Manipulating the Difference between the Day and Night Temperatures Can Enhance the Quality of Astragalus membranaceus and Codonopsis lanceolata Plug Seedlings



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Contents of Soluble Sugar and Starch
2.3. Contents of Total Phenols and Flavonoids
2.4. Hydrogen Peroxide Content
2.5. Assessments of the Chlorophyll Content and Stomatal Conductance
2.6. Gene Expression Analysis
2.7. Data Collection and Analysis
3. Results
3.1. Growth, Development, and Morphology
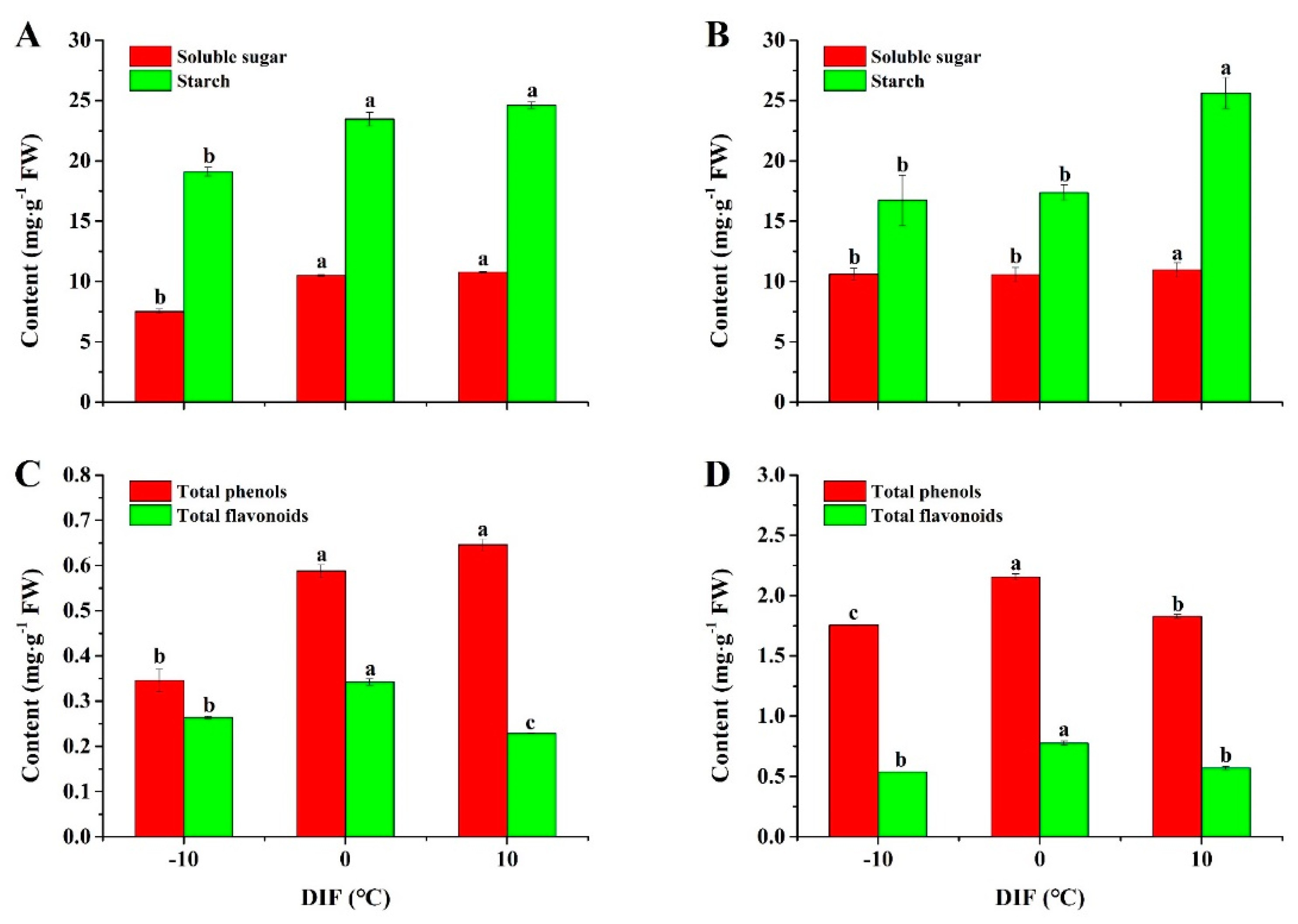
3.2. Contents of Soluble Sugar and Starch
3.3. Contents of Total Phenols and Flavonoids
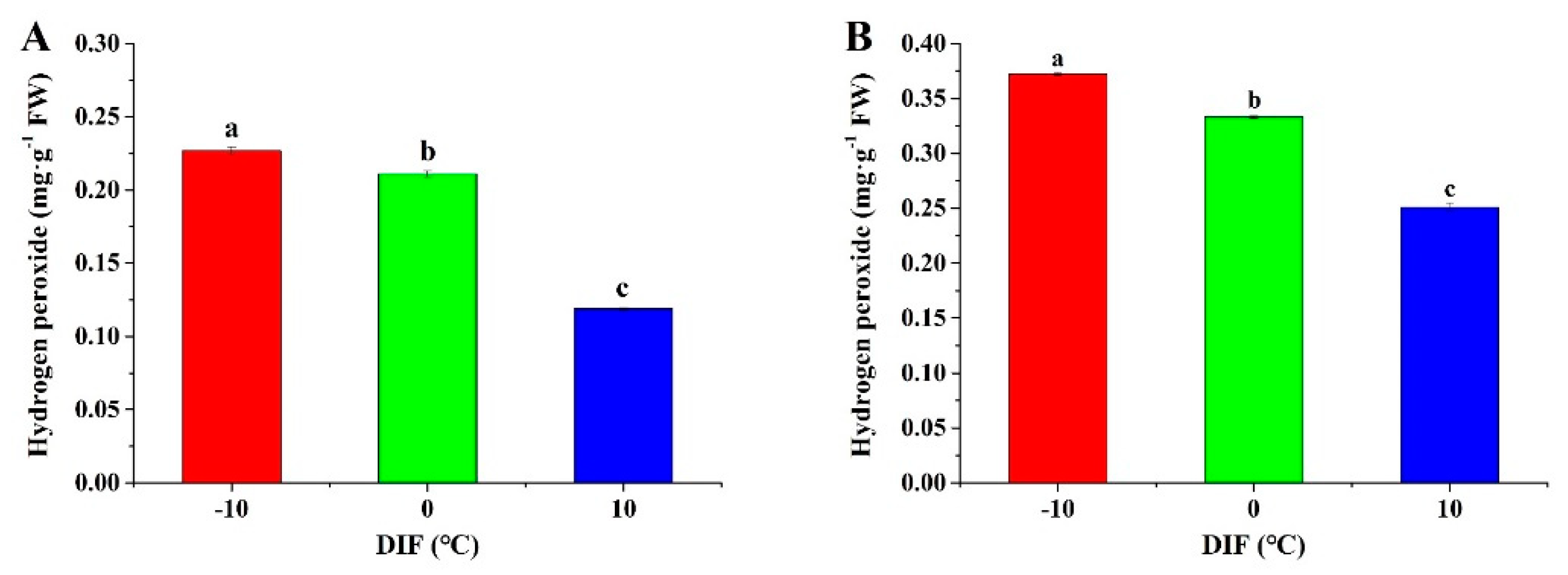
3.4. Hydrogen Peroxide Content
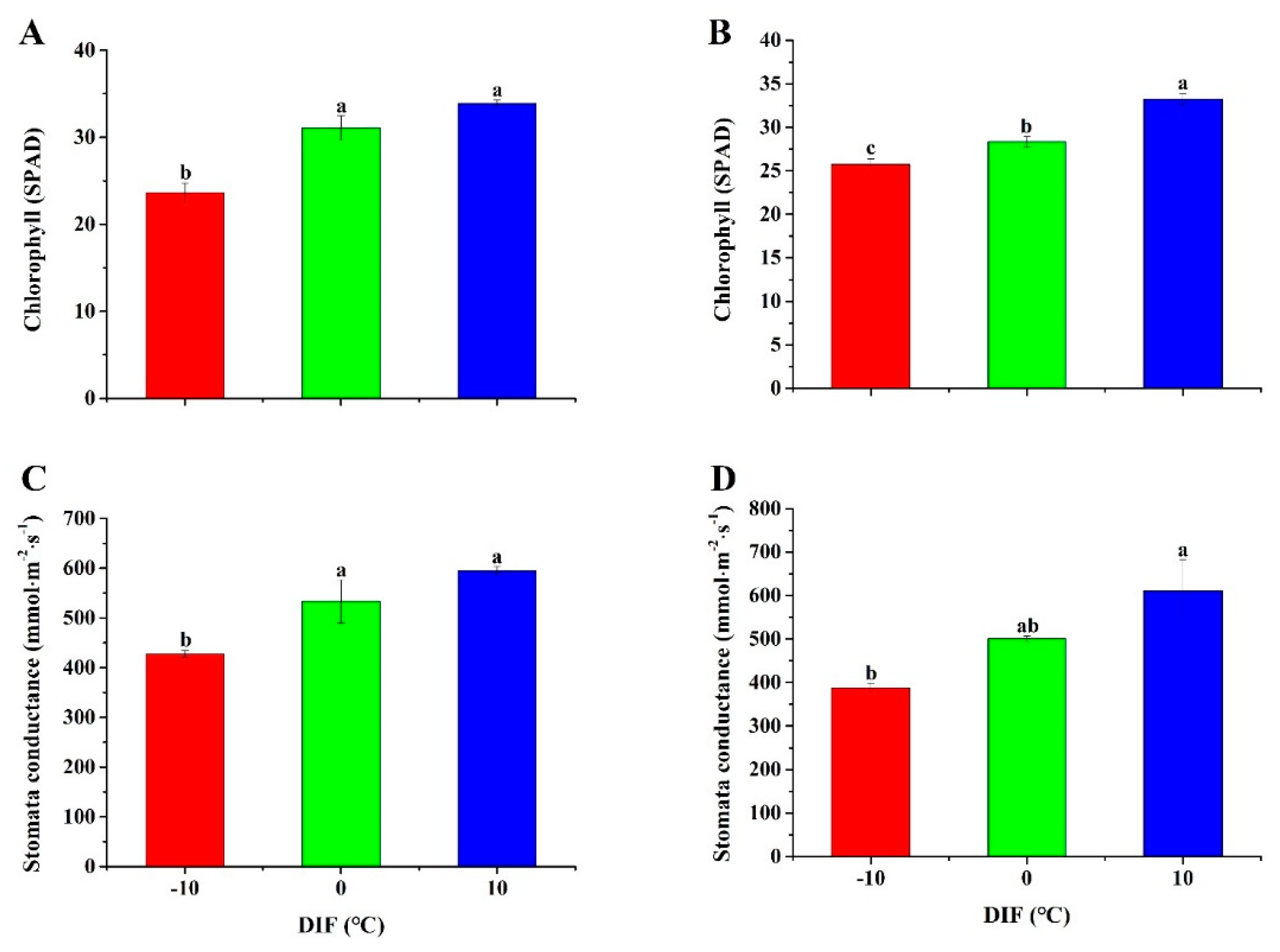
3.5. Chlorophyll Content and Stomatal Conductance
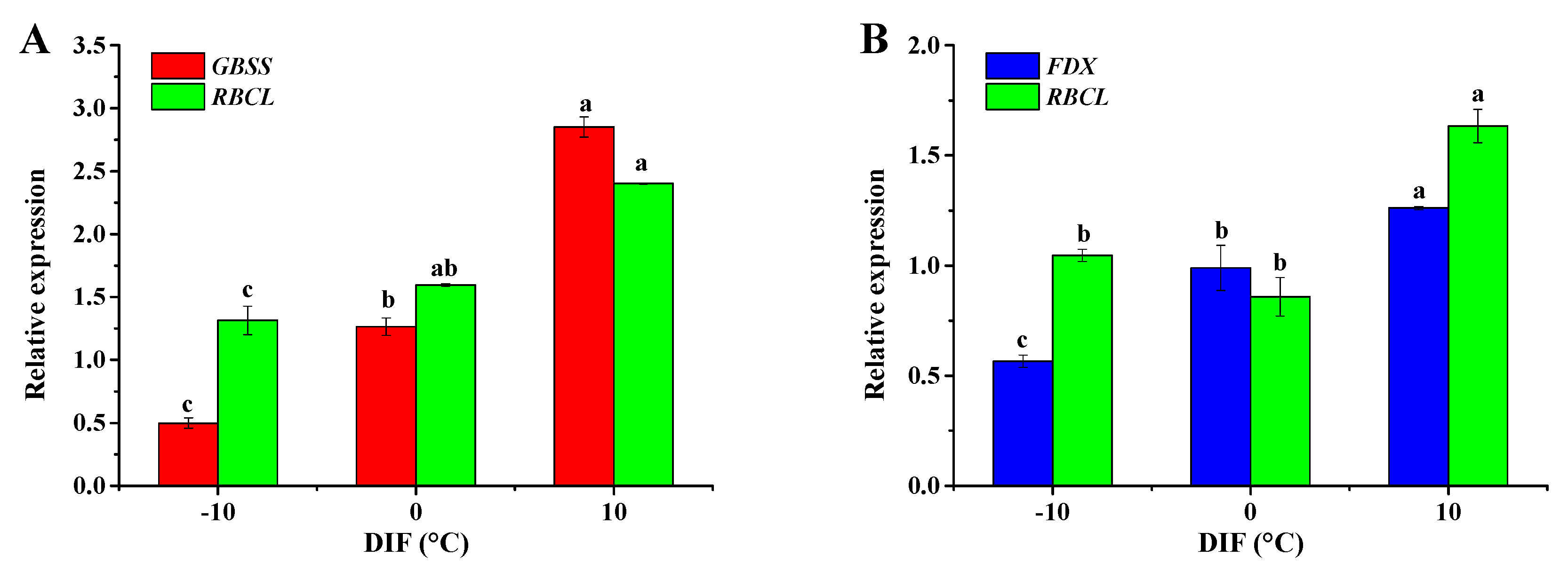
3.6. Gene Expression Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yang, J.; Jia, M.M.; Zhang, X.J.; Wang, P.L. Calycosin attenuates MPTP-induced Parkinson’s disease by suppressing the activation of TLR/NF-kappa b and mapk pathways. Phytother. Res. 2019, 33, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Han, A.Y.; Lee, Y.S.; Kwon, S.; Lee, H.S.; Lee, K.W.; Seol, G.H. Codonopsis lanceolata extract prevents hypertension in rats. Phytomedicine 2018, 39, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Boo, H.O.; Kim, H.H.; Barasch, D.; Nemirovski, A.; Lee, M.S.; Gorinstein, S.; Ku, Y.G. Codonopsis lanceolata and Nelumbo nucifera Gaertn. root extracts for functional food: Metabolic profiling by MS, FTIR and fluorescence and evaluation of cytotoxicity and Anti-Obesity properties on 3T3-L1 cell line. Eur. Food Res. Technol. 2017, 243, 689–700. [Google Scholar] [CrossRef]
- Moon, K.G.; Um, I.S.; Jeon, S.H.; Cho, Y.S.; Kim, Y.G.; Rho, I.R. Effect of organic fertilizer application on growth characteristics and saponin content in Codonopsis lanceolata. Hortic. Environ. Biotechnol. 2018, 59, 125–130. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Seong, E.S.; Yu, C.Y.; Kim, S.H.; Chung, I.M. Evaluation of phenolic compounds and antimicrobial activities in transgenic Codonopsis lanceolata plants via overexpression of the gamma-tocopherol methyltransferase (gamma-tmt) gene. S. Afr. J. Bot. 2017, 109, 25–33. [Google Scholar] [CrossRef]
- Li, Y.; Guo, S.; Zhu, Y.; Yan, H.; Qian, D.W.; Wang, H.Q.; Yu, J.Q.; Duan, J.A. Flowers of Astragalus membranaceus var. mongholicus as a novel high potential by-Product: Phytochemical characterization and antioxidant activity. Molecules 2019, 24, 434. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Zhang, Z.; Xu, X.; Wang, K.; Cheng, H.; Cao, B. Different mechanisms to obtain higher fruit growth rate in two Cold-Tolerant cucumber (Cucumis sativus L.) lines under low night temperature. Sci. Hortic. 2009, 119, 357–361. [Google Scholar] [CrossRef]
- Shi, W.; Yin, X.; Struik, P.C.; Xie, F.; Schmidt, R.C.; Jagadish, K.S.V. Grain yield and quality responses of tropical hybrid rice to high Night-Time temperature. Field Crops Res. 2016, 190, 18–25. [Google Scholar] [CrossRef]
- Shi, W.J.; Muthurajan, R.; Rahman, H.; Selvam, J.; Peng, S.B.; Zou, Y.B.; Jagadish, K.S.V. Source-Sink dynamics and proteomic reprogramming under elevated night temperature and their impact on rice yield and grain quality. New Phytol. 2013, 197, 825–837. [Google Scholar] [CrossRef]
- Robert, B. Controlling height with temperature. HortTechnology 1998, 8, 535–539. [Google Scholar]
- Bakken, A.K.; Flønes, M. Morphology and field performance of brassica transplants propagated under different day and night temperature regimes. Sci. Hortic. 1995, 61, 167–176. [Google Scholar] [CrossRef]
- Thingnaes, E.; Torre, S.; Ernstsen, A.; Moe, R. Day and night temperature responses in arabidopsis: Effects on gibberellin and auxin content, cell size, morphology and flowering time. Ann. Bot. 2003, 92, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Heins, R.D. Photoperiod and the difference between day and night temperature influence stem elongation kinetics in Verbena bonariensis. J. Am. Soc. Hortic. Sci. 2000, 125, 576–580. [Google Scholar] [CrossRef]
- Moe, R.; Heins, R.D.; Erwin, J. Stem elongation and flowering of the long-day plant Campanula-Isophylla moretti in response to day and night temperature alternations and light quality. Sci. Hortic. 1991, 48, 141–151. [Google Scholar] [CrossRef]
- Mortensen, L.M. Effects of Day-Night Temperature-Variations on growth, morphogenesis and flowering of Kalanchoe blossfeldiana v Poelln at different CO2 concentrations, daylengths and photon flux densities. Sci. Hortic. 1994, 59, 233–241. [Google Scholar] [CrossRef]
- Lund, J.B.; Andreassen, A.; Ottosen, C.O.; Aaslyng, J.M. Effect of a dynamic climate on energy consumption and production of Hibiscus rosa-sinensis L. in greenhouses. HortScience 2006, 41, 384–388. [Google Scholar] [CrossRef]
- Lyngved, R.; Snipen, L.G.; Iversen, T.H.; Hvoslef-Eide, A.K. Influence of potential growth factors on the production of proembryogenic masses of Cyclamen persicum Mill. in bioreactors. Sci. Hortic. 2008, 118, 53–59. [Google Scholar] [CrossRef]
- Grimstad, S.O.; Frimanslund, E. Effect of different day and night temperature regimes on greenhouse cucumber young plant production, flower bud formation and early yield. Sci. Hortic. 1993, 53, 191–204. [Google Scholar] [CrossRef]
- Yuan, X.K.; Yang, Z.Q. The effect of endogenous hormones on plant morphology and fruit quality of tomato under difference between day and night temperature. Hortic. Sci. 2018, 45, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Kojoh, K.; Matsuzawa, H.; Wakagi, T. Zinc and an N-Terminal extra stretch of the ferredoxin from a thermoacidophilic archaeon stabilize the molecule at high temperature. Eur. J. Biochem. 1999, 264, 85–91. [Google Scholar] [CrossRef]
- Schuermann, P.; Buchanan, B.B. The ferredoxin/thioredoxin system of oxygenic photosynthesis. Antioxid. Redox Signal. 2008, 10, 1235–1273. [Google Scholar] [CrossRef] [PubMed]
- Spreitzer, R.J.; Salvucci, M.E. Rubisco: Structure, regulatory interactions, and possibilities for a better enzyme. Annu. Rev. Plant Biol. 2002, 53, 449–475. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M.; Schulze, D. Does rubisco control the rate of photosynthesis and Plant-Growth—An exercise in molecular ecophysiology. Plant Cell Environ. 1994, 17, 465–487. [Google Scholar] [CrossRef]
- Merida, A.; Rodriguez-Galan, J.M.; Vincent, C.; Romero, J.M. Expression of the granule-bound starch synthase I (Waxy) gene from snapdragon is developmentally and circadian clock regulated. Plant Physiol. 1999, 120, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Visser, R.G.F.; Stolte, A.; Jacobsen, E. Expression of a chimeric granule-bound starch Synthase-Gus gene in transgenic potato plants. Plant Mol. Biol. 1991, 17, 691–699. [Google Scholar] [CrossRef]
- Umemoto, T.; Nakamura, Y.; Ishikura, N. Activity of starch synthase and the amylose content in rice endosperm. Phytochemistry 1995, 40, 1613–1616. [Google Scholar] [CrossRef]
- Zhao, H.; Dai, T.; Jiang, D.; Cao, W. Effects of high temperature on key enzymes involved in starch and protein formation in grains of two wheat cultivars. J. Agron. Crop Sci. 2008, 194, 47–54. [Google Scholar] [CrossRef]
- Hurkman, W.J.; McCue, K.F.; Altenbach, S.B.; Korn, A.; Tanaka, C.K.; Kotharia, K.M.; Johnson, E.L.; Bechtel, D.B.; Wilson, J.D.; Anderson, O.D.; et al. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci. 2003, 164, 873–881. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Dzhibladze, T.G.; Egorova, E.A. Elevated temperatures inhibit ferredoxin-dependent cyclic electron flow around photosystem I. Russ. J. Plant Physiol. 2005, 52, 578–583. [Google Scholar] [CrossRef]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Liu, Y.; Ren, X.; Jeong, H.; Wei, H.; Jeong, B. Growth and physiological responses of Adenophora triphylla (Thunb.) A.DC. Plug seedlings to day and night temperature regimes. Agronomy 2018, 8, 173. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Park, Y.G.; Jeong, B.R. In vitro propagation, phytochemical analysis, and evaluation of free radical scavenging property of Scrophularia kakudensis Franch tissue extracts. BioMed Res. Int. 2015, 2015, 480564. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Liu, Y.; Jeong, H.; Jeong, B. Supplementary light source affects the growth and development of Codonopsis lanceolata seedlings. Int. J. Mol. Sci. 2018, 19, 3074. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Hwang, S.J. The growth and development of ‘mini chal’ tomato plug seedlings grown under various wavelengths using light emitting diodes. Agronomy 2019, 9, 157. [Google Scholar] [CrossRef]
- Oh, H.J.; Park, J.E.; Park, Y.G.; Jeong, B.R. Growth and quality of plug seedlings of three indigenous medicinal plants as affected by ionic strength of the nutrient solution. Hortic. Environ. Biotechnol. 2014, 55, 63–69. [Google Scholar] [CrossRef]
- Santos-Moura, S.D.; Alves, E.U.; Ursulino, M.M.; Bruno, R.D.A.; dos Anjos Neto, A.P. Effect of shading on Dimorphandra gardneriana Tul. seedling production. Biosci. J. 2018, 34, 1147–1157. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Li, Y.S.; Li, P.; Zhang, F.M.; Thomas, B.W. Effect of difference between day and night temperature on tomato (Lycopersicon esculentum Mill.) root activity and low molecular weight organic acid secretion. Soil Sci. Plant Nutr. 2016, 62, 423–431. [Google Scholar] [CrossRef]
- Kim, H.C.; Ku, Y.G.; Lee, Y.B.; Lee, J.H.; Choi, J.H.; Bae, J.H. Early growth of sweet pepper by difference between day and night temperature after planting. Korean J. Hortic. Sci. 2013, 31, 552–557. [Google Scholar]
- Bachman, G.R.; McMahon, M.J. Day and night temperature differential (DIF) or the absence of Far-Red light alters cell elongation in ‘celebrity white’ petunia. J. Am. Soc. Hortic. Sci. 2006, 131, 309–312. [Google Scholar] [CrossRef]
- William, B.M.; Hammer, P.A.; Terri, I.K. Reversed greenhouse temperatures alter carbohydrate status in Lilium longiflorum Thunb. ‘nellie white’. J. Am. Soc. Hortic. Sci. 1993, 118, 736–740. [Google Scholar]
- Tombesi, S.; Cincera, I.; Frioni, T.; Ughini, V.; Gatti, M.; Palliotti, A.; Poni, S. Relationship among night temperature, carbohydrate translocation and inhibition of grapevine leaf photosynthesis. Environ. Exp. Bot. 2019, 157, 293–298. [Google Scholar] [CrossRef]
- Mohammed, A.R.; Cothren, J.T.; Chen, M.H.; Tarpley, L. 1-Methylcyclopropene (1-MCP)-Induced alteration in leaf photosynthetic rate, chlorophyll fluorescence, respiration and membrane damage in rice (Oryza sativa L.) under high night temperature. J. Agron. Crop Sci. 2015, 201, 105–116. [Google Scholar] [CrossRef]
- Pyl, E.T.; Piques, M.; Ivakov, A.; Schulze, W.; Ishihara, H.; Stitt, M.; Sulpice, R. Metabolism and growth in arabidopsis depend on the daytime temperature but are Temperature-Compensated against cool nights. Plant Cell 2012, 24, 2443–2469. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, S.S.; Ibrahim, R.; Damalas, C.A.; Noorhosseini, S.A. Effects of gamma stress and carbon dioxide on eight bioactive flavonoids and photosynthetic efficiency in Centella asiatica. J. Plant Growth Regul. 2017, 36, 957–969. [Google Scholar] [CrossRef]
- Ali, M.; Abbasi, B.H.; Ahmad, N.; Ali, S.S.; Ali, S.; Ali, G.S. Sucrose-Enhanced biosynthesis of medicinally important antioxidant secondary metabolites in cell suspension cultures of Artemisia absinthium L. Bioprocess Biosyst. Eng. 2016, 39, 1945–1954. [Google Scholar] [CrossRef]
- Zhang, Y.; Huber, D.J.; Hu, M.; Jiang, G.; Gao, Z.; Xu, X.; Jiang, Y.; Zhang, Z. Delay of postharvest browning in litchi fruit by melatonin via the enhancing of antioxidative processes and oxidation repair. J. Agric. Food Chem. 2018, 66, 7475–7484. [Google Scholar] [CrossRef]
- da Graca, J.P.; Ueda, T.E.; Janegitz, T.; Vieira, S.S.; Salvador, M.C.; de Oliveira, M.C.N.; Zingaretti, S.M.; Powers, S.J.; Pickett, J.A.; Birkett, M.A.; et al. The natural plant stress elicitor Cis-Jasmone causes Cultivar-Dependent reduction in growth of the stink bug, Euschistus heros and associated changes in flavonoid concentrations in soybean, Glycine max. Phytochemistry 2016, 131, 84–91. [Google Scholar] [CrossRef]
- He, X.; Yoon, W.B.; Park, S.J.; Park, D.S.; Ahn, J. Effects of pressure level and processing time on the extraction of total phenols, flavonoids, and phenolic acids from deodeok (Codonopsis lanceolata). Food Sci. Biotechnol. 2011, 20, 499–505. [Google Scholar] [CrossRef]
- Visser, R.G.F.; Somhorst, I.; Kuipers, G.J.; Ruys, N.J.; Feenstra, W.J.; Jacobsen, E. Inhibition of the expression of the gene for Granule-Bound starch synthase in potato by antisense constructs. Mol. Gen. Genet. 1991, 225, 289–296. [Google Scholar] [CrossRef]
- Mehla, N.; Sindhi, V.; Josula, D.; Bisht, P.; Wani, S.H. An introduction to antioxidants and their roles in plant stress tolerance. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 1–23. [Google Scholar]
- Xiao, F.; Yang, Z.Q.; Han, W.; Li, Y.X.; Qiu, Y.X.; Sun, Q.; Zhang, F.M. Effects of day and night temperature on photosynthesis, antioxidant enzyme activities, and endogenous hormones in tomato leaves during the flowering stage. J. Hortic. Sci. Biotechnol. 2018, 93, 306–315. [Google Scholar] [CrossRef]
- Yuan, X.K. Effect of day/night temperature difference on chlorophyll content, photosynthesis and fluorescence parameters of tomato at fruit stage. Photosynthetica 2016, 54, 475–477. [Google Scholar] [CrossRef]
- Agrawal, M.; Krizek, D.T.; Agrawal, S.B.; Kramer, G.F.; Lee, E.H.; Mirecki, R.M.; Rowland, R.A. Influence of inverse Day-Night temperature on ozone sensitivity and selected morphological and Physiological-Responses of cucumber. J. Am. Soc. Hortic. Sci. 1993, 118, 649–654. [Google Scholar] [CrossRef]
- Wang, Y.; Noguchi, K.; Ono, N.; Inoue, S.; Terashima, I.; Kinoshita, T. Overexpression of plasma membrane H+-ATPase in guard cells promotes light-induced stomatal opening and enhances plant growth. Proc. Natl. Acad. Sci. USA 2014, 111, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Correlation between strawberry (Fragaria ananassa Duch.) productivity and Photosynthesis-Related parameters under various growth conditions. Front. Plant Sci. 2016, 7, 1607. [Google Scholar] [CrossRef]
- Gago, J.; Daloso, D.D.M.; Figueroa, C.M.; Flexas, J.; Fernie, A.R.; Nikoloski, Z. Relationships of leaf net photosynthesis, stomatal conductance, and mesophyll conductance to primary metabolism: A multispecies Meta-Analysis approach. Plant Physiol. 2016, 171, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising CO2: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| 18S | 5′-CTCAACCATAAA CGATGCCGACC-3′ | 5′-AGTTTCAGCCTTGCGACCATACTCC-3′ |
| AmGBSS | 5′-ATAACATAGCGTATCAGGG-3′ | 5′-CTCGGTCAGATTCTAACACT-3′ |
| AmRBCL | 5′-TGGCTGTTCCTATCGTCA-3′ | 5′-AAGTAATCTCCCTTTCTCCT-3′ |
| β-Actin | 5′-CGAGAAGAGCTACGAGCTACCCGATGG-3′ | 5′-CTCGGTGCTAGGGCAGTGATCTCTTTGCT-3′ |
| ClFDX | 5′-CTTCGGCGTTTCTTCGT-3′ | 5′-CTGCCAAACCCTTGATAACT-3′ |
| ClRBCL | 5′-GCTTACCCATTAGACCTTT-3′ | 5′-GGGACGACCATACTTGTT-3′ |
| DIF z (°C) | Length (cm) | Dry Weight (mg) | Leaf Area (cm−2) | No. of Leaves | Stem Diameter (mm) | Root DW: Shoot DW Ratio | Shoot Dry Weight Per Shoot Length (g·m−1) | Root Dry Weight Per Root Length (g·m−1) | Dickson’s Quality Index (×10−4) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | Leaf | ||||||||
| −10 | 9.2 ± 0.3 b y | 3.3 ± 0.2 | 18.9 ± 1.4 c | 2.0 ± 0.1 c | 4.2 ± 0.3 c | 3.5 ± 0.3 c | 3.0 ± 0.0 c | 0.62 ± 0.03 c | 0.10 ± 0.01 | 0.21 ± 0.01 b | 0.06 ± 0.00 b | 1.3 ± 0.1 c |
| 0 | 8.8 ± 0.3 b | 4.3 ± 0.2 | 28.0 ± 0.5 b | 2.8 ± 0.3 b | 6.4 ± 0.4 b | 4.6 ± 0.3 b | 3.5 ± 0.2 b | 0.78 ± 0.03 b | 0.10 ± 0.01 | 0.32 ± 0.01 a | 0.07 ± 0.01 b | 2.5 ± 0.2 b |
| 10 | 10.2 ± 0.2 a | 4.4 ± 0.5 | 34.8 ± 2.7 a | 4.3 ± 0.4 a | 7.5 ± 0.4 a | 6.2 ± 0.3 a | 3.8 ± 0.2 a | 0.90 ± 0.02 a | 0.13 ± 0.02 | 0.34 ± 0.02 a | 0.10 ± 0.02 a | 3.2 ± 0.2 a |
| F-test | ** | NS | *** | *** | *** | *** | ** | *** | NS | *** | * | *** |
| DIF z (°C) | Length (cm) | Dry Weight (mg) | Leaf Area (cm−2) | Specific Leaf Weight (g·m−2) | Stem Diameter (mm) | Root DW: Shoot DW Ratio | Shoot Dry Weight Per Shoot Length (g·m-1) | Root Dry Weight Per Root Length (g·m−1) | Dickson’s Quality Index (×10−4) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | Leaf | ||||||||
| −10 | 16.5 ± 0.4 | 3.9 ± 0.4 | 56.6 ± 3.5 b y | 11.1 ± 1.8 b | 8.3 ± 0.8 b | 6.0 ± 0.5 b | 13.8 ± 0.5 b | 1.51 ± 0.07 c | 0.19 ± 0.03 | 0.34 ± 0.02 b | 0.28 ± 0.04 b | 5.9 ± 0.4 c |
| 0 | 17.0 ± 0.5 | 3.9 ± 0.3 | 77.7 ± 8.1 ab | 17.3 ± 2.7 ab | 10.5 ± 1.2 b | 7.4 ± 0.8 b | 14.1 ± 0.7 b | 1.82 ± 0.09 b | 0.22 ± 0.02 | 0.45 ± 0.04 a | 0.44 ± 0.05 a | 9.8 ± 1.6 b |
| 10 | 17.9 ± 0.5 | 4.9 ± 0.4 | 98.9 ± 10.3 a | 20.5 ± 2.2 a | 14.4 ± 1.3 a | 8.5 ± 0.5 a | 16.8 ± 0.8 a | 2.06 ± 0.07 a | 0.21 ± 0.02 | 0.55 ± 0.05 a | 0.47 ± 0.03 a | 14.2 ± 0.8 a |
| F-test | NS | NS | ** | * | ** | * | * | *** | NS | ** | * | *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Ren, X.; Jeong, B.R. Manipulating the Difference between the Day and Night Temperatures Can Enhance the Quality of Astragalus membranaceus and Codonopsis lanceolata Plug Seedlings. Agronomy 2019, 9, 654. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9100654
Liu Y, Ren X, Jeong BR. Manipulating the Difference between the Day and Night Temperatures Can Enhance the Quality of Astragalus membranaceus and Codonopsis lanceolata Plug Seedlings. Agronomy. 2019; 9(10):654. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9100654
Chicago/Turabian StyleLiu, Ya, Xiuxia Ren, and Byoung Ryong Jeong. 2019. "Manipulating the Difference between the Day and Night Temperatures Can Enhance the Quality of Astragalus membranaceus and Codonopsis lanceolata Plug Seedlings" Agronomy 9, no. 10: 654. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9100654