
Genomic Diversity, Antimicrobial Resistance, Plasmidome, and Virulence Profiles of Salmonella Isolated from Small Specialty Crop Farms Revealed by Whole-Genome Sequencing

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
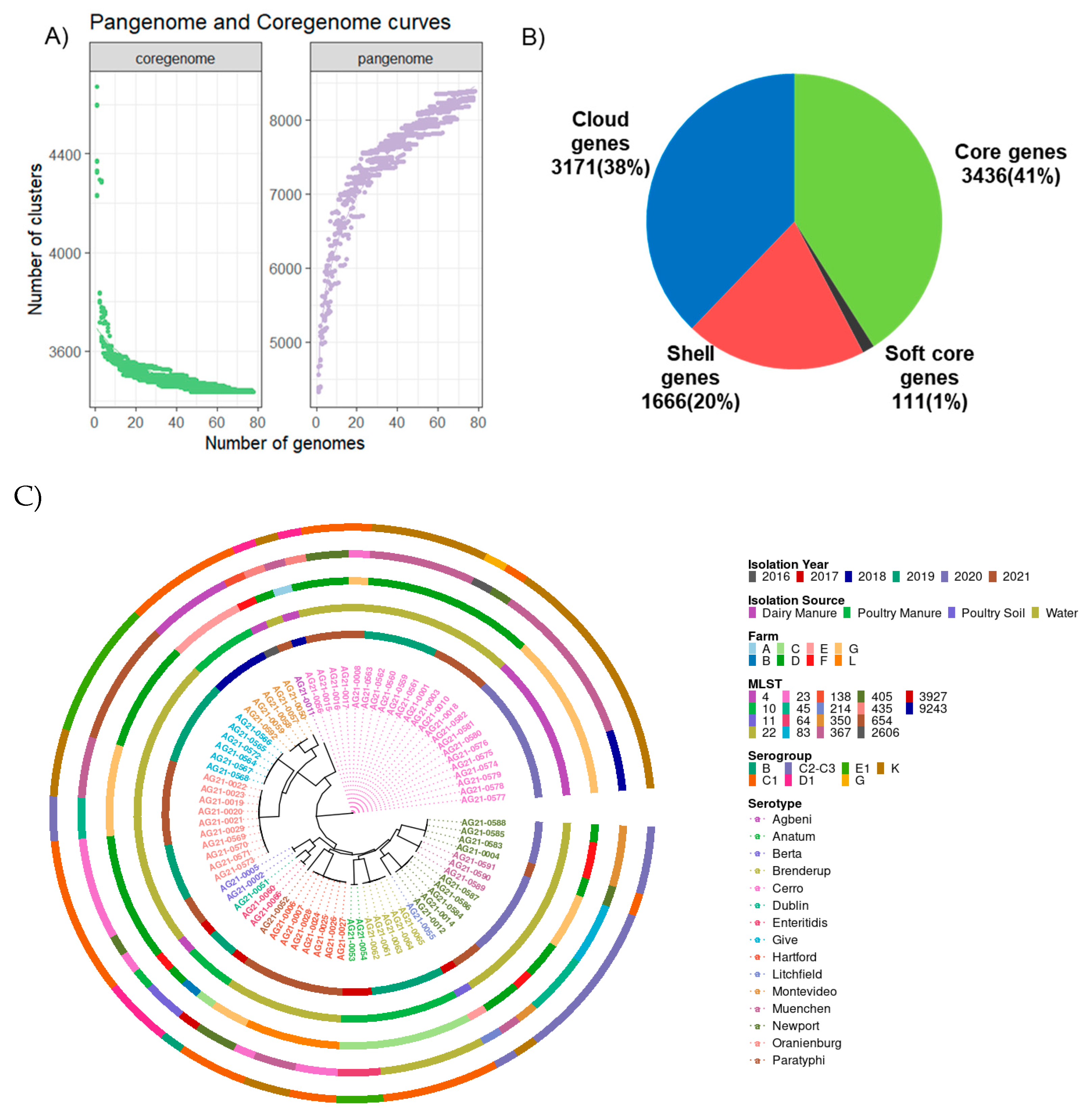
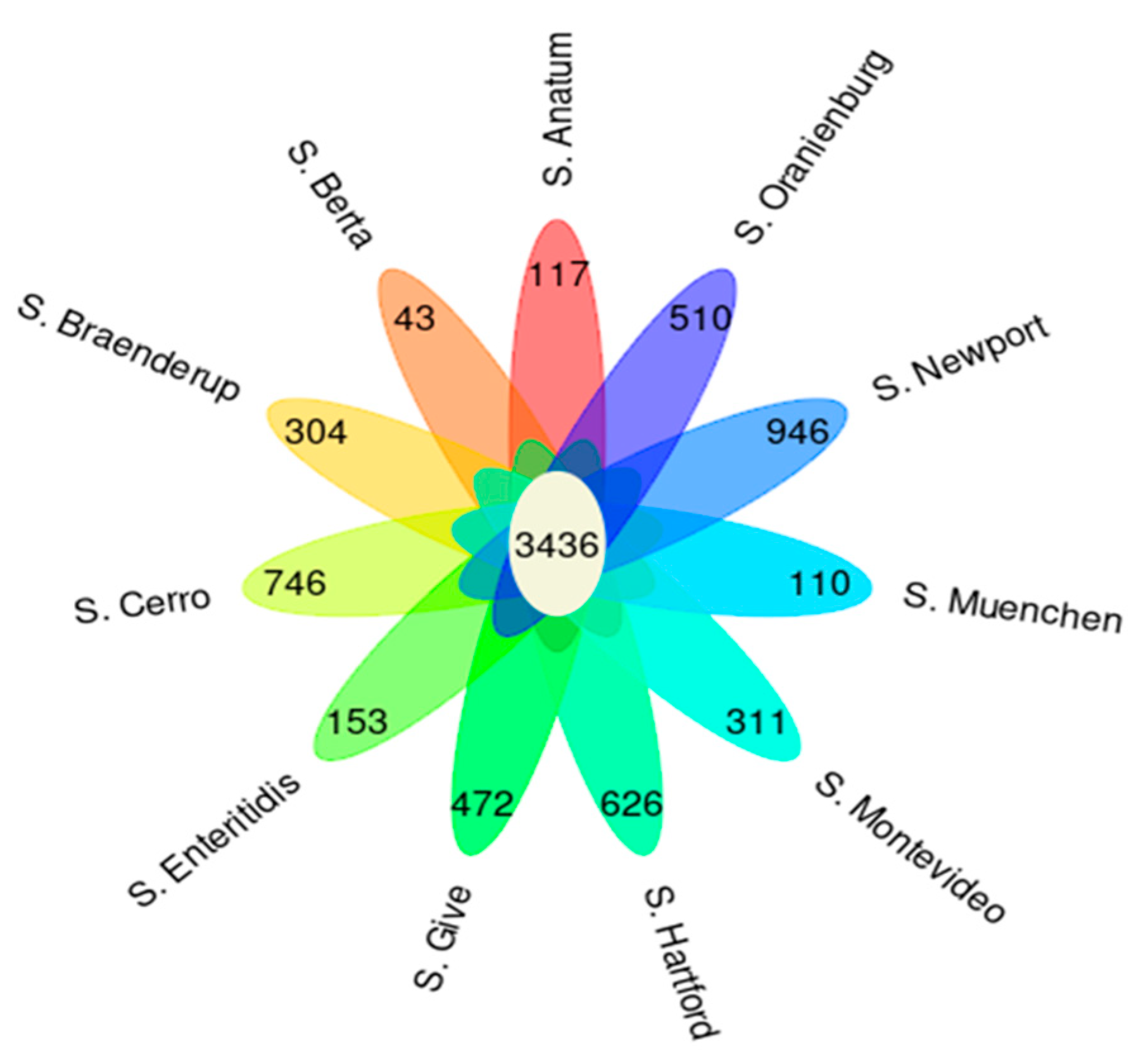
2.1. Genomic Features of Salmonella Revealed 15 Serotypes Belonging to 18 Sequence Types with 3436 Core Genes
2.2. A phylogenetic Tree Showed That All Serotypes except S. Newport form Monophyletic Clades
2.3. Pan-Genome-Wide Association Study (Pan-GWAS)
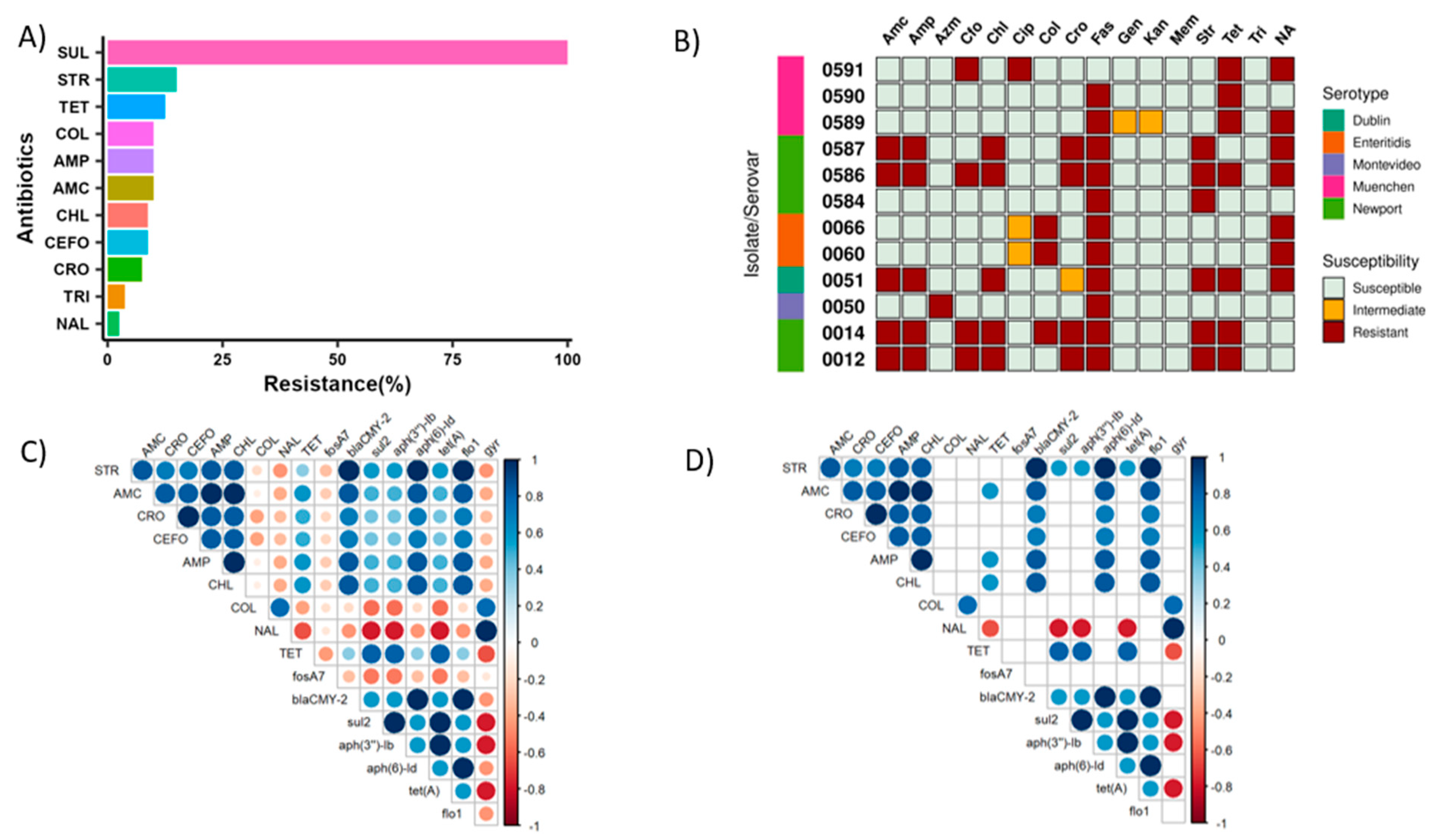
2.4. Antimicrobial Resistance Genes and Detection of the ACSSuT Cassette
2.5. Phenotypic Antimicrobial Susceptibility Testing Revealed Colistin Resistance in Some Isolates
2.6. Association between Phenotypic and Genotypic Antibiotic Resistance
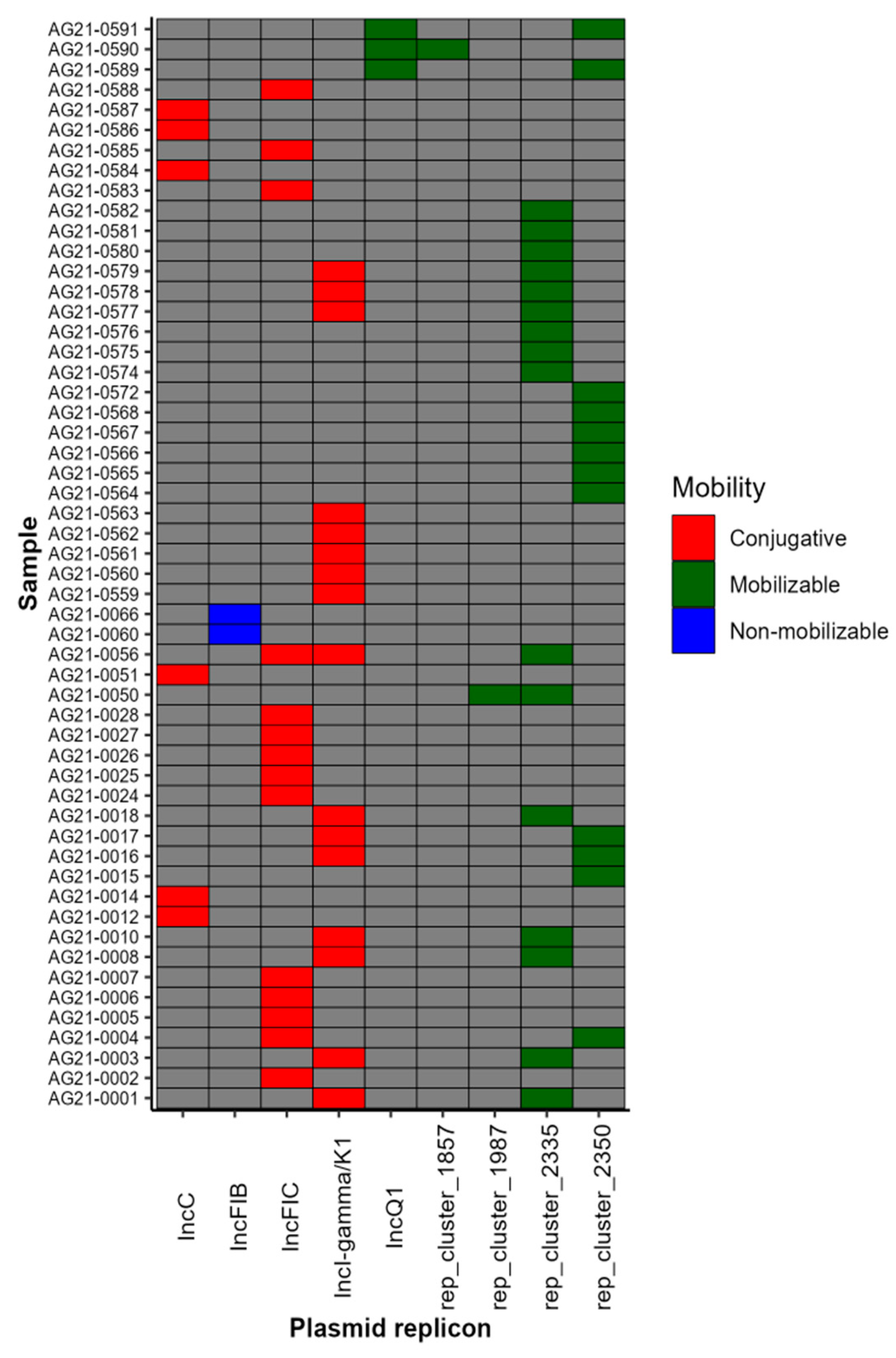
2.7. Plasmid Profile Revealed the Presence of Nine Plasmid Replicons
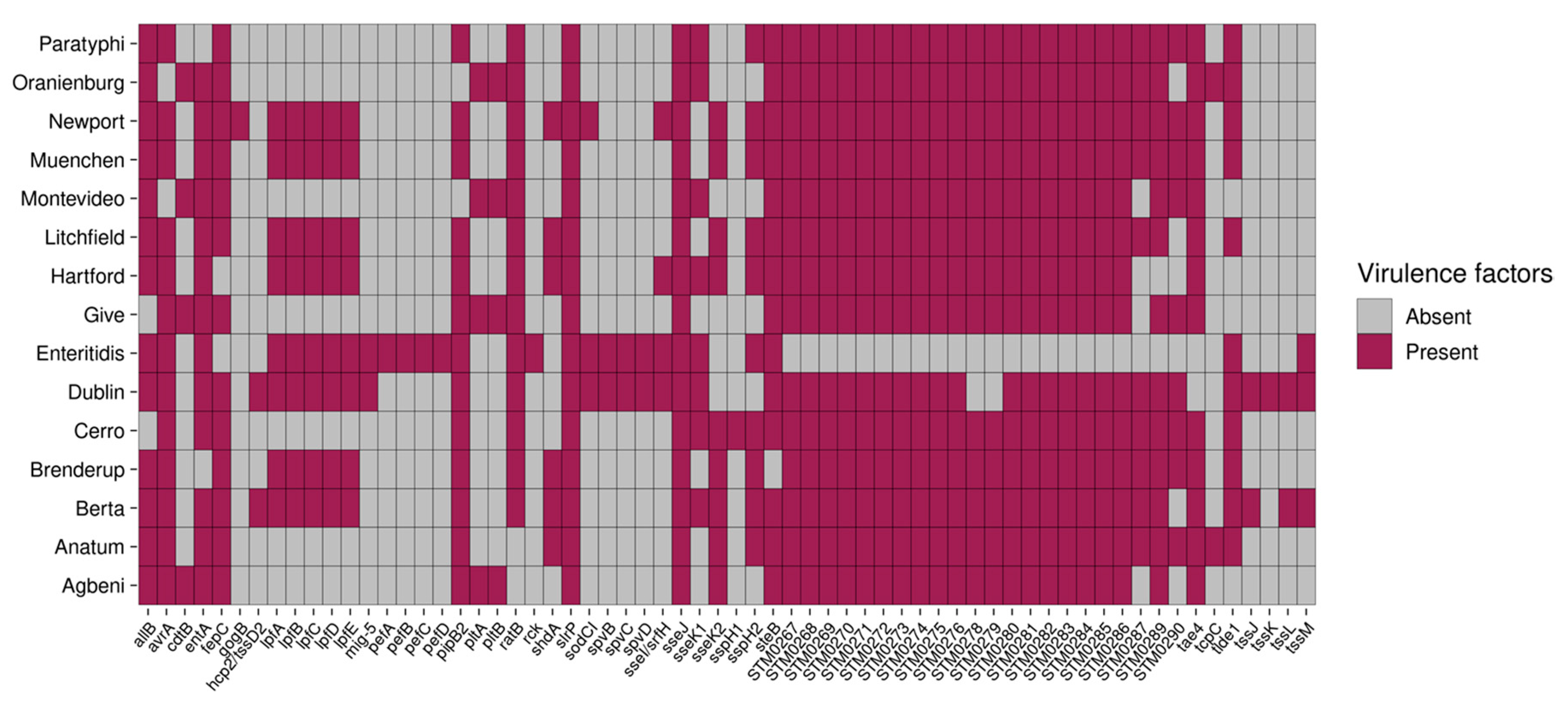
2.8. S. Enteritidis and S. Dublin Have Unique Virulence Genes
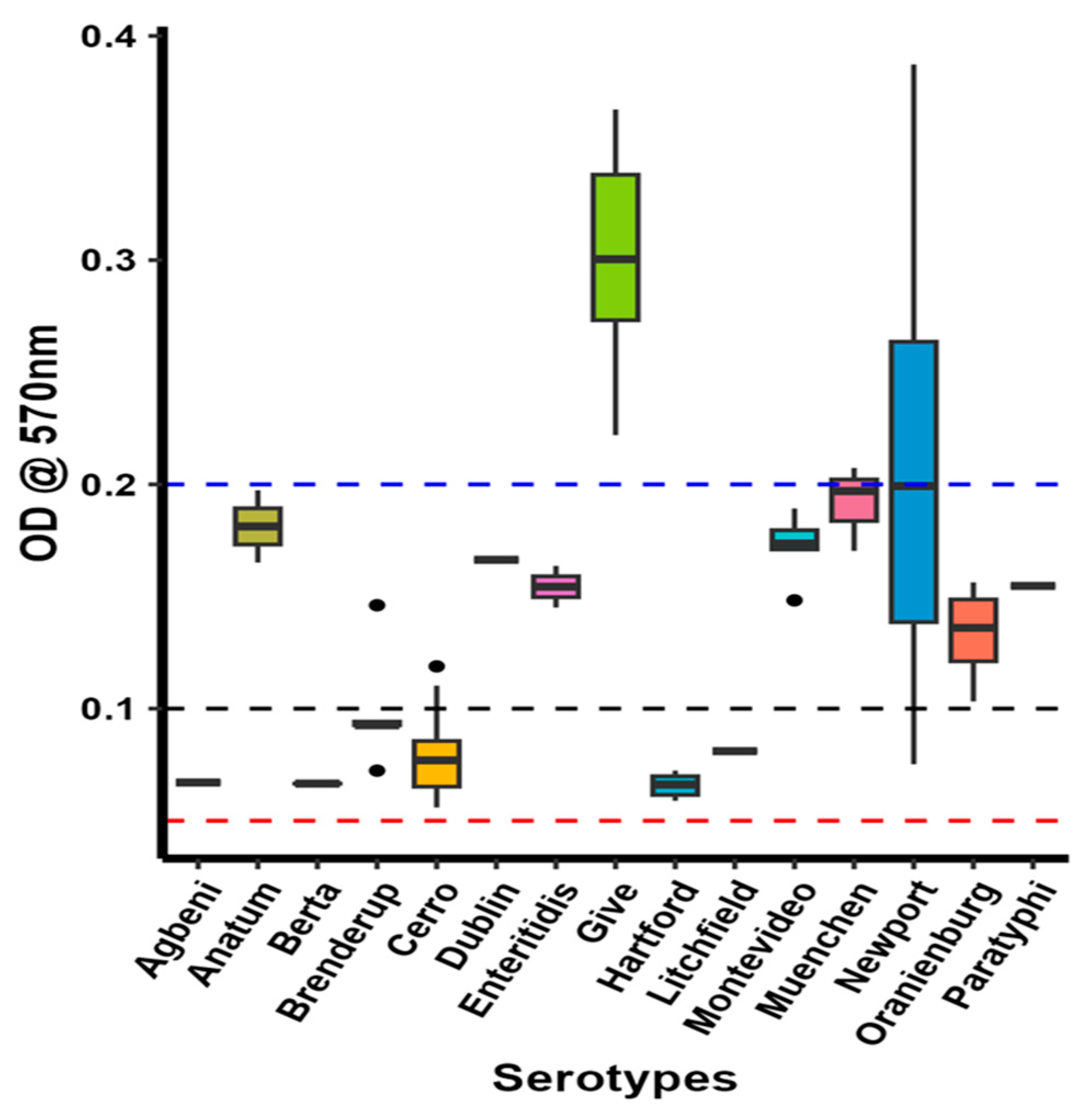
2.9. S. Give Is a Strong Biofilm Producer
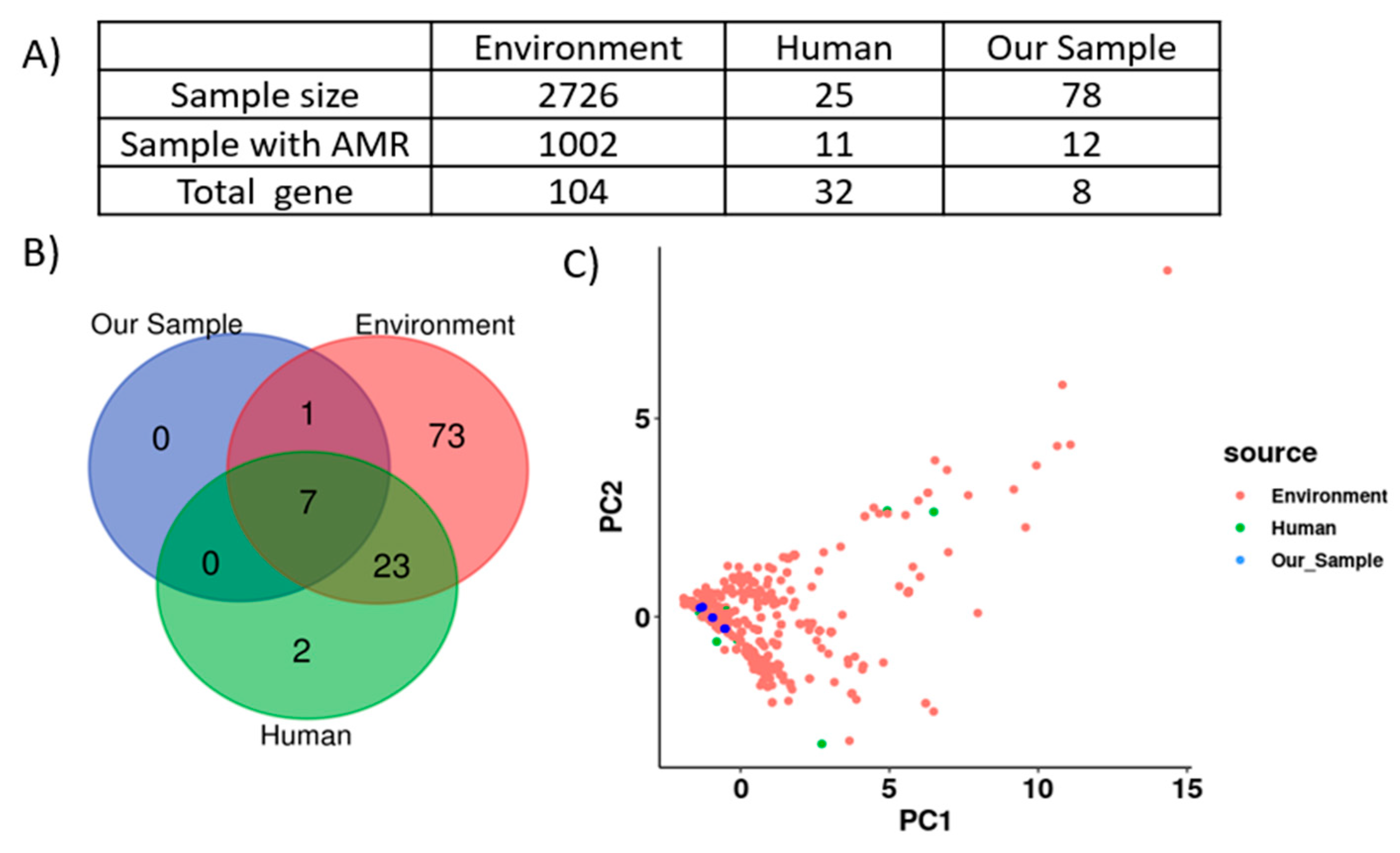
2.10. Comparative Genomic Analysis Revealed SSCFs as Potential Reservoirs for AMR Gene Transmission to Humans
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Sample Processing to Isolate Salmonella
4.3. DNA Extraction
4.4. Genome Assembly and Annotation
4.5. Pan-Genome Construction
4.6. Pan-Genome-Wide Association Studies
4.7. Serotypes, Sequence Type (ST), Average Nucleotide Identity (ANI), and Phylogenetic Structure
4.8. In Silico Antimicrobial Resistance Determinant (AMR), Virulence, and Plasmids
4.9. Phenotypic Antimicrobial Susceptibility Testing (AST)
4.10. Biofilm Assay
4.11. Comparative Genomic Analysis of AMR Genes Using Additional Salmonella Genomes
4.12. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nair, S.; Patel, V.; Hickey, T.; Maguire, C.; Greig, D.R.; Lee, W.; Godbole, G.; Grant, K.; Chattaway, M.A. Real-Time PCR Assay for Differentiation of Typhoidal and Nontyphoidal Salmonella. J. Clin. Microbiol. 2019, 57, e00167-19. [Google Scholar] [CrossRef]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Waldner, L.L.; MacKenzie, K.D.; Köster, W.; White, A.P. From exit to entry: Long-term survival and transmission of Salmonella. Pathogens 2012, 1, 128–155. [Google Scholar] [CrossRef]
- Rodriguez-Rivera, L.; Wright, E.; Siler, J.; Elton, M.; Cummings, K.; Warnick, L.; Wiedmann, M. Subtype analysis of Salmonella isolated from subclinically infected dairy cattle and dairy farm environments reveals the presence of both human-and bovine-associated subtypes. Vet. Microbiol. 2014, 170, 307–316. [Google Scholar] [CrossRef]
- Guibourdenche, M.; Roggentin, P.; Mikoleit, M.; Fields, P.I.; Bockemühl, J.; Grimont, P.A.; Weill, F.-X. Supplement 2003–2007 (No. 47) to the white-Kauffmann-Le minor scheme. Res. Microbiol. 2010, 161, 26–29. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Im, J.; Lee, J.-S.; Jeon, H.J.; Mogeni, O.D.; Kim, J.H.; Rakotozandrindrainy, R.; Baker, S.; Marks, F. The global burden and epidemiology of invasive non-typhoidal Salmonella infections. Hum. Vaccines Immunother. 2019, 15, 1421–1426. [Google Scholar] [CrossRef]
- Ray, L.C.; Collins, J.P.; Griffin, P.M.; Shah, H.J.; Boyle, M.M.; Cieslak, P.R.; Dunn, J.; Lathrop, S.; McGuire, S.; Rissman, T. Decreased Incidence of Infections Caused by Pathogens Transmitted Commonly Through Food During the COVID-19 Pandemic—Foodborne Diseases Active Surveillance Network, 10 US Sites, 2017–2020. Morb. Mortal. Wkly. Rep. 2021, 70, 1332. [Google Scholar] [CrossRef]
- Hoffmann, S.; Batz, M.B.; Morris, J.G., Jr. Annual cost of illness and quality-adjusted life year losses in the United States due to 14 foodborne pathogens. J. Food Prot. 2012, 75, 1292–1302. [Google Scholar] [CrossRef]
- Dutil, L.; Irwin, R.; Finley, R.; Ng, L.K.; Avery, B.; Boerlin, P.; Bourgault, A.-M.; Cole, L.; Daignault, D.; Desruisseau, A. Ceftiofur resistance in Salmonella enterica serovar Heidelberg from chicken meat and humans, Canada. Emerg. Infect. Dis. 2010, 16, 48. [Google Scholar] [CrossRef]
- Jibril, A.H.; Okeke, I.N.; Dalsgaard, A.; Kudirkiene, E.; Akinlabi, O.C.; Bello, M.B.; Olsen, J.E. Prevalence and risk factors of Salmonella in commercial poultry farms in Nigeria. PLoS ONE 2021, 15, e0238190. [Google Scholar] [CrossRef]
- Velasquez, C.G.; Macklin, K.S.; Kumar, S.; Bailey, M.; Ebner, P.E.; Oliver, H.F.; Martin-Gonzalez, F.S.; Singh, M. Prevalence and antimicrobial resistance patterns of Salmonella isolated from poultry farms in southeastern United States. Poult. Sci. 2018, 97, 2144–2152. [Google Scholar] [CrossRef]
- Plumb, I.D.; Schwensohn, C.A.; Gieraltowski, L.; Tecle, S.; Schneider, Z.D.; Freiman, J.; Cote, A.; Noveroske, D.; Kolsin, J.; Brandenburg, J.; et al. Outbreak of Salmonella Newport infections with decreased susceptibility to azithromycin linked to beef obtained in the United States and soft cheese obtained in Mexico—United States, 2018–2019. Morb. Mortal. Wkly. Rep. 2019, 68, 713. [Google Scholar] [CrossRef]
- Algarni, S.; Ricke, S.C.; Foley, S.L.; Han, J. The dynamics of the antimicrobial resistance mobilome of Salmonella enterica and related enteric bacteria. Front. Microbiol. 2022, 13, 859854. [Google Scholar] [CrossRef]
- Huston, C.L.; Wittum, T.E.; Love, B.C.; Keen, J.E. Prevalence of fecal shedding of Salmonella spp in dairy herds. J. Am. Vet. Med. Assoc. 2002, 220, 645–649. [Google Scholar] [CrossRef]
- Chousalkar, K.K.; Willson, N.-L. Nontyphoidal Salmonella infections acquired from poultry. Curr. Opin. Infect. Dis. 2022, 35, 431–435. [Google Scholar] [CrossRef]
- Pornsukarom, S.; Thakur, S. Assessing the impact of manure application in commercial swine farms on the transmission of antimicrobial resistant Salmonella in the environment. PLoS ONE 2016, 11, e0164621. [Google Scholar] [CrossRef]
- Greene, S.; Daly, E.; Talbot, E.; Demma, L.; Holzbauer, S.; Patel, N.; Hill, T.; Walderhaug, M.; Hoekstra, R.; Lynch, M. Recurrent multistate outbreak of Salmonella Newport associated with tomatoes from contaminated fields, 2005. Epidemiol. Infect. 2008, 136, 157–165. [Google Scholar] [CrossRef]
- Rothrock Jr, M.J.; Guard, J.Y.; Oladeinde, A. Salmonella diversity along the farm-to-fork continuum of pastured poultry flocks in the southeastern United States. Front. Anim. Sci. 2021, 2, 761930. [Google Scholar] [CrossRef]
- Fossler, C.P.; Wells, S.J.; Kaneene, J.B.; Ruegg, P.L.; Warnick, L.D.; Bender, J.B.; Eberly, L.E.; Godden, S.M.; Halbert, L.W. Herd-level factors associated with isolation of Salmonella in a multi-state study of conventional and organic dairy farms: I. Salmonella shedding in cows. Prev. Vet. Med. 2005, 70, 257–277. [Google Scholar] [CrossRef]
- Li, B.; Vellidis, G.; Liu, H.; Jay-Russell, M.; Zhao, S.; Hu, Z.; Wright, A.; Elkins, C.A. Diversity and antimicrobial resistance of Salmonella enterica isolates from surface water in southeastern United States. Appl. Environ. Microbiol. 2014, 80, 6355–6365. [Google Scholar] [CrossRef]
- Li, C.; Tyson, G.H.; Hsu, C.H.; Harrison, L.; Strain, E.; Tran, T.T.; Tillman, G.E.; Dessai, U.; McDermott, P.F.; Zhao, S. Long-read sequencing reveals evolution and acquisition of antimicrobial resistance and virulence genes in Salmonella enterica. Front. Microbiol. 2021, 12, 777817. [Google Scholar] [CrossRef]
- Kuang, X.; Hao, H.; Dai, M.; Wang, Y.; Ahmad, I.; Liu, Z.; Zonghui, Y. Serotypes and antimicrobial susceptibility of Salmonella spp. isolated from farm animals in China. Front. Microbiol. 2015, 6, 602. [Google Scholar] [CrossRef]
- Deblais, L.; Jang, H.; Kauffman, M.; Gangiredla, J.; Sawyer, M.; Basa, S.; Poelstra, J.W.; Babu, U.S.; Harrison, L.M.; Hiett, K.L. Whole genome characterization of thermophilic Campylobacter species isolated from dairy manure in small specialty crop farms of Northeast Ohio. Front. Microbiol. 2023, 14, 1074548. [Google Scholar] [CrossRef]
- Hailu, W.; Helmy, Y.A.; Carney-Knisely, G.; Kauffman, M.; Fraga, D.; Rajashekara, G. Prevalence and antimicrobial resistance profiles of foodborne pathogens isolated from dairy cattle and poultry manure amended farms in northeastern Ohio, the United States. Antibiotics 2021, 10, 1450. [Google Scholar] [CrossRef]
- Pires, A.F.; Kukielka, E.A.; Haghani, V.; Stover, J.K.; Ramos, T.D.M.; Kim, J.V.S.; Jay-Russell, M.T. Survey of farmers market managers in California: Food safety perspectives. J. Ext. 2020, 58, 17. [Google Scholar] [CrossRef]
- Andoh, L.A.; Dalsgaard, A.; Obiri-Danso, K.; Newman, M.J.; Barco, L.; Olsen, J.E. Prevalence and antimicrobial resistance of Salmonella serovars isolated from poultry in Ghana. Epidemiol. Infect. 2016, 144, 3288–3299. [Google Scholar] [CrossRef]
- Graham, J.P.; Eisenberg, J.N.; Trueba, G.; Zhang, L.; Johnson, T.J. Small-scale food animal production and antimicrobial resistance: Mountain, molehill, or something in-between? Environ. Health Perspect. 2017, 125, 104501. [Google Scholar] [CrossRef]
- Chen, J.; Ed-Dra, A.; Zhou, H.; Wu, B.; Zhang, Y.; Yue, M. Antimicrobial resistance and genomic investigation of non-typhoidal Salmonella isolated from outpatients in Shaoxing city, China. Front. Public Health 2022, 10, 988317. [Google Scholar] [CrossRef]
- Fortini, D.; Owczarek, S.; Dionisi, A.M.; Lucarelli, C.; Arena, S.; Carattoli, A.; Enter-Net Italia Colistin Resistance Study Group; Villa, L.; García-Fernández, A. Colistin Resistance Mechanisms in Human Salmonella enterica Strains Isolated by the National Surveillance Enter-Net Italia (2016–2018). Antibiotics 2022, 11, 102. [Google Scholar] [CrossRef]
- Li, Y.; Kang, X.; Ed-Dra, A.; Zhou, X.; Jia, C.; Müller, A.; Liu, Y.; Kehrenberg, C.; Yue, M. Genome-based assessment of antimicrobial resistance and virulence potential of isolates of non-pullorum/gallinarum Salmonella Serovars recovered from dead poultry in China. Microbiol. Spectr. 2022, 10, e00965-22. [Google Scholar] [CrossRef]
- Cummings, K.J.; Warnick, L.D.; Davis, M.A.; Eckmann, K.; Gröhn, Y.T.; Hoelzer, K.; MacDonald, K.; Root, T.P.; Siler, J.D.; McGuire, S.M. Farm animal contact as risk factor for transmission of bovine-associated Salmonella subtypes. Emerg. Infect. Dis. 2012, 18, 1929. [Google Scholar] [CrossRef]
- Carroll, L.M.; Buehler, A.J.; Gaballa, A.; Siler, J.D.; Cummings, K.J.; Cheng, R.A.; Wiedmann, M. Monitoring the Microevolution of Salmonella enterica in Healthy Dairy Cattle Populations at the Individual Farm Level Using Whole-Genome Sequencing. Front. Microbiol. 2021, 12, 763669. [Google Scholar] [CrossRef]
- Micallef, S.A.; Goldstein, R.E.R.; George, A.; Kleinfelter, L.; Boyer, M.S.; McLaughlin, C.R.; Estrin, A.; Ewing, L.; Beaubrun, J.J.-G.; Hanes, D.E. Occurrence and antibiotic resistance of multiple Salmonella serotypes recovered from water, sediment and soil on mid-Atlantic tomato farms. Environ. Res. 2012, 114, 31–39. [Google Scholar] [CrossRef]
- Tack, D.M.; Ray, L.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Rissman, T.; Jervis, R.; Lathrop, S.; Muse, A.; Duwell, M. Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 US Sites, 2016–2019. Morb. Mortal. Wkly. Rep. 2020, 69, 509. [Google Scholar] [CrossRef]
- Boyd, D.; Cloeckaert, A.; Chaslus-Dancla, E.; Mulvey, M.R. Characterization of variant Salmonella genomic island 1 multidrug resistance regions from serovars Typhimurium DT104 and Agona. Antimicrob. Agents Chemother. 2002, 46, 1714–1722. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, S.; White, D.G.; Schroeder, C.M.; Lu, R.; Yang, H.; McDermott, P.F.; Ayers, S.; Meng, J. Characterization of multiple-antimicrobial-resistant Salmonella serovars isolated from retail meats. Appl. Environ. Microbiol. 2004, 70, 1–7. [Google Scholar] [CrossRef]
- Cloeckaert, A.; Chaslus-Dancla, E. Mechanisms of quinolone resistance in Salmonella. Vet. Res. 2001, 32, 291–300. [Google Scholar] [CrossRef]
- Gharaibeh, M.H.; Shatnawi, S.Q. An overview of colistin resistance, mobilized colistin resistance genes dissemination, global responses, and the alternatives to colistin: A review. Vet. World 2019, 12, 1735. [Google Scholar] [CrossRef]
- Silva, C.; Puente, J.L.; Calva, E. Salmonella virulence plasmid: Pathogenesis and ecology. Pathog. Dis. 2017, 75, ftx070. [Google Scholar] [CrossRef]
- Guiney, D.G.; Fierer, J. The role of the spv genes in Salmonella pathogenesis. Front. Microbiol. 2011, 2, 129. [Google Scholar] [CrossRef]
- Kuijpers, A.F.; Bonacic Marinovic, A.A.; Wijnands, L.M.; Delfgou-van Asch, E.H.; van Hoek, A.H.; Franz, E.; Pielaat, A. Phenotypic prediction: Linking in vitro virulence to the genomics of 59 Salmonella enterica strains. Front. Microbiol. 2019, 9, 3182. [Google Scholar] [CrossRef]
- Miller, R.; Wiedmann, M. Dynamic duo—The Salmonella cytolethal distending toxin combines ADP-ribosyltransferase and nuclease activities in a novel form of the cytolethal distending toxin. Toxins 2016, 8, 121. [Google Scholar] [CrossRef]
- Wang, Y.-T.; Lei, C.-W.; Liu, S.-Y.; Chen, X.; Gao, Y.-F.; Zhang, Y.; Tang, Y.; Zhang, A.; Yang, X.; Wang, H.-N. Tracking Salmonella enterica by whole genome sequencing of isolates recovered from broiler chickens in a poultry production system. Int. J. Food Microbiol. 2021, 350, 109246. [Google Scholar] [CrossRef]
- Grinnell, M.; Provo, G.; Marsden-Haug, N.; Stigi, K.A.; DeBess, E.; Kissler, B.; Crarey, E.; Tate, H.; Pringle, J.; Grass, J. Outbreak of Salmonella Heidelberg infections linked to a single poultry producer—13 states, 2012–2013. Morb. Mortal. Wkly. Rep. 2013, 62, 553. [Google Scholar]
- Rajagopal, R.; Mini, M. Outbreaks of salmonellosis in three different poultry farms of Kerala, India. Asian Pac. J. Trop. Biomed. 2013, 3, 496–500. [Google Scholar] [CrossRef]
- You, Y.; Rankin, S.C.; Aceto, H.W.; Benson, C.E.; Toth, J.D.; Dou, Z. Survival of Salmonella enterica serovar Newport in manure and manure-amended soils. Appl. Environ. Microbiol. 2006, 72, 5777–5783. [Google Scholar] [CrossRef]
- Liao, C.H.; Shollenberger, L. Survivability and long-term preservation of bacteria in water and in phosphate-buffered saline. Lett. Appl. Microbiol. 2003, 37, 45–50. [Google Scholar] [CrossRef]
- Toth, J.D.; Aceto, H.W.; Rankin, S.C.; Dou, Z. Survival characteristics of Salmonella enterica serovar Newport in the dairy farm environment. J. Dairy Sci. 2011, 94, 5238–5246. [Google Scholar] [CrossRef]
- Uesugi, A.R.; Danyluk, M.D.; Mandrell, R.E.; Harris, L.J. Isolation of Salmonella Enteritidis phage type 30 from a single almond orchard over a 5-year period. J. Food Prot. 2007, 70, 1784–1789. [Google Scholar] [CrossRef]
- Wilson, A.; Fox, E.M.; Fegan, N.; Kurtböke, D.Í. Comparative genomics and phenotypic investigations into antibiotic, heavy metal, and disinfectant susceptibilities of Salmonella enterica strains isolated in Australia. Front. Microbiol. 2019, 10, 1620. [Google Scholar] [CrossRef]
- Cummings, K.J.; Warnick, L.D.; Elton, M.; Rodriguez-Rivera, L.D.; Siler, J.D.; Wright, E.M.; Gröhn, Y.T.; Wiedmann, M. Salmonella enterica serotype Cerro among dairy cattle in New York: An emerging pathogen? Foodborne Pathog. Dis. 2010, 7, 659–665. [Google Scholar] [CrossRef]
- Cohn, A.R.; Orsi, R.H.; Carroll, L.M.; Liao, J.; Wiedmann, M.; Cheng, R.A. Salmonella enterica serovar Cerro displays a phylogenetic structure and genomic features consistent with virulence attenuation and adaptation to cattle. Front. Microbiol. 2022, 13, 4725. [Google Scholar] [CrossRef]
- Tang, B.; Elbediwi, M.; Nambiar, R.B.; Yang, H.; Lin, J.; Yue, M. Genomic Characterization of Antimicrobial-Resistant Salmonella enterica in Duck, Chicken, and Pig Farms and Retail Markets in Eastern China. Microbiol. Spectr. 2022, 10, e01257-22. [Google Scholar] [CrossRef]
- Brennan, M.L.; Kemp, R.; Christley, R.M. Direct and indirect contacts between cattle farms in north-west England. Prev. Vet. Med. 2008, 84, 242–260. [Google Scholar] [CrossRef]
- Ford, L.; Carter, G.P.; Wang, Q.; Seemann, T.; Sintchenko, V.; Glass, K.; Williamson, D.A.; Howard, P.; Valcanis, M.; Castillo, C.F.S. Incorporating whole-genome sequencing into public health surveillance: Lessons from prospective sequencing of Salmonella Typhimurium in Australia. Foodborne Pathog. Dis. 2018, 15, 161–167. [Google Scholar] [CrossRef]
- Sangal, V.; Harbottle, H.; Mazzoni, C.J.; Helmuth, R.; Guerra, B.; Didelot, X.; Paglietti, B.; Rabsch, W.; Brisse, S.; Weill, F.-X. Evolution and population structure of Salmonella enterica serovar Newport. J. Bacteriol. 2010, 192, 6465–6476. [Google Scholar] [CrossRef]
- Saidenberg, A.B.; Stegger, M.; Semmler, T.; Rocha, V.G.; Cunha, M.P.; Souza, V.A.; Cristina Menão, M.; Milanelo, L.; Petri, B.S.S.; Knöbl, T. Salmonella Newport outbreak in Brazilian parrots: Confiscated birds from the illegal pet trade as possible zoonotic sources. Environ. Microbiol. Rep. 2021, 13, 702–707. [Google Scholar] [CrossRef]
- Paudyal, N.; Pan, H.; Wu, B.; Zhou, X.; Zhou, X.; Chai, W.; Wu, Q.; Li, S.; Li, F.; Gu, G. Persistent asymptomatic human infections by Salmonella enterica serovar Newport in China. Msphere 2020, 5, 10–1128. [Google Scholar] [CrossRef]
- Hur, J.; Kim, J.H.; Park, J.H.; Lee, Y.-J.; Lee, J.H. Molecular and virulence characteristics of multi-drug resistant Salmonella Enteritidis strains isolated from poultry. Vet. J. 2011, 189, 306–311. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Wiuff, C.; Mølbak, K.; Threlfall, E.J. Is it time to change fluoroquinolone breakpoints for Salmonella spp.? Antimicrob. Agents Chemother. 2003, 47, 827–829. [Google Scholar] [CrossRef]
- Baucheron, S.; Le Hello, S.; Doublet, B.; Giraud, E.; Weill, F.-X.; Cloeckaert, A. ramR mutations affecting fluoroquinolone susceptibility in epidemic multidrug-resistant Salmonella enterica serovar Kentucky ST198. Front. Microbiol. 2013, 4, 213. [Google Scholar] [CrossRef] [PubMed]
- Koide, K.; San, L.L.; Pachanon, R.; Park, J.-H.; Ouchi, Y.; Kongsoi, S.; Utrarachkij, F.; Nakajima, C.; Suzuki, Y. Amino acid substitution Ser83Ile in GyrA of DNA gyrases confers high-level quinolone resistance to nontyphoidal Salmonella without loss of supercoiling activity. Microb. Drug Resist. 2021, 27, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Elbediwi, M.; Li, Y.; Paudyal, N.; Pan, H.; Li, X.; Xie, S.; Rajkovic, A.; Feng, Y.; Fang, W.; Rankin, S.C. Global burden of colistin-resistant bacteria: Mobilized colistin resistance genes study (1980–2018). Microorganisms 2019, 7, 461. [Google Scholar] [CrossRef] [PubMed]
- Jovčić, B.; Novović, K.; Filipić, B.; Velhner, M.; Todorović, D.; Matović, K.; Rašić, Z.; Nikolić, S.; Kiškarolj, F.; Kojić, M. Genomic characteristics of colistin-resistant Salmonella enterica subsp. enterica Serovar infantis from poultry farms in the Republic of Serbia. Antibiotics 2020, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Dia, N.M.; Gautret, P.; Benkouiten, S.; Belhouchat, K.; Drali, T.; Parola, P.; Brouqui, P.; Memish, Z.; Raoult, D.; et al. Acquisition of extended-spectrum cephalosporin-and colistin-resistant Salmonella enterica subsp. enterica serotype Newport by pilgrims during Hajj. Int. J. Antimicrob. Agents 2015, 45, 600–604. [Google Scholar] [CrossRef]
- Saleh, N.M.; Hesham, M.S.; Amin, M.A.; Samir Mohamed, R. Acquisition of colistin resistance links cell membrane thickness alteration with a point mutation in the lpxD gene in Acinetobacter baumannii. Antibiotics 2020, 9, 164. [Google Scholar] [CrossRef]
- Oliva, M.; Monno, R.; D’Addabbo, P.; Pesole, G.; Dionisi, A.; Scrascia, M.; Chiara, M.; Horner, D.; Manzari, C.; Luzzi, I. A novel group of IncQ1 plasmids conferring multidrug resistance. Plasmid 2017, 89, 22–26. [Google Scholar] [CrossRef]
- Solghan, S.M.; Dumas, N.B.; Root, T.P.; Quinlan, T.M.; Armstrong, L.R.; Spina, N.L.; Zansky, S.M. Multidrug-resistant nontyphoidal Salmonella in New York state’s foodborne diseases active surveillance network counties. Foodborne Pathog. Dis. 2010, 7, 167–173. [Google Scholar] [CrossRef]
- Cao, G.; Allard, M.; Hoffmann, M.; Muruvanda, T.; Luo, Y.; Payne, J.; Meng, K.; Zhao, S.; McDermott, P.; Brown, E. Sequence analysis of IncA/C and IncI1 plasmids isolated from multidrug-resistant Salmonella Newport using single-molecule real-time sequencing. Foodborne Pathog. Dis. 2018, 15, 361–371. [Google Scholar] [CrossRef]
- Carraro, N.; Durand, R.; Rivard, N.; Anquetil, C.; Barrette, C.; Humbert, M.; Burrus, V. Salmonella genomic island 1 (SGI1) reshapes the mating apparatus of IncC conjugative plasmids to promote self-propagation. PLoS Genet. 2017, 13, e1006705. [Google Scholar] [CrossRef]
- Fernández-Alarcón, C.; Singer, R.S.; Johnson, T.J. Comparative genomics of multidrug resistance-encoding IncA/C plasmids from commensal and pathogenic Escherichia coli from multiple animal sources. PLoS ONE 2011, 6, e23415. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Santos, R.L.; Keestra, A.M.; Adams, L.G.; Bäumler, A.J. Salmonella enterica serotype Typhimurium ShdA is an outer membrane fibronectin-binding protein that is expressed in the intestine. Mol. Microbiol. 2002, 43, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Boyen, F.; Pasmans, F.; Donne, E.; Van Immerseel, F.; Morgan, E.; Adriaensen, C.; Hernalsteens, J.-P.; Wallis, T.; Ducatelle, R.; Haesebrouck, F. The fibronectin binding protein ShdA is not a prerequisite for long term faecal shedding of Salmonella typhimurium in pigs. Vet. Microbiol. 2006, 115, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhuang, L.; Wang, C.; Zhang, P.; Zhang, T.; Shao, H.; Han, X.; Gong, J. Virulence gene distribution of Salmonella Pullorum isolates recovered from chickens in China (1953–2015). Avian Dis. 2018, 62, 431–436. [Google Scholar] [CrossRef]
- Fierer, J. Extra-intestinal Salmonella infections: The significance of spv genes. Clin. Infect. Dis. 2001, 32, 519–520. [Google Scholar] [CrossRef]
- Hsu, C.-H.; Li, C.; Hoffmann, M.; McDermott, P.; Abbott, J.; Ayers, S.; Tyson, G.H.; Tate, H.; Yao, K.; Allard, M. Comparative genomic analysis of virulence, antimicrobial resistance, and plasmid profiles of Salmonella Dublin isolated from sick cattle, retail beef, and humans in the United States. Microb. Drug Resist. 2019, 25, 1238–1249. [Google Scholar] [CrossRef]
- Cao, G.; Zhao, S.; Kuang, D.; Hsu, C.-H.; Yin, L.; Luo, Y.; Chen, Z.; Xu, X.; Strain, E.; McDermott, P. Geography shapes the genomics and antimicrobial resistance of Salmonella enterica Serovar Enteritidis isolated from humans. Sci. Rep. 2023, 13, 1331. [Google Scholar] [CrossRef]
- Figueiredo, R.; Card, R.; Nunes, C.; AbuOun, M.; Bagnall, M.C.; Nunez, J.; Mendonça, N.; Anjum, M.F.; da Silva, G.J. Virulence characterization of Salmonella enterica by a new microarray: Detection and evaluation of the cytolethal distending toxin gene activity in the unusual host S. Typhimurium. PLoS ONE 2015, 10, e0135010. [Google Scholar] [CrossRef]
- Miller, R.A.; Wiedmann, M. The cytolethal distending toxin produced by nontyphoidal Salmonella serotypes Javiana, Montevideo, Oranienburg, and Mississippi induces DNA damage in a manner similar to that of serotype Typhi. MBio 2016, 7, e02109-16. [Google Scholar] [CrossRef]
- Rodriguez-Rivera, L.D.; Bowen, B.M.; den Bakker, H.C.; Duhamel, G.E.; Wiedmann, M. Characterization of the cytolethal distending toxin (typhoid toxin) in non-typhoidal Salmonella serovars. Gut Pathog. 2015, 7, 19. [Google Scholar] [CrossRef]
- Villa-Rojas, R.; Zhu, M.J.; Paul, N.C.; Gray, P.; Xu, J.; Shah, D.H.; Tang, J. Biofilm forming Salmonella strains exhibit enhanced thermal resistance in wheat flour. Food Control 2017, 73, 689–695. [Google Scholar] [CrossRef]
- Steenackers, H.; Hermans, K.; Vanderleyden, J.; De Keersmaecker, S.C. Salmonella biofilms: An overview on occurrence, structure, regulation and eradication. Food Res. Int. 2012, 45, 502–531. [Google Scholar] [CrossRef]
- Den Bakker, H.C.; Moreno Switt, A.I.; Govoni, G.; Cummings, C.A.; Ranieri, M.L.; Degoricija, L.; Hoelzer, K.; Rodriguez-Rivera, L.D.; Brown, S.; Bolchacova, E. Genome sequencing reveals diversification of virulence factor content and possible host adaptation in distinct subpopulations of Salmonella enterica. BMC Genom. 2011, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Avila-Novoa, M.-G.; Guerrero-Medina, P.-J.; Navarrete-Sahagún, V.; Gómez-Olmos, I.; Velázquez-Suárez, N.-Y.; De la Cruz-Color, L.; Gutiérrez-Lomelí, M. Biofilm formation by multidrug-resistant serotypes of Salmonella isolated from fresh products: Effects of nutritional and environmental conditions. Appl. Sci. 2021, 11, 3581. [Google Scholar] [CrossRef]
- Moraes, J.O.; Cruz, E.A.; Souza, E.G.; Oliveira, T.C.; Alvarenga, V.O.; Peña, W.E.; Sant’Ana, A.S.; Magnani, M. Predicting adhesion and biofilm formation boundaries on stainless steel surfaces by five Salmonella enterica strains belonging to different serovars as a function of pH, temperature and NaCl concentration. Int. J. Food Microbiol. 2018, 281, 90–100. [Google Scholar] [CrossRef]
- Castañeda, M.d.R.A.; Sarnacki, S.H.; Llana, M.N.; Guerra, A.G.L.; Giacomodonato, M.N.; Cerquetti, M.C. Dam methylation is required for efficient biofilm production in Salmonella enterica serovar Enteritidis. Int. J. Food Microbiol. 2015, 193, 15–22. [Google Scholar] [CrossRef]
- Baek, J.; Yoon, H. Cyclic di-GMP Modulates a Metabolic Flux for Carbon Utilization in Salmonella enterica Serovar Typhimurium. Microbiol. Spectr. 2023, 11, e03685-22. [Google Scholar] [CrossRef]
- Houot, L.; Chang, S.; Pickering, B.S.; Absalon, C.; Watnick, P.I. The phosphoenolpyruvate phosphotransferase system regulates Vibrio cholerae biofilm formation through multiple independent pathways. J. Bacteriol. 2010, 192, 3055–3067. [Google Scholar] [CrossRef]
- Davies, R.; Wales, A. Investigations into Salmonella contamination in poultry feedmills in the United Kingdom. J. Appl. Microbiol. 2010, 109, 1430–1440. [Google Scholar] [CrossRef]
- Freschi, C.R.; Carvalho, L.F.d.O.; Oliveira, C.J.B.d. Comparison of DNA-extraction methods and selective enrichment broths on the detection of Salmonella Typhimurium in swine feces by polymerase chain reaction (PCR). Braz. J. Microbiol. 2005, 36, 363–367. [Google Scholar] [CrossRef]
- Rahn, K.; De Grandis, S.; Clarke, R.; McEwen, S.; Galan, J.; Ginocchio, C.; Curtiss Iii, R.; Gyles, C. Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell. Probes 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Ribeiro, P.G.; Torres Jiménez, M.F.; Andermann, T.; Antonelli, A.; Bacon, C.D.; Matos-Maraví, P. A bioinformatic platform to integrate target capture and whole genome sequences of various read depths for phylogenomics. Mol. Ecol. 2021, 30, 6021–6035. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness. In Gene Prediction; Springer: Berlin/Heidelberg, Germany, 2019; pp. 227–245. [Google Scholar]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner; Lawrence Berkeley National Lab. (LBNL): Berkeley, CA, USA, 2014.
- Fenske, G.J.; Thachil, A.; McDonough, P.L.; Glaser, A.; Scaria, J. Geography shapes the population genomics of Salmonella enterica Dublin. Genome Biol. Evol. 2019, 11, 2220–2231. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.-I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Brynildsrud, O.; Bohlin, J.; Scheffer, L.; Eldholm, V. Rapid scoring of genes in microbial pan-genome-wide association studies with Scoary. Genome Biol. 2016, 17, 238. [Google Scholar] [CrossRef]
- Greening, S.S.; Zhang, J.; Midwinter, A.C.; Wilkinson, D.A.; McDougall, S.; Gates, M.C.; French, N.P. The genetic relatedness and antimicrobial resistance patterns of mastitis-causing Staphylococcus aureus strains isolated from New Zealand dairy cattle. Vet. Sci. 2021, 8, 287. [Google Scholar] [CrossRef]
- Zhang, S.; den Bakker, H.C.; Li, S.; Chen, J.; Dinsmore, B.A.; Lane, C.; Lauer, A.; Fields, P.I.; Deng, X. SeqSero2: Rapid and improved Salmonella serotype determination using whole-genome sequencing data. Appl. Environ. Microbiol. 2019, 85, e01746-19. [Google Scholar] [CrossRef]
- Uelze, L.; Grützke, J.; Borowiak, M.; Hammerl, J.A.; Juraschek, K.; Deneke, C.; Tausch, S.H.; Malorny, B. Typing methods based on whole genome sequencing data. One Health Outlook 2020, 2, 3. [Google Scholar] [CrossRef]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef]
- Gardner, S.N.; Slezak, T.; Hall, B.G. kSNP3. 0: SNP detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinformatics 2015, 31, 2877–2878. [Google Scholar] [CrossRef]
- Bernard, G.; Greenfield, P.; Ragan, M.A.; Chan, C.X. k-mer similarity, networks of microbial genomes, and taxonomic rank. Msystems 2018, 3, e00257-18. [Google Scholar] [CrossRef]
- Bernard, G.; Chan, C.X.; Ragan, M.A. Alignment-free microbial phylogenomics under scenarios of sequence divergence, genome rearrangement and lateral genetic transfer. Sci. Rep. 2016, 6, 28970. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Brown, C.T.; Irber, L. sourmash: A library for MinHash sketching of DNA. J. Open Source Softw. 2016, 1, 27. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef]
- Robertson, J.; Nash, J.H. MOB-suite: Software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb. Genom. 2018, 4, e000206. [Google Scholar] [CrossRef]
- Matlock, W.; Chau, K.K.; AbuOun, M.; Stubberfield, E.; Barker, L.; Kavanagh, J.; Pickford, H.; Gilson, D.; Smith, R.P.; Gweon, H.S. Genomic network analysis of environmental and livestock F-type plasmid populations. ISME J. 2021, 15, 2322–2335. [Google Scholar] [CrossRef]
- Douarre, P.-E.; Mallet, L.; Radomski, N.; Felten, A.; Mistou, M.-Y. Analysis of COMPASS, a new comprehensive plasmid database revealed prevalence of multireplicon and extensive diversity of IncF plasmids. Front. Microbiol. 2020, 11, 483. [Google Scholar] [CrossRef]
- Patel, J.B.; Cockerill, F.; Bradford, P.A. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Fifth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- El Hag, M.; Feng, Z.; Su, Y.; Wang, X.; Yassin, A.; Chen, S.; Peng, D.; Liu, X. Contribution of the csgA and bcsA genes to Salmonella enterica serovar Pullorum biofilm formation and virulence. Avian Pathol. 2017, 46, 541–547. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Salmonella-Positive Samples | Farm |
|---|---|---|
| 2016 | 1 | Dairy manure |
| 2017 | 2 | Poultry manure |
| 2017 | 1 | Dairy manure |
| 2017 | 1 | Soil |
| 2018 | 1 | Poultry manure |
| 2018 | 1 | Dairy manure |
| 2019 | 3 | Poultry manure |
| 2019 | 3 | Water |
| 2020 | 3 | Water |
| 2020 | 3 | Dairy manure |
| 2021 | 10 | Water |
| Total | 29 |
| Serotype | Coverage | Size (Mb) | Contigs | GC % | BUSCO | N50 | Completeness | Contamination | Gene |
|---|---|---|---|---|---|---|---|---|---|
| S. Agbeni | 35.4 | 4.5 | 42 | 52.2 | 98.65 | 382,607 | 100.0 | 0.3 | 4292 |
| S. Anatum | 66.9 | 4.6 | 24 | 52.2 | 98.65 | 562,293 | 100.0 | 0.0 | 4338 |
| S. Berta | 69.4 | 4.7 | 41 | 52.2 | 98.65 | 289,914 | 100.0 | 0.0 | 4465 |
| S. Brenderup | 76.0 | 4.7 | 27 | 52.2 | 98.65 | 497,120 | 100.0 | 0.1 | 4419 |
| S. Cerro | 69.3 | 4.6 | 61 | 52.2 | 98.65 | 174,627 | 99.9 | 0.1 | 4431 |
| S. Dublin | 55.9 | 4.9 | 37 | 52.1 | 98.65 | 406,024 | 99.7 | 0.3 | 4824 |
| S. Enteritidis | 64.9 | 4.6 | 32.5 | 52.1 | 98.65 | 440,347 | 100.0 | 0.2 | 4444 |
| S. Give | 78.7 | 4.5 | 29 | 52.2 | 98.65 | 454,909 | 100.0 | 0.0 | 4378 |
| S. Hartford | 36.8 | 4.8 | 60 | 52.1 | 98.65 | 236,095 | 99.4 | 0.1 | 4592 |
| S. Litchfield | 76.3 | 4.4 | 52 | 52.2 | 98.65 | 252,267 | 100.0 | 0.0 | 4391 |
| S. Montevideo | 71.3 | 4.6 | 31.8 | 52.3 | 98.65 | 433,986 | 100.0 | 0.2 | 4348 |
| S. Muenchen | 78.2 | 4.6 | 29 | 52.2 | 98.65 | 521,499 | 100.0 | 0.0 | 4345 |
| S. Newport | 62.7 | 4.8 | 36 | 52.1 | 98.65 | 563,829 | 100.0 | 0.1 | 4691 |
| S. Oranienburg | 53.2 | 4.6 | 33 | 52.1 | 98.65 | 542,191 | 100.0 | 0.0 | 4335 |
| S. Paratyphi | 82.4 | 4.6 | 29 | 52.3 | 98.65 | 369,141 | 100.0 | 0.5 | 4309 |
| Serotype | Number | Serogroup | MLST |
|---|---|---|---|
| S. Anatum | 2 | E1 | 64 |
| S. Agbeni | 1 | G | 2606 |
| S. Braenderup | 5 | C1 | 22 |
| S. Berta | 2 | D1 | 435 |
| S. Cerro | 23 | K | 367, 9243 |
| S. Dublin | 1 | D1 | 10 |
| S. Enteritidis | 2 | D1 | 11 |
| S. Give | 6 | E1 | 654 |
| S. Hartford | 7 | C1 | 405 |
| S. Litchfield | 1 | C2–C3 | 214 |
| S. Montevideo | 5 | C1 | 4, 138 |
| S. Muenchen | 3 | C2–C3 | 83 |
| S. Oranienburg | 10 | C1 | 23 |
| S. Newport | 9 | C2–C3 | 45, 350 |
| S. Paratyphi B var. L(+) tartrate+ | 1 | B | 3927 |
| Sample_Id | Serotypes | Gene | Antibiotic Class |
|---|---|---|---|
| AG21-0051 | Dublin | blaCMY-2, sul2, aph(3″)-Ib, aph(6)-Id, tet(A), floR | Cephalosporin, sulfonamide, streptomycin, streptomycin, tetracycline, chloramphenicol/florfenicol |
| AG21-0060 | Enteritidis | gyrA_D87Y | Quinolone |
| AG21-0066 | Enteritidis | gyrA_D87Y | Quinolone |
| AG21-0012 | Newport | floR, tet(A), aph(6)-Id, aph(3″)-Ib, sul2, blaCMY-2 | Chloramphenicol/florfenicol, tetracycline, streptomycin, streptomycin, sulfonamide, cephalosporin |
| AG21-0014 | Newport | floR, tet(A), aph(6)-Id, aph(3″)-Ib, sul2, blaCMY-2 | Chloramphenicol/florfenicol, tetracycline, streptomycin, streptomycin, sulfonamide, cephalosporin |
| AG21-0584 | Newport | floR, tet(A), aph(6)-Id, aph(3″)-Ib, sul2, blaCMY-2 | Chloramphenicol/florfenicol, tetracycline, streptomycin, streptomycin, sulfonamide, cephalosporin |
| AG21-0586 | Newport | floR, tet(A), aph(6)-Id, aph(3″)-Ib, sul2, blaCMY-2 | Chloramphenicol/florfenicol, tetracycline, streptomycin, streptomycin, sulfonamide, cephalosporin |
| AG21-0587 | Newport | floR, tet(A), aph(6)-Id, aph(3″)-Ib, sul2, blaCMY-2 | Chloramphenicol/florfenicol, tetracycline, streptomycin, streptomycin, sulfonamide, cephalosporin |
| AG21-0050 | Montevideo | fosA7 | Fosfomycin |
| AG21-0589 | Muenchen | aph(3″)-Ib, sul2, tet(A) | Streptomycin, sulfonamide, tetracycline |
| AG21-0590 | Muenchen | aph(3″)-Ib, sul2, tet(A) | Streptomycin, sulfonamide, tetracycline |
| AG21-0591 | Muenchen | aph(3″)-Ib, sul2, tet(A) | Streptomycin, sulfonamide, tetracycline |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhandari, M.; Poelstra, J.W.; Kauffman, M.; Varghese, B.; Helmy, Y.A.; Scaria, J.; Rajashekara, G. Genomic Diversity, Antimicrobial Resistance, Plasmidome, and Virulence Profiles of Salmonella Isolated from Small Specialty Crop Farms Revealed by Whole-Genome Sequencing. Antibiotics 2023, 12, 1637. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12111637
Bhandari M, Poelstra JW, Kauffman M, Varghese B, Helmy YA, Scaria J, Rajashekara G. Genomic Diversity, Antimicrobial Resistance, Plasmidome, and Virulence Profiles of Salmonella Isolated from Small Specialty Crop Farms Revealed by Whole-Genome Sequencing. Antibiotics. 2023; 12(11):1637. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12111637
Chicago/Turabian StyleBhandari, Menuka, Jelmer W. Poelstra, Michael Kauffman, Binta Varghese, Yosra A. Helmy, Joy Scaria, and Gireesh Rajashekara. 2023. "Genomic Diversity, Antimicrobial Resistance, Plasmidome, and Virulence Profiles of Salmonella Isolated from Small Specialty Crop Farms Revealed by Whole-Genome Sequencing" Antibiotics 12, no. 11: 1637. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12111637