
Characterization of Mollisols after Long-Term N Fertilization at Successive Rates in Continuous and Rotated Corn Systems



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site Description
2.2. Treatments and Field Management Practices
2.3. Soil Sampling and Determinations
2.4. Statistical Analyses
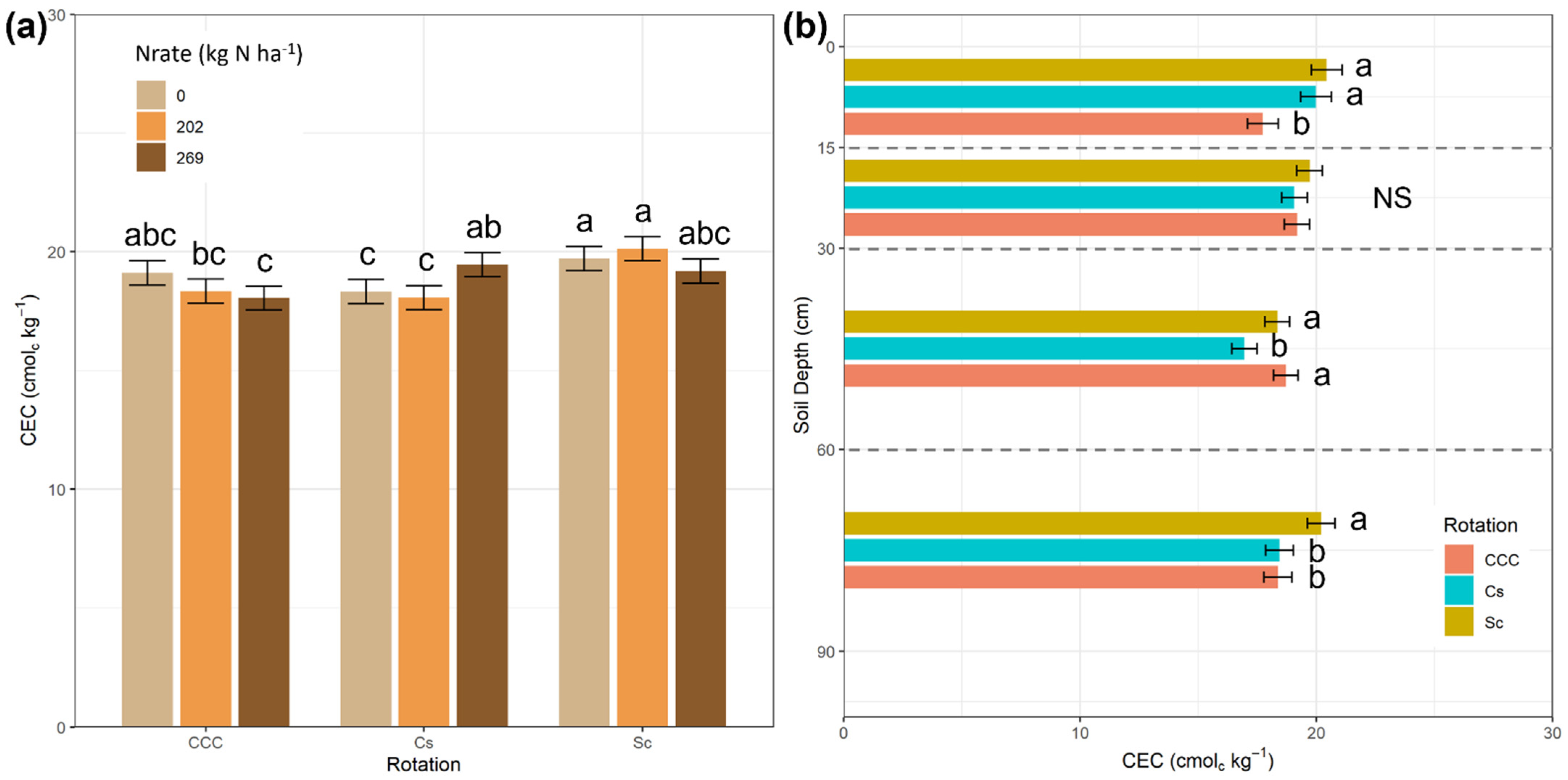
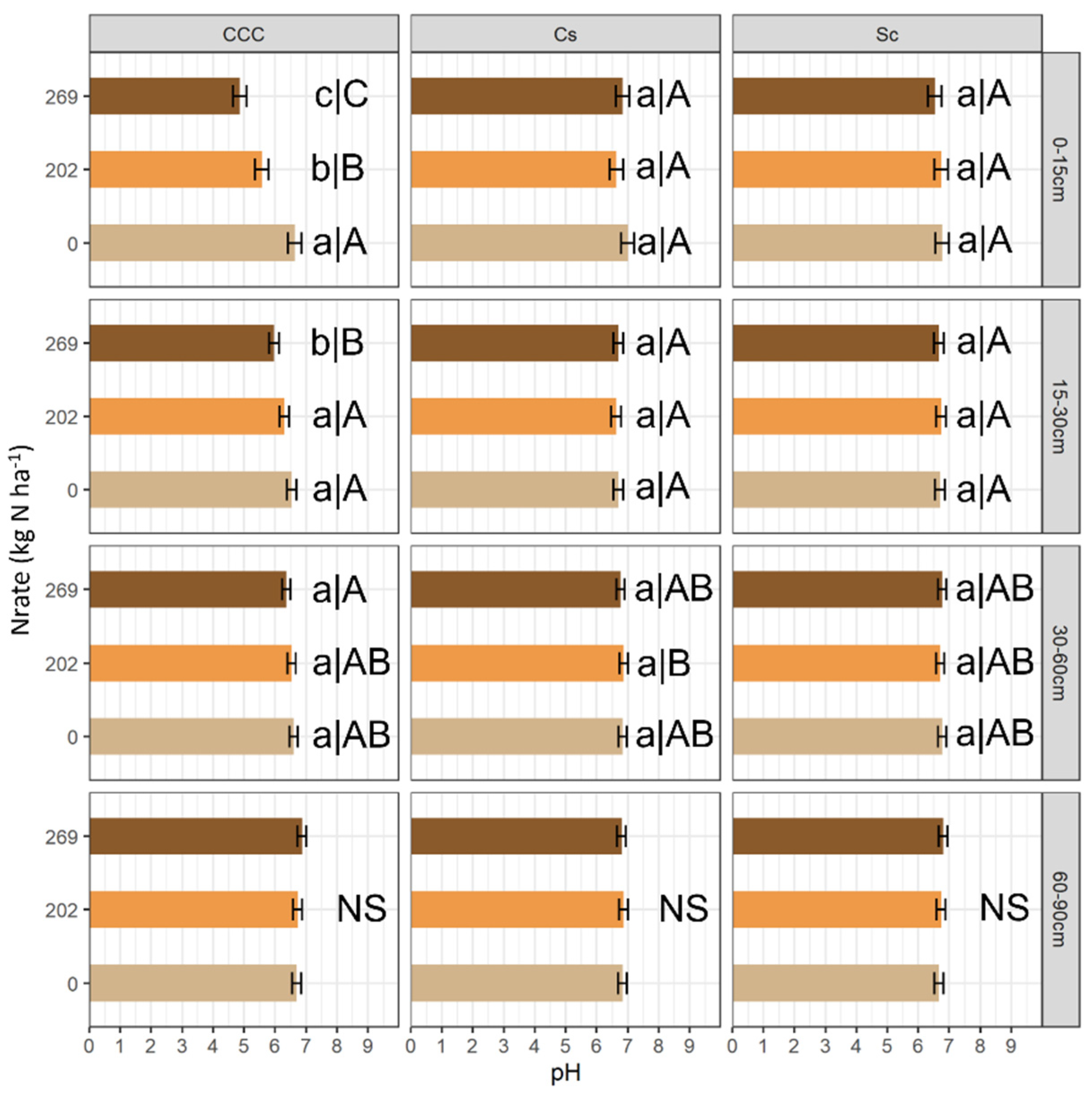
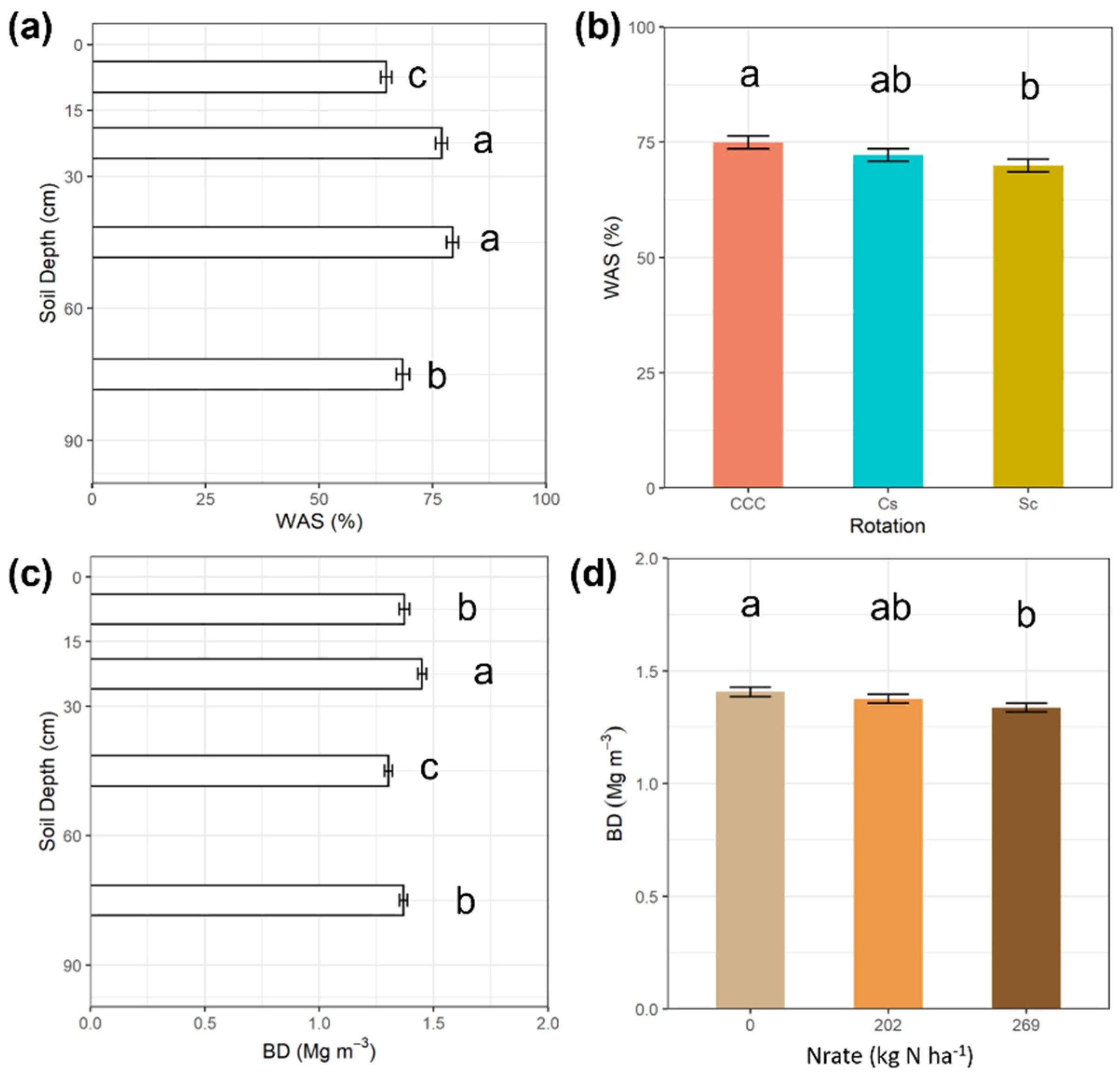
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Datnoff, L.E.; Elmer, W.H.; Huber, D.M. Mineral Nutrition and Plant Disease; American Phytopathological Society: St. Paul, MN, USA, 2007; p. 278. [Google Scholar]
- FAO. World Fertilizer Trends and Outlook to 2022; FAO: Rome, Italy, 2019. [Google Scholar]
- Liu, J.; You, L.; Amini, M.; Obersteiner, M.; Herrero, M.; Zehnder, A.J.; Yang, H. A high-resolution assessment on global nitrogen flows in cropland. Proc. Natl. Acad. Sci. USA 2010, 107, 8035–8040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- USDA-NASS. Crop Production; USDA-NASS: Washington, DC, USA, 2020.
- USDA-ERS (Ed.) Fertilizer Use and Price; USDA-ERS: Washington, DC, USA, 2020.
- Hatfield, J.L.; Wright-Morton, L.; Hall, B. Vulnerability of grain crops and croplands in the Midwest to climatic variability and adaptation strategies. Clim. Change 2018, 146, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Socolar, Y.; Goldstein, B.R.; de Valpine, P.; Bowles, T.M. Biophysical and policy factors predict simplified crop rotations in the US Midwest. Environ. Res. Lett. 2021, 16, 054045. [Google Scholar] [CrossRef]
- Khan, M.N.; Mohammad, F. Eutrophication: Challenges and Solutions. In Eutrophication: Causes, Consequences and Control; Ansari, A.A., Gill, S.S., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 2, pp. 1–15. [Google Scholar]
- Davidson, E.A.; Kanter, D. Inventories and scenarios of nitrous oxide emissions. Environ. Res. Lett. 2014, 9, 105012. [Google Scholar] [CrossRef]
- Baumhardt, R.L.; Stewart, B.A.; Sainju, U.M. North American Soil Degradation: Processes, Practices, and Mitigating Strategies. Sustainability 2015, 7, 2936–2960. [Google Scholar] [CrossRef] [Green Version]
- Karlen, D.L.; Veum, K.S.; Sudduth, K.A.; Obrycki, J.F.; Nunes, M.R. Soil health assessment: Past accomplishments, current activities, and future opportunities. Soil Till. Res. 2019, 195, 104365. [Google Scholar] [CrossRef]
- Jagadamma, S.; Lal, R.; Hoeft, R.G.; Nafziger, E.D.; Adee, E.A. Nitrogen fertilization and cropping systems effects on soil organic carbon and total nitrogen pools under chisel-plow tillage in Illinois. Soil Till. Res. 2007, 95, 348–356. [Google Scholar] [CrossRef]
- Liebig, M.A.; Varvel, G.E.; Doran, J.W.; Wienhold, B.J. Crop sequence and nitrogen fertilization effects on soil properties in the Western Corn Belt. Soil Sci. Soc. Am. J. 2002, 66, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Russell, A.E.; Laird, D.A.; Mallarino, A.P. Nitrogen fertilization and cropping system impacts on soil quality in Midwestern Mollisols. Soil Sci. Soc. Am. J. 2006, 70, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Stone, D.L.R.; Whitney, D.A.; Janssen, K.A.; Long, J.H. Soil properties after twenty years of fertilization with different nitrogen sources. Soil Sci. Soc. Am. J. 1991, 55, 1097–1100. [Google Scholar] [CrossRef]
- Behnke, G.D.; Zuber, S.M.; Pittelkow, C.M.; Nafziger, E.D.; Villamil, M.B. Long-term crop rotation and tillage effects on soil greenhouse gas emissions and crop production in Illinois, USA. Agric. Ecosyst. Environ. 2018, 261, 62–70. [Google Scholar] [CrossRef]
- Congreves, K.A.; Hayes, A.; Verhallen, E.A.; Van Eerd, L.L. Long-term impact of tillage and crop rotation on soil health at four temperate agroecosystems. Soil Till. Res. 2015, 152, 17–28. [Google Scholar] [CrossRef]
- Hoss, M.; Behnke, G.D.; Davis, A.S.; Nafziger, E.D.; Villamil, M.B. Short corn rotations do not improve soil quality, compared with corn monocultures. Agron. J. 2018, 110, 1274–1288. [Google Scholar] [CrossRef] [Green Version]
- Zuber, S.M.; Behnke, G.D.; Nafziger, E.D.; Villamil, M.B. Crop rotation and tillage effects on soil physical and chemical properties in Illinois. Agron. J. 2015, 107, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Barak, P.; Jobe, B.O.; Krueger, A.R.; Peterson, L.A.; Laird, D.A. Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant Soil 1997, 197, 61–69. [Google Scholar] [CrossRef]
- Obour, A.K.; Mikha, M.M.; Holman, J.D.; Stahlman, P.W. Changes in soil surface chemistry after fifty years of tillage and nitrogen fertilization. Geoderma 2017, 308, 46–53. [Google Scholar] [CrossRef]
- Jagadamma, S.; Lal, R.; Hoeft, R.G.; Nafziger, E.D.; Adee, E.A. Nitrogen fertilization and cropping system impacts on soil properties and their relationship to crop yield in the central Corn Belt, USA. Soil Till. Res. 2008, 98, 120–129. [Google Scholar] [CrossRef]
- Russell, A.E.; Cambardella, C.A.; Laird, D.A.; Jaynes, D.B.; Meek, D.W. Nitrogen fertilizer effects on soil carbon balances in Midwestern U.S. agricultural systems. Ecol. Appl. 2009, 19, 1102–1113. [Google Scholar] [CrossRef] [Green Version]
- Bowles, T.M.; Atallah, S.S.; Campbell, E.E.; Gaudin, A.C.; Wieder, W.R.; Grandy, A.S. Addressing agricultural nitrogen losses in a changing climate. Nat. Sustain. 2018, 1, 399–408. [Google Scholar] [CrossRef]
- Ajwa, H.A.; Tabatabai, M.A. Decomposition of different organic materials in soils. Biol. Fertil. Soils 1994, 18, 175–182. [Google Scholar] [CrossRef]
- Martens, D.A. Management and crop residue influence soil aggregate stability. J. Environ. Qual. 2000, 29, 723–727. [Google Scholar] [CrossRef] [Green Version]
- Illinois State Water Survey. Illinois Climate Normals; Illinois State Water Survey: Champaign, IL, USA, 2010. [Google Scholar]
- Soil Survey Staff; NRCS; USDA. Web Soil Survey. Available online: http://websoilsurvey.sc.egov.usda.gov/ (accessed on 10 November 2020).
- Nafziger, E. Illinois Agronomy Handbook; University of Illinois Extension: Urbana, IL, USA, 2009; Volume 224. [Google Scholar]
- Blake, G.R.; Hartge, K.H. Bulk Density. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; Volume 5, pp. 363–375. [Google Scholar]
- Schulte, E.; Hopkins, B. Estimation of soil organic matter by weight loss-on-ignition. Soil Org. Matter Anal. Interpret. 1996, 46, 21–31. [Google Scholar]
- Konen, M.E.; Jacobs, P.M.; Burras, C.L.; Talaga, B.J.; Mason, J.A. Equations for predicting soil organic carbon using loss-on-ignition for north central US soils. Soil Sci. Soc. Am. J. 2002, 66, 1878–1881. [Google Scholar] [CrossRef]
- McLean, E. Soil pH and lime requirement. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; American Society of Agronomy: Madison, WI, USA, 1983; Volume 9, pp. 199–224. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Ross, D.S.; Ketterings, Q. Recommended methods for determining soil cation exchange capacity. Recomm. Soil Test. Proced. Northeast. USA 1995, 493, 62. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Oliver, S. SAS for Mixed Models; SAS Institute: Cary, NC, USA, 2006. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; 4.1.0; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Tang, C.; Rengel, Z. Role of plant cation/anion uptake ratio in soil acidification. In Handbook of Soil Acidity, 1st ed.; Rengel, Z., Ed.; CEC Press: New York, NY, USA, 2003. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. Nitrogen transformations in modern agriculture and the role of biological nitrification inhibition. Nat. Plants 2017, 3, 17074. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C. Nitrate leaching in temperate agroecosystems: Sources, factors and mitigating strategies. Nutr. Cycl. Agroecosystems 2002, 64, 237–256. [Google Scholar] [CrossRef]
- Lin, B.; Sakoda, A.; Shibasaki, R.; Suzuki, M. A modelling approach to global nitrate leaching caused by anthropogenic fertilisation. Water Res. 2001, 35, 1961–1968. [Google Scholar] [CrossRef]
- Behnke, G.D.; Pittelkow, C.; Nafziger, E.; Villamil, M.B. Exploring the relationships between greenhouse gas emissions, yields, and soil properties in cropping systems. Agriculture 2018, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Riggins, C.W.; Rodríguez-Zas, S.; Zabaloy, M.C.; Villamil, M.B. Long-term N fertilization imbalances potential N acquisition and transformations by soil microbes. Sci. Total Environ. 2019, 691, 562–571. [Google Scholar] [CrossRef]
- Rengel, Z. Soil pH, Soil Health and Climate Change. In Soil Health and Climate Change; Singh, B.P., Cowie, A.L., Chan, K.Y., Eds.; Springer: Berlin, Germany; Heidelberg, Germany, 2011; pp. 69–85. [Google Scholar]
- Villamil, M.B.; Kim, N.; Riggins, C.W.; Zabaloy, M.C.; Allegrini, M.; Rodríguez-Zas, S.L. Microbial Signatures in Fertile Soils Under Long-Term N Management. Front. Soil Sci. 2021, 1, 24. [Google Scholar] [CrossRef]
- Lal, R. World crop residues production and implications of its use as a biofuel. Environ. Int. 2005, 31, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Necpálová, M.; Anex, R.P.; Kravchenko, A.N.; Abendroth, L.J.; Del Grosso, S.J.; Dick, W.A.; Helmers, M.J.; Herzmann, D.; Lauer, J.G.; Nafziger, E.D.; et al. What does it take to detect a change in soil carbon stock? A regional comparison of minimum detectable difference and experiment duration in the north central United States. J. Soil. Water. Conserv. 2014, 69, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Lal, R. Soil carbon dynamics in cropland and rangeland. Environ. Pollut. 2002, 116, 353–362. [Google Scholar] [CrossRef]
- Brown, K.H.; Bach, E.M.; Drijber, R.A.; Hofmockel, K.S.; Jeske, E.S.; Sawyer, J.E.; Castellano, M.J. A long-term nitrogen fertilizer gradient has little effect on soil organic matter in a high-intensity maize production system. Glob. Change Biol. 2014, 20, 1339–1350. [Google Scholar] [CrossRef]
- Chen, R.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Lin, X.; Blagodatskaya, E.; Kuzyakov, Y. Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories. Glob. Change Biol. 2014, 20, 2356–2367. [Google Scholar] [CrossRef]
- Poffenbarger, H.J.; Barker, D.W.; Helmers, M.J.; Miguez, F.E.; Olk, D.C.; Sawyer, J.E.; Six, J.; Castellano, M.J. Maximum soil organic carbon storage in Midwest US cropping systems when crops are optimally nitrogen-fertilized. PLoS ONE 2017, 12, e0172293. [Google Scholar] [CrossRef]
- Wilpiszeski, R.L.; Aufrecht, J.A.; Retterer, S.T.; Sullivan, M.B.; Graham, D.E.; Pierce, E.M.; Zablocki, O.D.; Palumbo, A.V.; Elias, D.A. Soil aggregate microbial communities: Towards understanding microbiome interactions at biologically relevant scales. Appl. Environ. Microbiol. 2019, 85, e00324-19. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, P.; Delgado-Baquerizo, M.; Jeffries, T.C.; Trivedi, C.; Anderson, I.C.; Lai, K.; McNee, M.; Flower, K.; Pal Singh, B.; Minkey, D.; et al. Soil aggregation and associated microbial communities modify the impact of agricultural management on carbon content. Environ. Microbiol. 2017, 19, 3070–3086. [Google Scholar] [CrossRef]
- Lynch, J.M.; Bragg, E. Microorganisms and Soil Aggregate Stability; Springer: New York, NY, USA, 1985; Volume 2, pp. 133–171. [Google Scholar]
- Trivedi, P.; Rochester, I.J.; Trivedi, C.; Van Nostrand, J.D.; Zhou, J.; Karunaratne, S.; Anderson, I.C.; Singh, B.K. Soil aggregate size mediates the impacts of cropping regimes on soil carbon and microbial communities. Soil Biol. Biochem. 2015, 91, 169–181. [Google Scholar] [CrossRef]
- Halvorson, A.D.; Reule, C.A.; Follett, R.F. Nitrogen fertilization effects on soil carbon and nitrogen in a dryland cropping system. Soil Sci. Soc. Am. J. 1999, 63, 912–917. [Google Scholar] [CrossRef]
- Behnke, G.D.; Zabaloy, M.C.; Riggins, C.W.; Rodríguez-Zas, S.; Huang, L.; Villamil, M.B. Acidification in corn monocultures favor fungi, ammonia oxidizing bacteria, and nirK-denitrifier groups. Sci. Total Environ. 2020, 720, 137514. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.W. The effect of parent material and soil development on nutrient cycling in temperate ecosystems. Biogeochemistry 1988, 5, 71–97. [Google Scholar] [CrossRef]
- Simonsson, M.; Andersson, S.; Andrist-Rangel, Y.; Hillier, S.; Mattsson, L.; Öborn, I. Potassium release and fixation as a function of fertilizer application rate and soil parent material. Geoderma 2007, 140, 188–198. [Google Scholar] [CrossRef]
- Yu, Q.; Wu, W.; Verburg, P.H.; van Vliet, J.; Yang, P.; Zhou, Q.; Tang, H. A survey-based exploration of land-system dynamics in an agricultural region of Northeast China. Agric. Syst. 2013, 121, 106–116. [Google Scholar] [CrossRef]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zhang, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | CEC | pH | SOC | WAS | BD | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | ||
| Rot | |||||||||||
| CCC | 18.51 | 0.44 | 6.31 | 0.12 | 20.08 | 0.63 | 74.99 | 1.38 | 1.34 | 0.03 | |
| Cs | 18.62 | 6.79 | 19.14 | 72.22 | 1.38 | ||||||
| Sc | 19.68 | 6.71 | 18.17 | 69.95 | 1.40 | ||||||
| Nrate | |||||||||||
| 0 | 19.06 | 0.38 | 6.73 | 0.08 | 18.90 | 0.58 | 71.42 | 1.38 | 1.41 | 0.02 | |
| 202 | 18.85 | 6.59 | 19.08 | 73.26 | 1.38 | ||||||
| 269 | 18.90 | 6.49 | 19.41 | 72.49 | 1.34 | ||||||
| D | |||||||||||
| 0–15 | 19.39 | 0.45 | 6.40 | 0.09 | 25.56 | 0.48 | 64.80 | 1.21 | 1.37 | 0.02 | |
| 15–30 | 19.33 | 0.40 | 6.55 | 0.08 | 23.28 | 0.49 | 76.96 | 1.30 | 1.45 | 0.02 | |
| 30–60 | 18.01 | 0.39 | 6.69 | 0.07 | 17.37 | 0.44 | 79.39 | 1.32 | 1.30 | 0.02 | |
| 60–90 | 19.01 | 0.42 | 6.78 | 0.07 | 10.31 | 0.22 | 68.40 | 1.48 | 1.37 | 0.02 | |
| Source of Variation | df | CEC | pH | SOC | WAS | BD | |||||
| Rot | 2 | 0.108 | 0.0616 | 0.1250 | 0.0167 | 0.3104 | |||||
| Nrate | 2 | 0.709 | 0.0057 | 0.8108 | 0.5158 | 0.0240 | |||||
| Rot × Nrate | 4 | 0.002 | 0.0124 | 0.9789 | 0.2068 | 0.9287 | |||||
| D | 3 | 0.001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||||
| Rot × D | 6 | 0.003 | <0.0001 | 0.2840 | 0.8282 | 0.3100 | |||||
| Nrate × D | 6 | 0.414 | 0.0033 | 0.3125 | 0.3743 | 0.5247 | |||||
| Rot × Nrate × D | 12 | 0.914 | 0.0351 | 0.3482 | 0.5991 | 0.9474 |
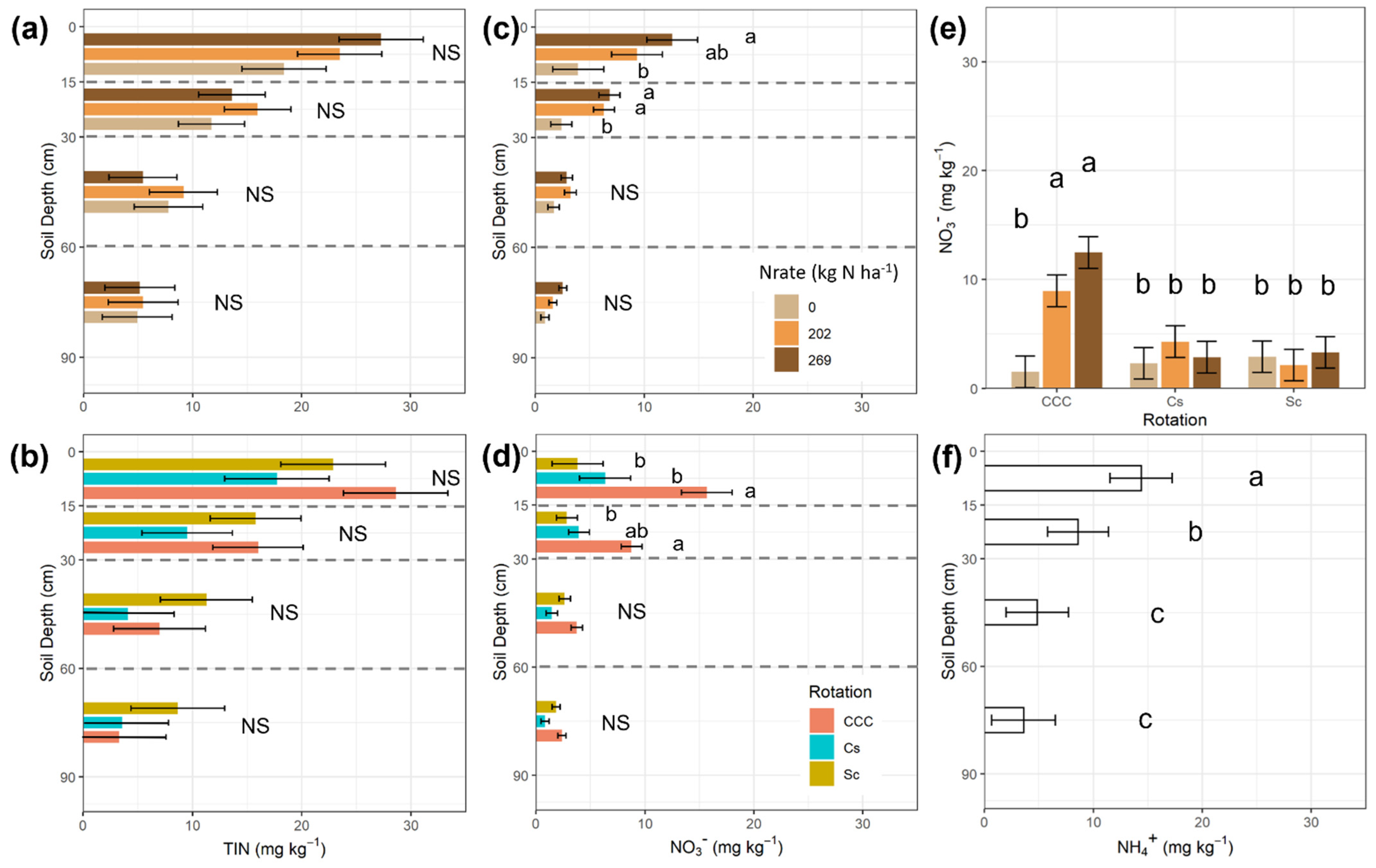
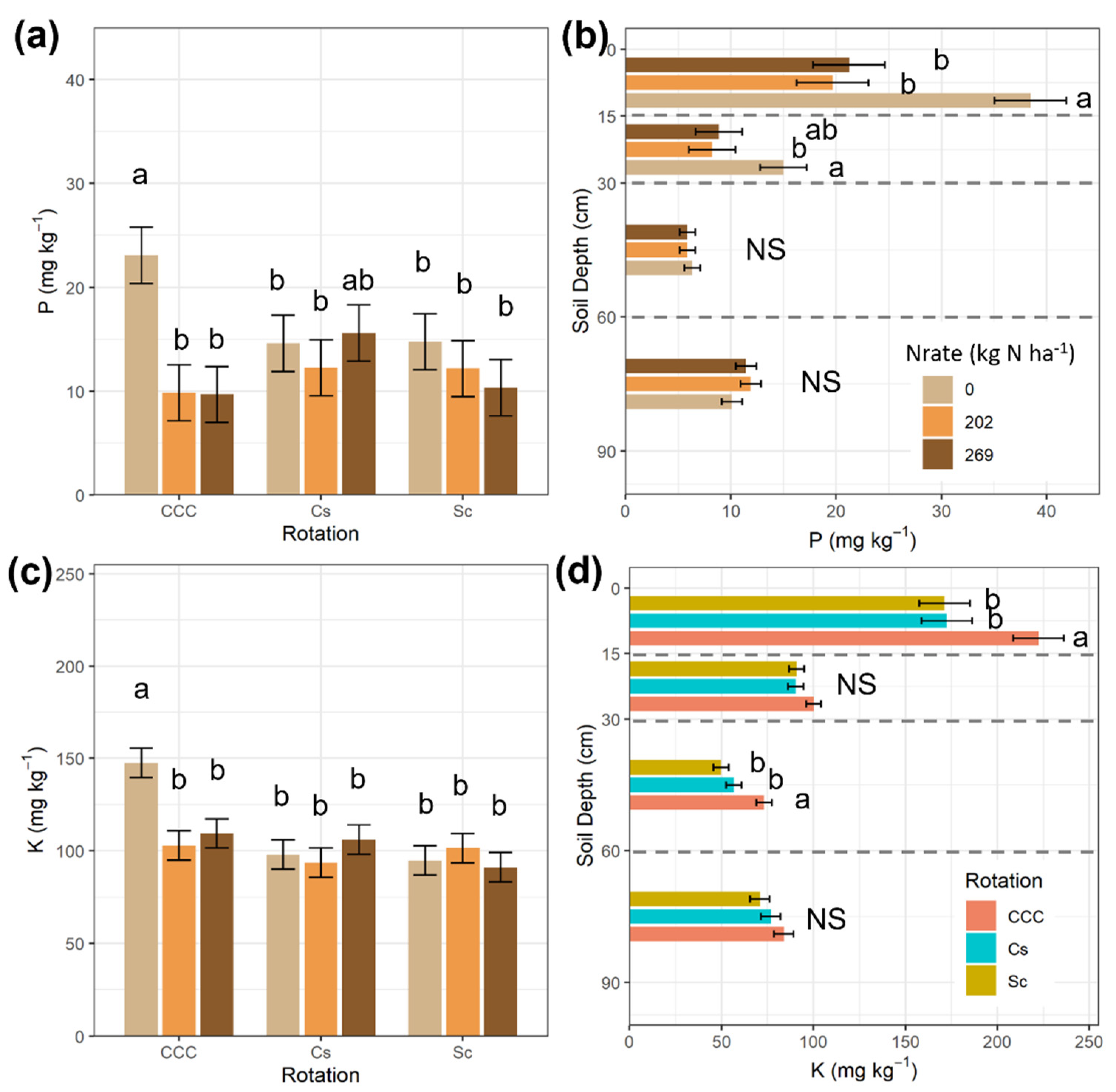
| Treatments | NO3− | NH4+ | TIN | P | K | Yield | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |||
| Rot | ||||||||||||||
| CCC | 7.65 | 0.84 | 6.15 | 4.74 | 13.72 | 4.16 | 14.19 | 1.56 | 119.86 | 4.62 | 8.44 | 0.40 | ||
| Cs | 3.15 | 5.54 | 8.73 | 14.14 | 99.11 | 11.05 | ||||||||
| Sc | 2.78 | 11.86 | 14.65 | 12.42 | 95.75 | |||||||||
| Nrate | ||||||||||||||
| 0 | 2.24 | 0.84 | 8.42 | 3.05 | 10.70 | 3.06 | 17.47 | 1.56 | 113.39 | 4.57 | 6.89 | 0.41 | ||
| 202 | 5.12 | 8.39 | 13.52 | 11.42 | 99.22 | 11.10 | ||||||||
| 269 | 6.21 | 6.74 | 12.88 | 11.86 | 102.11 | 11.25 | ||||||||
| D | ||||||||||||||
| 0–15 | 8.62 | 1.35 | 14.38 | 2.86 | 23.06 | 2.99 | 26.44 | 1.96 | 188.70 | 7.92 | ||||
| 15–30 | 5.19 | 0.56 | 8.58 | 2.80 | 13.77 | 2.65 | 10.70 | 1.28 | 93.81 | 2.39 | ||||
| 30–60 | 2.61 | 0.30 | 4.85 | 2.87 | 7.46 | 2.68 | 6.04 | 0.44 | 59.93 | 2.41 | ||||
| 60–90 | 1.69 | 0.21 | 3.59 | 2.92 | 5.17 | 2.72 | 11.15 | 0.57 | 77.19 | 3.05 | ||||
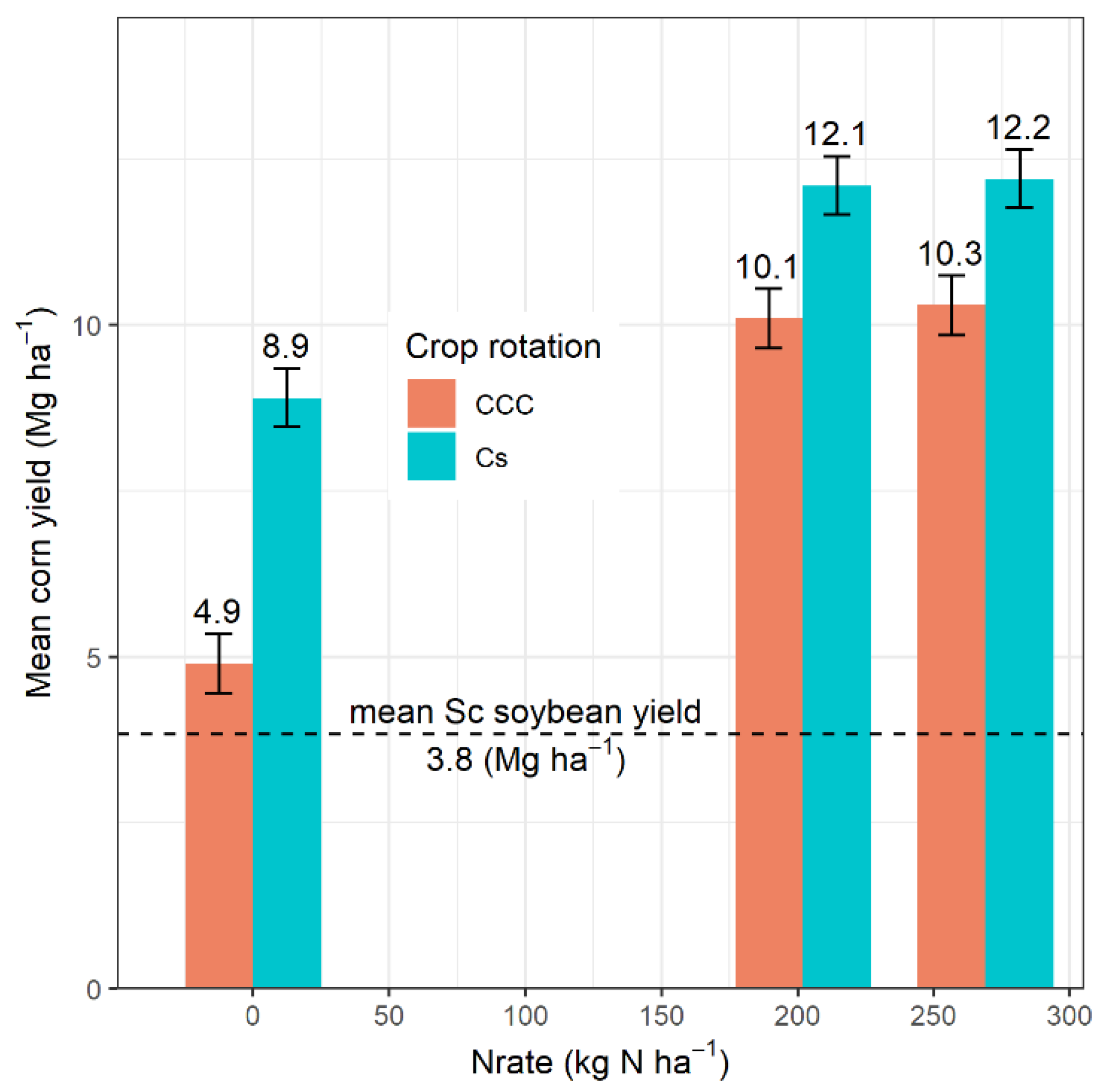
| Source of Variation | df | NO3− | NH4+ | TIN | P | K | df | Yield | ||||||
| Rot | 2 | 0.0005 | 0.6122 | 0.5638 | 0.6632 | 0.0047 | 1 | <0.0001 | ||||||
| Nrate | 2 | 0.0075 | 0.6428 | 0.5520 | 0.0207 | 0.0908 | 2 | <0.0001 | ||||||
| Rot × Nrate | 4 | 0.0055 | 0.4296 | 0.2296 | 0.0973 | 0.0171 | 2 | <0.0001 | ||||||
| D | 3 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||||||||
| Rot × D | 6 | 0.0097 | 0.8515 | 0.0972 | 0.6705 | 0.0870 | ||||||||
| Nrate × D | 6 | 0.0643 | 0.2630 | 0.0547 | 0.0029 | 0.3401 | ||||||||
| Rot × Nrate × D | 12 | 0.1996 | 0.7622 | 0.6334 | 0.1265 | 0.2185 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.; Behnke, G.D.; Villamil, M.B. Characterization of Mollisols after Long-Term N Fertilization at Successive Rates in Continuous and Rotated Corn Systems. Agronomy 2022, 12, 625. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030625
Kim N, Behnke GD, Villamil MB. Characterization of Mollisols after Long-Term N Fertilization at Successive Rates in Continuous and Rotated Corn Systems. Agronomy. 2022; 12(3):625. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030625
Chicago/Turabian StyleKim, Nakian, Gevan D. Behnke, and María B. Villamil. 2022. "Characterization of Mollisols after Long-Term N Fertilization at Successive Rates in Continuous and Rotated Corn Systems" Agronomy 12, no. 3: 625. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030625