hortic. 2016, 1108, 271–278. [Google Scholar] [CrossRef]
- Mulcahy, D.N.; Mulcahy, D.L.; Dietz, D. Biochar soil amendment increases tomato seedling resistance to drought in sandy soils. J. Arid Environ. 2013, 88, 222–225. [Google Scholar] [CrossRef]
- Di Lonardo, S.; Baronti, S.; Vaccari, F.P.; Albanese, L.; Battista, P.; Miglietta, F.; Bacci, L. Biochar-based nursery substrates: The effect of peat substitution on reduced salinity. Urban For. Urban Green. 2017, 23, 27–34. [Google Scholar] [CrossRef]
- De Tender, C.A.; Debode, J.; Vandecasteele, B.; D’Hose, T.; Cremelie, P.; Haegeman, A.; Ruttink, T.; Dawyndt, P.; Maes, M. Biological, physicochemical and plant health responses in lettuce and strawberry in soil or peat amended with biochar. Appl. Soil Ecol. 2016, 107, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Braud, A.; Sautel, M.; Afonso, S.; Beaudet, E.; Grosbellet, C. Cucumber culture on wood fiber substrate amended with Greenchar®. Acta Hortic. 2019, 1266, 103–108. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Frenkel, O.; Tsechansky, L.; Elad, Y.; Graber, E.R. Immobilization and deactivation of pathogenic enzymes and toxic metabolites by biochar: A possible mechanism involved in soilborne disease suppression. Soil Biol. Biochem. 2018, 121, 59–66. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Elad, Y.; Cytryn, E.; Graber, E.R.; Frenkel, O. Activating biochar by manipulating the bacterial and fungal microbiome through pre-conditioning. New Phytol. 2018, 219, 363–377. [Google Scholar] [CrossRef] [Green Version]
- Frenkel, O.; Jaiswal, A.K.; Elad, Y.; Lew, B.; Kammann, C.; Graber, E.R. The effect of biochar on plant diseases: What should we learn while designing biochar substrates? J. Environ. Eng. Landsc. Manag. 2017, 25, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Elad, Y.; David, D.R.; Harel, Y.M.; Borenshtein, M.; Kalifa, H.B.; Silber, A.; Graber, E.R. Induction of systemic resistance in plants by biochar, a soil-applied carbon sequestering agent. Phytopathology 2010, 100, 913–921. [Google Scholar] [CrossRef] [Green Version]
- Meller Harel, Y.; Elad, Y.; Rav-David, D.; Borenstein, M.; Shulchani, R.; Lew, B.; Graber, E.R. Biochar mediates systemic response of strawberry to foliar fungal pathogens. Plant. Soil 2012, 357, 245–257. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Graber, E.R.; Elad, Y.; Frenkel, O. Biochar as a management tool for soilborne diseases affecting early stage nursery seedling production. Crop. Prot. 2019, 120, 34–42. [Google Scholar] [CrossRef]
- Copley, T.R.; Aliferis, K.A.; Jabaji, S. Maple bark biochar affects Rhizoctonia solani metabolism and increases damping-off severity. Phytopathology 2015, 105, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- San Bautista, A.; Rueda, R.; Pascual, B.; Maroto, J.V.; López-Galarza, S. Influence of different substrates and nutrient solutions on the yields and the incidence of abiotic disorders of broccoli. Acta Hortic. 2005, 697, 275–280. [Google Scholar] [CrossRef]
- Gómez-Bellot, M.J.; Lorente, B.; Sánchez-Blanco, M.J.; Ortuño, M.F.; Nortes, P.A.; Alarcón, J.J. Influence of mixed substrate and arbuscular mycorrhizal fungi on photosynthetic efficiency, nutrient and water status and yield in tomato plants irrigated with saline reclaimed waters. Water 2020, 12, 438. [Google Scholar] [CrossRef] [Green Version]
- Shameer, S.; Prasad, T.N.V.K.V. Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant. Growth Regul. 2018, 84, 603–615. [Google Scholar] [CrossRef]
- Maboko, M.M.; Bertling, I.; du Plooy, C.P. Arbuscular mycorrhiza has limited effects on yield and quality of tomatoes grown under soilless cultivation. Acta Agric. Scand. Sect. B Soil Plant. Sci. 2013, 63, 261–270. [Google Scholar] [CrossRef]
- Waller, P.L.; Thornton, C.R.; Farley, D.; Groenhof, A. Pathogens and other fungi in growing media constituents. Acta Hortic. 2008, 779, 361–366. [Google Scholar] [CrossRef]
- Hyder, N.; Sims, J.J.; Wegulo, S.N. In vitro suppression of soilborne plant pathogens by coir. Horttechnology 2009, 19, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Sriram, S.; Savitha, M.J.; Ramanujam, B. Trichoderma-enriched coco-peat for the management of Phytophthora and Fusarium diseases of chilli and tomato in nurseries. J. Biol. Control. 2010, 24, 311–316. [Google Scholar]
- Kipngeno, P.; Losenge, T.; Maina, N.; Kahangi, E.; Juma, P. Efficacy of Bacillus subtilis and Trichoderma asperellum against Pythium aphanidermatum in tomatoes. Biol. Control. 2015, 90, 92–95. [Google Scholar] [CrossRef]
- Jalali, Z.; Shoor, M.; Nemati, S.H.; Rouhany, H. Trichoderma harzianum and Fe spray improve growth properties of Spathiphyllum sp. J. Ornam. Plants 2014, 4, 145–152. [Google Scholar]
- Fiasconaro, M.L.; Lovato, M.E.; Antolín, M.C.; Clementi, L.A.; Torres, N.; Gervasio, S.; Martín, C.A. Role of proline accumulation on fruit quality of pepper (Capsicum annuum L.) grown with a K-rich compost under drought conditions. Sci. Hortic. 2019, 249, 280–288. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K.; et al. Understanding the biostimulant action of vegetal-derived protein hydrolysates by high-throughput plant phenotyping and metabolomics: A case study on tomato. Front. Plant. Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Trevisan, S.; Francioso, O.; Quaggiotti, S.; Nardi, S. Humic substances biological activity at the plant-soil interface: From environmental aspects to molecular factors. Plant. Signal. Behav. 2010, 5, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Altissimo, A.; Franceschi, C.; Nardi, S. Phenol-containing organic substances stimulate phenylpropanoid metabolism in Zea mays. J. Plant. Nutr. Soil Sci. 2011, 174, 496–503. [Google Scholar] [CrossRef]
- Ros, M.; Hernandez, M.T.; Garcia, C.; Bernal, A.; Pascual, J.A. Biopesticide effect of green compost against Fusarium wilt on melon plants. J. Appl. Microbiol. 2005, 98, 845–854. [Google Scholar] [CrossRef]
- Yogev, A.; Raviv, M.; Hadar, Y.; Cohen, R.; Katan, J. Plant waste-based composts suppressive to diseases caused by pathogenic Fusarium oxysporum. Eur. J. Plant. Pathol. 2006, 116, 267–278. [Google Scholar] [CrossRef]
- De Corato, U.; Patruno, L.; Avella, N.; Lacolla, G.; Cucci, G. Composts from green sources show an increased suppressiveness to soilborne plant pathogenic fungi: Relationships between physicochemical properties, disease suppression, and the microbiome. Crop. Prot. 2019, 124, 104870. [Google Scholar] [CrossRef]
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144. [Google Scholar] [CrossRef]
- Pane, C.; Spaccini, R.; Piccolo, A.; Celano, G.; Zaccardelli, M. Disease suppressiveness of agricultural greenwaste composts as related to chemical and bio-based properties shaped by different on-farm composting methods. Biol. Control. 2019, 137, 104026. [Google Scholar] [CrossRef]
- Malandraki, I.; Tjamos, S.E.; Pantelides, I.S.; Paplomatas, E.J. Thermal inactivation of compost suppressiveness implicates possible biological factors in disease management. Biol. Control. 2008, 44, 180–187. [Google Scholar] [CrossRef]
- Hoitink, H.A.J.; Boehm, M.J. Biocontrol within the context of soil microbial communities: A substrate-dependent phenomenon. Annu. Rev. Phytopathol. 1999, 37, 427–446. [Google Scholar] [CrossRef]
- Pane, C.; Sorrentino, R.; Scotti, R.; Molisso, M.; Di Matteo, A.; Celano, G.; Zaccardelli, M. Alpha and beta-diversity of microbial communities associated to plant disease suppressive functions of on-farm green composts. Agriculture 2020, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Scotti, R.; Mitchell, A.L.; Pane, C.; Finn, R.D.; Zaccardelli, M. Microbiota characterization of agricultural green waste-based suppressive composts using omics and classic approaches. Agriculture 2020, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- De Corato, U.; Salimbeni, R.; De Pretis, A.; Patruno, L.; Avella, N.; Lacolla, G.; Cucci, G. Microbiota from ‘next-generation green compost’ improves suppressiveness of composted Municipal-Solid-Waste to soil-borne plant pathogens. Biol. Control. 2018, 124, 1–17. [Google Scholar] [CrossRef]
- St. Martin, C.C.G.; Bekele, I.; Eudoxie, G.D.; Bristol, D.; Brathwaite, R.A.I.; Campo, K.R. Modelling response patterns of physico-chemical indicators during high-rate composting of green waste for suppression of Pythium ultimum. Environ. Technol. 2014, 35, 590–601. [Google Scholar] [CrossRef]
- Scheuerell, S.J.; Sullivan, D.M.; Mahaffee, W.F. Suppression of seedling damping-off caused by Pythium ultimum, P. irregulare, and Rhizoctonia solani in container media amended with a diverse range of Pacific Northwest compost sources. Phytopathology 2005, 95, 306–315. [Google Scholar] [CrossRef] [Green Version]
- Burger, D.W.; Hartz, T.K.; Forister, G.W. Composted green waste as a container medium amendment for the production of ornamental plants. HortScience 1997, 32, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Chilosi, G.; Aleandri, M.P.; Bruni, N.; Tomassini, A.; Torresi, V.; Muganu, M.; Paolocci, M.; Vettraino, A.; Vannini, A. Assessment of suitability and suppressiveness of on-farm green compost as a substitute of peat in the production of lavender plants. Biocontrol Sci. Technol. 2017, 27, 539–555. [Google Scholar] [CrossRef]
- van der Gaag, D.J.; van Noort, F.R.; Stapel-Cuijpers, L.H.M.; de Kreij, C.; Termorshuizen, A.J.; van Rijn, E.; Zmora-Nahum, S.; Chen, Y. The use of green waste compost in peat-based potting mixtures: Fertilization and suppressiveness against soilborne diseases. Sci. Hortic. 2007, 114, 289–297. [Google Scholar] [CrossRef]
- De Corato, U.; Salimbeni, R.; De Pretis, A. Suppression of soil-borne pathogens in container media amended with on-farm composted agro-bioenergy wastes and residues under glasshouse condition. J. Plant. Dis. Prot. 2018, 125, 213–226. [Google Scholar] [CrossRef]
- Blaya, J.; López-Mondéjar, R.; Lloret, E.; Pascual, J.A.; Ros, M. Changes induced by Trichoderma harzianum in suppressive compost controlling Fusarium wilt. Pestic. Biochem. Physiol. 2013, 107, 112–119. [Google Scholar] [CrossRef] [PubMed]
- López-López, N.; Segarra, G.; Vergara, O.; López-Fabal, A.; Trillas, M.I. Compost from forest cleaning green waste and Trichoderma asperellum strain T34 reduced incidence of Fusarium circinatum in Pinus radiata seedlings. Biol. Control. 2016, 95, 31–39. [Google Scholar] [CrossRef]
- Wolna-Maruwka, A.; Piechota, T.; Niewiadomska, A.; Dach, J.; Szczech, M.; Jȩdryczka, M.; Pilarska, A.A. An assessment of adaptive and antagonistic properties of Trichoderma sp. strains in vegetable waste composts. Arch. Environ. Prot. 2017, 43, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N. Sustainable peat alternative growing media. Acta Hortic. 2012, 927, 973–980. [Google Scholar] [CrossRef]
- Van Gerrewey, T.; Ameloot, N.; Navarrete, O.; Vandecruys, M.; Perneel, M.; Boon, N.; Geelen, D. Microbial activity in peat-reduced plant growing media: Identifying influential growing medium constituents and physicochemical properties using fractional factorial design of experiments. J. Clean. Prod. 2020, 256, 120323. [Google Scholar] [CrossRef]
- Ehret, D.L.; Helmer, T. A new wood fibre substrate for hydroponie tomato and pepper crops. Can. J. Plant. Sci. 2009, 89, 1127–1132. [Google Scholar] [CrossRef] [Green Version]
- Vandecasteele, B.; Muylle, H.; De Windt, I.; Van Acker, J.; Ameloot, N.; Moreaux, K.; Coucke, P.; Debode, J. Plant fibers for renewable growing media: Potential of defibration, acidification or inoculation with biocontrol fungi to reduce the N drawdown and plant pathogens. J. Clean. Prod. 2018, 203, 1143–1154. [Google Scholar] [CrossRef]


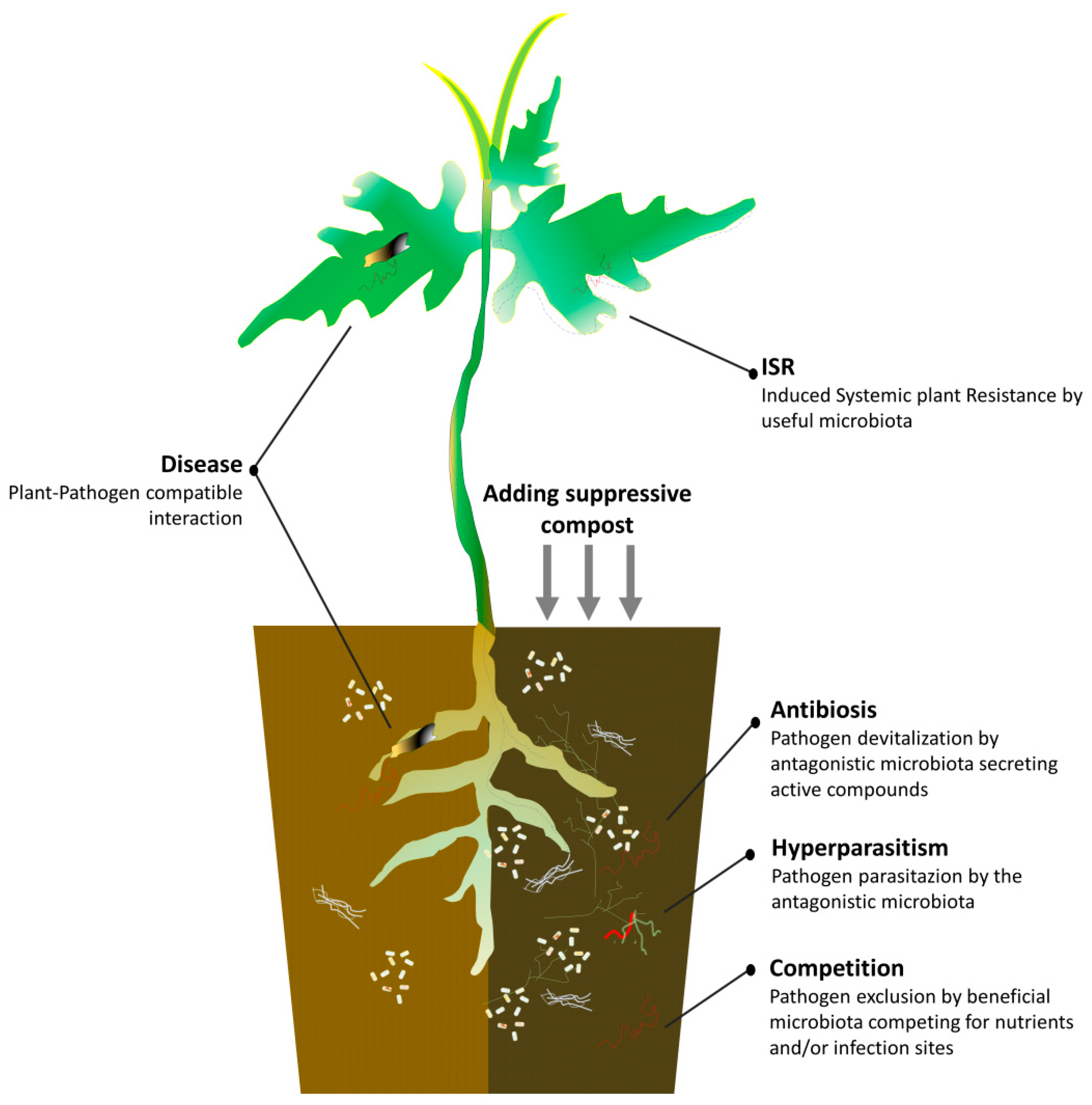{kind=link}
| Parameters | Optimal Range | Biochar | Coir | Green Compost | Wood Fiber |
|---|---|---|---|---|---|
| C-to-N ratio | 15–20 [84,85] | High [86,87] | High [38] | Optimal/high depending on raw material [15,88] | High [15,89] |
| pH | 5.5–6.5 [90] | Generally high [15,34] | Optimal, but can vary much (4.9–6.9) [15,38] | High [15] | Optimal [15,37] |
| EC | <0.5 dS m−1 (1:1.5 v v−1 water extract) [90] | Generally optimal [34,91] | High, but variable depending on washing and/or leaching treatment [15,38] | High [15] | Optimal [15,37] |
| CEC | 0–50 meq 100 g−1 [90] | Generally high, but depending on raw material and processing [87] | Low [15,38] | Generally high [88] | Optimal [15] |
| Nutrient content | Low (negligible) [15] | Low, except for K, Ca, and Mg (depending on particle size) [34] | Low N and P, high K [15,38] | High [3,15] | Low, but depending on plant species [15,89] |
| Bulk density | <0.40 g cm−3 [90] | Optimal [28] | Optimal [15,90] | Generally optimal [92] | Optimal [15,24] |
| Total porosity | >85 % v v−1 [90] | Optimal [91] | Optimal [15,90] | Generally low [15,37] | Optimal [15] |
| Available water 1 | 20–30 % v v−1 [90] | Variable depending on the particle size [28,91] | Optimal [15,90] | Generally low [15,37] | Generally low [15,37] |
| Air capacity | 20–30 % v v−1 [90] | Variable depending on the particle size [28] | Variable, optimal on the average [15,90] | Optimal [6] | Usually high [15,24] |
| Wettability 2 | <60 [44] | Optimal [28] | Optimal [15,44] | Optimal [93] | Optimal [24,44] |
| Weed, pest, and disease | None [44] | Optimal–low [28] | Optimal–low [15] | Optimal–low [15,24] | Optimal [3,15] |
| Physical and biological stability | High [3,37,94] | Optimal [28] | Optimal [15] | Variable depending on raw material [24] | Optimal, but tends to compress easily [3,15,89] |
| Disease suppressive properties | Desirable [95] | Low–medium [28] | Low [15,96,97] | High [3,98,99] | Always low [100,101,102] |
| Availability (technical) | Desirable at local/national level [24,93] | Locally available, but low quantity [28,91] | High quantity, availability can be limited depending on production sites [37] | Locally available, but low quantity [3] | Locally available, but low quantity [3] |
| Standardization and quality | High [103] | Generally low depending on raw material [91] | Generally optimal [15] | Variable depending on raw material [3,24,37] | Generally medium depending on raw material [3,15] |
| Price 3 | ~50 € m−3 | High [28] | Optimal–high | Low [24] | Optimal–high [3] |
| N | P | K | Source |
|---|---|---|---|
| (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | |
| 12.3–15.2 | 0.2–0.48 | 12.1–19.9 | [70] |
| 12.2 | 2.6 | 7.3 | [142] |
| 16.3 | Na | Na | [76] |
| Na | 1.8–2.5 | 5.6–11.1 | [143] |
| 15.4 | 3.1 | 8.6 | [144] |
| Element | Green Compost | Extraction Ratio | Compost Tea | Element Extracted from 1 L of Compost | Extraction Efficiency | Source | ||
|---|---|---|---|---|---|---|---|---|
| Compost | Extractant (Water) | |||||||
| (g kg−1 DW) | (mg L sub−1) | (L) | (L) | (mg L−1) | (mg) | (%) | ||
| N | 12.2 | 4270 | 1 | 10 | 9.5 | 95 | 2.2 | [142] |
| P | 2.6 | 910 | 1 | 10 | 3 | 30 | 3.3 | |
| K | 7.3 | 2555 | 1 | 10 | 196.6 | 1966 | 76.9 | |
| Ca | 66.1 | 23,135 | 1 | 10 | 48.7 | 487 | 2.1 | |
| Mg | 7.4 | 2590 | 1 | 10 | 21.2 | 212 | 8.2 | |
| N | 14.1 | Na | Na | [70,145] | ||||
| P | 0.5 | Na | Na | |||||
| K | 19.9 | 6965 | 1 | 4 | 960.6 | 3842.2 | 55.2 | |
| Ca | 44.3 | 15,505 | 1 | 4 | 51.5 | 205.8 | 1.3 | |
| Mg | 11.8 | 4130 | 1 | 4 | 48.9 | 195.6 | 4.7 | |
| Na | 2.2 | 770 | 1 | 4 | 39.8 | 159 | 20.6 | |
| N | 15.2 | Na | Na | [70,145] | ||||
| P | 0.5 | Na | Na | |||||
| K | 19.2 | 6720 | 1 | 4 | 935.8 | 3743 | 55.7 | |
| Ca | 49 | 17,150 | 1 | 4 | 51.6 | 206.4 | 1.2 | |
| Mg | 12.3 | 4305 | 1 | 4 | 44.7 | 178.8 | 4.2 | |
| Na | 1.5 | 525 | 1 | 4 | 25.3 | 101 | 19.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atzori, G.; Pane, C.; Zaccardelli, M.; Cacini, S.; Massa, D. The Role of Peat-Free Organic Substrates in the Sustainable Management of Soilless Cultivations. Agronomy 2021, 11, 1236. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061236
Atzori G, Pane C, Zaccardelli M, Cacini S, Massa D. The Role of Peat-Free Organic Substrates in the Sustainable Management of Soilless Cultivations. Agronomy. 2021; 11(6):1236. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061236
Chicago/Turabian StyleAtzori, Giulia, Catello Pane, Massimo Zaccardelli, Sonia Cacini, and Daniele Massa. 2021. "The Role of Peat-Free Organic Substrates in the Sustainable Management of Soilless Cultivations" Agronomy 11, no. 6: 1236. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061236